行政院國家科學委員會專題研究計畫 成果報告
斑馬魚肌肉調控蛋白 myf-5 基因上游近端序列之結合轉錄
因子及其調控機制(2/2)
研究成果報告(完整版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2313-B-002-028- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學分子與細胞生物學研究所 計 畫 主 持 人 : 蔡懷楨 計畫參與人員: 博士班學生:李鴻杰、林正勇 碩士班學生:羅方懿 報 告 附 件 : 出席國際會議研究心得報告及發表論文 處 理 方 式 : 本計畫可公開查詢中 華 民 國 96 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
斑馬魚肌肉調控蛋白
myf-5 基因上游近端序列之結合
轉錄因子及其調控機制(2/2)
計畫類別:■ 個別型計畫 □ 整合型計畫
計畫編號:NSC 95-2313-B-002-028-
執行期間:
2006 年 8 月 1 日至 2007 年 7 月 31 日
計畫主持人:
蔡懷楨
共同主持人:
計畫參與人員:
成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告
本成果報告包括以下應繳交之附件:
■赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年■二年後可公開查詢
執行單位:
國立臺灣大學分子與細胞生物學研究所
中 華 民 國 96 年 10 月 30 日
中文摘要:
Myf5 是一種 basic helix-loop-helix 轉錄因子,負責控制胚胎發育時期肌肉組織的分化, myf-5 的表現具有組織專一性與時期專一性的特性。在哺乳類與魚類中對於 myf-5 基因的調
控機制至今尚不清楚。利用顯微注射方法研究 myf-5 promoter 的結果顯示,在 myf-5 上游
-82/-62 序列為一重要的 cis-acting element,對 myf-5 基因的開啟有很大的影響 (Chen et al., 2003),但是到底是何種轉錄因子結合在我們發現的調控區上,目前不得而知。本文利用 yeast one hybrid system 找出與斑馬魚 myf-5 基因上游調控序列-82/-62 casette 結合的轉錄因子- Forkhead box d3 (Foxd3)。in vitro transactivation assay 證明 Foxd3 能促進斑馬魚 myf5 啟動子 活化。在注射foxd3-morpholino (MO)的胚胎中,發現 myf5 只侷限在 presomitic mesoderm 表 現,在體節與其鄰近的adaxial cells 則無法偵測到。Foxd3 對 myf5 的調控是有時間性的: 在 受精後12-18 小時的過程中,Foxd3 負責維持 Myf5 在體節與其鄰近的 adaxial cells 的表現。 此外,myod 的表現則未受到影響。在注射 pax3-MO 的胚胎中,發現 foxd3 的表現幾乎完全 消失,而myf5 的表現情況則與注射 foxd3-morpholino (MO)的胚胎類似。在同時注射 pax3-MO 與foxd3 mRNA 的胚胎中,發現 myf5 在體節與其鄰近 adaxial cells 的表現能被拯救(rescue)。 我們推論在胚發育的過程中,Pax3 控制 Foxd3 表現,之後 Foxd3 再調控 Myf5 的表現,進
而促進肌肉的發育。由於Foxd3 之前主要被認為是調控神經脊細胞發育的轉錄因子,因此
Abstract:
Myf5, one of the basic helix-loop-helix transcription factors, controls muscle differentiation and is expressed in somites during early embryogenesis. However, the transcription factors bound to the cis-elements of myf5 are poorly understood. In this study, we used the yeast one-hybrid assay and found that Forkhead box d3 (Foxd3) interacted specifically with the -82/-62 cassette, a key element directing somite-specific expression of myf5. The dual-luciferase assay revealed that the expression of Foxd3 potently transactivated the myf5 promoter. Knocking down foxd3 with morpholino oligonucleotide (MO) resulted in a dramatic down-regulation of myf5 in somites and adaxial cells but not in the presomitic mesoderm. Foxd3 mediation of myf5 expression is stage-dependent, maintaining myf5 expression in the somites and adaxial cells during the 7- to 18-somite stage. On the other hand, myod expression remained unchanged in foxd3 morphants. Furthermore, in the pax3 morphant, the expression of foxd3 was down-regulated greatly and the expression of myf5 was similar to that of the foxd3 morphant.Co-injection of foxd3 mRNA and pax3-MOs greatly restored the expression of myf5 in the somites and adaxial cells, suggesting that pax3 induces foxd3 expression, which then induces the expression of myf5. This report is the first study to show that Foxd3, a well-known regulator in neural crest development, is also involved in myf5 regulation.
前言:
In vertebrates, the specification of muscle progenitor cells in the somites depends on
inductive signals emanating from adjacent tissues, such as the neural tube, the notochord, and the
dorsal and lateral ectoderm. In response to inducers, muscle precursor cells (myoblasts) start
expressing several transcriptional activators that control the expression of muscle structural genes.
A family of muscle regulatory factors (MRFs) with a basic DNA binding motif and a basic
helix-loop-helix dimerization domain has been identified in mammals, birds, frogs, fish, insects,
and nematodes (Michelson et al., 1990; Hopwood et al., 1991; Buonanno et al., 1992; Saitoh et
al., 1993; Krause et al., 1994; Chen et al., 2000, 2001). Proteins in this family include Myod
(Davis et al., 1987), myogenin (Braun et al., 1989a; Edmondson and Olson 1989; Wright et al.,
1989), Myf5 (Braun et al., 1989b), and MRF4/herculin/Myf6 (Rhodes and, Konieczny 1989;
Braun et al., 1990; Miner and Wold 1990).
In mice, Myf5 is activated at different anatomical sites in the embryo under the control
of distinct, cis-acting regulatory elements (Hadchouel et al., 2000; Summerbell et al., 2000;
Carvajal et al., 2001). An enhancer, 6.6 kb upstream, is required for myf5 expression in the
epaxial domain (Gustafsson et al., 2002). A 270-bp core enhancer, about 57 kb upstream, directs
myf5 expression in limbs and maintains myf5 expression in somites (Buchberger et al., 2003). Another enhancer directs myf5 expression in cervical somites and restricts myf5 transcription in
the myotome. In Xenopus, two negative regulatory elements have been identified in the Xmyf5
promoter, which controls Xmyf5 expression. An interferon regulatory factor-like DNA binding
element down-regulates Xmyf5 expression in differentiating myocytes (Mei et al., 2001), and a
Wnt/ß-catenin signals (Yang et al., 2002). A T-box binding site mediates dorsal activation of
Xmyf5 transcription and is involved in the regulation of muscle development (Lin et al., 2003). In zebrafish myf5, the upstream sequence -82 to -1 (-82/-1) directs reporter gene expression
specifically in the somites but the -62/-1 segment cannot (Chen et al., 2001). Recently, Chen et al.
(2003) demonstrated that the -82/-62 regulatory cassette in zebrafish myf5 is a cis-element that is
able to direct somite-specific expression and repress nonspecific expression during early
embryogenesis. Although promoter analysis and identification of cis-regulatory elements have
been carried out with mouse, Xenopus, and zebrafish myf5, the trans-acting factors that actually
bind to cis-acting elements in myf5 remain unknown.
Somite patterning is under the control of a variety of signals provided by the dorsal
neural tube, notochord, floor plate, surface ectoderm, and lateral plate mesoderm (Münsterberg et
al., 1995; Marcelle et al., 1995; Pourquié et al., 1996; Hirsingeret al., 1997; Yamaguchi 1997;
Currie and Ingham 1998; Tajbakhsh et al., 1998; Reshef et al., 1998). Several transcription factors
and signaling modulators, such as bone morphogenetic protein 4 (BMP4; Cossu et al., 1996),
Noggin (Hirsinger et al., 1997), Wnt (Ikeya and Takada, 1998; Tajbakhsh et al., 1998; Borycki et
al., 1999), Sonic Hedgehog (Shh; Coutelle et al., 2001), Gli (Gustafsson et al., 2002), and Pax3
(Marcelle et al., 1995; Maroto et al., 1997; Tajbakhsh et al., 1997), play important roles in
mediating the response of signals from surrounding tissues to induce expression of MRFs. The
regulatory network of mouse myf5 has been elucidated (Cossu and Borello, 1999; Buckingham,
2001; Roth et al., 2003; Tajbakhsh, 2003), and it is proposed that pax3 regulates mouse myf5
However, the detailed molecular interaction among factors in the regulatory network during the
time of commitment to modulate myf5 expression has yet to be revealed.
研究目的:
In this study, using a yeast one-hybrid assay, we determined that Foxd3 interacted
specifically with the -82/-62 regulatory element of zebrafish myf5. Foxd3 also plays an important
role in maintaining myf5 expression in the somites and adaxial cells. This Foxd3 mediation of
myf5 is stage-dependent. Furthermore, we demonstrate that the expression of foxd3 in the pax3
morphant was down-regulated greatly and that the expression of myf5 was similar to that of the
foxd3 morphant. Injection of foxd3 mRNA rescued the defects caused by pax3 morpholino oligonucleotide (MO), suggesting that pax3 induces foxd3 expression, which then induces the
expression of myf5. This article is the first study to demonstrate that Foxd3 mediates myf5
expression and is involved in myogenesis during zebrafish embryogenesis.
研究方法:
Yeast one-hybrid screening
Yeast one-hybrid screening was performed according to the protocol of the manufacturer
(Clontech). The bait plasmids pHISi-6×(-82/-62) and pLacZi-6×(-82/-62) were constructed using
synthetic DNA oligomers containing six repeats of the myf5 -82/-62 cassette. Four plasmids were
used in the selection procedure as negative controls. They included p53HIS, which contains the
pHISi-m4m5 and pLacZi-m4m5, which contain four repeats of the myf5 -82/-62 cassette in which
the -70/-62 sequence was mutated to GAAGTTAAC (m4m5; Chen et al., 2003). The cDNA was
inserted into a plasmid, pGADT7-Rec, by homologous recombination in yeast. Transformed
plasmids were recovered with a yeast plasmid isolation kit (Clontech). The plasmids isolated
from each clone were transformed into Escherichia coli DH5α cells for amplification.
Fish embryos
Zebrafish (AB strain) were maintained at 28.5°C under a 14-h light/10-h dark photoperiod. After
fertilization, eggs were collected and cultured in an aquarium. The number of embryonic
cleavages was counted, and somite formation was observed under a fluorescent stereomicroscope
MZ FLIII (Leica).
Knockdown microinjection of zebrafish embryos
MOs were obtained from Gene Tools. The sequences of MOs were designed as follows:
foxd3-MO1, TGCTGCTGGAGCAACCCAAGGTAAG, antisense nucleotides 160 to 184 of zebrafish foxd3 cDNA (GenBank accession no. AF052249); foxd3-MO2,
CACTGGTGCCTCCAGACAGGGTCAT, antisense nucleotides 197 to 221 of zebrafish foxd3
cDNA; foxd3-MO-sense, ATGACCCTGTCTGGAGGC- ACCAGTG, sense nucleotides 197 to
221 of zebrafish foxd3 cDNA; pax3-MO1, GCTAATGCGGTCATATCTCCTCTGC, antisense
nucleotides 266 to 290 of zebrafish pax3 cDNA (GenBank accession no. NM131277); pax3-MO2,
ACGAAAAAAGGATGCACGAAGCACT, antisense nucleotides 241 to 265 of zebrafish pax3
cDNA; myf5-MO, TACGTCCATGATTGGTTTGGTGTTG, antisense nucleotides 28 to 52 of
at a stock concentration of 1 mM and diluted to the desired concentration for microinjection into
each embryo.
Electrophoretic mobility shift assay
The -82/-62 cassette, the mutated sequence of -82/-62 (sequence at -70/-62 was mutated to
GAAGTTAAC; m4m5) and a nonspecific sequence (Non-30fr) were used as oligonucleotide
probes for the binding assay (Chen et al., 2003). All probes were labeled with γ-[32P]ATP (3,000
μCi/ml) using T4 polynucleotide kinase (NEB). Probes, recombinant Foxd3 protein (50 or 500
ng), and 1 μg of poly(dIdC) were added to the reaction buffer (10 mM Tris at pH 7.5, 50 mM
NaCl, 0.5 mM ethylenediaminetetraacetic acid pH 8.0, 0.5 mM dithiothreitol, 5% glycerol) and
incubated at 30°C for 30 min. Unlabeled -82/-62 cassette, Non-30fr, and m4m5 were used for
competitive inhibition. After reacting, all products were analyzed by 6% acrylamide gel
electrophoresis (79:1 acrylamide:bisacrylamide). After transferring the bands to a 3M filter and
drying the gel, X-ray film was exposed to the sample for 2 days.
Cloning zebrafish Foxd3 cDNA and plasmid constructions
Full-length cDNA coding for zebrafish foxd3 was obtained by polymerase chain reaction (PCR)
from a cDNA library of 14-18 hours postfertilization (hpf) zebrafish embryos using the 5′-primer
CTCGAGATGACCCTGTCTGGAGGCACC and the 3′-primer
CTCGAGTCATTGAGAAGGCCATT in which an XhoI site was included. The PCR products
were first ligated into pGEMT-easy vector (Promega), then digested by XhoI, and subcloned into
a pET-15b vector (Novagen). The coding region of zebrafish foxd3 was amplified by PCR using
and the 3′-primer TCATTGAGAAGGCCATTTCGATACCG. The PCR products were subcloned
into the pGEMT-easy vector (Promega), digested by KpnI and NotI, then cloned into the pCMVm
vector, which contains the CMV promoter and enhancer (Chen et al., 2003). Plasmids
pZMYP-2937E and pZMYP-6212E, to which green fluorescent protein was fused with the
upstream 3 and 6 kb of zebrafish myf5, respectively, were described previously (Chen et al.,
2001). Plasmid pRL-ZMYP3.0, containing an upstream 2.9 kb of zebrafish myf5 that was
recovered from an AgeI-PstI-cut pZMYP-2937E, was subcloned into the NheI-PstI-cut
phRL-Null vector (Promega) in which the NheI was blunted. Plasmid pRL-ZMYP6.0, containing
the upstream 6.2 kb of myf5 that was recovered from an AgeI-PstI-cut pZMYP-6212E, was
subcloned into the phRL-Null vector by using the same strategy.
Preparation of recombinant proteins in vitro
E. coli BL21 (DE3)/pLysS containing zebrafish foxd3 cDNA was cultured to produce recombinant Foxd3. Following induction, E. coli was treated with 0.5 mM
isopropyl-1-thio-β-D-galacto-pyranoside for 4 h at 37 °C. After the cells were lysed, recombinant
Foxd3 was purified with a Ni-NTA spin column (Qiagen)
Cell culture
Monkey kidney COS-1 cells were cultured in Dulbecco's modified Eagle’s medium (DMEM,
Biowest) containing 10% fetal bovine serum (Biowest), which was heat-inactivated by incubating
for 30 min at 56°C, supplemented with 1× penicillin/streptomycin/glutamine (Biowest), and then
incubated at 37°C in an atmosphere of 5% CO2 and 95% air. Fresh culture medium was provided
embryonal carcinoma cell line P19 was cultured in alpha minimum essential medium (Gibco),
7.5% bovine calf serum (Biowest), 2.5% fetal bovine serum, and 1×
penicillin/streptomycin/glutamine. Medium was renewed at least every 48 h and subcultured
every 2 or 3 days.
Dual-luciferase assay
About 1×105 cells were seeded onto each well of six-well plates 24 h prior to transfection. Cells
were transfected by the lipofectamine method (Invitrogen) according to the manufacturer’s
instruction. Transfection mixtures normally contained 6 µl of lipofectamine and 1 to 2 µg of
plasmid constructs of firefly luciferase and Renilla luciferase (RL). After a 48-h transfection,
cells were harvested for luciferase assay by using the Dual-Luciferase Reporter Assay System
(Promega). Luciferase activity was measured from three separated experiments in a Luminoskan
Ascent (Thermo Labsystems).
RNA in situ hybridization
Whole-mount in situ hybridization of whole embryos was performed by using digoxigenin (DIG)
-labeled riboprobes of myf5, foxd3, myod, myogenin, and α-actin. We followed the procedures as
described by Chen et al. (2001), except that phosphate-buffered saline with 0.1% Tween 20, 2
mg/ml bovine serum albumin, 5% sheep serum, and 1% dimethyl sulfoxide were used in the
blocking solution. To determine the co-localization of myf5 and foxd3 transcripts, double in situ
hybridizations were performed following the scheme of Jowett (2001), except that foxd3 was
labeled with fluorescein-UTP (Roche) and myf5 was labeled with DIG-UTP.
Capped mRNAs of foxd3, myf5, and red fluorescent protein (RFP) were synthesized according to
the protocol of the manufacturer (Epicentre). The resulting mRNA was diluted to11 or 22 ng/µl
for foxd3 mRNA, 130 or 260 ng/µl for myf5 mRNA, and 44 ng/µl for RFP mRNA with distilled
water, and 2.3 nl of each was injected into 1-cell stage embryos. In the rescue experiments,
embryos were observed at 13- to 16-somite stage in terms of morphological defects and the
expression patterns of target genes by using whole mount in situ hybridization.
結果:
Foxd3 is the cognate protein bound to the myf5 -82/-62 cassette
Yeast one-hybrid screening was used to identify the factor bound to the -82/-62 cassette of
zebrafish myf5. Six copies of the -82/-62 cassette were inserted in the region upstream of the
Saccharomyces cerevisiae selection marker YM4271. A cDNA library constructed from mRNA of 15-18 hpf embryos was screened, and 8.5 x 103 clones were grown on minimal medium. About
162 colonies that not only were able to grow on the selective medium but that also were
lacZ-positive were isolated. Then, their insert DNA fragments were cloned and sequenced. After these cDNA sequences were identified from the gene bank using the BLAST procedure, we
selected 83 putative colonies containing the full-length cDNA and back-transformed them into
yeasts. Finally, there were 17 colonies containing cDNA fragments that were able to interact
specifically with the -82/-62 bait. One of these colonies containing foxd3 cDNA was chosen for
further study because this cDNA insert was somite-positive after in situ hybridization. The
binding specificity of Foxd3 and -82/-62 cassette was evaluated with two assays. First,
medium only when they contained the wild-type myf5 -82/-62 cassette. Recombinant yeasts
harboring the mutated sequence within -82/-62 did not grow (Fig. 1A). Second, β-galactosidase
activity was detected in yeasts containing Foxd3 fused with the activation domain when the myf5
-82/-62 cassette was upstream of lacZ. However, β-galactosidase activity was not detectable in
yeasts containing mutated sequences of the myf5 -82/-62 cassette (Fig. 1B).
Electrophoretic mobility shift assay demonstrates that recombinant Foxd3 binds to the -82/-62 cassette
Electrophoretic mobility shift assay (EMSA) was used to determine whether the myf5
-82/-62 cassette is able to bind Foxd3 in vitro. Recombinant Foxd3 produced by E. coli interacted
specifically with the -82/-62 probe, producing the shifted band shown on the gel (Fig. 2). In
addition, the shifted band of DNA-protein complex was lost completely when excess amounts of
cold -82/-62 oligonucleotides were added. But, neither the nonspecific DNA competitor (Non30fr;
Fig. 2, lane 7) nor the mutated -82/-62 competitor (m4m5; Fig. 2, lane 4) interfered by forming a
specific complex between recombinant Foxd3 and the -82/-62 probe. Thus, the interaction
between Foxd3 and the myf5 -82/-62 cassette is specific.
Transactivation of the myf5 promoter by Foxd3
To test the functional consequences of Foxd3 interactions with the myf5 -82/-62 cassette, we
performed transient transfection assays with a luciferase reporter gene under the control of the
zebrafish myf5 promoter in the COS-1 and P19 cell lines. The upstream 3-kb or 6-kb region of
zebrafish myf5 was cloned to the phRL-Null vector. Results showed that the luciferase activity in
COS-1 cells in the presence of recombinant Foxd3 was 1.8-fold (pRL-ZMYP3.0) or 2.4-fold
Fig. 3). This case was also the finding when we used the P19 cells: the luciferase activity was
increased 3.65-fold (pRL-ZMYP3.0) or 4.16-fold (pRL-ZMYP6.0) by the presence of Foxd3 (n =
4, P < 0.05; Fig. 3). Thus, we conclude that the interaction between Foxd3 and the myf5 promoter
resulted in transactivation of gene expression.
Expression patterns of myf5 and foxd3 were colocalized
The pattern of foxd3 expression in zebrafish embryos varied with developmental stage.
During the 4- to 6-somite stage, foxd3 was transcribed in the floor plate, presumptive neural crest
cells, and tail bud (Fig. 4A). During the 7- to 9-somite stage, foxd3 expression was first detected
in the somites (Fig. 4B, arrowhead) and the signals also appeared in the posterior premigratory
crests and tail bud. During the 11- to 16-somite stage, foxd3 was expressed strongly in the
somites and the signals in the neural crest cells were down-regulated prior to neural crest
migration (Fig. 4C, D). However, faint signals in the lateral head and in the loose cluster of
foxd3-positive cells in the posterior head increased incrementally with the formation of each pair of somites (Fig. 4C, D, arrow). In embryos with 16-28 somites, foxd3 expression in the somites
was high but was down-regulated rapidly in the somites after the 28-somite stage (data not
shown). Use of double in situ hybridization to detect both the myf5 and the foxd3 transcripts at the
11- to 13-somite stage revealed that the expression domains of myf5 and foxd3 coincided greatly
in somites and adaxial cells (Figs. 4E and F), indicating that the expression patterns of foxd3 and
myf5 were co-localized.
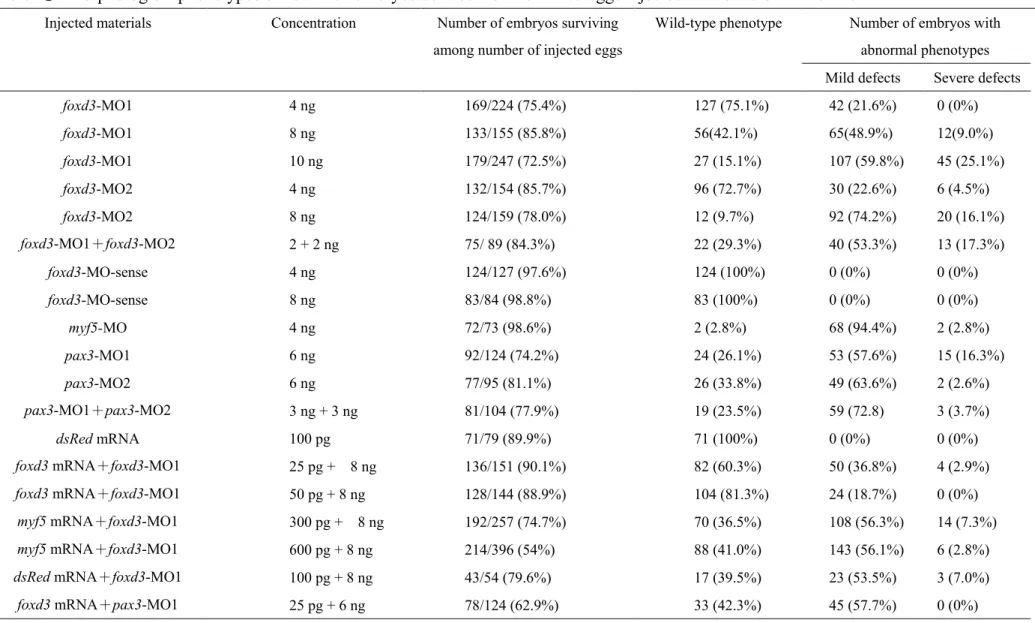
In the foxd3 morphant, morphological defects were dose-dependent
one complementary to 25 bp of the 5′-untranslated region of foxd3 mRNA (foxd3-MO1) and the
other complementary to 25 bp after AUG of foxd3 mRNA (foxd3-MO2). Embryos that received
two types of foxd3-MOs displayed similar defective phenotypes, whereas embryos that received
only the control MO (foxd3-MO-sense) developed normally, even when we injected it at a
concentration as high as 8 ng/embryo (Table 1). When 4-10 ng of foxd3-MOs were injected,
morphants with 12-14 somites displayed defects in the head and the tail bud (Figs. 5A, B).
Although the frequency of segmentation defects did not differ in wild-type and
foxd3-MO1-injected embryos, the somites of foxd3 morphants became broader than the somites of wild-type embryos (Figs. 5C, D). Most abnormalities were mild and included a raised tail, a
reduced head, and wider somites with an irregular boundary.Minor abnormalities led to serious
defects that retarded development. It is interesting to note that the defects caused by injecting two
foxd3-MOs were dose-dependent and synergistic (Table 1). To confirm whether the foxd3-MO-induced defects were specific, we co-injected synthetic foxd3 mRNA and foxd3-MO1. The foxd3-MO1 was used because it was complementary to a sequence of the 5′-untranslated
region, i.e., it blocked the endogenous foxd3 mRNA but not the microinjected foxd3 mRNA.
Synthetic foxd3 mRNA (25-50 pg) largely rescued the morphological defects induced by
foxd3-MO1 (Table 1), suggesting that the foxd3-MO-induced defects were specific.
Effects of foxd3 on myf5 expression are stage-dependent
To determine whether the states of differentiation of the somites were affected in foxd3
morphants, we assayed the expression of a number of genes that are normally expressed in
morphants and wild-type embryos were the same, but no signals were detected in the somites and
adaxial cells during the 8- to 10-somite stage (Figs. 6A, G).During the 11- to 13-somite stage,
myf5 gene expression in morphants was inactivated in the somites and adaxial cells (located on the lateral portions of somites 1-9) but signals in the PSM were not lost (Figs. 6B, H). Meanwhile,
myf5 expression decreased greatly in the somites and the adaxial cells during the 14- to 16-somite stage, but myf5 transcripts were expressed normally in the PSM (Figs. 6C, I). In foxd3 morphants,
myf5 was expressed normally in the PSM: these embryos did not lose myf5 expression patterns in somites 0 and -1 (Figs. 6G-J, arrowheads). However, the myf5 signals greatly decreased in the
newly formed somites and in the completely formed somites, indicating that foxd3 functions to
maintain myf5 gene expression in the somites but not in the PSM during somitogenesis.
After the 17- to 19-somite stage, endogenous myf5 expression was down-regulated
and differences in myf5 expression between wild-type embryos and foxd3 morphants became
negligible (Figs. 6D, J). This down-regulated expression pattern persisted for wild-type embryos
and foxd3 morphants after the 20- to 22-somite stage (Figs. 6E, 6F vs 6K, 6L). However, at these
stages, the expression level of foxd3 in the somites was still strong. This evidence clearly
demonstrated that the regulation of myf5 through foxd3 was stage-dependent. Moreover, we also
found an unexpected ectopic expression of myf5 in the tail bud in foxd3 morphants with raised
tails (Fig. 6H, arrow), suggesting that Foxd3 may play other roles in the tail bud.
Foxd3 modulates expression of myf5 but not myod
We compared the expression of two genes involved in somitogenesis in foxd3
myf5) and α-actin (the structural protein in somites) clearly were down-regulated in the somites but were expressed normally in adaxial cells (Figs. 7C, D, G, H). However, myod expression in
the somites and adaxial cells remained unchanged (Fig. 7B, F). Furthermore, we noticed that 10
somites were positive for myod staining in wild-type embryos, whereas 6 were positive in the
foxd3-MO-injected embryos. This difference may be due to development delay in MO-treated embryos. Thus, we suggest that Foxd3 specifically regulated the expression of myf5 but not
myod.
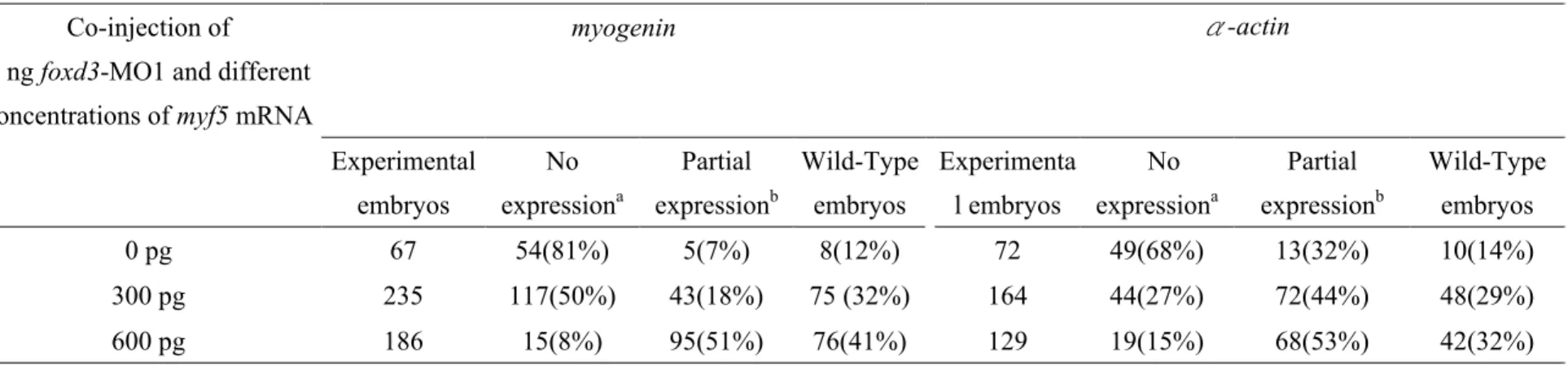
foxd3 knockdown morphant can be rescued by injecting myf5 mRNA
To determine whether the foxd3 morphant phenotype could be rescued by myf5, we co-injected
foxd3-MO1 and synthetic myf5 mRNA. A series of different concentrations of myf5 mRNA was injected together with 8 ng of foxd3-MO1 into eggs. The expression of myogenin and α-actin was
rescued partially by injecting myf5 mRNA compared to the expression in embryos that were
injected with foxd3-MO1 alone (Table 2; Figs. 7G vs I; 7H vs J). Co-injection of myf5 mRNA
effectively rescued the expression of myogenin (from 7% to 51% of defects) and α-actin (from
32% to 60% of defects) at the concentration range of 300-600 pg (Table 2). Moreover, embryos
that were injected with myf5-MO showed reduced expression of myogenin but not of myod (Fig.
7I). The expression pattern of myogenin in foxd3 morphants and in myf5 morphants was similar.
Taken together, our data suggest that myf5 mRNA effectively rescues the foxd3 morphant
phenotype and that the foxd3-MO1 used in this study specifically inhibits myf5 expression.
Molecular control of pax3, foxd3, and myf5
myogenesis, we injected pax3-MOs into embryos to repress pax3 expression. Like the strategy of
using foxd3-MOs, two pax3-MOs were designed: pax3-MO1 and pax3-MO2. Embryos that
received pax3-MO1 displayed severe convergence/extension phenotypes. The defects caused by
these two MOs were similar, dose-dependent, and synergistic (Table 1). We found that foxd3
expression was down-regulated significantly (Figs. 8D vs E). Meanwhile, myf5 expression was
restricted in the PSM and weak in somites 0 and -1 and in the adaxial cells on the sides of somites
0 and -1 (Figs. 8A, B). The patterns of myf5 expression were similar in morphants derived from
embryos treated with foxd3 mRNA and pax3-MO (Figs. 6H, 7E, 8B). However, in pax3
morphants, myod clearly was down-regulated in the somites but was expressed normally in
adaxial cells (Figs. 8F, G). Furthermore, we found that, when 23 pg of foxd3 mRNA and 6 ng of
pax3-MO1 were co-injected, the expression of myf5 was greatly restored in the somites (Fig. 8C), but it did not rescue myod expression in the somites (Fig. 8H). Based on these findings, we
conclude that (1) foxd3 specifically regulates expression of myf5 but not myod, (2) pax3 acts as an
upstream regulator of foxd3, and (3) down-regulation of myf5 in pax3 morphants occurs because
foxd3 is not expressed.
討論:
The winged helix transcription factor forkhead gene was first described in Drosophila
(Weigel et al., 1989). Rodent HNF3 transcription factor is very similar (Weigel and Jäckle 1990;
Lai et al., 1990, 1991). Forkhead domains have been reported in organisms ranging from yeasts
binding domain, more than 10 different classes of forkhead genes have been described. Some
classes have been divided into subclasses a-d (Kaufmann and Knöchel 1996). Foxd3 (CWH3,
Hfh2, fkd6) is in class V.
foxd3 is expressed in the presumptive neural crest region in both chick and mouse embryos and plays a role in neural crest differentiation in multiple systems (Freyaldenhoven et al.,
1997; Hromas et al., 1999; Kos et al., 2001). Overexpression of foxd3 in a line of myeloid cells
prevents them from maturing into granulocytes (Xu et al., 1998). Ectopic expression of foxd3 in
the neural tube of chicks changes the fate of cells into neural crest-like cells and can interfere
with subsequent differentiation (Dottori et al., 2001). In addition to the somatic mesoderm,
zebrafish foxd3 is transcribed in the somitic mesoderm, paraxial mesoderm, and tail bud
(Odenthal and Nüsslein-Volhard, 1998; Fig. 4). Whether foxd3 is involved in myogenesis is
unknown. Previously, we demonstrated that a cis-element (-82/-62) of zebrafish myf5 drives
somite-specific expression and represses nonspecific expression during the early development of
zebrafish embryos (Chen et al., 2003). In this study, we discovered that the winged helix
transcription factor foxd3 interacts specifically with the myf5 -82/-62 cassette (Fig. 2). The
dual-luciferase assay revealed that the expression of Foxd3 potently transactivated the myf5
promoter (Fig. 3) and that foxd3 plays an important role in mediating myf5 expression during
somitogenesis.
Foxd3 is necessary for maintaining myf5 expression in somites
Many regulatory modules are thought to be responsible for directing the
somites are induced by factors secreted from a variety of adjacent tissues, such as Shh (Fan and
Tessier-Lavigne 1994), Wnts (Münsterberg et al., 1995; Stern et al., 1995), Pax3 (Maroto et al.,
1997; Tajbakhsh et al., 1997), and BMP (Pourquié et al., 1996; Dietrich et al., 1998). These
environmental signals affect the initiation and continued expression of myf5 in the somites. Here,
we find that Foxd3, another regulatory module, has a novel function in myf5 expression due to the
finding that down-regulation of foxd3 suppresses myf5 expression in the somites.myf5 expression
decreases dramatically in the somites and adaxial cells in the Foxd3 knockdown embryos (Figs.
4C, I). Addition of myf5 mRNA effectively rescues the expression of both myogenin and α-actin
(Fig. 7) in the somites of foxd3 morphants, indicating that foxd3 modulates myf5 expression
specifically. However, myf5 expression remains unchanged in the PSM of Foxd3 knockdown
embryos. The endogenous foxd3 is not expressed in the PSM (Odenthal and Nüsslein-Volhard
1998; Fig. 4). Therefore, we propose that foxd3 functions in newly formed somites but not in the
PSM.
The expression of zebrafish myf5 is stage-specific and is restricted, particularly in the
posterior part of each somite (Chen et al., 2001; Coutelle et al., 2001). Very little is known about
the regulators that are involved in this delicate expression of myf5. In this report, we demonstrate
clearly that Foxd3 is a trans-acting factor that binds directly at the upstream cis-elements of
zebrafish myf5 gene. Knockdown of Foxd3 level leads to a reduction of myf5 transcripts in the
newly formed somites but not in the PSM (Fig. 6). Thus, we propose that Foxd3 functions to
maintain the continued expression of myf5 in the somites but does not function to initiate myf5
posterior part of newly formed somites. Misexpression of foxd3 leads to the ectopic expression of
myf5 but not myod (data not show). These results indicate that foxd3 may function to restrict myf5 expression in the posterior part of somites. In addition, myod was expressed normally in the
posterior part of the somites in the foxd3 morphants, suggesting that the mechanism of restricting
expression of myf5 is independent of myod.
Compared with the expression pattern of myf5, foxd3 transcripts reached a relatively
high level at the 9- to 18-somite stage, then foxd3 was down-regulated rapidly after 24 hpf
(Odenthal and Nüsslein-Volhard 1998; data not show). Similarly, myf5 transcripts increased
substantially until the 16- to 18-somite stage and then declined gradually to an undetectable level
by 26 hpf (Chen et al., 2001). In this report, we find that myf5 expression patterns in the wild-type
embryos and in the foxd3 morphants are similar after the 20- to 22-somite stage, although the
expression level of foxd3 in the somites was still strong at these stages. However, zebrafish myf5
transcripts in the somites became weaker and weaker after the 14- to 16-somite stage, and the
transcripts were present only in the PSM close to the tail bud by 24 hpf (Chen et al., 2001). Taken
together, we propose that the foxd3 modulation on myf5 expression is stage-dependent. Foxd3 is
required for myf5 activation in the somites between the 7- to 18-somite stage, suggesting different
factors and mechanisms are involved in myf5 down-regulation or perturbed Foxd3 functions in
myf5 activation after the 17-somite stage.
We also found that foxd3 morphants express myf5 ectopically in the tail bud (Fig. 4H).
However, cells located in the tail bud do not express myf5 or myod. Endogenous foxd3 also was
mediates myf5 expression in the somites differently than in the tail bud. The function of foxd3
may be similar to the Wnt signal, which has different effects on the cranial paraxial mesoderm
and on the trunk (Tzahor et al., 2003).Second, the effect may be due to a convergence-extension
defect during gastrulation. Zebrafish mutants, such as spadetail, have a severe defect in
convergence-extension of the trunk paraxial mesoderm. They lose trunk somites, resulting in
paraxial mesoderm cells accumulating in the tail (Kimmel et al. 1989). These hypotheses merit
further investigation.
In foxd3 morphants, the morphological changes we find by 24 hpf are that somites
become wider, with an irregular somite boundary (Figs. 5C, D). But these embryos are able to
twitch as normal as wild-type embryos. We also observe that the morphology of foxd3 morphants
remains unchanged during 3 to 5 days post-fertilization. Moreover, only embryos that received
both myf5- and myod-MO lost somites (data not show). Nevertheless, embryos that received
foxd3-, myf5-, or myod-MO alone still were able to develop somites normally. Taken together, these findings may be because myf5 and myod have complementary functions during
somitogenesis. The function of zebrafish Myf5 is redundant during somitogenesis. However,
whether Myf5 and Myod still play complementary roles in the muscle system other than somite,
such as craniofacial muscle development, is worthwhile to study.
In the caudal region, paraxial mesoderm is produced by gastrulation in the primitive
streak or tail bud. The foxd3-MO-injected embryos displayed abnormal heads, defective tails, and
wider somites (Figs. 5). Moreover, myf5 mRNA did not rescue the morphological defects induced
convergence-extension defect during gastrulation, but may be not related during somitogenesis.
In zebrafish, Odenthal and Nüsslein-Volhard (1998) demonstrated that there is a strong
expression of foxd3 in the involuting dorsal mesoderm. Mutation in zebrafish trilobite (tri) or
knypek (kny) gene affects the convergence-extension movements. Somites in the tri, kny, or kny;tri mutant appear substantially wider in their mediolateral dimension (Solnica-Krezel et al., 1996; Marlow et al., 1998; Sepich et al., 2000; Henry et al., 2000). In mouse, Foxd3-/- embryos
are dead after implantation at approximately 6.5 days post coitum, with a loss of epiblast cells,
expansion of proximal extraembryonic tissues, and a distal, mislocalized anterior organizing
center, suggesting that Foxd3 is required for maintaining embryonic cells of the early mouse
embryo (Hanna et al, 2002). Taken together, it is highly possible that zebrafish Foxd3 may
function during gastrulation. Loss of Foxd3 function causes abnormal heads, abnormal tails, and
wider somites, evident as the defective phenotype displayed by foxd3 morphants.
Regulation of myf5 by pax3 is mediated by foxd3
Pax3 belongs to the family of paired-box-containing transcription factors. It is
expressed in developing somites, the dorsal spinal cord, mesencephalon, and neural crest
derivatives. Heterozygous Splotch mice are characterized by pigmentation defects due to a
disorder in neural crest formation. Furthermore, homozygous embryos exhibit neural defects,
including spina bifida and exencephaly (Tremblay and Gruss 1994). All neural crest derivatives
caudal to the boundary of the hindbrain and spinal cord are lost. In addition, foxd3 is not
expressed in the caudal region of Splotch mutants, where dorsal root ganglia and sympathetic
pax3 (Dottori et al., 2001). Ectopic expression of pax3 throughout the neural tube alters the dorsal–ventral characteristics of the neural tube, represses floor plate formation, and decreases
motor neuron differentiation in transgenic mice embryos (Tremblay et al., 1996). Forced
expression of pax3 can activate expression of both myod and myf5 in paraxial mesoderm cultures
and in neural tube explants. In addition, genetic and in vitro analysis determined that pax3
regulation of mouse myf5 is indirect (Maroto et al., 1997; Tajbakhsh et al., 1997; Roth et al.,
2003).
pax3 plays a distinct role in the development of myogenic precursors and is thought to function upstream of myod (Marcelle et al., 1995; Maroto et al., 1997; Tajbakhsh et al., 1997).
The Splotch mutant lacks limb musculature (Franz et al., 1993; Bober et al., 1994), demonstrating
that pax3 is necessary for the migration of muscle precursor cells (Bober et al., 1994; Goulding et
al., 1994; Marcelle et al., 1995). myod is not activated in the Splotch/myf5 double null mutant,
suggesting that myod acts genetically downstream of pax3 and myf5 in the establishment of
skeletal muscle in the body (Tajbakhsh et al., 1997). Relaix et al. (2003) showed that pax3 cannot
directly activate the distal myod enhancer, indicating that pax3 activates myod indirectly. Thus,
whether Pax3 acts directly on myod in mammals remains unclear. In zebrafish, the expression of
pax3 is observed first in somites of embryos at the 6- to 8-somite stage. Transcripts are detected in most of the somites until the 14- to 16-somite stage; subsequently, expression is reduced (Seo
et al. 1998). In this report, embryos that received pax3-MOs display severe
convergence/extension phenotypes. This consequence may be because pax3 is detected initially at
morphants display defective convergence/extension phenotypes, and foxd3 expression is
down-regulated greatly in pax3-MO1-injected zebrafish embryos, suggesting that pax3 functions
upstream of foxd3. However, the stripe pattern of myf5 can be rescued by foxd3 mRNA in pax3
morphants (Fig. 8), but the convergence/extension phenotype can not (Table 1). Actually, the
molecular mechanism about pax3 and foxd3 involved in gastrulation are still unclear at the
present study. Taken together, we speculate that foxd3 may be one of the downstream target
genes of pax3 during gastrulation. Over-expression of foxd3 in pax3 morphants could not rescue
all the functions that pax3 plays. Besides, it is also reasonable to speculate that Pax3 and Foxd3
seems to be acting in independent ways during gastrulation.
In pax3 morphants, myf5 expression is restricted to the PSM, as is the case in foxd3
morphants. In addition, the pax3 (Seo et al. 1998) and foxd3 (Fig. 4) transcripts are not detectable
in the PSM. Over-expression of foxd3 mRNA in pax3 morphants rescues the stripe patterns of
myf5 in the somites, indicating that these factors are permissive for the expression of the target genes, but not instructive. Taken together, these findings suggest that myf5 down-regulation
occurs in pax3 morphants, because foxd3 is not expressed. Pax3 and Foxd3 function to maintain
myf5 expression in the newly formed somites. Moreover, our analysis of zebrafish foxd3 and pax3 morphants demonstrated that foxd3 specifically regulates expression of myf5 but not myod (Fig. 7,
8). These data suggest that Pax3 may activate the expression of myf5 and myod through a
different regulatory mechanism. Therefore, we propose a model in which the signaling cascade
involved in muscle development begins with pax3 inducing foxd3 expression, which then induces
Delicate network of myf5 regulation during embryogenesis
The function of cassette -82/-62 is to recruit a trans-factor to drive somite-specific
expression. Several studies have found that a transcription factor interacts with a ubiquitous
factor to drive tissue-specific expression. In neuron- specific expression, a neuron-restricted
transcription factor, MASH1, interacts with CBF to drive tissue-specific expression (Mandolesi et
al., 2002). Cardiac-specific expression is directed by the interaction of a heart-specific factor,
myocardin, with a ubiquitous serum response factor bound to a CArG box (Wang et al., 2001).
We found that the trans-factor Foxd3 binds to the -82/-62 cis-element and has a novel function
during myogenesis. This function is totally distinct from previously known functions of Foxd3 in
neural crest development. We hypothesize that Foxd3 interacts with different cofactors in the
somites and neural crest cells. In addition, we think that the effect of Foxd3 in the tail bud is
different from its effect in the somites. This difference occurs because different cofactors bind to
cassette -82/-62. As a result, Foxd3 plays a unique role in regulating myf5 expression. These
hypotheses merit further investigation.
A trans-acting factor can function as both an activator and a repressor, depending on
its binding sequence and/or its interaction with a specific cofactor. Transcription factors YY1
(Shrivastava and Calame 1994; Shi et al., 1997) and NF-Y (Peng and Jahroudi 2002) function in
this way. Moreover, the ectopic expression of Foxd3 in the chick neural tube induces expression
of migratory neural crest markers but suppresses interneuron differentiation (Dottori et al., 2001).
Over-expression of Foxd3 prevents the migration of neural crest cells along the dorsolateral
found that Foxd3 has opposite effects in the somites and tail bud and can function as both an
activator and a repressor. Recently, we reported that a cis-acting element in the downstream
region (+502/+835) of zebrafish myf5 intron 1 represses myf5 expression (Lin et al., 2004). On
the basis of these results, we hypothesize that the zebrafish myf5 -82/-62 cassette, the intron 1
+502/+835 element, Foxd3 and other unknown factors, and even a cis-element located in the
distal upstream region form a huge complex that orchestrates the spatiotemporal expression of
myf5 during somitogenesis.
In summary, we characterized signals that modulate somitogenesis. Our results reveal
that the winged helix transcription factor Foxd3 plays an important role in maintaining myf5
expression in the newly formed somites and the adaxial cells. We propose a model of gene
regulation in which pax3 induces foxd3 expression, which then induces the expression of myf5.
However, because inhibition of foxd3 translation does not completely abolish myf5 expression in
the PSM, we speculate that factors other than Foxd3 are involved in regulating myf5 expression in
the PSM.
參考文獻:
本計畫之相關研究內容已於2006 年發表於國際期刊Developmental Biology 290, 359-372.
Blagden, C.S., Currie, P.D., Ingham, P.W., Hughes, S.M., 1997. Notochord induction of zebrafish slow muscle mediated by Sonic hedgehog. Genes Dev. 11, 2163-2175.
Bober, E., Franz, T., Arnold, H.H., Gruss, P., Tremblay, P., 1994. Pax-3 is required for the development of limb muscles: a possible role for the migration of dermomyotomal muscle progenitor cells. Development 120, 603-612.
Borycki, A.G., Brown, A.M., Emerson, C.P.Jr., 2000. Study of skeletal myogenesis in cultures of unsegmented paraxial mesoderm. Development 127, 2075-2087.
Borycki, A.G., Brunk, B., Tajbakhsh, S., Buckingham, M., Chiang, C., Emerson, C.P.Jr., 1999. Sonic hedgehog controls epaxial muscle determination through Myf5 activation. Development
126, 4053-4063.
Braun, T., Bober, E., Buschhausen-Denker, G., Kohtz, S., Grzeschik, K.H., Arnold, H.H., 1989a. Differential expression of myogenic determination genes in muscle cells: possible autoactivation by the Myf gene products. EMBO J. 8, 3617-3625.
Braun, T., Bober, E., Winter, B., Rosenthal, N., Arnold, H.H., 1990. Myf-6, a new member of the human gene family of myogenic determination factors: evidence for a gene cluster on chromosome 12. EMBO J. 9, 821-831.
Braun, T., Buschhausen-Denker, G., Bober, E., Tannich, E., Arnold, H.H. 1989b. A novel human muscle factor related to but distinct from MyoD1 induces myogenic conversion in 10T1/2 fibroblasts. EMBO J. 8, 701-709.
Buchberger, A., Nomokonova, N., Arnold, H.H., 2003. Myf5 expression in somites and limb buds of mouse embryos is controlled by two distinct distal enhancer activities. Development 130, 3297-3307.
Buckingham, M., 2001. Skeletal muscle formation in vertebrates. Curr Opin Genet Dev. 11, 440-448.
Buonanno, A., Apone, L., Morasso, M.I., Beers, R., Brenner, H.R., Eftimie, R., 1992. The MyoD family of myogenic factors is regulated by electrical activity: isolation and characterization of a mouse Myf5 cDNA. Nucleic Acids Res. 20, 539-544.
Carvajal, J.J., Cox, D., Summerbell, D., Rigby, P.W., 2001. A BAC transgenic analysis of the Mrf4/Myf5 locus reveals interdigitated elements that control activation and maintenance of gene expression during muscle development. Development 128, 1857-1868.
Chen, Y.H., Lee, H.C., Liu, C.F., Lin, C.Y., Tsai, H.J., 2003. Novel regulatory sequence -82/-62 functions as a key element to drive the somite-specificity of zebrafish myf5. Dev. Dyn. 228, 41-50.
Chen YH, Lee WC, Cheng CH, Tsai HJ., 2000. Muscle regulatory factor gene: zebrafish (Danio rerio) myogenin cDNA. Comp. Biochem. Physiol. B 127, 97-103.
Chen, Y.H., Lee, W.C., Liu, C.F., and Tsai, H.J., 2001. Molecular Structure, Dynamic Expression, and Promoter Analysis of Zebrafish (Danio rerio) myf-5 Gene. Genesis 29, 22-35.
Cossu, G., Borello, U., 1999. Wnt signaling and the activation of myogenesis in mammals. EMBO J. 18, 6867-6872.
Cossu, G., Tajbakhsh, S., Buckingham, M., 1996. How is myogenesis initiated in the embryo? Trends Genet. 12, 218-223.
Coutelle, O., Blagden, C.S., Hampson, R., Halai, C., Rigby, P.W., Hughes, S.M., 2001. Hedgehog signalling is required for maintenance of myf5 and myoD expression and timely terminal differentiation in zebrafish adaxial myogenesis. Dev. Biol. 236, 136-150.
Currie, P.D., Ingham, P.W., 1998. The generation and interpretation of positional information within the vertebrate myotome. Mech. Dev. 73, 3-21.
Davis, R.L., Weintraub, H., Lassar, A.B., 1987. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell 51, 987-1000.
Dietrich, S., Schubert, F.R., Healy, C., Sharpe, P.T., Lumsden, A., 1998. Specification of the hypaxial musculature. Development 125, 2235-2249.
Dottori, M., Gross, M.K., Labosky, P., Goulding, M., 2001. The winged-helix transcription factor Foxd3 suppresses interneuron differentiation and promotes neural crest cell fate. Development
128, 4127-4138.
Edmondson, D.G., Olson, E.N., 1989. A gene with homology to the myc similarity region of MyoD1 is expressed during myogenesis and is sufficient to activate the muscle differentiation program. Genes Dev. 3, 628-640.
Fan, C.M., Tessier-Lavigne, M., 1994. Patterning of mammalian somites by surface ectoderm and notochord: evidence for sclerotome induction by a hedgehog homolog. Cell 79, 1175-1186. Franz, T., Kothary, R., Surani, M.A., Halata, Z., Grim, M., 1993. The Splotch mutation interferes
with muscle development in the limbs. Anat. Embryol. (Berl.) 187, 153-160.
Freyaldenhoven, B.S., Freyaldenhoven, M.P., Iacovoni, J.S., Vogt, P.K., 1997. Avian winged helix proteins CWH-1, CWH-2 and CWH-3 repress transcription from Qin binding sites. Cancer Res. 57, 123-129.
Goulding, M., Lumsden, A., and Paquette, A.J., 1994. Regulation of Pax-3 expression in the dermomyotome and its role in muscle development. Development 120, 957-971.
Gustafsson, M.K., Pan, H., Pinney, D.F., Liu, Y., Lewandowski, A., Epstein, D.J., Emerson, C.P. Jr., 2002. Myf5 is a direct target of long-range Shh signaling and Gli regulation for muscle specification. Genes Dev. 16, 114-126.
Hadchouel, J., Tajbakhsh, S., Primig, M., Chang, T.H., Daubas, P., Rocancourt, D., Buckingham, M., 2000. Modular long-range regulation of Myf5 reveals unexpected heterogeneity between skeletal muscles in the mouse embryo. Development 127, 4455-4467.
Hanna, L.A., Foreman, R.K., Tarasenko, I.A., Kessler, D.S., Labosky, P.A., 2002. Requirement for Foxd3 in maintaining pluripotent cells of the early mouse embryo. Genes Dev. 16, 2650-2661.
Henry, C.A., Hall, L.A., Burr, Hille, M., Solnica-Krezel, L., Cooper, M.S., 2000. Somites in zebrafish doubly mutant for knypek and trilobite form without internal mesenchymal cells or compaction. Curr. Biol. 7, 1063-1066.
Hirsinger, E., Duprez, D., Jouve, C., Malapert, P., Cooke, J., Pourquié, O., 1997. Noggin acts downstream of Wnt and Sonic Hedgehog to antagonize BMP4 in avian somite patterning. Development 124, 4605-4614.
Hopwood, N.D., Pluck, A., Gurdon, J.B., 1991. Xenopus Myf5 marks early muscle cells and can activate muscle genes ectopically in early embryos. Development 111, 551-560.
Hromas, R., Ye, H., Spinella, M., Dmitrovsky, E., Xu, D., Costa, R.H., 1999. Genesis, a Winged Helix transcriptional repressor, has embryonic expression limited to the neural crest, and stimulates proliferation in vitro in a neural development model. Cell Tissue Res. 297, 371-382.
Ikeya, M., Takada, S., 1998. Wnt signaling from the dorsal neural tube is required for the formation of the medial dermomyotome. Development 125, 4969-4976.
Kaufmann, E., Knöchel, W., 1996. Five years on the wings of fork head. Mech. Dev. 57, 3-20. Jowett, T., 2001. Double in Situ hybridization techniques in zebrafish. Methods 23, 345-358. Kimmel, C.B., Kane, D.A., Walker, C., Warga, R.M., Rothman, M.B., 1989. Amutation that
changes cell movement and cell fate in the zebrafish embryo. Nature 337, 358-362.
Kos, R., Reedy, M. V., Johnson, R. L., Erickson, C.A., 2001. The winged-helix transcription factor FoxD3 is important for establishing the neural crest lineage and repressing melanogenesis in avian embryos. Development 128, 1467-1479.
Krause, M., Harrison, S.W., Xu, S.Q., Chen, L., Fire, A., 1994. Elements regulating cell- and stage-specific expression of the C. elegans MyoD family homolog hlh-1. Dev. Biol. 166, 133-148.
Lai, E., Clark, K.L., Burley, S.K., Darnell, J.E.Jr., 1993. Hepatocyte nuclear factor 3/fork head or "winged helix" proteins: a family of transcription factors of diverse biologic function. Proc. Natl. Acad. Sci. U. S. A. 90, 10421-10423.
Lai, E., Prezioso, V.R., Smith, E., Litvin, O., Costa, R.H., Damell, J.E.Jr., 1990. HNF-3A, a hepatocyte-enriched transcription factor of novel structure is regulated transcriptionally. Genes Dev. 4, 1427-1436.
Lai, E., Prezioso, V.R., Tao, W.F., Chen, W.S., Damell, J.E.Jr., 1991. Hepatocyte nuclear factor 3 alpha belongs to a gene family in mammals that is homologous to the Drosophila homeotic gene fork head. Genes Dev. 5, 416-427.
Lin, C.Y., Chen, Y.H., Lee, H.C., Tsai, H.J., 2004. Novel cis-element in intron 1 represses somite expression of zebrafish myf5. Gene 334, 63-72
Lin, G.F., Geng, X., Chen, Y., Qu, B., Wang, F., Hu, R., Ding, X., 2003. T-box binding site mediates the dorsal activation of myf-5 in Xenopus gastrula embryos. Dev. Dyn. 226, 51-58. Mandolesi, G., Gargano, S., Pennuto, M., Illi, B., Molfetta, R., Soucek, L., Mosca, L., Levi, A.,
Jucker, R., Nasi, S., 2002. NGF-dependent and tissue-specific transcription of vgf is regulated by a CREB-p300 and bHLH factor interaction. FEBS Lett. 510, 50-56.
Marcelle, C., Stark, M.R., Bronner-Fraser, M., 1997. Coordinate actions of BMPs, Wnts, Shh and noggin mediate patterning of the dorsal somite. Development 124, 3955-3963.
Marcelle, C., Wolf, J., Bronner-Fraser, M., 1995. The in vivo expression of the FGF receptor FREK mRNA in avian myoblasts suggests a role in muscle growth and differentiation. Dev. Biol. 172, 100-114.
Marlow, F., Zwartkruis, F., Malicki, J., Neuhauss, S.C., Abbas, L., Weaver, M., Driever, W., Solnica-Krezel, L., 1998. Functional interactions of genes mediating convergent extension, knypek and trilobite, during the partitioning of the eye primordium in zebrafish. Dev. Biol. 203, 382-399.
Maroto, M., Reshef, R., Munsterberg, A.E., Koester, S., Goulding, M., Lassar, A.B., 1997. Ectopic Pax-3 activates MyoD and Myf5 expression in embryonic mesoderm and neural tissue. Cell 89, 139-148.
Mei, W., Yang, J., Tao, Q., Geng, X., Rupp, R.A., Ding, X., 2001. An interferon regulatory factor-like binding element restricts Xmyf-5 expression in the posterior somites during Xenopus myogenesis. FEBS Lett. 505, 47-52.
Michelson, A.M., Abmayr, S.M., Bate, M., Arias, A.M., Maniatis, T., 1990. Expression of a MyoD family member prefigures muscle pattern in Drosophila embryos. Genes Dev. 4, 2086-2097.
Miner, J.H., Wold, B., 1990. Herculin, a fourth member of the MyoD family of myogenic regulatory genes. Proc. Natl. Acad. Sci. U. S. A. 87, 1089-1093.
Münsterberg, A.E., Kitajewski, J., Bumcrot, D.A., McMahon, A.P., Lassar, A.B., 1995. Combinatorial signaling by Sonic hedgehog and Wnt family members induces myogenic bHLH gene expression in the somite. Genes Dev. 9, 2911-2922.
208, 245-258.
Peng, Y., Jahroudi, N., 2002. The NFY transcription factor inhibits von Willebrand factor promoter activation in non-endothelial cells through recruitment of histone deacetylases. Blood 99, 2408-2417.
Petropoulos, H., Gianakopoulos, P.J., Ridgeway, A.G., Skerjanc, I.S., 2004. Disruption of Meox or Gli activity ablates skeletal myogenesis in P19 cells. J. Biol. Chem. 279, 23874-23881. Pourquié, O., Fan, C.M., Coltey, M., Hirsinger, E., Watanabe, Y., Breant, C., Francis-West, P.,
Brickell, P., Tessier-Lavigne, M., Le Douarin, N.M., 1996. Lateral and axial signals involved in avian somite patterning: a role for BMP4. Cell 84, 461-471.
Relaix, F., Polimeni, M., Rocancourt, D., Ponzetto, C., Schafer, B.W., Buckingham, M., 2003. The transcriptional activator PAX3-FKHR rescues the defects of Pax3 mutant mice but induces a myogenic gain-of-function phenotype with ligand-independent activation of Met signaling in vivo. Genes Dev. 17, 2950-2965.
Reshef, R., Maroto, M., Lassar, A.B., 1998. Regulation of dorsal somitic cell fates: BMPs and Noggin control the timing and pattern of myogenic regulator expression. Genes Dev. 12, 290-303.
Rhodes, S.J., Konieczny, S.F., 1989. Identification of MRF4: a new member of the muscle regulatory factor gene family. Genes Dev. 3, 2050-2061.
Roth, J.F., Shikama, N., Henzen, C., Desbaillets, I., Lutz, W., Marino, S., Wittwer, J., Schorle, H., Gassmann, M., Eckner, R., 2003. Differential role of p300 and CBP acetyltransferase during myogenesis: p300 acts upstream of MyoD and Myf5. EMBO J. 22, 5186-5196.
Saitoh, O., Fujisawa-Sehara, A., Nabeshima, Y., Periasamy, M., 1993 Expression of myogenic factors in denervated chicken breast muscle: isolation of the chicken Myf5 gene. Nucleic Acids Res. 21, 2503-2509.
Seo, H.C., Saetre, B.O., Havik, B., Ellingsen, S., Fjose, A., 1998. The zebrafish Pax3 and Pax7 homologues are highly conserved, encode multiple isoforms and show dynamic segment-like expression in the developing brain. Mech. Dev. 70, 49-63.
Shi, Y., Lee, J.S., Galvin, K.M., 1997. Everything you have ever wanted to know about Yin Yang 1... Biochim. Biophys. Acta. 1332, F49-F66.
Shrivastava, A., Calame, K., 1994. An analysis of genes regulated by the multi-functional transcriptional regulator Yin Yang-1. Nucleic Acids Res. 22, 5151-5155.
Sepich, D.S., Myers, D.C., Short, R., Topczewski, J., Marlow, F., Solnica-Krezel, L., 2000. Role of the zebrafish trilobite locus in gastrulation movements of convergence and extension. Genesis 27, 159-173.
Stern, H.M., Brown, A.M., Hauschka, S.D., 1995. Myogenesis in paraxial mesoderm: preferential induction by dorsal neural tube and by cells expressing Wnt-1. Development 121, 3675-3686. Solnica-Krezel, L., Stemple, D.L., Mountcastle-Shah, E., Rangini, Z., Neuhauss, S.C., Malicki, J.,
Schier, A.F., Stainier, D.Y., Zwartkruis, F., Abdelilah, S., Driever, W., 1996. Mutations affecting cell fates and cellular rearrangements during gastrulation in zebrafish. Development 123, 67-80.
Summerbell, D., Ashby, P.R., Coutelle, O., Cox, D., Yee, S., Rigby, P.W., 2000. The expression of Myf5 in the developing mouse embryo is controlled by discrete and dispersed enhancers specific for particular populations of skeletal muscle precursors. Development 127,
3745-3757.
Tajbakhsh, S., 2003. Stem cells to tissue: molecular, cellular and anatomical heterogeneity in skeletal muscle. Curr Opin Genet Dev. 13, 413-422.
Tajbakhsh, S., Borello, U., Vivarelli, E., Kelly, R., Papkoff, J., Duprez, D., Buckingham, M., Cossu, G., 1998. Differential activation of Myf5 and MyoD by different Wnts in explants of mouse paraxial mesoderm and the later activation of myogenesis in the absence of Myf5. Development 125, 4155-4162.
Tajbakhsh, S., Rocancourt, D., Cossu, G., Buckingham, M., 1997. Redefining the genetic hierarchies controlling skeletal myogenesis: Pax-3 and Myf5 act upstream of MyoD. Cell 89, 127-138.
Tremblay, P., Pituello, F., Gruss, P., 1996. Inhibition of floor plate differentiation by Pax3: evidence from ectopic expression in transgenic mice. Development 122, 2555-2567.
Tremblay, P., Gruss, P., 1994. Pax: genes for mice and men. Pharmacol. Ther. 61, 205-226.
Tzahor, E., Kempf, H., Mootoosamy, R.C., Poon, A.C., Abzhanov, A., Tabin, C.J., Dietrich, S., Lassar, A.B., 2003. Antagonists of Wnt and BMP signaling promote the formation of vertebrate head muscle. Genes Dev. 17, 3087-3099.
Wang, D., Chang, P.S., Wang, Z., Sutherland, L., Richardson, J.A., Small, E., Krieg, P.A., Olson, E.N., 2001. Activation of cardiac gene expression by myocardin, a transcriptional cofactor for serum response factor. Cell 105, 851-862.
Weigel, D., Jäckle, H., 1990. The fork head domain: a novel DNA binding motif of eukaryotic transcription factors? Cell 63, 455-456.
Weigel, D., Jurgens, G., Kuttner, F., Seifert, E., Jackle, H., 1989. The homeotic gene fork head encodes a nuclear protein and is expressed in the terminal regions of the Drosophila embryo. Cell 57, 645-658.
Wright, W.E., Sassoon, D.A., Lin, V.K., 1989. Myogenin, a factor regulating myogenesis, has a domain homologous to MyoD. Cell 56, 607-617.
Xu, D., Yoder, M., Sutton, J., Hromas, R., 1998. Forced expression of Genesis, a winged helix transcriptional repressor isolated from embryonic stem cells, blocks granulocytic differentiation of 32D myeloid cells. Leukemia 12, 207-212.
Yamaguchi, T. P., 1997. New insights into segmentation and patterning during vertebrate somitogenesis. Curr. Opin. Genet. Dev. 7, 513-518.
Yang, J., Mei, W., Otto, A., Xiao, L., Tao, Q., Geng, X., Rupp, R.A., Ding, X., 2002. Repression through a distal TCF-3 binding site restricts Xenopus myf-5 expression in gastrula mesoderm. Mech. Dev. 115, 79-89.
結果圖片之文字說明:
Figure 1. The yeast one-hybrid system was used to identify Foxd3 bound to the myf5 -82/-62 cassette. (A) The yeast one-hybrid assay of clones transfected with the plasmids indicated the
following: pHISi-6×(-82/-62), which contains six repeats of -82/-62; pGADT7-zfoxd3, which contains foxd3 with the GAL4 activation domain; pHISi-m4m5, which contains the mutated -82/-62 cassette; p53HIS/pGAD53, the positive control; p53HIS/pGAD424, the negative control. Yeasts that harbored plasmids containing wild-type -82/-62 grew under growth-inhibiting conditions (histidine and leucine were absent, 60 mM of 3-amino-1,2,4-triazole was present) when Foxd3 fused to the activation domain was expressed. Yeasts containing plasmids with mutated -82/-62 did not grow. (B) The colony-lift filter method was used to perform ß-galactosidase assays. Yeasts were transformed, as indicated, with each of the following plasmids: pLacZi-6×(-82/-62), the bait plasmid that contains six repeats of -82/-62 and carries the lacZ reporter gene; pLacZi-m4m5, which contains four repeats of the mutated -82/-62 sequence; p53BLUE/pGAD53m, the positive control; p53BLUE/pGAD424, the negative control. Positive ß-galactosidase activity, shown in blue, was detected only when the foxd3 activation domain fusion was expressed in yeasts containing wild-type -82/-62. No activity was detected in yeasts with mutated -82/-62.
Figure 2. Binding between purified recombinant Foxd3 and 32P-radiolabeled oligonucleotide probes was studied with the electrophoretic mobility shift assay. The arrow
indicates the shifted band formed by double-stranded oligonucleotide and recombinant Foxd3. Three probes were used: cassette -82/-62, mutated -82/-62 (m4m5, in which the -70/-62 sequence was mutated), and a nonspecific DNA sequence (Non-30fr). Radiolabeled cassette -82/-62, without added nuclear extracts, was the negative control (lane 1). The shifted bands were abolished completely when excess amount of unlabeled -82/-62 oligonucleotide was added (lanes 4 and 5). Recombinant Foxd3 did not bind to m4m5 (lane 8) or Non30fr (lane 9). In addition, m4m5 and Non30fr competitors did not compete for binding with Foxd3 and cassette -82/-62 (lanes 6 and 7).
Figure 3. Transactivation of the myf5 promoter by Foxd3
Cultured cell lines COS-1 and P19 were used to study whether Foxd3 transactivates the expression of zebrafish myf5. In the dual-luciferase assay, luciferase activity was represented as the fold increase compared to the absence of recombinant Foxd3. In COS-1 cells, the luciferase activity increased 1.8-fold (pRL-ZMYP3.0) or 2.4-fold (pRL-ZMYP6.0) when the recombinant Foxd3 was present. In the P19 cell line, the luciferase activity increased 3.65-fold (pRL-ZMYP3.0) or 4.16-fold (pRL-ZMYP6.0) when Foxd3 was present. All transfections and luciferase assays were performed independently at least three times. Data are means + S.D. Asterisks indicate a mean is significantly different (P < 0.05).
Figure 4. Temporal and spatial expression of foxd3 in zebrafish embryos at different somite stages. (A) At the 4- to 6-somite stage, foxd3 was transcribed in the floor plate (FP), presumptive
neural crest cells (NC), and tail bud (TB). (B) At the 7- to 9-somite stage, foxd3 transcripts were first detected in the somites (arrowhead) and the signals were weak in migrating neural crest cells. In addition to the somitic mesoderm, foxd3 mRNA also was found in the tail bud and posterior premigratory crest (PC). (C, D) At the 11- to 16-somite stage, foxd3 transcription increased incrementally after each pair of somites was formed. foxd3 transcripts in the lateral head were down-regulated, but foxd3 expression was strong in the somites and cranial ganglia posterior to the otic vesicle (arrow). (E, F) In the 11- to 13-somite stage of wild-type embryos, double in situ hybridization using red-labeled myf5 and blue-labeled foxd3 probes was used to show that expression of these genes was colocalized in the posterior part of the somites. Scale bars: 100 μm.
Figure 5. The effect of inhibiting Foxd3 protein translation on somitogenesis in zebrafish embryos. Lateral and dorsal views of wild-type (left panels) and foxd3-MO1-injected (right
panels) 12- to 14-somite stage embryos.Embryos that were injected with 10 ng of foxd3-MO1 displayed an abnormal phenotype, including an abnormal tail bud, a reduced head (A vs B), and wider somites with an irregular somite boundary (C vs D). Scale bars: 100 μm.
Figure 6. Whole-mount in situ hybridizations showing gene expression in wild-type (A-F) and the foxd3-MO-injected embryos (G-L) at different somite stages. In embryos with 8 to 19
somites, myf5 expression in the somites and adaxial cells of foxd3-MO1-injected (10ng) embryos was much lower than in wild-type embryos. Weak myf5 signals appeared in somites 0 and -1 (arrowheads) and in presomitic mesoderm. Ectopic expression of myf5 was observed in the tail bud (asterisk in H). By the 20- to 24-somite stage, myf5 expression patterns in foxd3-MO-injected and wild-type embryos were similar (E, F vs K, L). Scale bars: 100 μm.
Figure 7. Effect of inhibiting Foxd3 protein synthesis on myf5, myod, myogenin, and α-actin expression in embryos with 10-12 somites. In foxd3-MO1-injected (10 ng) embryos, myf5
expression in the somites, adaxial cells, and presomitic mesoderm was reduced dramatically (A vs E) but myod expression was unchanged (B vs F). In foxd3-MO1-injected embryos, the expression of myogenin (C vs G) and α-actin (D vs H) was abolished in the somites, except in adaxial cells. Embryos injected with 4 ng of myf5-MO exhibited reduced myogenin expression in the somites, a finding similar to the defective phenotype of foxd3 morphants (I vs G). Co-injection of 600 ng/µl myf5-capped mRNA rescued expression of myogenin (J) and α-actin (K) in foxd3-MO1-injected embryos. Scale bars: 100 μm.
Figure 8. The effect of inhibiting Pax3 protein synthesis on the expression of foxd3, myf5, and myod in the zebrafish embryos at 5 to 8 somites. Probes were used to detect myf5 (A, B,
C), foxd3 (D, E), and myod (F, G, H) in the wild-type embryos (A, D, F), the pax3-MO1-injected (6 ng) embryos (B, E, G), and the embryos co-injected with 6 ng of pax3-MO1 and 25 pg of foxd3 mRNA (C, H). (B) In pax3-MO1-injected embryos, myf5 expression was restricted in the presomitic mesoderm (PSM) and weak in somites 0 and -1 and in the adaxial cells on the sides of somites 0 and -1. (E) The shape of the neural plate became abnormal in the pax3-MO1 morphants, and foxd3 expression was weak in their neural fold (arrow) and tail bud (arrowhead). (G) Meanwhile, myod expression was down-regulated in the somites, but it was expressed in adaxial
cells. In the rescue experiment, co-injection of foxd3 mRNA and pax3-MO1 restored myf5 expression in the somites (C), whereas the myod expression was not rescued (H). Scale bars: 100 μm.
Figure 9. Models of the myf5 regulation network during somitogenesis in mice and zebrafish.
(a) myod functions downstream of pax3 (Tajbakhsh et al., 1997). (b) pax3 activates myod indirectly (Relaix et al., 2003).