Abstract.
20-Fluoro-6,7-methylenedioxy-2-phenyl-4-quino-lone (CHM-1) has been reported to induce cell cycle
arrest and apoptosis in many types of cancer cells.
However, there is no available information to show CHM-1
affecting DNA damage and expression of associated repair
genes. Herein, we investigated whether or not CHM-1
induced DNA damage and affected DNA repair gene
expression in U-2 OS human osterogenic sarcoma cells.
The comet assay showed that incubation of U-2 OS cells
with 0, 0.75, 1.5, 3 and 6 μM of CHM-1 led to a longer
DNA migration smear (comet tail). DNA gel electrophoresis
showed that 3 μM of CHM-1 for 24 and 48 h treatment
induced DNA fragmentation in U-2 OS cells. Real-time
PCR analysis showed that treatment with 3 μM of CHM-1
for 24 h reduced the mRNA expression levels of ataxia
telangiectasia mutated (ATM), ataxia-telangiectasia and
Rad3-related (ATR), breast cancer 1, early onset (BRCA1),
14-3-3sigma (14-3-3σ), DNA-dependent serine/ threonine
protein kinase (DNA-PK) and O
6-methylguanine-DNA
methyltransferase (MGMT) genes in a time-dependent
manner. Taken together, the results indicate that CHM-1
caused DNA damage and reduced DNA repair genes in
U-2 OS cells, which may be the mechanism for
CHM-1-inhibited cell growth and induction of apoptosis.
Much evidence has shown that DNA damage is implicated
in age-related diseases. For example, DNA single-strand
breaks (SSBs) in neocortex of people with Alzheimer disease
(AD) are two-fold higher than in the controls (1). Moreover,
cortical neurons in AD patients contain DNA SSBs and DNA
double-strand breaks (DSBs) in situ (2). It is also reported
that DNA damage is involved in apoptosis of tumor cells (3).
Quinolone derivatives such as 2-phenyl-4-quinolones have
been shown to induce cytotoxicity in many human cancer
cell lines and to inhibit platelet aggregation (4-9);
synthesized 2-phenyl-4-quinolone series compounds
inhibited tubulin polymerization and acted as anti-mitotic
agents (4-8); and synthesized
2-phenylpyrroloquinolin-4-ones inhibited the growth of hepatocellular carcinoma in
vitro and in vivo (10). Recently,
20-fluoro-7-methylenedioxy-2-phenyl-4-quinolone (CHM-1) has been shown to act as an
anti-invasive agent in hepatocellular carcinoma cells (11).
However, there is no available information to address
whether CHM-1 induces DNA damage in U-2 OS human
osterogenic sarcoma cells. Therefore, in this study, we
investigated the effects of CHM-1 on DNA damage and
DNA repair genes in U-2 OS cells.
Correspondence to: Jing-Gung Chung, Department of Biological
Science and Technology, China Medical University, No 91, Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C. Tel: +886 422053366 ext. 2161, Fax: +886 422053764, e-mail: [email protected]
Key Words: CHM-1, DNA damage, DNA repair, gene expression,
U-2 OS human osteogenic sarcoma cells.
The Novel Quinolone CHM-1 Induces DNA
Damage and Inhibits DNA Repair Gene Expressions
in a Human Osterogenic Sarcoma Cell Line
HUNG-YI CHEN
1, HSU-FENG LU
2, JAI-SING YANG
3, SHENG-CHU KUO
4, CHYI LO
5,6, MEI-DUE YANG
7,
TSAN-HUNG CHIU
8, FU-SHIN CHUEH
9, HENG-CHIEN HO
10, YANG-CHING KO
11,12and JING-GUNG CHUNG
13,14Schools of
1Pharmacy,
5Nursing and
6Chinese Medicine, Departments of
3Pharmacology,
10
Biochemistry and
13Biological Science and Technology,
4
Graduate Institute of Pharmaceutical Chemistry, China Medical University, Taichung 404, Taiwan, R.O.C.;
2Department of Clinical Pathology, Cheng Hsin General Hospital, Taipei 112, Taiwan, R.O.C.;
Departments of
7Surgery and
8OBS/GYN, China Medical University Hospital, Taichung 404, Taiwan, R.O.C.;
11
Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine,
St. Martin De Porres Hospital, Chiayi 600, Taiwan, R.O.C.;
12
Department of Nursing, Chung Jen College of Nursing,
Health Sciences and Management, Chiayi 622, Taiwan, R.O.C.;
Departments of
9Health and Nutrition Biotechnology and
Materials and Methods
Cell culture. The U-2 OS human osterogenic sarcoma cell line was
obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan). U-2 OS cells were plated onto 75 cm2
tissue culture flasks with 90% McCoy’s 5a medium (Gibco BRL, Grand Island, NY, USA). The cell medium with 2 mM L-glutamine was adjusted to contain 10% fetal bovine serum (FBS; Gibco BRL/Invitrogen, Grand Island, NY, USA), and 1% penicillin-streptomycin (100 Units/ml penicillin and 100 μg/ml penicillin-streptomycin) and grown at 37˚C under a humidified 5% CO2atmosphere (12).
Flow cytometric assay for the percentage of viable human osterogenic sarcoma cells. Approximately 2×105cells/well of U-2
OS cells in 12-well plates were incubated with CHM-1 at final concentrations of 0, 0.75, 1.5, 3 and 6 μM, vehicle (1 μl dimethyl sulfoxide; DMSO) and 5 μM of H2O2 (positive control) for 24 hours, or cells were treated with 3 μM of CHM-1 for 0, 12, 24, 36 and 48 hours. Cells in 5 ml tubes from each treatment were stained with propidium iodide (PI, 5 μg/ml) and immediately analyzed by flow cytometry (Becton-Dickinson, San Jose, CA, USA) for the percentage of viable cells as previously described (13-14).
Comet assay for examining DNA damage in U-2 OS cells. U-2 OS
cells (2×105/well) in 12-well plates with cell medium were
incubated with CHM-1 at final concentrations of 0, 5, 10, 25 and 30 μM, vehicle (1 μl DMSO) and 5 μM of H2O2(positive control) grown at 37°C in 5% CO2and 95% air. At the end of incubation, cells were harvested for the examination of DNA damage using the comet assay as described elsewhere (13-14). Briefly, glass slides were pre-coated with 1% agarose, about 3×104cells per gel for each
treatment were centrifuged (500×g, 5 min at 4˚C) and the cell pellet was then suspended in 170 μl of warm (37˚C) 0.5% agarose and two 80 μl aliquots placed onto a glass slide which placed in lysis buffer (100 mM EDTA, 2.5 M NaCl, 10 mM Tris–HCl and 1% Triton X-100, adjusted to pH 10 with NaOH) for 2 hours. Then slides were washed twice with ice-cold deionised water and were transferred to
an electrophoresis tank which contained cold electrophoresis buffer (300 mM NaOH, 1 mM EDTA, pH 13) and incubated for 20 min to allow unwinding of the DNA. Then electrophoresis was carried out at 30 V and 300 mA for 20 min before slides were removed and flooded with neutralization buffer (0.4 M Tris-HCl, pH 7.5), and rinsed twice with deionised water. Slides were stained with 1 ml of PI (2.5 μg/ml) for 20 min (15). Comets were visualized and photographed by use of a fluorescence microscope as previously described (16-17). PI-stained DNA tails in the individual nucleus were quantified by TriTek Comet Score V 1.5 software (TriTek Crop., Sumerduck, VA, USA).
DNA gel electrophoresis for examining DNA damage in U-2 OS cells. U-2 OS cells (1×106/well) in 6-well plates with cell medium
were incubated with 3 μM of CHM-1 for 24 and 48 hours. At the end of incubation, cells were harvested in phosphate-buffered saline (PBS) by centrifugation and were lysed in a digestion buffer containing 0.5% sarkosyl, 0.5 mg/ml proteinase K, 50 mM Tris-HCl Table I. Primers used in real-time PCR in this study. The DNA sequence
was evaluated using Primer Express software.
Primer name Primer sequence
Human ATM F TTTACCTAACTGTGAGCTGTCTCCAT R ACTTCCGTAAGGCATCGTAACAC Human ATM F GGGAATCACGACTCGCTGAA
R CTAGTAGCATAGCTCGACCATGGA Human BRCA1 F CCAGGGAGTTGGTCTGAGTGA
R ACTTCCGTAAGGCATCGTAACAC Human 14-3-3σ F GCCATGGACATCAGCAAGAA
R GGCTGTTGGCGATCTCGTA Human DNA-PK F CCAGCTCTCACGCTCTGATATG
R CAAACGCATGCCCAAAGTC Human MGMT F CCTGGCTGAATGCCTATTTCC
R TGTCTGGTGAACGACTCTTGCT Human GAPDH F ACACCCACTCCTCCACCTTT
R TAGCCAAATTCGTTGTCATACC Each assay was conducted at least twice to ensure reproducibility. F, Forward; R, reverse.
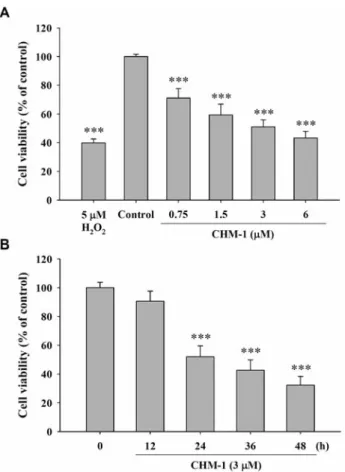
Figure 1. CHM-1 affected the percentage of viable human osterogenic
sarcoma U-2 OS cells. U-2 OS cells (2×105cells/well) were placed in 12-well plates and were incubated with CHM-1 at final concentrations of 0, 0.75, 1.5, 3 and 6 μM, vehicle (1 μl DMSO) and 5 μM of H2O2 (positive control) for 24 hours (A), or cells were treated with 3 μM CHM-1 for 0, 12, 24, 36 and 48 hours (B). Cells from each treatment were stained with propidium iodide (5 μg/ml) and analyzed by flow cytometry (Becton-Dickinson, San Jose, CA, USA) as previously described. *** p<0.001 Compared to untreated control.
(pH 8.0) and 10 mM EDTA at 55˚C overnight. Cells were then treated with 0.5 μg/ml RNase A for 2 hours at 37˚C. The genomic DNA was extracted by phenol-chloroform-isoamyl alcohol extraction (25:24:1) as previously described (14). The extracted DNA from each treatment was resuspended with 50 μl TBE buffer. Approximately 1 μg/μl (12 μl) of DNA was loaded into each well and DNA gel electrophoresis was performed at 50 V for 90 min using 2% agarose. After ethidium bromide staining, the cells then were photographed under fluorescence light as previously described (14, 16-17).
Real-time PCR of ATM, ATR, BRCA1, 14-3-3σ, DNA-PK and MGMT in U-2 OS cells. U-2 OS cells (1×106/well) in 6-well plates were
incubated with 3 μM of CHM-1 for 24 hours. At the end of incubation, cells were harvested in PBS by centrifugation then the total RNA from each sample was extracted by using the Qiagen RNeasy Mini Kit (Qiagen Inc., Valencia, CA, USA) as described previously (15, 18). RNA samples were reverse-transcribed for 30 min at 42˚C with High Capacity cDNA Reverse Transcription Kit according to the standard protocol of the supplier (Applied Biosystems, Foster City, CA, USA). The quantitative PCR from each sample was performed as
follows: 2 min at 50˚C, 10 min at 95˚C, and 40 cycles of 15 s at 95˚C, 1 min at 60˚C using 1 μl of the cDNA reverse-transcribed as described above, 2X SYBR Green PCR Master Mix (Applied Biosystems) and 200 nM of forward and reverse primers as shown in Table I. Each assay was run on an Applied Biosystems 7300 Real-Time PCR system in triplicates and expression fold-changes were derived using the comparative CTmethod (19).
Statistical analysis. Student’s t-test was used to analyze differences
between exposure to CHM-1 and the untreated (control) group. All data are presented as the means±SD of three experiments and
p<0.05 was considered significantly.
Results
CHM-1 reduced the percentage of viable U-2 OS cells. The
U-2 OS cells were exposed to different concentrations of
CHM-1 for 24
hoursor were treated with 3 μM of CHM-1
for different time periods. The cells were collected for the
determination of percentage of viable U-2 OS cells and the
Figure 2. CHM-1-induced DNA damage in U-2 OS cells was examined by comet assay. U-2 OS cells (2×105cells/well; 12-well plates) were incubated with different concentrations of CHM-1 for 24 hours or cells were treated with 3 μM of CHM-1 for different time periods and DNA damage was determined by comet assay as described in the Materials and Methods. Representative images of cells are shown in panel A and C, and panel B and D show comet length (fold). *p<0.05 Compared to untreated control.
results are shown in Figure 1A and B, which indicate that
CHM-1 reduced the percentage of viable cells and these
effects took place in a dose- and time-dependent manner.
CHM-1-induced DNA damage in U-2 OS cells as examined
by comet assay. In earlier studies, it had been shown that
CHM-1 induced cytotoxic effects on U-2 OS cells (20). In
the present study, we investigated whether or not CHM-1
induced DNA damage in U-2 OS cells. The results from the
comet assay are shown in Figure 2 and indicate that CHM-1
induced DNA damage in U-2 OS cells. These effects are
dose dependent (Figure. 2A and B). However, the long
incubation of U-2 OS cells with 3 μM of CHM-1 led to a
longer DNA migration smear (comet tail) (Figure 2C and D),
indicating that CHM-1 induced DNA damage in U-2 OS
cells in a time-dependent manner.
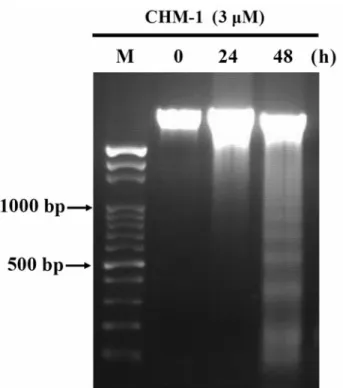
CHM-1-induced DNA fragmentation in U-2 OS cells was
determined by DNA gel electrophoresis. DNA fragmentation
is a characteristic of apoptosis (21). Here, we isolated DNA
from U-2 OS cells after treatment with 3 μM of CHM-1 for
24 and 48 hours before DNA was used for agarose gel
electrophoresis. The results shown in Figure 3 indicate that
CHM-1 induced DNA damage and fragmentation in U-2 OS
cells, which also indicated the occurrence of apoptosis
(Figure 3). The longer time (48 hours) of incubation of U-2
OS cells with CHM-1 led to more DNA damage and
fragments than that of short time (24 hours) incubation. This
finding suggests that CHM-1 appears to significantly induce
apoptosis of U-2 OS cells.
CHM-1 inhibited the relative expression levels of DNA damage
and repair genes in U-2 OS cells as shown by real-time PCR.
For investigating whether CHM-1 affected DNA damage and
repair gene expressions, U-2 OS cells were treated with 3 μM
CHM-1 for 0 and 24
hours. Total RNA was isolated from each
sample and associated gene expressions were examined by
real-time PCR (Figure 4). Expression levels of ATM, ATR,
BRCA1, 14-3-3σ, DNA-PK and MGMT mRNA were decreased
on 24
hourstreatment when compared with the control group.
Discussion
In our primary studies, we have demonstrated that CHM-1
induced cell cycle arrest and apoptosis in many human
cancer cell lines (20, 22). However, there is no available
information to show CHM-1 affected DNA damage and
DNA repair associated gene expression in U-2 OS cells.
Herein, we used the comet assay (single-cell gel
electrophoresis) to measure the levels of DNA damage from
U-2 OS cells after exposure to different concentrations of
CHM-1 for various time periods. It is well-documented that
the comet assay is a highly sensitive technique for DNA
damage examination (19, 23-25). The results showed that
CHM-1 induced a significant increase in the tail moment of
the comets of U-2 OS cells. H
2O
2was used as positive
control and showed significant tail movement.
Figure 3. CHM-1-induced DNA fragmentation in U-2 OS cells was
examined by DNA gel electrophoresis. U-2 OS cells were incubated with 3 μM CHM-1 for 0, 24 and 48 hours, the cells were harvested and DNA was extracted from each treatment before DNA fragmentation was examined by DNA gel electrophoresis as described in the Materials and Methods.
Figure 4. CHM-1-inhibited expression of DNA damage and repair genes in
U-2 OS cells were examined by real-time PCR. The total RNA was extracted from the U-2 OS cells after treatment with 3 μM of CHM-1 for 0 and 24 hours. RNA samples were reverse-transcribed cDNA and real-time PCR carried out as described in the Materials and Methods. The experiments of ATM, ATR, BRCA-1, 14-3-3σ, DNA-PK and MGMT genes related to GAPDH are presented. Data represent the mean±SD of three experiments. *p<0.05 and ***p<0.001 compared to untreated control.
These findings are in agreement with other reports which
showed that CHM-1 induced DNA damage in human
osterogenic sarcoma cells (20) and murine colorectal
adenocarcinoma cells (22). Other reports already showed that
strand-break formation during the process of excision repair
may also cause DNA migration measurable in the comet
assay (26-27). In cells, DNA repair can reduce DNA damage
by eliminating DNA lesions. Results (Figure 3) from DNA
gel electrophoresis demonstrated that CHM-1 indeed induced
DNA fragmentation (apoptosis) in U-2 OS cells.
Actually, in our primary studies, we had already
documented the cytotoxic effects of CHM-1 on other cells
such as osterogenic sarcoma cells (20) and osterogenic
sarcoma and, including U-2 OS cells (Figure. 1). Our earlier
studies also showed that the reduction of cancer cell numbers
by CHM-1 may be achieved through the induction of
apoptosis or by antiproliferative effect (20, 22). Our results
also demonstrated that CHM-1 inhibited expression of DNA
repair genes such as ATM, ATR, BRCA1, 14-3-3σ, DNA-PK
and MGMT (Figure 4) in examined U-2 OS cells. Further
studies are needed to establish the role of the interaction of
CHM-1 with DNA in carcinogenesis.
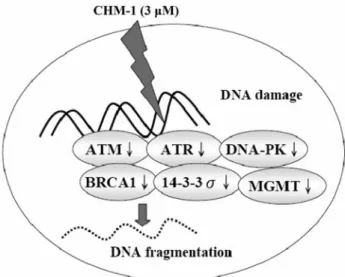
In conclusion, the CHM-1 itself may induce DNA damage
in U-2 OS cells via the inhibition of expression of DNA
repair genes such as ATM, ATR, BRCA1, 14-3-3σ, DNA-PK
and MGMT, subsequent to DNA damage (Figure 5).
Acknowledgements
This work was supported by grant CMU98-S-04 from China
Medical University, Taichung, Taiwan, R.O.C.
References
1 Mullaart E, Boerrigter ME, Ravid R, Swaab DF and Vijg J: Increased levels of DNA breaks in cerebral cortex of Alzheimer's disease patients. Neurobiol Aging 11: 169-173, 1990.
2 Adamec E, Vonsattel JP and Nixon RA: DNA strand breaks in Alzheimer's disease. Brain Res 849: 67-77, 1999.
3 Button M, Jenkin GR, Bowman KJ, Harrington CF, Brewer TS, Jones GD and Watts MJ: DNA damage in earthworms from highly contaminated soils: Assessing resistance to arsenic toxicity by use of the comet assay. Mutat Res 696: 95-100, 2010. 4 Kuo SC, Lee HZ, Juang JP, Lin YT, Wu TS, Chang JJ, Lednicer D, Paull KD, Lin CM, Hamel E and Lee KH: Synthesis and cytotoxicity of 1,6,7,8-substituted 2-(4’-substituted phenyl)-4-quinolones and related compounds: identification as antimitotic agents interacting with tubulin. J Med Chem 36: 1146-1156, 1993. 5 Li L, Wang HK, Kuo SC, Wu TS, Lednicer D, Lin CM, Hamel E and Lee KH: Antitumor agents. 150. 2’,3’,4’,5’,5,6,7-Substituted 2-phenyl-4-quinolones and related compounds: their synthesis, cytotoxicity, and inhibition of tubulin polymerization. J Med Chem 37: 1126-1135, 1994.
6 Xia Y, Yang ZY, Xia P, Bastow KF, Tachibana Y, Kuo SC, Hamel E, Hackl T and Lee KH: Antitumor agents. 181. Synthesis and biological evaluation of 6,7,2’,3’,4’'-substituted-1,2,3,4-tetrahydro-2-phenyl-4-quinolones as a new class of antimitotic antitumor agents. J Med Chem 41: 1155-1162, 1998.
7 Xia Y, Yang ZY, Xia P, Hackl T, Hamel E, Mauger A, Wu JH and Lee KH: Antitumor Agents. 211. Fluorinated 2-phenyl-4-quinolone derivatives as antimitotic antitumor agents. J Med Chem 44: 3932-3936, 2001.
8 Chen YC, Lu PH, Pan SL, Teng CM, Kuo SC, Lin TP, Ho YF, Huang YC and Guh JH: Quinolone analogue inhibits tubulin polymerization and induces apoptosis via Cdk1-involved signaling pathways. Biochem Pharmacol 74: 10-19, 2007. 9 Huang LJ, Hsieh MC, Teng CM, Lee KH and Kuo SC: Synthesis
and antiplatelet activity of phenyl quinolones. Bioorg Med Chem
6: 1657-1662, 1998.
10 Ferlin MG, Chiarelotto G, Gasparotto V, Dalla Via L, Pezzi V, Barzon L, Palu G and Castagliuolo I: Synthesis and in vitro and in
vivo antitumor activity of 2-phenylpyrroloquinolin-4-ones. J Med
Chem 48: 3417-3427, 2005.
11 Wang SW, Pan SL, Peng CY, Huang DY, Tsai AC, Chang YL, Guh JH, Kuo SC, Lee KH and Teng CM: CHM-1 inhibits hepatocyte growth factor-induced invasion of SK-Hep-1 human hepatocellular carcinoma cells by suppressing matrix metalloproteinase-9 expression. Cancer Lett 257: 87-96, 2007. 12 Lu KH, Lue KH, Chou MC and Chung JG: Paclitaxel induces
apoptosis via caspase-3 activation in human osteogenic sarcoma cells (U-2 OS). J Orthop Res 23: 988-994, 2005.
13 Chen JC, Lu KW, Tsai ML, Hsu SC, Kuo CL, Yang JS, Hsia TC, Yu CS, Chou ST, Kao MC, Chung JG and Wood WG: Gypenosides induced G0/G1 arrest via CHk2 and apoptosis through endoplasmic reticulum stress and mitochondria-dependent pathways in human tongue cancer SCC-4 cells. Oral Oncol 45: 273-283, 2009.
14 Lee JH, Li YC, Ip SW, Hsu SC, Chang NW, Tang NY, Yu CS, Chou ST, Lin SS, Lino CC, Yang JS and Chung JG: The role of Ca2+in baicalein-induced apoptosis in human breast
MDA-MB-231 cancer cells through mitochondria- and caspase-3-dependent pathway. Anticancer Res 28: 1701-1711, 2008.
Figure 5. The possible signaling pathway for CHM-1-reduced
expression of DNA damage and repair genes in a human osterogenic sarcoma cell line (U-2 OS).
15 Banerjee B, Vadiraj HS, Ram A, Rao R, Jayapal M, Gopinath KS, Ramesh BS, Rao N, Kumar A, Raghuram N, Hegde S, Nagendra HR and Prakash Hande M: Effects of an integrated yoga program in modulating psychological stress and radiation-induced genotoxic stress in breast cancer patients undergoing radiotherapy. Integr Cancer Ther 6: 242-250, 2007.
16 Button M, Jenkin GR, Bowman KJ, Harrington CF, Brewer TS, Jones GD and Watts MJ: DNA damage in earthworms from highly contaminated soils: Assessing resistance to arsenic toxicity by use of the Comet assay. Mutat Res 2009.
17 Ip SW, Liao SS, Lin SY, Lin JP, Yang JS, Lin ML, Chen GW, Lu HF, Lin MW, Han SM and Chung JG: The role of mitochondria in bee venom-induced apoptosis in human breast cancer MCF7 cells. In Vivo 22: 237-245, 2008.
18 Lin JG, Chen GW, Li TM, Chouh ST, Tan TW and Chung JG: Aloe-emodin induces apoptosis in T24 human bladder cancer cells through the p53-dependent apoptotic pathway. J Urol 175: 343-347, 2006.
19 Lin SS, Huang HP, Yang JS, Wu JY, Hsia TC, Lin CC, Lin CW, Kuo CL, Gibson Wood W and Chung JG: DNA damage and endoplasmic reticulum stress mediated curcumin-induced cell cycle arrest and apoptosis in human lung carcinoma A-549 cells through the activation caspases cascade- and mitochondrial-dependent pathway. Cancer Lett 272: 77-90, 2008.
20 Hsu SC, Yang JS, Kuo CL, Lo C, Lin JP, Hsia TC, Lin JJ, Lai KC, Kuo HM, Huang LJ, Kuo SC, Wood WG and Chung JG: Novel quinolone CHM-1 induces apoptosis and inhibits metastasis in a human osterogenic sarcoma cell line. J Orthop Res 27: 1637-1644, 2009.
21 Hansen CM, Hansen D, Holm PK, Larsson R and Binderup L: Cyanoguanidine CHS 828 induces programmed cell death with apoptotic features in human breast cancer cells in vitro. Anticancer Res 20: 4211-4220, 2000.
22 Chou LC, Yang JS, Huang LJ, Wu HC, Lu CC, Chiang JH, Chen KT, Kuo SC and Chung JG: The synthesized 2-(2-fluorophenyl)-6,7-methylenedioxyquinolin-4-one (CHM-1) promoted G2/M arrest through inhibition of CDK1 and induced apoptosis through the mitochondrial-dependent pathway in CT-26 murine colorectal adenocarcinoma cells. J Gastroenterol 44: 1055-1063, 2009. 23 Ashby J, Tinwell H, Lefevre PA and Browne MA: The single
cell gel electrophoresis assay for induced DNA damage (comet assay): measurement of tail length and moment. Mutagenesis 10: 85-90, 1995.
24 Pool-Zobel BL, Lotzmann N, Knoll M, Kuchenmeister F, Lambertz R, Leucht U, Schroder HG and Schmezer P: Detection of genotoxic effects in human gastric and nasal mucosa cells isolated from biopsy samples. Environ Mol Mutagen 24: 23-45, 1994. 25 Donatus IA, Sardjoko and Vermeulen NP: Cytotoxic and
cytoprotective activities of curcumin. Effects on paracetamol-induced cytotoxicity, lipid peroxidation and glutathione depletion in rat hepatocytes. Biochem Pharmacol 39: 1869-1875, 1990. 26 Tice RR, Andrews PW and Singh NP: The single-cell gel assay:
a sensitive technique for evaluating intercellular differences in DNA damage and repair. Basic Life Sci 53: 291-301, 1990. 27 Olive PL, Banath JP and Durand RE: Detection of etoposide
resistance by measuring DNA damage in individual Chinese hamster cells. J Natl Cancer Inst 82: 779-783, 1990.