一個新的調控 Serratia marcescens 表面移行及毒素因子訊
號傳遞系統之研究
計畫類別: 個別型計畫 計畫編號: NSC92-2314-B-002-356- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學醫學院醫事技術學系暨研究所 計畫主持人: 賴信志 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 12 月 22 日
Regulates Swarming Motility and Cell Envelope Architecture in Response to
2
Exogenous Saturated Fatty Acids
3 4
Hsin-Chih Lai1,2,*, Po-Chi Soo1, Jun-Rong Wei1, Wen-Ching Yi1, Shwu-Jen Liaw1,2,
5
Yu-Tze Horng1, Shi-Ming Lin3, Shen-Wu Ho1,2, Simon Swift4 and Paul Williams5
6 7
1School and Graduate Institute of Medical Technology, National Taiwan University
8
College of Medicine, Taipei, Taiwan, R.O.C. 2Department of Laboratory Medicine,
9
National Taiwan University Hospital and National Taiwan University College of 10
Medicine, Taipei, Taiwan, R.O.C. 3Center for Optoelectronic Biomedicine, National
11
Taiwan University College of Medicine. 4Division of Molecular Medicine, Faculty
12
of Medical and Health Sciences, University of Auckland, Auckland, New 13
Zealand.5Institute of Infection, Immunity and Inflammation, Centre for Biomolecular
14
Sciences, University of Nottingham, Nottingham, NG7 2RD, United Kingdom 15
16 17
*For correspondence. Dr. Hsin-Chih Lai, Graduate Institute of Medical Technology,
18
College of Medicine, National Taiwan University, No.1. Chan-Der Street, Taipei 100, 19
Taiwan (R.O.C.). 1 Tel. +886 2 2397 0800 (ext 6931) 2 Fax +886 2 2371 1574 3 Email: [email protected] 4 5
Running title: Swarming regulation in Serratia marcescens. 6
Keywords: Serratia marcescens, two-component system, swarming motility, fatty 7
acids. 8
ABSTRACT 1
Serratia marcescens swarms at 30°C, but not at 37°C on a nutrient rich (LB)
2
agar surface. Mini-Tn5 mutagenesis of S. marcescens CH-1 yielded a mutant (WC100) 3
that swarms not only vigorously at 37°C, but also earlier and faster than the parent 4
strain at 30°C. Analysis of this mutant revealed that the transposon had inserted into a 5
gene (rssA) predicted to encode a bacterial two component signal transduction sensor 6
kinase, upstream of which, a potential response regulator gene (rssB) was located. 7
rssA and rssB insertion-deletion mutants were constructed through homologous
8
recombination and both exhibited similar swarming phenotypes on LB swarming agar 9
whereby swarming not only occurs at 37°C, but also initiates at a lower cell density, 10
on a surface of higher agar concentration and more rapidly than the parent strain at 11
30°C. Both mutants also exhibited increased haemolysin activity and altered cell 12
surface topologies compared with the parent CH-1 strain. Temperature and certain 13
saturated fatty acids (SFAs) were found to negatively regulate S. marcescens 14
swarming via the action of RssA-RssB. Analysis of the respective fatty acid profiles 15
of the parent, rssA and rssB mutants grown at 30oC or 37oC and different nutrition
16
conditions revealed a relationship between cellular fatty acid composition and 17
swarming phenotypes. The cellular fatty acid profile is further observed to be affected 18
by RssA and RssB. SFA-dependent inhibition of swarming was also observed in 19
Proteus mirabilis and Salmonella typhimurium, suggesting that either SFAs per se or
1
the modulation of cellular fatty acid composition and hence homeostasis of membrane 2
fluidity may be a conserved mechanism for regulating swarming motility in Gram 3
negative bacteria. 4
INTRODUCTION 1
When inoculated onto LB medium solidified with 0.8% w/v agar at 30°C, 2
Serratia marcescens exhibits a characteristic swarming phenotype in which short
3
motile vegetative rods at the colony margin differentiate into elongated, aseptate and 4
hyperflagellate swarm cells which migrate coordinately and rapidly away from the 5
colony (11). Phenotypically, the process of swarming cell differentiation and 6
migration in S. marcescens may be divided into two separate phases: (i) the lag period 7
prior to the onset of swarming behavior and the induction of swarm cell 8
differentiation at the colony edge and (ii) active motile swarming migration (or 9
translocation) from the colony edge. Individual bacterial cells do not possess the 10
intrinsic ability to swarm as swarming behavior in Serratia is a result of the 11
coordinated, multicellular effort of groups ofdifferentiated cells functioning through 12
close cell-cell interactions(11). 13
Swarming is the result of the regulated expression of gene networks required to 14
initiate the complex processes underlying the required morphological and 15
physiological changes (11, 13, 21, 22). Development of a Serratia surface swarming 16
colony requires the processingand integration of multiple environmental, cell-to-cell 17
and intracellularsignals involving surface contact and local high bacterial population 18
densities (2). The flagellar master operon (flhDC) and an N-acylhomoserine 19
lactone-dependent quorum-sensing systemare global regulators of flagellar motility 1
and cell population density, respectively (14, 24, 33, 35). Cellular differentiation is 2
only one partof this process which requires a lag period prior to thecommencement 3
of swarmingmigration, during which time cellular proliferation occurs up to the 4
required population density, and a large amount of biosurfactant was produced (1, 24, 5
32). An additional layer of regulation of surface migration may also be exerted 6
through the post-transcriptional regulator RsmA (5). When over-expressed, rsmA 7
inhibits the formation of a spreadingcolony in S. marcescens (4) and this effect may 8
be mediated via repression of AHL-dependent quorum-sensing, since overexpression 9
of rsmA in P. aeruginosa has been reported negatively regulate AHL synthase 10
expression (38). 11
Many regulatory pathways involved in control of swarming remain have yet to 12
be characterized at the molecular level. For example, exposure of the cells to surfaces 13
with a specific viscosity is likely to require recognition by an unknown sensor. 14
Furthermore, the physiological signals that provide essential stimuli for the initiation 15
of swarming and the underlying signal transduction pathways have also not yet been 16
characterized. S. marcescens swarming is known to be a temperature-dependent 17
behaviour, which occurs at 30°C, but not at 37°C. Bacteria inoculated onto identical 18
plates and incubated at 37°C did not initiate swarming even after 48h culture and 19
formed only small defined colonies. This may be a consequence of reduced flagellar 1
motility (35) and/or a reduction of biosurfactant production following temperature 2
upshift. To begin unravelling the underlying regulatory mechanism(s), we have 3
utilized mini-Tn5 mutagenesis to screen a group of S. marcescens mutants that 4
demonstrated swarming proficiency at 37°C. In the process of characterizingone of 5
these mutants, we identified a bacterial two-component signal transduction system 6
(23) which we termed RssA-RssB. This two component system functions in the 7
coordination of swarmingmigration which is independent of surface flhDC flagellar 8
motility, AHL-dependent quorum-sensing, and the production of biosurfactants, 9
factors which have previously been identified as regulators of population surface 10
migration behavior in Serratia (15, 24). Here we present evidence to show that 11
swarming of S. marcescens is inhibited by presence of saturated fatty acids (SFAs) 12
and that this effect is mediated via the RssA-RssB two component system. The 13
cellular fatty acids composition, haemolysin production and cell surface topography 14
were also shown to be regulated by RssA-RssB. The negative regulatory effect of 15
myristic acid on bacterial swarming was also observed in bacteria such as Salmonella 16
typhimurium and Proteus mirabilis, suggesting existence of a common regulatory
17
pathway in Gram negative bacterial swarming. 18
MATERIALS AND METHODS
1
Bacterial strains, plasmids and culture conditions. S. marcescens CH-1 is a
2
clinical isolate and is functionally wild-type forswimming motility and swarming 3
migration behavior. No AHL quorum sensing signals were detected from CH-1 cells. 4
S. marcescens CH-1∆A and CH-1∆B are rssA and rssB knock-out mutants,
5
respectively, via the insertion of an Ω cassette (Smr) from pHP45Ω (39) and
6
subsequent homologous recombination. Bacterial strains Salmonella typhimurium 7
SJW1103 and Proteus mirabilis P19 are both wild-type for motility and chemotaxis 8
and were gifts from R. Macnab (Yale University) and C. Hughes (Cambridge 9
University), respectively. E. coli strains (JM109; CC118 λpir ; S17-1 λpir; Top10 F’ 10
and XL-1 Blue) were cultured at 37 oC and S. marcescens at 30 oC in L-broth (LB)
11
medium (Difco) (40) unless otherwise indicated in the text. M9 salt solution (40) was 12
used to make minimal growth medium (MGM), into which 0.8% Eiken agar was 13
incorporated to make MGM plates. In cases of mixed cultures, e.g., conjugations, 14
incubations were at 30oC.
15
The pUT::mimi-Tn5-Km1 construct (10) was used in the transposon mutagenesis 16
assays for screening super-swarming mutants. pZErO2.1 (Kmr), pCR2.1 (Apr; 17
Kmr )(Invitrogen), pBCSK+ (Cmr) and pBluescript SKII+ (Stratagene) constructs 18
were used as DNA fragment cloning vectors. pPC300(PflhDC::luxCDABE) and 19
pSA400(PshlBA::luxCDABE) constructs, used as bioreporters for monitoring the 20
promoter activity of PflhDC and PshlBA, respectively, were generated by insertion 21
of a PCR amplified promoter region upstream of luxCDABE, derived from pSB1075 1
(47), into the pACYC184(Tcr; Cmr) vector (9). The PCR primer pairs were: 2 5’GATATCCAGCCTCAGGCGGAGGG3’/5’GGATCCATTCCCCATCCCGACAG 3 ACTA3’ (PflhDCSm) and 4 5’GCAGCAGCGCCGGTATAAGCAC3’/5’ATCGCCAGCGCAGCGGCCAGTT3’ 5
(PshlBA). pWC200 (Tcr, Cmr ) was constructed by PCR cloning of a 1766 bp DNA 6
fragment containing full length rssA into a pACYC184 vector, the expression of 7
which was driven by its native promoter. Full length rssA PCR was via the primer 8
pair 5’ACCATTATTTTCCAGGTGCT3’/5’ATACAGAGTGTCGATAATTT3’. The 9
pWC201 (Cmr) was constructed by cloning a 1014 bp DNA fragment containing the 10
complete rssB structural gene into pBCSK(Stratagene) which was also expressed via 11
its native promoter. The rssB primers were: 12
5’TGCGGCCTGCGCGCAGGCGC3’/5’AGAATATTGGCGATGCCTGC3’. 13
pWC202 containing the complete 2.5 Kb rssB-rssA genes amplified by the primer pair 14
5’TGCGGCCTGCGCGCAGGCGC3’/5’ATACAGAGTGTCGATAATTT3’. 15
Swimming motility for S. marcescens was examined on motility agar [LB 16
solidified with 0.35% Eiken agar (Eiken, Japan)] by sterile needlepoint inoculation 17
from an overnight culture into the center of the agar plate. Swarming motility for S. 18
marcescens was examined on swarming agar plates (LB solidified with 0.8% Eiken
19
agar) by inoculating 5 µl of an overnight broth culture onto the center of the agar plate, 20
with some modifications for S. typhimurium SJW1103 and P. mirabilis P19 described 21
in the figure legends. Swimming motility and swarming migration distances were 22
recorded at hourly intervals to enable comparisons. Swarm cell differentiation i.e., the 23
overproduction of flagella, cellular elongation and polyploidy, was also examined 24
microscopically as described (29, 35). Briefly, for cell elongation assays, after 25
overnight culture, bacteria were diluted 1:100 and cultured in 20ml LB broth at 30oC 1
and 37oC with vigorous shaking at 200 rpm. An Olympus BH2 light microscope was 2
used for observation of individual bacterial cells at 1000 X magnification. Images 3
were captured using an SPOT CCD camera integrated with a PC computer 4
(Diagnostics Instruments). Bacterial colonies were examined under light microscopy 5
at a magnification of 1X or 30X using an Olympus SZH microscope. For determining 6
bacterial growth rates, hourly increases in the optical density of broth cultures at 7
600nm were measured. 8
Enzymes and chemicals. DNA restriction and modification enzymes were
9
purchased from Roche (Germany). Taq polymerase and PCR-related products were 10
obtained from either Perkin Elmer (USA) or Takara Biomedicals (Japan). Other 11
laboratory grade chemicals were purchased from Sigma (USA), Merck (Germany) or 12
BDH (UK). 13
Recombinant DNA techniques. Unless otherwise indicated, standard protocols
14
were used for DNA/DNA hybridization, plasmid and chromosomal DNA preparation, 15
transformation, electroporation, PCR, restriction digestion, agarose gel electrophoresis, 16
DNA recovery from agarose gels, DNA ligation (40), and conjugation (10). Southern 17
blotting analysis of chromosomal DNA was performed using nylon membranes 18
(HybondN+; Amersham) and a DIG High Prime labeling kit according to the 19
recommendations of the manufacturer(Roche). PCR DNA amplicons were cloned by 20
pCRTMII® and the TA Cloning Kit (Invitrogen). DNA sequencing and analysis was 21
performed using a Perkin-Elmer Autosequencer model 377 with a Taq DyeDeoxyTM 1
terminator cycle sequencing kit (Applied Biosystems). The DNA sequences of PCR 2
products were confirmed by sequencing both strands from two or three independent 3
reactions. 4
Analysis of DNA and protein sequences. Deduced DNA and protein sequences
5
were compared with GenBank DNA or non-redundant protein sequence databases, 6
respectively, using blastn or blastx via the NCBI internet homepage 7
(http://www.ncbi.nlm.nih.gov/). Protein sequence identities were analyzed by ExPASy 8
proteomics tools (DAS, Tmpred, SOSUI, PredictProtein and ProtScale) in the 9
ExPASy Molecular Biology Server of the Swiss Institute of Bioinformatics (SIB) 10
(http://tw.expasy.org). 11
Screening of super-swarming mutants by Mini-Tn5 mutagenesis. For the
12
effective transfer of the pUT-miniTn5-Km1 recombinant plasmid from E. coli to S. 13
marcescens CH-1 by conjugation (10), the S. marcescens CH-1 recipient strain and E.
14
coli S17-1 λ-pir donor strain, carrying the pUT derivative recombinant plasmid, were
15
grown overnight with vigorous shaking at 30oC in 10 ml LB broth, with the addition
16
of streptomycin 50 µgml-1 and kanamycin 50 µgml-1 for E. coli cultures. Mating was
17
achieved by mixing 100µl of each bacterial suspension together, followed by the 18
addition of 5 ml of 10 mM MgSO4. The mixture was then filtered through a Type HA
Filter membrane (Millipore) using a negative-pressure pump (Stratagene). The 1
drained membrane was subsequently placed onto the agar surface of a normal 1.2% 2
LB plate and incubated for 8-18 hour at 30oC. The bacteria were then suspended in 5
3
ml MgSO4 broth, that can be kept at 4 oC for one week, and spread onto modified LB
4
agar plates (0.04% NaCl, 2% glycerol, 0.5% yeast extract, 1% Bacto-tryptone, 0.8% 5
Eiken agar, 50µgml-1 kanamycin and 13µgml-1 tetracycline) followed by incubation at
6
37 oC. Transconjugants that exhibited swarming migration at 37 oC were then selected.
7
A total of 6000 colonies were screened, from which 17 were finally selected. Southern 8
blot hybridization using labeled Km gene as a probe was performed to confirm 9
insertion of only one transposon copy in the mutants. 10
Construction of S. marcescens CH-1∆A and CH-1∆B insertion deletion
11
mutants. A PCR protocol was designed to introduce specific HindIII sites into rssA
12
and rssB genes for subsequent insertion of a HindIII digested Ω (Sm r) gene cassette
13
(39). The Primer pairs 14 5’GGTTTACGATCGAGACAACC3’/5’GGATCCGTGCGGGCGATCTG3’ (rssA) 15 and 16 5’GCATTGGAGCTGGCCGGCTTTA3’/5’GGATCCAAAGTCGTTCGGGCTGT 3’ 17
(rssB) were used to amplify the 5’-region of the gene to be inactivated. PCR 18
products were T-cloned into pCR2.1 (Stratagene), sub-cloned as an XbaI/BamHI 19
fragment into pZero2.1 (Invitrogen) and excised as a XbaI/HindIII fragment. A 1
second PCR product encompassing the 3’-region of the gene to be inactivated was 2
generated using primer pairs 3 5’GGATCCGCTCGACGGCAGCGAGGA3’/5’TCCGGCAAATCGATGATGAAGC 4 C3’ and 5 5’GGATCCCATCAGGTCATGCTGGATA3’/5’CTACTCTTTCTTCAGCAAATAGC 6
C3’ for rssA and rssB, respectively, and was T-cloned into pCR2.1 and excised as an 7
HindIII/EcoRI fragment. The 2 kb Sm-resistant Ω DNA fragment was excised from
8
pHP45Ω (39) as a HindIII fragment. The three DNA fragments were ligated to the 9
XbaI/EcoRI digested suicide vector pUT-mini-Tn5-Km (10). The resultant
10
pUT-rssA::Sm and pUT-rssB::Sm vectors were selected by streptomycin resistance in 11
the permissive E. coli strain CC118 and verified by restriction enzyme mapping. For 12
gene inactivation mutagenesis by homologous recombination, the respective plasmids 13
were transferred from the permissive host strain E. coli S17-1 pir to S. marcescens 14
CH-1 by conjugation and the transconjugants were spread on LB plates with 15
streptomycin (100 µg/ml) and tetracycline (13 µg/ml). Mutants with double cross over 16
events were selected by colony PCR screening. Southern hybridization using PCR 17
amplified rssA or rssB genes as probes was performed to confirm the mutant 18
genotypes (data not shown). The data confirmed that a double-crossover event had 19
occurred and the new strains were hence designated as CH-1∆A and CH-1∆B strains. 1
Complementation of super-swarming mutants. Transfer of pWC200,
2
pWC201, pWC202 and control plasmid into S. marcescens CH-1∆A or CH-1∆B 3
mutant via electroporation tested the effect of overexpressing rssA, rssB or rssB-rssA 4
in the mutant backgrounds for complementation assays. Transformants that were Cmr
5
were selected for further characterization of swarming and cell differentiation 6
behavior. 7
Detection of luciferase activity. The Autolumat LB 953 luminometer (EG&G,
8
Germany) with the 'replicates' program was used for bioluminescence measurement. 9
All procedures followed the protocols supplied by the manufacturer. 10
Measurement of hemolysin activity, surfactant production and flagellum
11
production. Cell-associated haemolysin (ShlA) activity was assayed as described (27)
12
and calculated in arbitrary hemolytic units (1 unit causing the release of 50 mg 13
hemoglobin/hr in the standard assay). A qualitative assay for surfactant production 14
was performed using the drop collapsing method (34). A semi-quantitative assay for 15
biosurfactant was performed using a TLC based assay as described by Matsuyama et 16
al., (37). To quantify the amount of flagellin, cells from surfaces of agar plates were
17
harvested by washing with LB and normalized to a fixed cell mass [OD(A600nm) x 18
cell suspension volume(ml) = 3]. Whole cell lysates were separated by SDS-PAGE 19
(40) followed by flagellin detection using anti Salmonella typhimurium polyclonal 1
antibodies (ViroStat) and ECL kit (Amersham). 2
Atomic force microscopy(AFM). Bacteria were cultured overnight in LB
3
medium and diluted 1:100 in fresh LB and incubated at 37℃ with vigorous shaking 4
(225rpm) for 2 hr. Prior to imaging, bacteria were gently washed with distilled water 5
and final concentrations of 104-105 cells/ml was used for AFM experiments.
6
Pre-cleaned slides were treated with poly-L-lysine 0.01% and left to dry. A 20µl 7
drop of bacterial suspension in distilled water was applied onto a treated slide. After 8
adsorption for 30 minutes, distilled water was added to remove the unadsorbed cell. 9
AFM experiments were performed using SOLVER BIO atomic force microscopy 10
(NT-MDT, Moscow, Russia). The procedures for AFM imaging are described in 11
Hansma et al. (20). Silicon nitrite tips were used, with a force constant of 5.5 N/m. 12
Imaging was carried out on dry samples. AFM images were generated at line 13
frequencies of between 1 and 3 Hz, with 256 lines per image. Images were obtained 14
using semi-contact (tapping) mode AFM. The data were analyzed with SMENA 15
software (Advanced Technologies Center, Russia). 16
Analysis of cellular fatty acids by GC-FAME. Both swarming and
17
non-swarming bacteria were seeded onto plates under growth conditions described in 18
the Results section. Extraction of cellular fatty acids was performed by standard 19
procedures (MIDI, USA). Samples were prepared for analysis by the MIDI 20
GC-FAME analysis system (Microbe Inotech Laboratories, USA). Three independent 1
experiments were performed and results obtained were averages of three experiments 2
(SEM< 10%). 3
RESULTS
1 2
A Serratia marcescens mutant defective in both temperature and temporal
3
control of swarming migration. To characterize the genetic determinants of
4
temperature-dependent regulation of swarming behavior in S. marcescens CH-1, 5
mini-Tn5 transposon mutagenesis experiments were performed to screen for mutants 6
that swarmed on modified LB plates at 37°C. A total of 17 out of 6000 7
transconjugants that showed such temperature-independent swarming behavior, were 8
obtained. One of these mutants, designated S. marcescens WC100, was selected for 9
further characterization. When inoculated on a 0.8% w/v or even a 1% w/v agar 10
swarming plate and incubated at 37oC, WC100 in contrast to the parent strain CH-1 11
exhibited swarming motility. This suggested that the temperature-dependent 12
regulation of swarming behavior had been abolished in this mutant. 13
Analysis of the locus which the transposon interrupted. Conventional
14
digestion and cloning a 3009-bp DNA fragment of the S. marcescens CH-1 genome 15
followed by sequencing with primers designed from within the I or O end of the 16
transposon, respectively (10) were used to identify S. marcescens WC100 DNA 17
flanking the mini-Tn5 insertion site. This analysis revealed that the mini-Tn5 insertion 18
in WC100 was within a 1407bp open reading frame (ORF), designated orf1, encoding 19
a putative 469-amino-acid (52.8 kDa) polypeptide with a calculated isoelectric point 1
of 5.45. Upstream of orf1 we identified an additional ORF, orf2, encoding a putative 2
219-amino-acid (24.7 kDa) in the same direction. Upstream of orf2 was a divergent 3
ORF, orf3, encoding a putative 231-amino-acid protein (Fig. 1A). 4
The deduced protein sequences were compared with non-redundant protein 5
sequence databases using blastn or blastx, via the NCBI internet homepage. 6
Computer-assisted homology searches using BLASTP (3)of protein sequences stored 7
in the DDBJ/EMBL/GenBank databanks indicatethat ORF1 is highly homologous to 8
members of the two-component sensor kinase family of proteins, including E. coli 9
EvgS (22% identity,41% similarity)(12). Orf2 shares homology to two-component 10
response-regulator elements, including a 39 % identity and 57% homology throughout 11
the alignment to a two-component response regulator PmrA of Pectobacterium 12
carotovorum subsp. carotovorum (25).
13
Orf1 and Orf2 were further observed to contain the domain regions highly 14
conserved in two-component sensor kinase and response regulator domains, 15
respectively. Domains suchas the H box (Met-242 to Val-257) (a phosphoaccepter 16
domain) and motifs N (Leu-353 to Arg-373), D/F (Gly-385 to Gly-414) and G 17
(Thr-415 to Lys-443) thatare associated with the sensor proteins (48) are maintained 18
withinRssA (Fig. 1B and 1D) (41). The amino acid phosphorylation domain including 19
conserved Asp-8, 9, 51 (the putative phosphorylation site) and Lys-101 and the 1
DNA-binding domain (42) which are typical of OmpR family proteins can be 2
identified in RssB (Fig. 1C and 1D). The results therefore suggest that RssA and RssB 3
may function as a typical sensory histidine protein kinase and response regulator pair 4
in S. marcescens CH-1. Orf3 shows high homology to a potential oxido-reductase 5
YgfF in E. coli (64% identity, 78% homology) (8). We have designated the gene 6
encoded by orf1, ‘rssA’, forregulation of Serratia swarming A. orf2 is denoted ‘rssB’ 7
and orf3 as ygfFSm. The nucleotide sequence of 3009bp encoding all three ORFs(rssA,
8
rssB and ygfFSm) has been submitted tothe DDBJ/EMBL/GenBank databases under
9
the accession no. AF465237. 10
Analysis of the deduced amino acid sequence of RssA from the ExPASy 11
proteomics tools at internet homepage http://tw.expasy.org identified two N-terminal 12
hydrophobic regions (Lys5-Trp33 and Gly162-Arg189), indicating thatRssA may 13
form two membrane helixesand localize as a inner membrane transmembrane protein 14
(Fig. 1D). A HAMP domain, which is predicted to play an active role in 15
transmembrane signal transduction (6), was identified between 33 and 162. 16
Additionally, the phospho-accepting domain was located near the middle of the 17
protein in the cytoplasm, and a predicted ATPase domain is observed between 353 and 18
443 towards the C-terminal (Fig. 1D). The predicted N-terminal phosphorylation 19
domain and carboxyl terminal DNA-binding motif (residues 189–201) of RssB are 1
shown in figure 1D. 2
Construction of rssA and rssB insertion-deletion mutants and
3
characterization of swarming-related phenotypes. Mutations were separately
4
introduced into rssA and rssB in CH-1 cells by insertion deletion via homologous 5
recombination, to form the CH-1∆A and CH-1∆B mutants, respectively. The 6
phenotypes of WC100, CH-1∆A and CH-1∆B mutants were characterized by their 7
growth dynamics, cell elongation pattern, cell densities and temperature-regulated 8
swarming behaviors. No differences were observed between WC100 and S. 9
marcescens CH-1∆A in any of these analyses. Effect of temperature on swarming of
10
CH-1, CH-1∆A and CH-1∆B on LB swarming plates were shown in figure 2A. 11
By analysis of the swarming fronts of S. marcescens CH-1∆A, CH-1∆B and 12
CH-1 at 30°C, using the method of Gygi et al. (18), the swarming behavior and 13
velocity of CH-1∆A, CH-1∆B and CH-1 was monitored. Depending on the moisture 14
content of the plates, CH-1 cells typicallyspent 4-5 h (at 30°C) in the initial lag phase, 15
which was approximately one hour longer than that of the CH-1∆A and CH-1∆B 16
mutants. Following the initiation of swarming, CH-1∆A cells migrate at an 17
increasingly higher velocity than CH-1 cells (Fig. 2B). The average velocity of S. 18
marcescens CH-1 swarming increased from 1 mm/h (between 5 to 6 h) to 3.3 mm/h
(between 7 to 8 h), whereas S. marcescens CH-1∆A increased from 2 mm/h to 5 1
mm/h during the same period (Fig. 2B). Due to a shorter lag phase and higher 2
migration velocity, CH-1∆A mutants therefore translocate a greater distance in the 3
same period than CH-1parental cells. Swarming of CH-1∆A mutants was essentially 4
the same at 37°C as that observed at 30oC (Fig. 2 A and 2B).
5
CH-1∆B exhibited a slower swarming phenotype than CH-1∆A at both 6
temperatures (Fig. 2 A and 2B). Both CH-1∆A and CH-1∆B cells swarmed at an agar 7
concentration of up to 1% w/v at 30°C (Fig. 2C), and on an average measured about 8
10% shorter than that of CH-1 cells following growth on 0.8% w/v LB agar surface 9
(data not shown). When incubatedat 30°C, both CH-1∆A and CH-1∆B cells initiated 10
swarming earlier than CH-1 cells and requiredca. 50-fold fewer cells to commence 11
migration (3.4 × 106 /50mm2 versus 1.7 × 108/50mm2 cells to initiate swarming).
12
Similar cell-population density dependent initiation of swarming for both strains was 13
also observed at 37°C (data not shown). 14
Transformation of plasmid pWC200 (pACYC184::rssA) expressing the rssA 15
gene restored CH-1∆A to the non swaming phenotype characteristic of the parent 16
strain CH-1 incubated at 37°C (data not shown), confirming rssA as the gene 17
responsible for the super-swarming phenotype of CH-1∆A. This observation also 18
suggested that RssA may function as a negative regulatorof swarming at 37°C. On the 19
other hand, rssB-rssA (pWC202, pACYC184::rssB-rssA) in trans, but not rssB 1
(pWC201, pACYC184::rssB) alone, inhibited CH-1∆B swarming at 37°C (data not 2
shown), suggesting rssB-rssA may form an operon and both genes were involved in 3
swarming regulation. 4
Factors such as swimming motility, the flagellar master operon flhDC and 5
biosurfactant which are known to be essential for swarming migration in Serratia 6
were examined to determine whether they are controlled by RssAB. A swimming 7
assay at 37°C showed that the motilities of CH-1, CH-1∆A and CH-1∆B were similar 8
(data not shown). Monitoring of flhDCSm promoter activity by plasmid pPC300
9
(PflhDCSm::luxCDABE) and Western blot analysis for quantification of flagellin
10
production on 0.8% w/v LB agar surfaces also did not identify any measurable 11
differences between mutants and parent (data not shown). These observations indicate 12
that, compared with the parent strain CH-1, expression of the flhDCSm regulon is
13
unaffected in either rssA or rssB mutant cells on swarming agar surfaces. 14
The super-swarming phenotype of CH-1∆A or CH-1∆B cells may be due to 15
over-production of biosurfactant, hence reducing the surface friction on swarming 16
plates. A drop-collapsing test (24) was performed to qualitatively assay the surface 17
tension of stationary phase culture suspensions from CH-1∆A, CH-1∆B and CH-1 18
cells grown on LB swarming plates at 37°C. No reproducible differences were 19
observed, (data not shown). Similar results were obtained using thin layer 1
chromatography (TLC) analysis(36). Taken together, the swarming behaviour of S. 2
marcescens CH-1∆A and CH-1∆B at 37°C is not due to an increase in surface
3
flagellar motility or biosurfactant production. 4
RssAB negatively regulates haemolysin production in S. marcescens. The
5
cell-surface associated haemolysin, ShlA has been identified as a major virulence 6
factor in S. marcescens (28). To determine whether RssAB regulated haemolysin 7
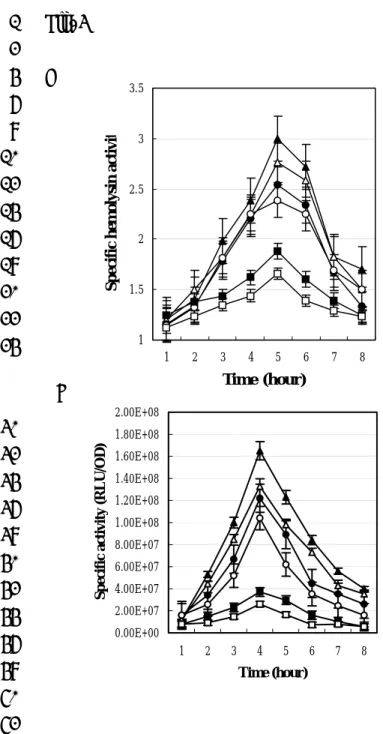
production we assayed both haemolytic activity (Fig. 3A) and shlA expression (Fig. 8
3B). Figure 3A shows that haemolytic activity in CH-1∆A and CH-1∆B cells was 9
higher than in CH-1 cells at both 30°C (1.7-fold and 1.4-fold increase for CH-1∆A 10
and CH-1∆B respectively) and 37°C (1.6-fold and 1.4-fold increase for CH-1∆A and 11
CH-1∆B respectively) after 5 h of growth into the late log phase. Further experiments 12
using a plasmid-borne shlBA::luxCDABE fusion (pSA400) as a reporter for 13
haemolysin gene (shlA) activity revealed a 4 to 8 fold and 3 to 4 fold increase in 14
shlBA promoter activity in CH-1∆A and CH-1∆AB, respectively, compared with the
15
parent strain (Fig. 3B). This data indicate that RssA-RssB is a negative regulator of 16
shlBA promoter activity.
17
RssA and RssB are involved in the determination of cellular surface
18
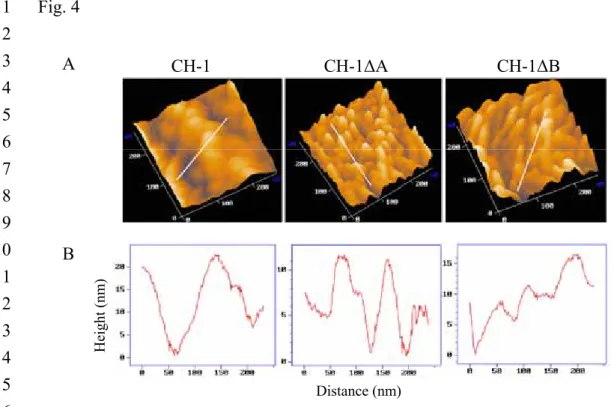
architecture. The detailed cellular surface topography of log phase CH-1, CH-1∆A
and CH-1∆B cells was studied using atomic force microscopy (AFM) by 1
systematically targeting regions within the cellular envelope. A larger scan area (3 x 3 2
µm2) was initially utilized to select suitable cells for higher-resolution imagery. In
3
each of the CH-1, CH-1∆A and CH-1∆B strains, more than twenty individual 4
vegetative cells were examined. Although some variations do exist in the calculation 5
of exact cellular size, a clear trend emerged that, on average, the calculated surface 6
texture parameter RMS (Root-Mean-Square) values for length and width of CH-1, 7
CH-1∆A and CH-1∆B cells were 1981 x 734 nm, 1518 x 555 nm and 1644 x 680 nm, 8
respectively. This was consistent with our previous observation that the average cell 9
dimensions of both CH-1∆A and CH-1∆B mutants were shorter than CH-1 cells under 10
1000X light microscopy. 11
The cell surface topography of both CH-1∆A and CH-1∆B mutants changed 12
when cultured in LB at 37°C (Fig. 4A), and characterization of these changes are 13
shown in figure 4B. CH-1 showed a mean valley-to-peak distance (Rmean) of 14
11.900nm, with a maximal value (Rmax) of 25.900nM, a surface roughness (Ra) of 15
2.652nm, and a root-mean-square (Rq) 3.344nm. Both CH-1∆A and CH-1∆B, 16
however, showed an Rmean of 9.075 and 7.775nm, an Rmax of 17.700 and 14.675nm, 17
an Ra of 1.999 and 1.545 nm, and an Rq of 2.240nm and 1.948nm, respectively. 18
These data suggest that RssA and RssB are either directly or indirectly involved in 19
determination of S. marcescens cell surface topology. 1
Inhibition of S. marcescens swarming by saturated fatty acids via RssA.
2
Although CH-1 cells do not swarm on 0.8% w/v LB agar plates at 37°C, we found 3
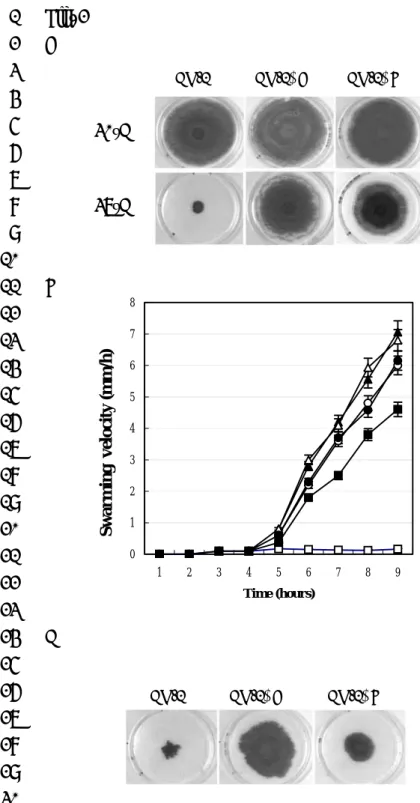
that CH-1 did exhibit ample migratory swarming on defined MGM plates (M9 4
minimal growth medium containing 0.8% Eiken agar) and on MGM plates containing 5
casamino acids (1%) at both 30 and 37°C (Fig. 5A), suggesting that a higher 6
environmental temperature per se is not inhibitory for S. marcescens CH-1 swarming 7
under these culture conditions. We also noted that CH-1 swarming began very quickly 8
after it was transferred from LB broth cultures to either MGM or MGM-casamino 9
acid plates at both temperatures (within 1 h and at a much lower colony cell 10
population density). This observation is very striking when compared with the 11
cell-population density dependent swarming behavior of CH-1 at 30°C and inhibition 12
of swarming at 37°C on normal LB swarming plates, indicating that the absence of 13
specific nutrients in the minimal medium promotes the early appearance of swarming. 14
Swarming assays on MGM-casamino acids (1%)/yeast extract (1%) plates further 15
revealed that whereas CH-1∆A swarmed at both temperatures, CH-1 swarming was 16
completely inhibited at 37°C (Fig. 5A). These observations suggested that some 17
components (or their metabolic derivatives) of yeast extract inhibited parental CH-1 18
cell swarming at 37°C. We therefore examined the effects of amino acids, glucose, 19
mono- and di- saccharides, saturated (SFAs) and unsaturated (UFAs) fatty acids, iron 1
starvation and pH on the swarming behavior of S. marcescens CH-1 (Fig. 5A and data 2
not shown). These data revealed that apart from certain fatty acids, none of the other 3
medium components or environmental conditions influenced the CH-1 swarming. 4
We found that whereas UFAs exerted no significant swarming-inhibition effects on 5
CH-1, swarming was completely inhibited by the addition of lauric or myristic acids 6
and slightly inhibited by the presence of palmitic or stearic acids (Fig. 5A). None of 7
the fatty acids tested showed significant inhibitory effects towards CH-1∆A swarming 8
(Fig. 5A). 9
To determine whether this SFA-dependent phenomenon was dose-dependent, 10
myristic acid at concentrations of 0.01%, 0.005%, 0.0025% and 0.00125% (w/v) was 11
added to MGM-casamino acid plates, followed by swarming assays at 37°C. While 12
there was no effect on CH-1∆A, the swarming lag times of CH-1 cells are delayed 13
with increasing concentrations of myristic acid and swarming was completely 14
inhibited by 0.01% (w/v) myristic acid (Fig. 5B). A similar myristic acid inhibitory 15
effect was also observed at 30°C on both LB swarming plates and MGM-casamino 16
acid plates, with lauric acid also exhibiting similar inhibitory effects (Fig. 5A). 17
Taken together, these data indicate that, in addition to temperature shift, initiation of 18
swarming is regulated by the availability of SFAs in a concentration-dependent 19
manner and such effect is likely to be mediated via RssA. 1
Swarming behavior is closely related to “swarming” or “non-swarming”
2
cellular fatty acid profiles. We postulated that the cellular fatty acid profile of CH-1
3
might be affected by environmental fatty acids which may in turn influence 4
RssA-RssB signaling leading to either a swarming or a non-swarming phenotypes. To 5
this end, the cellular fatty acid profiles (expressed as percentage of the total fatty acids 6
of CH-1 cultured at 37°C) of cells grown under different conditions were evaluated by 7
extraction and analysis using gas chromatography (GC). These growth conditions 8
included swarming conditions (MGM plates and MGM casamino acids (1%) plates) 9
and non-swarming conditions (LB plates and MGM-casamino acids (1%)/yeast 10
extracts (1%) plates). Major observed changes in the profiles of cells grown under 11
non-swarming conditions were increases in the content of lauric acid (12:0), myristic 12
acid (14:0) and palmitic acid (16:00) (Fig. 5C; Table 1). For cells grown in swarming 13
conditions, a concomitant increase in unsaturated fatty acids (especially 18:1 w7c) 14
and branch-chain fatty acids (especially 15:0 anteiso) were observed (Fig. 5C; Table 15
1). These results highlight the link between cellular fatty acid composition and 16
swarming behavior at constant temperatures in S. marcescens. 17
The observed changes in fatty acid profile under different nutrient conditions at 18
37°C that coincide with the swarming or non-swarming behavior of CH-1 supported 19
our hypothesis of a direct correlation between fatty acid profile and swarming 1
regulation. To confirm this finding, we collected both swarming and non-swarming 2
CH-1 cells cultured under different growth conditions for fatty acid profile analysis. 3
CH-1 swarming cells were collected at 30°C from LB swarming plates, 4
MGM-casamino acids (1%)/yeast extract (1%) plates, MGM-casamino acids 5
(1%)-myristoleic acids (0.01%), MGM-casamino acids(1%)-palmitoleic acids (0.01%) 6
and MGM-casamino acids (1%)-oleic acids (0.01%) (30°C), and at 30°C and 37°C 7
from MGM plates and MGM-casamino acids (1%) plates. For CH-1 non-swarming 8
cells, cells were collected at 30°C from MGM-casamino acids (1%)-lauric acid 9
(0.01%) plate, MGM-casamino acids (1%)-myristic acid (0.01%) plate and 10
MGM-casamino acids (1%)-palmitic acid (0.01%) plates, and at 37°C from LB 11
swarming plates. 12
The results are summarized in Table 1 and indicate that for CH-1 cells growing 13
at non-swarming conditions, including either a higher incubation temperature or in the 14
presence of growth nutrients containing SFAs, the ratio of high melting point to low 15
melting point fatty acids (12:0+14:0+16:0+18:0/10:0 3OH+15:0 anteiso+18:1 w7c) is 16
much higher than in cells grown under swarming conditions (Table 1). The percentage 17
of 12:0 showed the most significant difference (Fig. 5C). In contrast, CH-1 cells 18
grown under swarming conditions, including lower incubation temperatures or media 19
lacking SFAs, contain higher percentages of UFAs and branch-chain fatty acids, with 1
18:1 w7c and 15:0 anteiso contents showing the greatest variations (Fig. 5C). The 2
calculated average ratio was 2.7 for swarming cells and 6.2 for non-swarming cells. 3
The fatty acid profiles of the two super-swarming mutants, CH-1∆A and CH-1∆B, 4
inoculated either on LB swarming plates (30°C or 37°C), MGM, MGM-casamino 5
acids or CH-1∆A on MGM-casamino acids-myristic acid (0.01%) (37°C) were 6
basically similar (Table 1). 7
Taken together, these results suggest that the S. marcescens CH-1 cellular fatty 8
acid profile is affected by both temperature and medium fatty acid content and that the 9
profile is closely related to swarming phenotypes, with swarming cells containing 10
relatively more UFAs and branch-chain fatty acids and non-swarming cells containing 11
more SFAs. Thus the two different cellular fatty acid profiles could be classified as 12
“swarming” and “non-swarming” profiles. 13
RssA-RssB regulate cellular fatty acid composition. To determine whether
14
RssA-RssB influence cellular fatty acid composition, the cellular fatty acid 15
compositions of the corresponding mutants was determined by GC. Table 1 shows 16
that the fatty acid profile of CH-1 cells cultured on an LB agar surface at 37°C is 17
significantly different from both CH-1∆A and CH-1∆B swarming cells which showed 18
swarming fatty acid profiles at this temperature. For both mutants, the ratio of 19
saturated- to non- saturated fatty acids is 1.8 and 2.6 respectively whereas it is 5.6 for 1
the parent strain (Table 1). Furthermore, although CH-1, CH-1∆A and CH-1∆B all 2
showed swarming fatty acid profiles at 30°C, their overall fatty acid composition 3
differed (Table 1). Further experiments analyzing the fatty acid profile of the three 4
strains also found variance between them when cultured on MGM media at 37°C 5
(Table 1). These data indicate that, consistent with swarming regulation, cellular fatty 6
acid profiles are regulated via RssA-RssB. 7
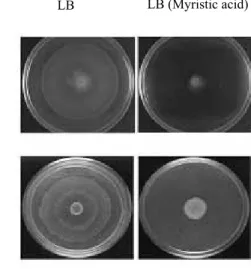
Myristic acid inhibits swarming in both P. mirabilis and S. typhimurium. To
8
determine whether myristic acid also influenced swarming migration in other Gram 9
negative bacteria, P. mirabilis P19 and S. typhimurium LT2 at 37°C were inoculated 10
onto 0.5 % or 2.0 % w/v Eiken agar LB plates supplemented with myristic acid at a 11
concentration of 0.01% (w/v). Figure 6 shows that myristic acid inhibited the 12
swarming of P. mirabilis and S. typhimurium LT2. These results indicate that fatty 13
acid-mediated inhibition of swarming motility is conserved among these Gram 14
negative bacterial genera. 15
DISCUSSION
1
Although there is an extensive literature on bacterial swarming motility, the 2
signals sensed and underlying genetically programmed molecular mechanisms are 3
still far from being fully understood. Here we have identified a two component 4
regulatory system, RssA-RssB which negatively controls swarming in response to 5
SFAs in S. marcescens strain CH-1. Mutation of the sensor, RssA or the response 6
regulator, RssB confers a “vigorous-swarming” phenotype on LB agar. This 7
phenotype is reminiscent of that observed by Belas et al.,(7) and Liaw et al., (31) 8
where precocious swarming mutants of P. mirabilis BB2000 and P19 which were 9
defective in the temporal control of swarming migration initiated swarming some 60 10
min earlier than the parent. In each case, the corresponding mutation was localized to 11
a gene termed rsbA, which codes for a putative sensor kinase. Despite the functional 12
similarities and their homology to the sensor kinase protein family, RsbA is composed 13
of 897 amino acids and RssA is composed of 469 amino acids, and both proteins do 14
not have significant homology or identity following amino acid sequence 15
comparisons, apart from a low percentage of identity within certain short polypeptide 16
sequences. P. mirabilis BB2000 and P19, in contrast to S. marcescens CH-1, swarm 17
on LB plates at 37oC (7, 30), suggesting that RsbA and RssA may not be functionally
18
equivalent at least with respect to their temperature dependency. In E. coli K12 19
swarming is regulated via a locus termed yojN-rcsB-rcsC (44) with the same genetic 1
organization as that found in P. mirabilis i.e. rsbA-rscB-rscC (7) where YojN is a 2
homologue of RsbA. This suggests that P. mirabilis and E. coli may use a common 3
signal transduction pathway to regulate swarming. 4
It is currently not clear whether RssA senses SFAs per se or the change in 5
membrane lipid composition in response to feeding of fatty acids, although the 6
regulatory effect of SFAs on Serratia swarming is identified in this report. If, like E. 7
coli and Salmonella, Serratia takes up fatty acids it is offered and incorporates these
8
into its membranes, this will affect membrane fatty acid composition. Although not 9
clear yet, evidences suggested that changes in cellular fatty acid profile which may 10
either directly or indirectly be involved in altering the ratio of SFAs to UFAs may be 11
sensed by RssA and RssB. For both mutants, the ratio of saturated- to non- saturated 12
fatty acids is 1.8 and 2.6 respectively whereas it is 5.6 for the parent strain when 13
cultured on LB plates at 37 oC (Table 1). These fatty acid profiles correspond to
14
“swarming” and “non-swarming” phenotypes respectively suggesting that changes in 15
membrane fluidity may be sensed via the RssA-RssB system, which may either 16
directly or indirectly be involved in altering the ratio of SFAs to UFAs. Furthermore, 17
these changes in fatty acid profiles and hence swarming phenotype can also be 18
modified by temperature shifts or by the provision of a SFA. For example, in LB at 19
30oC, CH-1 has a “swarming” fatty acid profile i.e. a ratio of saturated- to
1
non-saturated fatty acids of 2.0 whereas at 37 oC, it fails to swarm and has a fatty acid
2
ratio of 5.6 (Table 1). On a minimal medium (MGM plus casamino acids), CH-1 3
swarms at both temperatures but swarming can be inhibited by the exogenous 4
provision of lauric acid or myristic acid. These convert the fatty acid ratio from 2.5 5
(i.e. swarming profile) to 7.2 and 5.1 (i.e. non swarming profiles) respectively. 6
However, myristic acid was unable to inhibit swarming by the rssA mutant at 37oC,
7
which retained a swarming fatty acid profile with a ratio of 3.1. Thus as a putative 8
sensor kinase, RssA is likely to sense changes either in membrane fluidity or the ratio 9
of saturated to unsaturated fatty acids or even variations in the levels of specific fatty 10
acids either exogenously or within the cell envelope. To further clarify SFAs function, 11
a S. marcescens mutant defective in fatty acid transport would have to be constructed 12
and effect of SFAs on its swarming being characterized. 13
Since swarming and haemolysin production are upregulated in both rssA and rssB 14
mutants, it is possible that the response regulator RssB, when phosphorylated via 15
RssA, functions either as a repressor or as an activator of a repressor. RssA in S. 16
marcescens may sense different signaling states under varying growth temperatures
17
and nutritional conditions which lead to changes in membrane fatty acid composition 18
in order to maintain cellular homeostasis. This may be accomplished by regulating the 19
ratio of kinase to phosphatase activities of RssA, such that a kinase-dominant state is 1
present at high growth temperatures or in the case of SFA-rich nutrient availability. 2
RssA is predicted to possess two transmembrane domains and either the periplasmic 3
or cytoplasmic region may function to propagate a conformational change that is 4
sufficient to significantly alter its activity. Such a conformational change is likely to 5
be governed by the physical state of the membrane lipid bilayer. Lipids in biological 6
membranes are usually maintained in a fluid, liquid-crystalline state (46) and the 7
correct physical state of membrane lipids is essential for optimal membrane structure 8
and function. Temperature markedly affects membrane lipid composition in order to 9
maintain an appropriate liquid crystalline state. As the growth temperature increases, 10
the proportion of high-melting-point fatty acids in the membrane lipids increases. 11
This change from an less ordered to an ordered membrane composition may stimulate 12
activation of kinase activity in RssA, resulting in autophosphorylation of the 13
conserved histidine contained within its transmitter domain followed by 14
phosphotransfer to RssB and consequently down-regulation of down-stream genes 15
involved in swarming regulation. While the down-stream genes involved in 16
modulating the S. marcescens cellular fatty acid profile remains to be established, it is 17
conceivable that RssA-RssB is involved in a regulatory loop where either SFAs or 18
temperature upshifts influence swarming behavior via the action of RssA and RssB, 19
which also regulate cellular fatty acid composition. Since swarming migration in P. 1
mirabilis and S. typhimurium, in common with S. marcescens, can be inhibited by
2
SFAs, it is possible that the regulation of swarming through the control of membrane 3
fluidity via a two component system is conserved in Gram negative bacteria. Indeed 4
in S. typhimurium, a number of transposon mutants with defects in putative two 5
component regulatory systems and which exhibit aberrant swarming have been 6
described (45). Amongst these, a mutant defective in the rsbA homologue yjoN 7
showed substantially reduced flagellin levels. 8
An important question that remains is the identity of the molecular mechanism 9
underlying swarming regulation by RssA-RssB. Given that the Serratia rssA and rssB 10
mutants do not exhibit defects in known components of the swarming pathways such 11
as biosurfactant and flhDC, these findings imply the existence of additional signalling 12
pathways involving RssAB. Indeed, given that the average cell lengths of CH-1∆A 13
and CH-1∆B mutants are about 10% shorter than parental CH-1 cells and that 14
flhDCSm expression is not affected, this suggests that cell length determination is also
15
regulated by the RssA-RssB system, independent of flhDCSm. Furthermore, the cell
16
surface topography of rssA and rssB mutants is markedly different to that of the CH-1 17
wild type as revealed by AFM. In P. mirabilis, an acidic capsular polysaccharide 18
(Cmf-CPS) facilitates swarming migration (17). Cmf-CPS is proposed to enhance 19
growth medium surface fluidity by extracting water from the agar soreducing friction 1
and aiding bacterial surface translocation. Capsule synthesis under the control of 2
two-component system is also reported to be involved in regulation of swarming in E. 3
coli K12 (43). In S. marcescens, so far we have no evidence that the extracellular
4
polysaccharide (EPS) has contribution to swarming motility. Further experiments will 5
be required to determine whether RssA-RssB regulates swarming via the control of 6 EPS synthesis. 7 8 ACKNOWLEDGEMENTS 9
This work was supported by grants from the National Science Council (NSC, 10
grants number NSC-91-2314-B-002-258, NSC-92-2314-B-002-356), Technology 11
Development Program for Academia, Ministry of Economical Affairs (grant number 12
91-EC-17-A-10-S1-0013) and Environmental Protection Administration (EPA, grant 13
number EPA-91-NSC-01-B003) of Taiwan and by a joint Royal Society 14
U.K./National Science Council of Taiwan travel project grant which is gratefully 15
acknowledged. 16
Reference List 1
1. Alavi, M. and R. Belas. 2001. Surface sensing, swarmer cell differentiation, 2
and biofilm development. Methods Enzymol. 336:29-40. 3
2. Alberti, L. and R. M. Harshey. 1990. Differentiation of Serratia marcescens 4
274 into swimmer and swarmer cells. J.Bacteriol. 172:4322-4328. 5
3. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, 6
and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of
7
protein database search programs. Nucleic Acids Res. 25:3389-3402. 8
4. Ang, S., Y. T. Horng, J. C. Shu, P. C. Soo, J. H. Liu, W. C. Yi, H. C. Lai, K. T. 9
Luh, S. W. Ho, and S. Swift. 2001. The role of RsmA in the regulation of
10
swarming motility in Serratia marcescens. J.Biomed.Sci. 8:160-169. 11
5. Ang, S., Y. T. Horng, J. C. Shu, P. C. Soo, J. H. Liu, W. C. Yi, H. C. Lai, K. T. 12
Luh, S. W. Ho, and S. Swift. 2001. The role of RsmA in the regulation of
13
swarming motility in Serratia marcescens. J.Biomed.Sci. 8:160-169. 14
6. Appleman, J. A. and V. Stewart. 2003. Mutational analysis of a conserved 15
signal-transducing element: the HAMP linker of the Escherichia coli nitrate 16
sensor NarX. J.Bacteriol. 185:89-97. 17
7. Belas, R., R. Schneider, and M. Melch. 1998. Characterization of Proteus 1
mirabilis precocious swarming mutants: identification of rsbA, encoding a 2
regulator of swarming behavior. J.Bacteriol. 180:6126-6139. 3
8. Blattner, F. R., G. Plunkett, III, C. A. Bloch, N. T. Perna, V. Burland, M. 4
Riley, J. Collado-Vides, J. D. Glasner, C. K. Rode, G. F. Mayhew, J. Gregor,
5
N. W. Davis, H. A. Kirkpatrick, M. A. Goeden, D. J. Rose, B. Mau, and Y.
6
Shao. 1997. The complete genome sequence of Escherichia coli K-12. Science
7
277:1453-1474.
8
9. Chang, A. C. and S. N. Cohen. 1978. Construction and characterization of 9
amplifiable multicopy DNA cloning vehicles derived from the P15A cryptic 10
miniplasmid. J.Bacteriol. 134:1141-1156. 11
10. de Lorenzo, V. and K. N. Timmis. 1994. Analysis and construction of stable 12
phenotypes in gram-negative bacteria with Tn5- and Tn10-derived 13
minitransposons. Methods Enzymol. 235:386-405. 14
11. Eberl, L., S. Molin, and M. Givskov. 1999. Surface motility of serratia 15
liquefaciens MG1. J.Bacteriol. 181:1703-1712. 16
12. Eguchi, Y., T. Oshima, H. Mori, R. Aono, K. Yamamoto, A. Ishihama, and R. 17
two-component system in Escherichia coli. Microbiology 149:2819-2828. 1
13. Fraser, G. M. and C. Hughes. 1999. Swarming motility. Curr.Opin.Microbiol 2
2:630-635.
3
14. Givskov, M., J. Ostling, L. Eberl, P. W. Lindum, A. B. Christensen, G. 4
Christiansen, S. Molin, and S. Kjelleberg. 1998. Two separate regulatory
5
systems participate in control of swarming motility of Serratia liquefaciens MG1. 6
J.Bacteriol. 180:742-745. 7
15. Givskov, M., J. Ostling, L. Eberl, P. W. Lindum, A. B. Christensen, G. 8
Christiansen, S. Molin, and S. Kjelleberg. 1998. Two separate regulatory
9
systems participate in control of swarming motility of Serratia liquefaciens MG1. 10
J.Bacteriol. 180:742-745. 11
16. Gygi, D., M. M. Rahman, H. C. Lai, R. Carlson, J. Guard-Petter, and C. 12
Hughes. 1995. A cell-surface polysaccharide that facilitates rapid population
13
migration by differentiated swarm cells of Proteus mirabilis. Mol Microbiol 14
17:1167-1175.
15
17. Gygi, D., M. M. Rahman, H. C. Lai, R. Carlson, J. Guard-Petter, and C. 16
Hughes. 1995. A cell-surface polysaccharide that facilitates rapid population
17:1167-1175.
1
18. Gygi, D., M. M. Rahman, H. C. Lai, R. Carlson, J. Guard-Petter, and C. 2
Hughes. 1995. A cell-surface polysaccharide that facilitates rapid population
3
migration by differentiated swarm cells of Proteus mirabilis. Mol Microbiol 4
17:1167-1175.
5
19. Hansma, H. G., K. A. Browne, M. Bezanilla, and T. C. Bruice. 1994. Bending 6
and straightening of DNA induced by the same ligand: characterization with the 7
atomic force microscope. Biochemistry 33:8436-8441. 8
20. Hansma, H. G., K. A. Browne, M. Bezanilla, and T. C. Bruice. 1994. Bending 9
and straightening of DNA induced by the same ligand: characterization with the 10
atomic force microscope. Biochemistry 33:8436-8441. 11
21. Harshey, R. M. 1994. Bees aren't the only ones: swarming in gram-negative 12
bacteria. Mol Microbiol 13:389-394. 13
22. Harshey, R. M. 2003. Bacterial motility on a surface: many ways to a common 14
goal. Annu.Rev.Microbiol 57:249-273. 15
23. Hoch, J. A. and T. J. Silhavy. 1995. Two-component Signal Transduction, In 16
American Society for Microbiology Press, Washington, D. C. 17
24. Horng, Y. T., S. C. Deng, M. Daykin, P. C. Soo, J. R. Wei, K. T. Luh, S. W. 1
Ho, S. Swift, H. C. Lai, and P. Williams. 2002. The LuxR family protein SpnR
2
functions as a negative regulator of N-acylhomoserine lactone-dependent 3
quorum sensing in Serratia marcescens. Mol.Microbiol. 45:1655-1671. 4
25. Hyytiainen, H., S. Sjoblom, T. Palomaki, A. Tuikkala, and P. E. Tapio. 2003. 5
The PmrA-PmrB two-component system responding to acidic pH and iron 6
controls virulence in the plant pathogen Erwinia carotovora ssp. carotovora. Mol 7
Microbiol 50:795-807. 8
26. Koronakis, V., M. Cross, B. Senior, E. Koronakis, and C. Hughes. 1987. The 9
secreted hemolysins of Proteus mirabilis, Proteus vulgaris, and Morganella 10
morganii are genetically related to each other and to the alpha-hemolysin of 11
Escherichia coli. J.Bacteriol. 169:1509-1515. 12
27. Koronakis, V., M. Cross, B. Senior, E. Koronakis, and C. Hughes. 1987. The 13
secreted hemolysins of Proteus mirabilis, Proteus vulgaris, and Morganella 14
morganii are genetically related to each other and to the alpha-hemolysin of 15
Escherichia coli. J.Bacteriol. 169:1509-1515. 16
28. Kurz, C. L., S. Chauvet, E. Andres, M. Aurouze, I. Vallet, G. P. Michel, M. 17
Uh, J. Celli, A. Filloux, S. De Bentzmann, I. Steinmetz, J. A. Hoffmann, B. B.
Finlay, J. P. Gorvel, D. Ferrandon, and J. J. Ewbank. 2003. Virulence factors
1
of the human opportunistic pathogen Serratia marcescens identified by in vivo 2
screening. EMBO J. 22:1451-1460. 3
29. Lai, H. C., M. J. Lai, S. Lin-Chao, K. T. Lu, and S. W. Ho. 1997. Population 4
cell differentiation of Serratia marcescens on agar surface and in broth culture. 5
J.Microbiol Immunol.Infect. 30:242-254. 6
30. Liaw, S. J., H. C. Lai, S. W. Ho, K. T. Luh, and W. B. Wang. 2001. 7
Characterisation of p-nitrophenylglycerol-resistant Proteus mirabilis 8
super-swarming mutants. J.Med.Microbiol 50:1039-1048. 9
31. Liaw, S. J., H. C. Lai, S. W. Ho, K. T. Luh, and W. B. Wang. 2001. 10
Characterisation of p-nitrophenylglycerol-resistant Proteus mirabilis 11
super-swarming mutants. J.Med.Microbiol 50:1039-1048. 12
32. Lindum, P. W., U. Anthoni, C. Christophersen, L. Eberl, S. Molin, and M. 13
Givskov. 1998. N-Acyl-L-homoserine lactone autoinducers control production
14
of an extracellular lipopeptide biosurfactant required for swarming motility of 15
Serratia liquefaciens MG1. J.Bacteriol. 180:6384-6388. 16
33. Lindum, P. W., U. Anthoni, C. Christophersen, L. Eberl, S. Molin, and M. 17
of an extracellular lipopeptide biosurfactant required for swarming motility of 1
Serratia liquefaciens MG1. J.Bacteriol. 180:6384-6388. 2
34. Lindum, P. W., U. Anthoni, C. Christophersen, L. Eberl, S. Molin, and M. 3
Givskov. 1998. N-Acyl-L-homoserine lactone autoinducers control production
4
of an extracellular lipopeptide biosurfactant required for swarming motility of 5
Serratia liquefaciens MG1. J.Bacteriol. 180:6384-6388. 6
35. Liu, J. H., M. J. Lai, S. Ang, J. C. Shu, P. C. Soo, Y. T. Horng, W. C. Yi, H. C. 7
Lai, K. T. Luh, S. W. Ho, and S. Swift. 2000. Role of flhDC in the expression
8
of the nuclease gene nucA, cell division and flagellar synthesis in Serratia 9
marcescens. J.Biomed.Sci. 7:475-483. 10
36. Matsuyama, T., K. Kaneda, Y. Nakagawa, K. Isa, H. Hara-Hotta, and I. 11
Yano. 1992. A novel extracellular cyclic lipopeptide which promotes
12
flagellum-dependent and -independent spreading growth of Serratia marcescens. 13
J.Bacteriol. 174:1769-1776. 14
37. Matsuyama, T., K. Kaneda, Y. Nakagawa, K. Isa, H. Hara-Hotta, and I. 15
Yano. 1992. A novel extracellular cyclic lipopeptide which promotes
16
flagellum-dependent and -independent spreading growth of Serratia marcescens. 17
J.Bacteriol. 174:1769-1776. 18
38. Pessi, G., F. Williams, Z. Hindle, K. Heurlier, M. T. Holden, M. Camara, D. 1
Haas, and P. Williams. 2001. The global posttranscriptional regulator RsmA
2
modulates production of virulence determinants and N-acylhomoserine lactones 3
in Pseudomonas aeruginosa. J.Bacteriol. 183:6676-6683. 4
39. Prentki, P. and H. M. Krisch. 1984. In vitro insertional mutagenesis with a 5
selectable DNA fragment. Gene 29:303-313. 6
40. Sambrook, J., T. Maniatis, and E. F. Fritsch. 1989. Molecular cloning. A 7
laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. 8
41. Stock, A. M., V. L. Robinson, and P. N. Goudreau. 2000. 9
TWO-COMPONENT SIGNAL TRANSDUCTION. Annual Review of 10
Biochemistry 69:183-215. 11
42. Stock, J. B., A. J. Ninfa, and A. M. Stock. 1989. Protein phosphorylation and 12
regulation of adaptive responses in bacteria. Microbiol Rev. 53:450-490. 13
43. Takeda, S., Y. Fujisawa, M. Matsubara, H. Aiba, and T. Mizuno. 2001. A 14
novel feature of the multistep phosphorelay in Escherichia coli: a revised model 15
of the RcsC --> YojN --> RcsB signalling pathway implicated in capsular 16
synthesis and swarming behaviour. Mol Microbiol 40:440-450. 17
44. Takeda, S., Y. Fujisawa, M. Matsubara, H. Aiba, and T. Mizuno. 2001. A 1
novel feature of the multistep phosphorelay in Escherichia coli: a revised model 2
of the RcsC --> YojN --> RcsB signalling pathway implicated in capsular 3
synthesis and swarming behaviour. Mol Microbiol 40:440-450. 4
45. Toguchi, A., M. Siano, M. Burkart, and R. M. Harshey. 2000. Genetics of 5
swarming motility in Salmonella enterica serovar typhimurium: critical role for 6
lipopolysaccharide. J.Bacteriol. 182:6308-6321. 7
46. Vigh, L., B. Maresca, and J. L. Harwood. 1998. Does the membrane's 8
physical state control the expression of heat shock and other genes? Trends 9
Biochem.Sci. 23:369-374. 10
47. Winson, M. K., S. Swift, L. Fish, J. P. Throup, F. Jorgensen, S. R. Chhabra, 11
B. W. Bycroft, P. Williams, and G. S. Stewart. 1998. Construction and analysis
12
of luxCDABE-based plasmid sensors for investigating N-acyl homoserine 13
lactone-mediated quorum sensing. FEMS Microbiol Lett. 163:185-192. 14
48. Wolanin, P. M., P. A. Thomason, and J. B. Stock. 2002. Histidine protein 15
kinases: key signal transducers outside the animal kingdom. Genome Biol. 16
3:REVIEWS3013.
1
Table 1. Percentage of cellular fatty acids in both swarming and non-swarming S. 2
marcescens cells. Strains including S. marcescens CH-1, CH-1∆A, CH-1∆B were
3
inoculated under different culture conditions, leading to either swarming or 4
non-swarming phenotypes. The composition of cellular fatty acids was analyzed by 5
gas chromatography (MIDI, USA). Results are the means of three independent 6
determinations (SEM<10%). “Ratio*” means the ratio of the calculated fatty acid 7
composition (12:0+14:0+16:0+18:0/10:03OH+15:0 anteiso+18:1 w7c). 8
9
Percentage of total fatty acids
10:0 (3OH) 12:00 14:00 15:0 (anteiso) 16:00 18:1 (w7c) 18:00 Ratio*
swarming cells 2.7 (ave)
30CH-1/LB 4.26 7.52 2.95 25.96 17.08 2.92 2.0 30CH-1/MGM 2.9 8.08 6.72 30.42 10.22 0.83 2.5 30CH-1/MGM/Cas 3.44 6.08 6.41 28.66 8.82 2.5 30CH-1/MGM/Cas/YE 3.55 7.42 3.69 29.54 17.21 1.9 30CH-1/MGM/Cas/ C14:1 ∆9 2.42 9.2 2.81 30.82 10.89 1.99 3.2 30CH-1/MGM/Cas/ C16:1 ∆9 3.62 8.63 3.06 31.48 14.84 2.4
30CH-1/MGM/Cas/ C18:1 ∆9 3.13 8.18 2.47 29.52 13.64 2.17 2.7 30CH-1∆A/LB 2.71 6.21 1.9 29.21 15.85 2.1 30CH-1∆B/LB 3.41 6.2 2.47 27.37 17.63 1.8 37CH-1/MGM 2.02 9.09 5.57 38.99 6.83 4.0 37CH-1/MGM/Cas 3.21 1.9 7.17 1.38 33.53 7.58 1.19 3.6 37CH-1∆A/LB 4.28 7.04 7.48 25.92 13.58 1.8 37CH-1∆A/MGM 6.47 11.26 33.95 5.6 2.4 37CH-1∆A/MGM/Cas 5.99 1.64 32.47 7.7 1.64 4.3 37CH-1∆A/MGM/Cas/C14 1.12 13.14 1.49 35.42 14.96 0.85 3.1 37CH-1∆B/LB 2.91 7.49 3.75 33.98 13.57 2.6 37CH-1∆B/MGM 8.14 7.67 9.17 33.64 7.42 3.0 37CH-1∆B/MGM/Cas 3.18 2.32 7.65 1.34 35.01 8.11 1.02 3.6
non-swarming cells 6.2 (ave)
30CH-1/MGM/Cas/ C12 1.76 24.75 5.97 1.16 30.01 5.66 0.9 7.2 30CH-1/MGM/Cas/ C14 1.63 3.65 12.78 0.6 36.25 8.15 0.73 5.1 30CH-1/MGM/Cas/ C16 1.63 11.12 7.35 0.89 35.63 6.42 0.89 6.2 37CH-1/ LB 7.75 8.25 1.35 29.8 7.27 2.63 5.6 37CH-1/ MGM/ Cas/ YE 1.08 5.48 10.28 0.6 36.33 5.82 1.12 7.1 1
* Calculated as the total percentage of 12:00, 14:00, 16:00 and 18:00 divided by total 1
percentage of 10:0 3OH, 15:0 anteiso and 18:1 w7c. MGM, M9 minimal growth 2
medium; YE, yeast extract; Cas, cas amino acids. 3