Water stress, ammonium, and leaf senescence in detached rice leaves
Jaw-Neng Lin & Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (*Corresponding author)
Received 26 November 1997; accepted in revised form 12 March 1998
Key words: ammonium, leaf senescence, Oryza sativa, water stress
Abstract
Ammonium accumulation in relation to water stress-promoted senescence of detached rice leaves was investi-gated. The effect of water stress on the senescence of detached rice leaves is associated with the accumulation of ammonium. The accumulation of ammonium in detached rice leaves by water stress is attributed to a decrease in glutamine synthetase activity. Ammonium accumulation in detached rice leaves, induced by water stress, was accompanied by an increase in tissue sensitivity to ethylene which, in turn, accelerated leaf senescence.
Abbreviations: ABA = abscisic acid; GS = glutamine synthetase; RWC = relative water content; STS = silver
thiosulphate
1. Introduction
Water stress has many influences on nitrogen metabolism [3]. Nitrate reductase activity decreases with drought stress [21, 22]. In the leaves of bean, water stress is correlated with a decline of the activity of GS (EC 6.3.1.2) [2]. Decline in activity of GS in leaves by water stress may result, at least in part, in an accumulation of ammonium in leaves. Water stress has been shown to enhance leaf senescence [6, 11, 21]. Recently, we reported that ammonium accumula-tion is associated with ABA-promoted senescence of detached rice leaves [10]. We also reported that water stress resulted in a 4-fold increase in endogenous ABA in detached rice leaves 8 h after the start of incubation [7]. It appears that ammonium accumulation may also be associated with water stress-promoted senescence of detached rice leaves. In this study, the possible involvement of ammonium in the regulation of water stress-promoted senescence of detached rice leaves was investigated.
2. Materials and methods
Rice (Oryza sativa cv. Taichung Native 1) was cul-tured as previously described [14]. The apical 3 cm segments excised from the third leaves of 12 d old seedlings were used. A group of 10 segments was floated in a Petri dish containing 10 mL of test solu-tions. Incubation was carried out at 27 ◦C in light (40 µmol quanta m−2s−1). Water stress was applied by a previously described method [6]. Briefly, leaf seg-ments were exposed to the vapor above a solution of 0.5 M NaCl. Water stress was judged by RWC [19] and proline accumulation.
Proline was extracted and its concentration deter-mined by the method of Bates et al. [1]. Leaf segments were homogenized with 3% sulphosalicylic acid and the homogenate was centrifuged. The supernatant was treated with acetic acid and acid ninhydrin, boiled for 1 h and then absorbance at 520 nm was determined. Levels of proline are expressed as µmol g−1 initial fresh weight.
Leaf senescence was followed by measuring the decrease of chlorophyll and protein. Chlorophyll was determined according to Wintermans and De Mots [24] after extraction in 96% (v/v) ethanol. For
pro-tein determination, leaf segments were homogenized in 50 mM sodium phosphate buffer (pH 6.8). The extracts were centrifuged at 17,600 g for 20 min and the supernatant liquids were used for determination of protein by the Bradford method [5]. Protein level was expressed as mg−1g−1initial fresh weight.
Ammonium was extracted by homogenizing leaf segments in 0.3 mM sulphuric acid (pH 3.5). The homogenate was centrifuged for 10 min at 39,000 g and the supernatant was used for determination of ammonium as described previously [16]. Ammonium level was expressed as µmol g−1initial fresh weight. For nitrate determination, leaf segments were homog-enized with double distilled water. The homogenate was centrifuged for 25 min at 17,600 g. The super-natant was used for determination of nitrate by the method described by Hecht and Mohr [13]. Nitrate level was expressed as µmol g−1initial fresh weight.
For extraction of GS, leaf segments were homog-enized with 10 mM Tris-HCl buffer (pH 7.6, con-taining 1 mM MgCl2, 1 mM EDTA and 10 mM
2-mercaptoethanol) in a chilled pestle and mortar. The homogenate was centrifuged at 15,000 g for 30 min and the resulting supernatant was used for determina-tion of GS activity. The whole extracdetermina-tion procedure was carried out at 4◦C. GS was assayed by the method of Oaks et al. [20]. One unit of GS activity is defined as 1 µmol L-glutamate-γ -monohydroxamate formed per min.
When STS was used as a source of Ag+, leaf segments were treated with STS and test solution. A stock solution of STS was prepared by mixing equal volumes of 0.01 M AgNO3and 0.04 M Na2S2O3[17].
All experiments were repeated three times and within each experiment, treatments were replicated 4 times. Similar results and identical trends were obtained each time. The data reported here are from a single experiment.
3. Results and discussion
RWC of leaf segments exposed to the vapor above a 0.5 M solution of NaCl, with light exposure, decreased rapidly for 24 h, but subsequently remained unchanged (Figure 1). Proline accumulates in leaves under water stress [12]. Under our experimental con-dition, we also observed that the level of proline increased markedly in leaf segments exposed to the vapor above a 0.5 M solution of NaCl (Figure 1). Based on the results of RWC and proline level, we
Figure 1. Changes with time in RWC and proline level of detached
rice leaves. Values are averages with standard errors (n = 4).
conclude that the treatment in our study did indeed cause water stress.
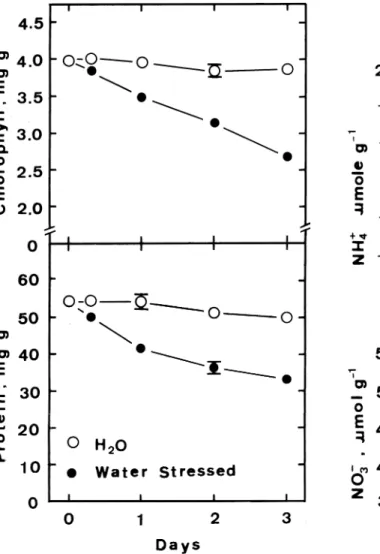
Figure 2 shows the time course of changes in amounts of chlorophyll and protein in leaf segments floated on water or subjected to water stress in light. In control leaf segments, the chlorophyll and protein con-tents showed little change during incubation in light. Levels of chlorophyll and protein fell significantly as a result of water stress. The senescence of detached rice leaves was followed by measuring the decrease in protein [10]. Clearly, our results are consistent with those by other investigators [11, 21] who reported that water stress enhanced leaf senescence.
Figure 3 shows the time course of changes in leaves of ammonium and nitrate, and the activity of GS in detached leaves floated on water or subjected to water stress in light. Ammonium in control leaves remained unchanged during the first day of light
incu-Figure 2. Changes with time in chlorophyll and protein levels
of detached rice leaves. Values are averages with standard errors (n = 4).
bation and slightly increased thereafter. It is clear that water stress significantly increases ammonium level in detached rice leaves. In plant leaves, ammonium is produced during nitrate assimilation, deamination of amino acids and photorespiration [18]. Figure 3 shows that water stress resulted in a decrease in nitrate level. This result suggests that water stress-induced ammonium accumulation may have resulted from the promotion of the reduction of nitrate. If nitrate is assumed to be the source of ammonium, then detached rice leaves ought to accumulate more ammonium if fed with additional nitrate. However, detached rice leaves pretreated with 50 mM KNO3for 24 h in light
con-tained almost the same level of ammonium as those pretreated with 50 mM KCl (Table 1). Thus, nitrate
Figure 3. Changes with time in ammonium and nitrate levels,
and GS activity of detached rice leaves. Values are averages with standard errors (n = 4).
Table 1. Effect of KNO3 pretreatment on water stress-induced
ammonium accumulation in detached rice leaves
Treatment NH+4 (µmol g−1)
KCL→ arrow 14.0± 0.5
KNO3→ water 13.7± 0.6
KCl→ water stress 24.8± 0.8
KNO3→ water stress 26.8± 0.6
Detached rice leaves were pretreated with either 50 mM KCl or KNO3for 24 h and then treated with either water or water stress in the light. Data represent averages and standard errors, n = 4.
is unlikely to be the source of ammonium in water stressed detached rice leaves. Furthermore, water stress has been shown to decrease nitrate reductase activity [21, 22]. Therefore, it is unlikely that water stress-induced ammonium resulted from the reduction of nitrate.
It has been shown that GS activity in leaves decreases with water stress [2]. Since GS is the pri-mary enzyme responsible for ammonium assimilation in plants [18], decreased GS activity by water stress may result, at least in part, in an accumulation of ammonium in leaves. We observed that water stress-treated rice leaves had a lower GS activity than the control leaves (Figure 3). It appears that water stress-induced ammonium accumulation is also attributed to the decrease in GS activity.
Inhibition of GS activity in detached rice leaves by water stress may also result in lowering the levels of glutamine and glutamic acid. Since addition of gluta-mine and glutamic acid had no effect on water stress-promoted senescence (data not shown), it appears that the lack of glutamine and glutamic acid is unlikely to be the reason for the promotion of senescence by water stress.
Previously, we proposed that senescence of detached rice leaves is mediated through an increase in tissue sensitivity to ethylene [15]. Recently, we provided evidence showing that ABA-, jasmonate-, and NH4Cl-promoted senescence is mediated through
increasing tissue sensitivity to ethylene [10, 23]. Water stress has been shown to increase endogenous ABA [7] and ammonium level in detached rice leaves (Figure 3). It is most likely that ammonium accumu-lation induced by water stress may change ethylene sensitivity of detached rice leaves, which in turn results in senescence promotion. This possibility was tested by using the ethylene action inhibitor, Ag+[4]. In the present study, STS was used as a source of Ag+. The effects of STS on water stress-promoted
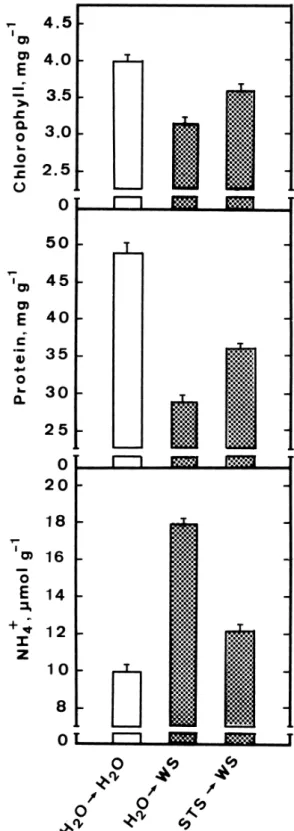
Figure 4. Effect of STS on water stress-promoted senescence and
ammonium accumulation of detached rice leaves. Leaf segments were pretreated with either water or STS (0.8 mM) for 12 h and then treated with either water or water stress for 24 h in the light. Values are averages with standard errors (n = 4).
senescence and ammonium accumulation of detached rice leaves are shown in Figure 4. STS was observed to inhibit, partially, water stress-promoted senescence and ammonium accumulation of detached rice leaves. In conclusion, the results of the present study provide further evidence to support our previous con-clusion that ammonium accumulation is associated with senescence of detached rice leaves [8–10].
Acknowledgements
This work has been financially supported by the National Science Council of the Republic of China. Part XXXXII of the series “Senescence of Rice Leaves”.
References
1. Bates LS, Waldren RP and Teare ID (1973) Rapid determi-nation of free proline for water stress studies. Plant Soil 39: 205–207
2. Becker TW, Hoppe M and Foch PH (1986) Evidence for the participation of dissimilatery processes in maintaining high carbon fluxes through the photosynthetic carbon reduction and oxidation cycles in water-stressed bean leaves. Photosyn-thetica 20: 153–157
3. Beevers L (1976) Nitrogen Metabolism in Plants. Great Britain: Edward Arnold Publishers
4. Beyer SM Jr (1976) A potent inhibitor of ethylene action in plants. Plant Physiol 58: 268–271
5. Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254 6. Chen CT and Kao CH (1990) Comparative study of the
metabolism of 1-aminocyclopropane-1-carboxylic acid and senescence of water-stressed and ABA-treated excised rice leaves. Plant Cell Physiol 31: 463–468
7. Chen CT and Kao CH (1993) Osmotic stress and water stress have opposite effects on putrescine and proline production in excised rice leaves. Plant Growth Regul 13: 197–202 8. Chen SJ and Kao CH (1996) Ammonium accumulation in
relation to senescence of detached maize leaves. Bot Bull Acad Sin 37: 255–259
9. Chen SJ and Kao CH (1997) Methyl jasmonate, ammonium, and leaf senescence in rice. J Plant Physiol (in press) 10. Chen SJ, Hung KT and Kao CH (1997) Ammonium
accumula-tion is associated with senescence of rice leaves. Plant Growth Regul 21: 195–201
11. Dwivedi S, Kar M and Mishra D (1979) Biochemical changes in exised leaves of Oryza sativa subjected to water stress. Physiol Plant 45: 35–40
12. Hanson AD and Hitz WD (1982) Metabolic responses of mesophytes to plant water deficits. Annu Rev Plant Physiol 33: 163–203
13. Hecht U and Mohr H (1990) Factors controlling nitrate and ammonium accumulation in mustard (Sinapis alba) seedlings. Physiol Plant 78: 379–387
14. Kao CH (1980) Senescence of rice leaves IV. Influence of benzyladenine on chlorophyll degradation. Plant Cell Physiol 21: 1255–1262
15. Kao CH and Yang SF (1983) Role of ethylene in the senes-cence of detached rice leaves. Plant Physiol 73: 881–885 16. Lin CC and Kao CH (1996) Disturbed ammonium assimilation
is associated with growth inhibition of roots in rice seedlings caused by NaCl. Plant Growth Regul 18: 233–238
17. Liu J, Mukherjee I and Reid DM (1990) Adventitious rooting in hypocotyls of sunflower (Helianthus annus L.) seedlings III. The role of ethylene. Physiol Plant 78: 268–276
18. Miflin BJ and Lea PJ (1976) The pathway of nitrogen assimi-lation in plants. Phytochemistry 15: 873–885
19. Mukherjee SP and Choudhuri MA (1983) Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol Plant 58: 166–170
20. Oaks A, Stulen J, Jones K, Winspear MJ and Boesel IL (1980) Enzymes of nitrogen assimilation in maize roots. Planta 148: 477–484
21. Shah CB and Loomis RS (1965) Ribonucleic acid and protein metabolism in sugar beet during drought. Physiol Plant 18: 240–254
22. Smith FW and Thompsons JF (1971) Regulation of nitrate reductase in excised barley roots. Plant Physiol 48: 219–233 23. Tsai F-Y, Hung KT and Kao CH (1996) An increase in
ethylene sensitivity is associated with jasmonate-promoted senescence of detached rice leaves. J Plant Growth Regul 15: 197–200
24. Wintermans JFGM and De Mots A (1965) Spectrophotometric characteristics of chlorophyll a and b and their pheophytins in ethanol. Biochem Biophys Acta 109: 448–453