國
立
交
通
大
學
電子物理研究所
博
士
論
文
應用奈米技術探討克雷白氏肺炎桿菌第三型線毛
的黏附與延展特性
Using Nanotechniques to Explore the Adhesion and Extension Properties of
Klebsiella pneumoniae Type 3 Fimbriae
研 究 生:詹佳翰
指導教授:徐 琅 教授
應用奈米技術探討克雷白氏肺炎桿菌第三型線毛的黏附與延展特性
Using nanotechniques to explore the adhesion and extension properties of
Klebsiella pneumoniae type 3 fimbriae
研 究 生:詹佳翰
Student:Chia-Han Chan
指導教授:徐 琅
Advisor:Long Hsu
國 立 交 通 大 學
電 子 物 理 研 究 所
博 士 論 文
A ThesisSubmitted to Department of Electrophysics College of Science
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of Philosophy
in
Department of Electrophysics October 2011
Hsinchu, Taiwan, Republic of China
i
應用奈米技術探討克雷白氏肺炎桿菌第三型線毛的黏附與延展特性
學生:詹佳翰
指導教授
:徐 琅
國立交通大學電子物理研究所 博士班
摘
要
細菌藉由其表面線毛黏附宿主細胞,因線毛具螺旋彈簧結構的伸縮特性,能降低環境沖 刷力的影響,使細菌與宿主保持緊密黏附。克雷白氏肺炎桿菌(Klebsiella pneumoniae)是一株伺 機性感染細菌,近年來臨床上發現多種抗藥性菌株,使得治療方式趨於困難,而其表面第三 型線毛引起細胞製造生物膜,進而產生抗藥性。本論文將以克雷白氏肺炎桿菌主要致病的因 子第三型線毛作為材料,利用雷射鑷夾(OT)、電子顯微鏡(TEM)、原子力顯微鏡(AFM)、 基因重組技術、基因標定技術、胺基酸定點突變技術等奈米技術,從生物物理的角度,探討 單一根第三型線毛的特性,並進一步分析影響線毛主要蛋白MrkA 組裝的重要殘基。我們發現 第三型線毛的長度約為1 微米,線毛的螺旋截面並不對稱,截面最大寬度 7.4 奈米和最小寬度 4.2 奈米。第三型線毛頂端黏附蛋白有四個黏附因子,其中 MrkDv3 與膠原蛋白的黏附力最大, 約為 4 皮牛頓,是黏附蛋白的重要因子。第三型線毛受力伸展曲線具有三個階段:(1)第一階 段表現線性的受力伸長特性,彈性係數為60.9 皮牛頓/微米,楊氏模數為 100 皮牛頓/微米2; (2)第二階段表現固定力伸展特性,野生株的線毛以固定力 66 ± 4 皮牛頓拆開並伸展數微米長; (3)第三階段表現非線性的受力伸長特性,其受力伸展曲線轉折點為特徵力 102 ± 9 皮牛頓。另 外,針對第三型線毛骨幹主要蛋白MrkA 單體,我們篩選十二個 MrkA 關鍵的胺基酸進行定點 突變,以分析其受力伸展物理特性的變化。其中G189A 突變株具特殊力學性能,其第二階段 的拆解力呈現持續增加的拆解力(57~68 皮牛頓),不同於其他突變株或一般株以固定 66 皮牛頓 力拆解。這樣的結果顯示,G189 胺基酸對 MrkA 的結構拆解力穩定度扮演重要的角色。本論 文提供一個研究各種線毛的生物力學性能的奈米技術平台,藉由線毛生物物理特性結果與發 現,能提供細菌感染致病的資訊,有助於傳統細菌感染與抗藥性的研究。
ii
Using nanotechniques to explore the adhesion and extension properties of
Klebsiella pneumoniae type 3 fimbriae
Student:Chia-Han Chan
Advisors:Dr. Long Hsu
Department of Electrophysics
National Chiao Tung University
ABSTRACT
Fimbriae exhibiting flexible and stretchable properties similar to springs may aid bacteria in
remaining attached to host cells by reducing the impact of the flushes in environment. Klebsiella
pneumoniae is a common pathogen causing both pneumonia and urinary tract infections. Most
pathogenic Klebsiella pneumoniae strains produce type 3 fimbriae, which are critical for bacterial biofilm formation. In this thesis, we investigated the structural and mechanical properties of
Klebsiella pneumoniae type 3 fimbriae, which constitute a known virulence factor for the bacterium,
and to identify residues in the major pilin MrkA important for pilus assembly. Type 3 fimbria was 1 micron in length, and the largest diameter was 7.4 nm and the smallest was 4.2 nm, have an elliptically cylindrical structure with the largest diameter and smallest diameter of each ellipse-like cross section in fimbrial structures by using transmission electron microscopy. We present type 3 fimbriae MrkD adhesin variants, mrkDV3-,was the best adhesin, and the adhesive force between mrkDV3-expressed fimbriae and collagen IV is about 4 pN.The force-extension curve of type 3 fimbria exhibited a three-phase:(1) the first spring-like phase stretched with increasing force, the spring constant kp of 60.9 N/μm, the estimated Young’s moduli (E) were 100 Mpa, (2) the second uncoiling phase, it started to uncoil and extended several micrometers at a fixed force of 66 ± 4 pN, and (3) the final nonlinear phase with a characteristic force of 102 ± 9.8 pN at the inflection point. In addition, we screened twelve critical residues on major pilins MrkA of type 3 fimbriae. By contrast, the G189A mutant remained capable of producing normal amounts of fimbriae. Further investigating the mechanical properties of the G189A fimbriae revealed that the uncoiling force for MrkA-MrkA interaction varied from 58 pN to 67 pN rather than be constant like the uncoiling force of 66 pN measured from the other mutants or wild type. The work has provided a method that allows us to understand of the biomechanical function of different types of pili. The investigation of pili brings information for the design of new drugs to prevent bacterial infections, which is increased bacterial resistance towards antibiotics.
iii
誌
謝
生命是一個無休止向前運轉的巨輪,我們所衝過的每一道關卡,將是下一個目標的起點。 感謝恩師 徐琅老師的栽培與包容,還有感謝師母一路關心與鼓勵,尤其待人處事方面,您們 的諄諄教誨都是我全方位的明燈,我會銘記在心,在往後的人生裡。感謝國家型奈米計畫的 老師們,尤其是生科系彭慧玲老師、清大生科系張晃猷老師、清大動機系劉承賢老師、清大 材料系游萃蓉老師,謝謝您們在我的研究上耐心的指導。感謝大師兄博睿學長為跨領域研究 開啟先河、生科系盈蓉學姊鼎力相助、最執著於生物物理研究的夥伴豐榮一路相挺,讓我勇 敢的渡過許多低潮。還有龍蝦團隊的夥伴們國良、欣瑜、建誠,謝謝你們幫忙催生這篇論文。 實驗室的夥伴,不打不相識的勝陽,終於要一起畢業,博真學姊、愛堂、哲良、東銘、士模、 頌揚、軒毫、文鴻、庭樵,希望大家繼續在跨領域的研究上發光發熱。最感謝的是我的愛妻 慧君,謝謝妳為我打理一切,使我在工作和學業上無後顧之憂,亦要在此對愛女欣叡及愛子 欣諺說聲抱歉這陣子對你們疏忽了,你們可愛的臉龐,就是我撐下去的動力。感謝我的家人 無怨無悔的付出以及支持,是我得以能完成學業,並且繼續朝我的理想邁進的重要支柱,最 後,再次謝謝一路幫助我的朋友們,在這人生短暫交會之際,謝謝你們綻放溫暖的光輝,照 亮我前行的道路,幫助我完成人生夢想,感恩。 詹佳翰謹至于 國立交通大學電子物理研究所 中華民國一百年十月 本論文獻給我最敬愛的父母、岳父母、家人以及朋友們,願你們永遠健康快樂
iv
目
錄
中文摘要 ……… i 英文摘要 ……… ii 致謝 ……… iii 目錄 ……… iv 表目錄 ……… v 圖目錄 ……… vi 一 緒論……… 1 二 原理……… 4 2-1 雷射鑷夾捕捉力的特性……… 4 2-2 雷射鑷夾捕捉力的光彈性係數校正……… 5 2-3 功率頻譜密度法……… 5 2-4 四象限光偵測器奈米級解析度的測量技術……… 8 三、 雷射鑷夾的架設……… 10 3-1 雷射鑷夾系統……… 10 3-2 微粒子位置檢測系統……… 12 3-3 樣品平台……… 13 3-4 系統控制程式……… 13 四、 實驗方法……… 15 4-1 第三型線毛與第四型膠原蛋白間黏附力……… 15 4-2 第三型線毛在應力下的機械特性……… 17 4-3 第三型線毛主要蛋白氨基酸序列突變的影響……… 23 五、 結 果 與 討 論……… 27 5-1 第三型線毛與第四型膠原蛋白間黏附力……… 27 5-2 第三型線毛在應力下的機械特性……… 32 5-3 第三型線毛主要蛋白氨基酸序列突變的影響……… 40 六、 結論與展望……… 49 參考文獻 ……… 51 附錄一 12 株點突變體的電子顯微鏡照片……… 55 附錄二 第三型線毛的串鏈模型……… 61
v
表 目 錄
表4-1 點突變所使用的引子………..……… 26 表5-1 常見材料的楊氏模數………..……… 34 表5-2 第三型線毛與其他型線毛的物理參數之比較……...……… 40 表5-3 點突變體重組線毛拆解力………...……… 46vi
圖 目 錄
圖1-1 第三型線毛的構成要素與其三維螺旋形結構示意圖…………...……… 3 圖2-1 微粒子偏移捕捉中心距離與四象限光偵測器訊號關係圖………...…… 4 圖2-2 利用功率頻譜密度測量雷射鑷夾光彈性示意圖………...…… 7 圖2-3 利用功率頻譜四象限光偵測器示意圖………...…… 9 圖3-1 雷射鑷夾系統的照片………...…… 11 圖3-2 雷射鑷夾系統示意圖………...…… 11 圖3-3 四象限光偵測器裝置圖………...…… 12 圖3-4 樣品平台裝置圖………...…… 13 圖3-5 「線毛受外力的伸展曲線」之操作面板……….…… 14 圖3-6 「線毛受外力的伸展曲線」之數據分析比較面板……….…… 14 圖4-1 雷射鑷夾測量線毛與膠原蛋白間黏附力實驗示意圖………...…… 16 圖4-2 雷射鑷夾測量線毛與膠原蛋白間黏附力的訊號曲線………...…… 17 圖4-3 利用雷射鑷夾操控黏附線毛的微粒子示意圖………...…… 19 圖4-4 單一方向線毛彈力實驗的訊號曲線………...…… 19 圖4-5 線毛長度量測實驗示意圖………...…… 20 圖4-6 線毛彈性以及楊氏模數實驗數據及示意圖………...…… 21 圖4-7 線毛受力而伸長的示意圖………...…… 22 圖4-8 第三型線毛骨幹蛋白胺基酸序列示意圖…………...……… 23 圖4-9 各種線毛的線毛單元體的胺基酸序列比較圖………...…… 25 圖5-1 四種大腸桿菌的線毛生長密度電子顯微鏡照片………...…… 27 圖5-2 線毛黏附力的數據統計圖………...…… 28 圖5-3 黏附力的群組週期分佈圖………...…… 30 圖5-4 線毛黏附數量與黏附力圖………...…… 31 圖5-5 第三型線毛原子力顯微鏡照片………...…… 32 圖5-6 第三型線毛電子顯微鏡照片………...…… 34 圖5-7 單一根第三型線毛之典型受力伸長關係圖……… 35 圖5-8 典型的第三型線毛受外力的伸長示意圖………...…… 36 圖5-9 典型的第三型線毛受外力的伸長曲線………...…… 37 圖5-10 兩根第三型線毛受外力的伸長曲線………...…… 38 圖5-11 兩根第三型線毛受外力的伸長示意圖………...……… 38 圖5-12 線毛的拆解力與特徵力頻率直方圖………...…… 39 圖5-13 主要蛋白MrkA 不同點突變體的生物膜形成能力……… 41 圖5-14 主要蛋白MrkA 不同點突變體的聚合體電泳圖……… 42 圖5-15 各種突變線毛電子顯微鏡圖………...……… 43 圖5-16 野生菌株與點突變體的線毛數量、平均長度、平均寬度等物理參數… 44 圖5-17 重組突變體單一線毛受力伸展曲線………...……… 46 圖5-18 重組突變體單一線毛受力伸展曲線比較………...……… 47 圖5-19 野生株與G189A 突變體線毛受力伸展(黑色)和收縮(灰色)曲線……… 481
第一章 緒論
許多細菌的感染與侵入都是從附著人類、動物或植物宿主的細胞表面或黏膜開始發 生,大部分的細菌利用它表面絲狀物線毛(或稱為纖毛)去貼附宿主細胞的表面,以感染 宿主細胞進而對其造成傷害[1-3]。細菌黏附宿主細胞,一般經由位於線毛頂端或主體的 黏附蛋白(adhesin)附著於宿主細胞,且藉由線毛骨幹類似於彈簧的拉伸性能,能幫助細 菌緊密黏附,以抵抗環境的衝擊力,例如在尿道或在呼吸道所產生拖曳力。由於不同種 類的線毛的形狀與機械特性有所不同,因而造成感染的疾病也各異。其中第一型線毛與 P 型線毛是大腸桿菌造成感染的主要媒介,其外觀較為粗直,最常被國外研究;而第三 型線毛則較為纖細彎曲,常見於引發的肺炎疾病的克雷白氏肺炎桿菌,過去幾乎沒有關 於第三型線毛機械特性方面的研究。本論文利用雷射鑷夾(OT)、電子顯微鏡(TEM)、 原子力顯微鏡(AFM)、基因重組技術、基因標定技術、胺基酸定點突變技術等奈米技 術,從生物物理的角度,探討單一根第三型線毛的特性。 過去,透過電子顯微鏡的觀察,線毛的外觀已經幾乎被確認。雖然這些靜態圖象提 供外觀結構的訊息,但是關於細菌線毛在活體的動態的功能的訊息卻無法得知。直到最 近,測量材料彈性之奈米技術的發展[4],第一型與 P 型線毛的結構和生物力學的角度分 析,如受力伸長特性,已被雷射鑷夾[5, 6]與原子力顯微鏡[7, 8]所定量測出,並用以解 釋線毛的可伸縮性將影響菌體在黏附宿主細胞時所能承受的剪力(例如尿液沖刷)。線毛 的典型尺寸是直徑約為2-10 奈米、長度約為 1-2 微米,黏附分子位於線毛頂端以黏附 宿主細胞,而線毛骨幹則由主要蛋白組裝成三維螺旋結構[9-11]。螺旋結構可以使線毛 伸展和收縮,而大多數機械特性取決於這螺旋結構的延展特性,用以抵抗外界環境流動 的剪切力。根據線毛受力伸展曲線分析顯示,線毛骨幹透過一串主要蛋白單元體蜿蜒連 接形成,類似一條螺旋電話線。那些主要蛋白單元體透過層與層之間連續的鍵結結合成 穩定架構,當線毛受外力時,線毛骨幹主要蛋白單元體一一被拉開,使線毛將以數倍長 度被拉長[12]。特別的是,而當外力停止時,線毛骨幹將恢復原狀,顯然像螺旋形一樣 的架構的展開完全可逆[13]。有報導指出,線毛展開的可逆現象,將受到環境酸鹼值影 響,而使線毛主要蛋白分子間交互作用力有明顯變化[14]。此外,線毛主要蛋白以及頂 端黏附蛋白對於細菌形成生物膜也扮演重要角色[15, 16]。因此,線毛受力伸展的量測, 對於線毛的黏附與延展特性的研究是重要的。2
多數的細菌皆擁有多種不同形式的線毛,譬如大腸桿菌有第一型線毛、P 型線毛等 [17],沙門氏菌(Salmonella)有第一型線毛與第三型線毛[18],奈瑟氏腦膜炎雙球菌 (Neisseria meningitidis)與奈瑟氏淋病雙球菌(Neisseria gonorrhoeae)則有第四型線毛 [19]。克雷白氏肺炎桿菌(Klebsiella pneumoniae)在臨床上是一株伺機性感染細菌,平常 少量存在於健康人體腸道中,在人體免疫不全時會造成感染,而引起肺炎、呼吸道感染、 肝膿瘍、尿道感染、敗血症及腦膜炎等症[20]。最近幾年來,臨床上發現克雷白氏菌具 有多種抗藥性的分離菌株[21],使得治療方式趨於困難。克雷白氏肺炎桿菌表面有第一 型和第三型線毛,其中第三型線毛被認為可能是參與致病的主要因子[22, 23],扮演著體 外與數種不同形式的細胞和細胞外基質蛋白質鍵結的角色,因此,認是第三型線毛結構 是重要的。 第三型線毛長約 1 - 2 μm,寬約 4 - 7 nm,構成第三型纖毛所需的五種蛋白質已被確 定。這些蛋白質分別由基因組mrkA、mrkB、mrkC、mrkD 與 mrkF 所轉譯[24, 25]。第三 型線毛的構成要素之示意圖顯示在圖1。mrkA 轉譯的蛋白 MrkA 組裝成第三型線毛骨幹 的主要單元體;mrkB and mrkC 所轉譯的 MrkB 和 MrkC 則與在細菌外膜的調控線毛組 裝有關,MrkB 則是位於周膜間隙的 chaperone,負責穩定線毛單元體並幫助其正常折疊, MrkC 為位於細菌外膜上的 usher,線毛單元體經此蛋白被運送至膜外,並固著在其上; mrkD 基因可以轉譯出位於線毛頂端的黏附蛋白 MrkD,是克雷白氏肺炎桿菌中與紅血球 凝集有關之黏附分子;最後,MrkF 經由實驗證實,與維持第三型線毛在細菌外膜上的 穩定性有關[26],如圖 1-1 所示。然而,不同於被詳加研究的第一型與 P 型線毛,第三 型線毛的生物物理特性至今仍大部分未知。第三型線毛組成蛋白的胺基酸序列與P 型線 毛組成蛋白的胺基酸序列差異大,且這類型線毛的結構組織與機械特性也仍尚未了解。 另外,由於蛋白質MrkA 是第三型線毛的主要蛋白分子,我們將主要蛋白 MrkA 進行特 定胺基酸之點突變,分析點突變對於線毛堆疊以及線毛正常功能的影響,進而推斷這些 胺基酸在線毛組裝的過程中所扮演的角色。研究線毛的組裝機制,除了可以更進一步了 解此制病因子的結構與功能、探討其致病原理外,同時也可作為分子生物學中,研究ㄧ 般單元體組裝成聚合體之調控機制的參考模型[27, 28]。
3 圖1-1 第三型線毛的構成要素與其三維螺旋形結構示意圖 本論文中,我們以克雷白氏主要致病因子第三型線毛為樣品,利用電子顯微鏡以及 原子力顯微鏡觀測線毛微結構包括直徑、長度與密度。接著,利用雷射鑷夾操控微粒子 去黏附單一線毛,並且將之拉長至最大可允許移動量,藉由線毛受力伸長的數據,分析 得到線毛的黏附以及延展特性。然後,比較第三型線毛與其他類型的線毛,就其結構和 力學性能之間的差異,藉此模擬細菌黏附寄主細胞後受到環境剪力,及其與宿主之間的 交互作用,期望未來得知克雷白氏肺炎桿菌入侵人體的方式。在第二章,我們介紹雷射 鑷夾的捕捉機制,並利用光功率頻譜密度(PSD)檢測器,據此量化雷射鑷夾橫向捕捉力, 與微生物位移量的關係。第三章介紹雷射鑷夾系統的架設。第四章測量第三型線毛線毛 頂端黏附分子的黏附力,測量單一根第三型線毛在應力作用下的機械特性,並進一步分 析第三型線毛主要蛋白單元體 Mrk A 胺基酸序列突變對其生物及機械特性的影響。最 後,第五章對這篇論文做一個總結,並敘述未來的展望。
4
第二章 原理
雷射鑷夾的基本概念是將一道雷射光束經顯微物鏡聚焦,在雷射聚焦處形成一穩定 的位能井,使得在此焦點附近的微粒子就會被吸引到焦點,而達到捕捉的目的。當微粒 子偏移聚焦處的均衡位置時,將承受一個隨偏移量成正比的虎克定律回復力。雷射鑷夾 系統提供奈米級位移解析度以及皮牛頓等級力的測量,非常適合於分子生物間交互作用 力的研究。 2-1 雷射鑷夾捕捉力的特性 雷射鑷夾的捕捉力就像線性彈簧的作用力一樣,彈簧彈力大小與彈簧伸長量 成正比,兩者的比值即為彈簧的彈性係數,而雷射鑷夾受力變化正比於捕獲物體 偏離平衡位置之位移量。因此,雷射鑷夾有如一『光學彈簧』,黏附在被捕捉的微粒 子上,具有固定的「捕捉力彈性係數」。在沒有其他外力作用下,被捕捉的微粒子停留 在彈簧的平衡點(即雷射光焦點)。如圖 2-1 所示,當被雷射鑷夾捕捉住的微粒子受到外 力F 時,微粒子圓心與雷射鑷夾捕捉中心的偏移距離(D)也隨之改變,就像對彈簧壓縮, 會引起彈簧形變,產生對應之彈力。根據「微粒子位移量」與「光學捕捉力」的關係, 定量分析雷射鑷夾的「光彈性係數」,進而校正雷射鑷夾捕捉力F(OT)。 圖 2-1 微粒子偏移捕捉中心距離與四象限光偵測器訊號關係圖 微粒子圓心與雷射鑷夾捕捉中心的偏移距離(D); 四象限光偵測器(QPD)訊號 Optical Tweezeers QPD D>0 F F(OT) F F(OT) Optical Tweezeers QPD D=0 PZT XYZ-Stage QPD Optical Tweezeers D > 0 PZT XYZ-Stage PZT XYZ-Stage5
2-2 雷射鑷夾捕捉力的光彈性係數校正
為了應用雷射鑷夾於生物黏附及延展實驗。我們首先要校正雷射鑷夾的光彈性係 數。我們利用功率頻譜密度分析法(Power spectrum density)量測雷射鑷夾光彈性係數,以 校正微粒子的偏移距離與雷射鑷夾捕捉力的關係。接著,測量微粒子的偏移量,再乘上 光彈性係數,即可得到雷射鑷夾施於微粒子的力量。 功率頻譜密度分析法的優點是直接利用雷射鑷夾捕捉一個微粒子,即可量出雷射鑷 夾作用於這顆微粒子的光彈性係數。當我們利用雷射鑷夾捕捉微粒子時,雖然微粒子會 受到捕捉力而限制雷射鑷夾在所建立的光學陷阱之中,但卻因為布朗運動的存在,使微 粒子並非穩定地停止在雷射鑷夾的捕捉中心,而會在捕捉中心的附近晃動。 為了得到奈米尺度的解析能力,我們利用四象限光偵測器(QPD)訊號分析的單一粒 子追蹤技術[39],測量微粒子在雷射鑷夾的捕捉中心晃動的範圍與頻率,即可得知雷射 鑷夾光彈性係數。雷射鑷夾的光彈性係數越大時,微粒子運動的範圍與頻率也越大。 單一粒子追蹤技術,主要是利用四象限光偵測器,偵測被雷射鑷夾捕捉之微粒子的位 置,其位置偏移所導致光強度在光偵測器,所造成的電壓值變化。光偵測器的訊號是以 電壓值(V)為單位,我們進一步以功率頻譜密度分析法校正光偵測器的訊號,分析其電 壓值與微粒子實際的位置(nm)的換算值(nm/V)。而這個換算值,對線毛黏附與延展特性 的實驗非常重要。因為在線毛實驗中,我們必須要知道微粒子的位移,來反推微粒子所 受的力以及線毛被拉開的長度等。總之,功率頻譜密度分析法是一個相當普遍且方便於 雷射鑷夾光彈性係數量測與四象限光偵測器(QPD)校正的方法。 2-3 功率頻譜密度法[29-31] 我們利用朗玆凡方程式(Langevin equation)來描述粒子在雷射鑷夾捕捉位能井中的 布朗運動:
)
(
)
(
)
(
)
(
t
kx
t
x
t
m
x
t
F
−
−
β
′
=
′′
, (2.3.1) 其中F(t)熱擾動形成的力,x(t)是微粒子在雷射鑷夾位能井中,因布朗運動而隨時間 t 的 所在位置,k 則為雷射鑷夾光彈性係數,kx(t)是雷射鑷夾捕捉力,x’(t) 是微粒子的速度, rπη
β
=6 表示水流黏滯力常數,η 為水黏滯係數,r 為微小粒子之半徑,β
x’(t) 則是微 粒子所受的的水流黏滯力的大小, m 是微粒子的質量,mx’’(t) 則是微粒子所受的合力。6 (2.3.1)式表示一個力平衡的方程式,其中粒子加速度的產生的力與液體的黏滯力以及雷 射鑷夾的彈簧恢復力,這與溶液分子隨機撞擊。 由於微粒子在水中的運動其實是一個阻尼很大的運動,所以當微粒子被水分子撞擊 時產生的作用力,很快就會達到力平衡,使微粒子到達其終端速度。因此,(2.3.1)式中 的三個力的合力m ′′ 項與其他幾項比起來是小很多,我們可以把最後一項 xx m ′′ 忽略為零,
)
(
)
(
)
(
t
kx
t
F
t
x
′
+
=
β
, (2.3.2) 我們將(2.3.2)式兩邊做傅立葉轉換後可以得到:)}
(
{
)}
(
{
)}
(
{
x
t
k
FT
x
t
FT
F
t
FT
′
+
⋅
=
⋅
β
, (2.3.3) 其中FT{}表示傅利葉轉換運算。又根據傅利葉轉換運算規則,)}
(
{
)
2
(
)}
(
{
x
t
i
f
FT
x
t
FT
′
=
π
,將此關係式代入(2.3.3)可改寫成:)}
(
{
)}
(
{
)}
(
{
)
2
(
i
f
FT
x
t
+
k
⋅
FT
x
t
=
FT
F
t
⋅
π
β
, (2.3.4) 我們將(2.3.4)式兩邊取絕對值平方後可以得到:[
2 2]
2 2)}
(
{
)}
(
{
)
2
(
f
FT
x
t
FT
F
t
k
+
π
β
=
, (2.3.5) 其中FT
{ t
x
(
)}
2代表的是微粒子位置分布x(t)的能量頻譜。因為 F(t)代表隨機的力量, 對應時間的平均數是零,其能量頻譜[31,32]為 2)}
(
{
)
(
f
FT
F
t
PSD
F=
=4β
kBT, (2.3.6) 其中 kB 是波茲曼常數,T 是絕對溫度。 將(2.3.6)中的4β
kBT 代入(2.3.5)式中的FT
{ t
x
(
)}
2可以得到x(t)的能量頻譜 2 2 2)
2
(
)}
(
{
)
(
f
k
t
F
FT
f
PSD
xπβ
+
=
= ⎥⎦ ⎤ ⎢⎣ ⎡ 2+ 2 2 2 ) 2 ( 4 4 f k T kBπβ
β
π
β
= 2[
2 2]
f f T k c B +β
π
, (2.3.7)7 其中
πβ
2
k
f
c=
為微粒子能量頻譜密度的截止頻率,β
=
6
πη
r
表示微粒子所受的水流 黏滯力常數,η 則是的水流黏滯係數。因此我們可以量測一微小球被雷射鑷夾捕捉的布朗運動,接著分析其Power Spectrum Density,量測 Power Spectrum Density 圖中的截止
頻率fc。由 fc =k/2πβ,帶入水阻力常數β =6πηr,即可推得此微小球所受雷射鑷夾捕 捉之光彈性係數k = 2πβ fc ,如圖 2-2。由上述我們可以得到雷射鑷夾光彈簧係數, 以及被捕捉的微粒子偏移捕捉中心時所對應的力,我們可以用來量測分子生物間微小的 作用力。 圖2-2 利用功率頻譜密度測量雷射鑷夾光彈性示意圖 (A)、(B)分別為 X、Y 方向微粒子在一均勻位能井中的布朗運動隨時間的關係圖; (C)、(D)由微粒子的布朗運動得到功率頻譜密度對頻率的關係圖,可校正雷射鑷夾 光彈性係數 Kx、Ky。
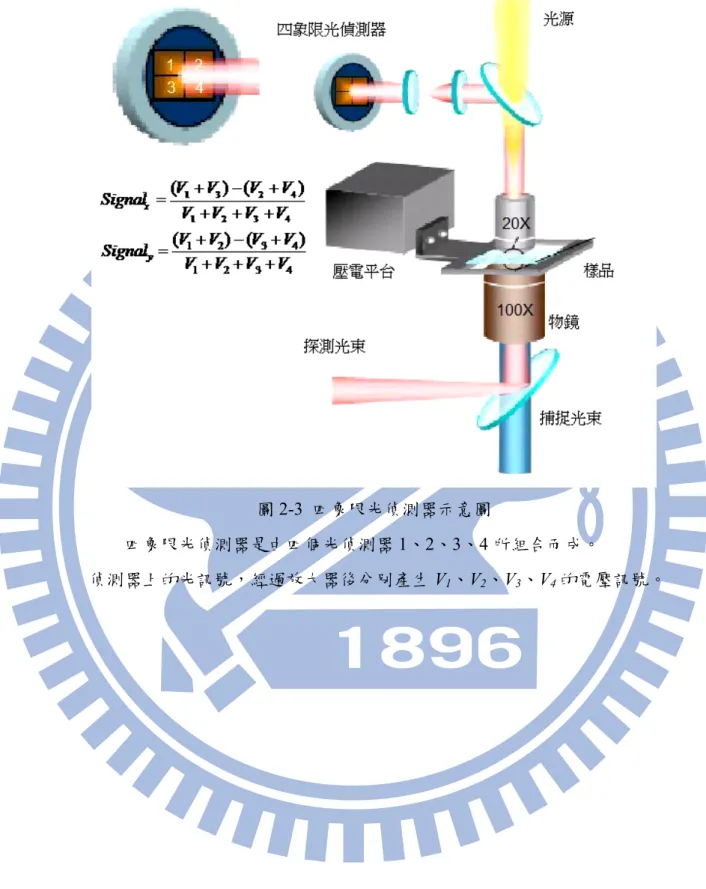
8 2-4 四象限光偵測器奈米級解析度的測量技術[39] 受限於光學的繞射極限,我們無法利用影像處理的方法來觀察被雷射鑷夾捕捉的微 粒子在溶液中微小的布朗運動(Brownian motion)。單一粒子追蹤技術,則為雷射鑷夾開 拓一個嶄新的里程碑,首先,我們利用雷射鑷夾操控一顆被捕捉的微粒子,然後去掃描 探測雷射光束,並用四象限光偵測器偵測穿過微粒子的探測雷射光束的光分佈,藉由偵 測微粒子在溶液中的微小運動,以反推微粒子與捕捉雷射光束焦點之相對位移。而這不 僅增加了雷射鑷夾對微小力量測的解析度,同時藉由空間解析與力解析的提升,雷射鑷 夾成為研究生物物理與機械特性的新工具。 當被捕捉的微粒子在垂直雷射鑷夾光軸方向的位移時,四象限光偵測器上的四個光 偵測器偵測到的光分佈也會產生對應的改變。因此,我們便可以由光強度訊號變化與微 粒子位移的關係,推測出微粒子與雷射鑷夾捕捉中心的相對的位移D,代入雷射鑷夾光 彈性係數,則可以得知雷射鑷夾捕捉力。 如圖 2-3 所示,四象限光偵測器是由四個光偵測器 1、2、3、4 所組合而成。當光 照射在四個偵測器上,經過放大器後分別產生V1、V2、V3、V4的電壓訊號。其中光強度 與電壓訊號成正比。當光斑往水平方向偏移時,四個光偵測器所偵測到的電壓訊號會產 生改變,其中x 方向的位移可以由四個光偵測器的偵測訊號 Sx = (V1+V )−(3 V2+V4);同 理,y 方向則是 Sy = (V1+V2) − (V +3 V4)。而在平行光軸方向,也就是 z 方向的位移,則 由於與四個偵測器所量到的總強度有關,便可以由Sz = (V1+V +3 V2+V4) 求得。 4 3 2 1 4 2 3 1 ) ( ) ( V V V V V V V V Sx + + + + − + = , (2.4.1) 4 3 2 1 4 3 2 1 ) ( ) ( V V V V V V V V Sy + + + + − + = , (2.4.2) 由(2.4.1)及(2.4.2)式得到光斑在水平方向以及垂直方向的光斑偏移量,我們可以進一步 得知微粒子相對雷射光焦點的位移 Dv = (Dx, Dy, Dz) ∝ Sv≡ (Sx, Sy, Sz),以及所對應的雷射 鑷夾捕捉力。
9
圖2-3 四象限光偵測器示意圖
四象限光偵測器是由四個光偵測器 1、2、3、4 所組合而成。
10
第三章 雷射鑷夾的架設
3-1 雷射鑷夾系統
我們的雷射鑷夾系統架設在一台倒立式顯微鏡平台上(DMIRB, Leica, Germany)。如
圖3-1 所示,雷射鑷夾所使用的捕捉光源為一台近紅外波長之固態雷射(10 W, 1064 nm,
IRCL-1064-1000-L, CrystaLaser, USA),其選擇考量在降低光學破壞生物組織的可能性。
雷射光束透過一組12X 擴束器(BE1)擴束後,再由半波長波板(HWP)和偏振分光鏡(PBS)
結合以改變雷射功率。將雷射光束透過顯微鏡邊開口引導到一台倒立式的顯微鏡 (DMIRB, Leica)。雷射光束進入顯微鏡後,經由分光鏡(DM1)將雷射光反射進入顯微物 鏡(OBJ1, 100X, NA=1.25, Leica),此即為一套雷射鑷夾系統。接著我們將微粒子以及細 菌放在兩片蓋玻片之間,再將這組玻片置於一個壓電平台(PZTstage)上作微小且精確的
位移操控,此一壓電平台最小步距1.2 奈米,而最大移動範圍為 80 微米。當雷射鑷夾捕
捉微粒子時,我們利用一個聚光物鏡(OBJ2, 20X, NA=0.5, PlanFluor, Nikon)收集微粒子 造成的雷射散射光,再利用一個聚焦透鏡(L1, f=50mm),將聚光物鏡的後焦平面上的雷 射散射光,成像在一個四象光偵測器(QPD, G6849, Hamamatsu)的位置上。圖 3-1 所示為 雷射鑷夾系統的照片,畫面中雷射鑷夾是架設在一個空氣中漂浮光學平台,以減少振動 的影響。攝影機安裝在顯微鏡的高分辨率成像轉接環,而該樣本影像即時顯示在左側屏 幕,而樣品數據採集以進行研究則顯示在屏幕右側。我們將四象光偵測器量得的電壓經 訊號放大器放大後,由電腦透過資料擷取卡(DAQcard)紀錄,經計算即可獲得微粒子之 位移量,如圖3-2 所示。 為了產生高度聚焦的捕捉光場,我們所使用的物鏡為 100 倍油鏡(NA=1.25, NPLAN, Leica, Germany),並且把我們的生物樣品固定在一個利用 PZT(P-611, PI, Germany)操控 的支架上。而為了建立三維單一粒子追蹤系統並延伸其追蹤範圍[37,39],我們引入一道 探測光束(10 mW, 633 nm);接著利用一片聚光鏡(20 倍, NA=0.5, PlanFluor, Nikon, Japan) 收集探測光經過乳膠珠微擾所產生的前向散射光,最後將散射光打在四象限光偵測器 (Si-PIN, S4349, Hamamatsu, Japan)上。
透過四象限光偵測器所收集到的電壓訊號加以分析,我們可以得到電壓訊號V 對乳i
膠珠偏移捕捉中心點的三維偏移量Δ 之關係(xi Δxi =βi ×Vi,i=x,y,z;βi為各軸的轉 換常數,其單位為[奈米/伏特]);接著我們了解乳膠珠在微小偏移量下所受到雷射鑷夾的
11 捕捉力可視為一組三維線性彈簧力F (i Fi =ki ×Δxi),於是我們將可以得到任何時刻乳膠 珠所受到的三維捕捉光之合力。又因為乳膠珠是連結在第三型線毛上,所以我們利用靜 力平衡的物理觀點將可求得當時線毛所受到的外力大小。 利用近紅外雷射光源通過高度聚焦的顯微物鏡產生捕捉光點,並利用PZT 致動器進行微 小掃描量測線毛受力實驗;接著引入一道氦氖雷射光束作為探測光束,並利用聚光鏡收 光使得前向散射光打在四象限光偵測器上,最後測得乳膠珠對捕捉中心的三維偏移量之 量測。 圖3-1 雷射鑷夾系統的照片 圖3-2 雷射鑷夾系統示意圖[33]
12 3-2 微粒子位置檢測系統 如上所述,引入一道氦氖雷射光束作為探測光束,並利用聚光鏡收光使得前向散射 光打在四象限光偵測器上,如圖3-3 所示。四象限光偵測器將探測光轉換為電流,以截 取微粒子對捕捉中心的三維偏移量之位移訊號,然後將微粒子位移訊號轉換成雷射鑷夾 捕捉力。在這個系統中,四象限光偵測器安裝在顯微鏡上的一個XY 微米定位平台,被 用來作為探測器。當微小球與雷射光焦點重合時,前向散射光斑會均勻的打在四象光偵 測器上。當微小球與雷射光焦點不重合時,如圖中有X 方向的位移時,則光斑不是均勻 的打在四象光偵測器上。我們藉由圖中公式推算前向散射光斑打在四象光偵測器上的位 置,即可反推微小球的奈米級位移。 圖3-3 四象限光偵測器裝置圖 為了量測雷射鑷夾的光彈性係數,功率頻捕密度分析法是一個相當簡單的方法。首 先,雷射鑷夾捕捉一個微粒子,利用四象限光偵測器測量微粒子,在雷射鑷夾捕捉下的 布朗運動,並知道水流黏滯係數,然後將四象限光偵測器偵測到的光訊號轉換為電壓訊 號,則我們就可以利用這個方法量得雷射鑷夾的光彈性係數。接著,利用這個方法校正 光偵測器量得的電壓值(伏特),與微粒子實際的位置(奈米)的換算值(奈米/伏特),我進而 得知微粒子對於雷射鑷夾捕捉中心的偏移量,以及所對應的雷射鑷夾捕捉力。
13 3-3 樣品平台 我們使用一個閉迴路奈米級 XY 定位壓電平台被安裝在顯微鏡的平台上,以提供樣 品在幾個奈米解析度的平移運動。透過電腦控制壓電平台精確的移動薄鋁製載具,如圖 3-4,進而操控在薄鋁製載具上方以兩個蓋玻片間距 0.17 毫米厚封裝的生物樣品。其中, 薄鋁製載具中央有一個洞在中間,以避免平台移動時,雷射鑷夾捕捉具光的顯微物鏡推 動樣品,造成截取訊號干擾。 圖3-4 樣品平台裝置圖 3-4 系統控制程式 本實驗使用 Labview 軟體操控閉迴路奈米級 XY 定位壓電平台。圖 3-5 為「線毛受 外力的伸展曲線」之操作面板,圖左方為微粒子黏附線毛時,線毛受外力伸展曲線的X 軸即時四象限光偵測器訊號截取;上方為四象限光偵測器電壓訊號所對應的即時雷射鑷 夾捕捉力,亦即線毛所受外力;中間為實驗全程的完整訊號;右方則是使用者自由選取 訊號截取範圍,進一步分析微粒子 X、Y、Z 軸方向的位移訊號,可以幫助使用者判定 線毛是否只受一維方向外力。其中藍色訊號為線毛X 軸方向受力伸展曲線,紅色訊號為 線毛 Y 軸方向受力伸展曲線,而綠色訊號為線毛 Z 軸方向受力伸展曲線。由圖可知線 毛在X 軸方向受力伸展曲線有左右對稱的訊號,此為線毛伸展與收縮曲線,而 Y、Z 軸 訊號則呈現趨於平緩的曲線,這表示線毛在Y、Z 軸方向不受力。 將四象限光偵測器偵測微粒子的位移訊號,換算成雷射鑷夾捕捉力,則可得到線毛 受外力的伸展曲線,如圖3-6 所示。我們進一步將數個線毛受外力的伸展曲線數據分析 比較,由圖中可以看到不同長度線毛的受力伸展曲線幾乎有類似的受力伸展行為。由上 述操作系統,我們將可以分析線毛受外力的機械特性。
14
圖3-5 「線毛受外力的伸展曲線」之操作面板
15
第四章 實驗方法
4-1 第三型線毛與第四型膠原蛋白間黏附力 克雷白氏肺炎桿菌(Klebsiella pneumoniae)在臨床上是一株伺機性感染細菌[30],常 在免疫不全的病人身上造成感染。當克雷白氏肺炎桿菌要進行感染時,第一步便是先黏 附於宿主細胞上。而第三型線毛被發現是用來黏附的一個重要黏附因子。在第三型線毛 上,已經被發現其中有一個叫 mrkD 的對偶基因是負責調控線毛頂端的 MrkD 黏著素 (adhesin)。雖然對於 MrkD 黏著素的受體還沒有很確定,但是已經發現第三型線毛會黏 著於第四型及第五型膠原蛋白。在這個實驗中,我們將用雷射鑷夾來量測不同的黏著素 對黏附於第四型膠原蛋白的影響。由於野生型(wild type)的克雷白氏肺炎桿菌一次會表 現出許多型的線毛,因此為了真的能量到第三型線毛的黏附力,我們是將載有第三型線 毛的基因轉殖到大腸桿菌(E. coli) JM 109 菌株上。這一株的大腸桿菌原來並不會表現第 三型線毛。 目前已知在第三型線毛頂端有四種黏附蛋白 mrkD-v1、mrkD-v2、mrkD-v3 和 mrkD-v4,在本實驗中我們將這四種頂端黏附蛋白分批轉殖在大腸桿菌 JM109 上,使每 一批的每隻大腸桿菌只表現出單一種頂端黏附蛋白的線毛。而膠原蛋白是大部分上皮細 胞的底膜結構聚合體,普遍存在於宿主表面的細胞受器。接著我們利用雷射鑷夾分別量 測這四種黏附蛋白與膠原蛋白的結合力,並找出線毛主要黏附蛋白分子。我們主要的目 的是藉由雷射鑷夾測量線毛與膠原蛋白之間的黏附力,進一步分析線毛頂端黏附力。 首先,我們將直徑 1 微米微粒子表面塗覆膠原蛋白,以代替宿主的細胞。接著我們 將線毛頂端具有單一種黏附蛋白的大腸桿菌固定於載玻片上,然後用雷射鑷夾捕捉一顆 表面塗覆膠原蛋白的微粒子,先從遠處慢慢靠近一隻大腸桿菌,但避免碰觸,隨即拉離, 圖4-1 顯示。倘若微粒子有黏到大腸桿菌的線毛,則在微粒子遠離大腸桿菌的過程中, 微粒子偏移雷射鑷夾捕捉中心的偏移量D 將由四象光偵測器即時反應。因為雷射鑷夾對 微粒子的捕捉力等於微粒子表面膠原蛋白與線毛之間的黏附力,所以我們可以根據已校 正的雷射鑷夾捕捉力與微粒子偏移量 D 的線性關係,由微粒子偏移量 D 來判斷線毛與 膠原蛋白之間的黏附力,圖4-2 則為其所對應的四象限光偵測器訊號曲線。16 起初,線毛尚未被拉直之前,微粒子不受線毛拉力,微粒子一直維持在雷射鑷夾的 捕捉中心位置,如圖4-1(A)所示。在此一階段,因為微粒子未偏移,即偏移量 D =0,所 以四象限光偵測器反應的偏移量D 訊號曲線隨時間呈現水平直線,如圖 4-2(A)所示。而 圖4-1(B)顯示微粒子剛黏附線毛,而線毛蜷曲尚未被拉直,四象限光偵測器的訊號曲線 仍維持平衡位置,如圖4-2(B)所示。如圖 4-1(C),當線毛被拉直之後,微粒子被線毛黏 著拉離而開始偏移雷射鑷夾的捕捉中心,此時雷射鑷夾的捕捉力隨微粒子偏移量D 的增 加而增大,因此,在這個拉拔階段,四象限光偵測器反應的偏移量D 隨時間的訊號曲線 為一個斜直線,當雷射鑷夾捕捉力達到恰可將微粒子與線毛分開的力量時,四象限光偵 測器反應的偏移量D 訊號曲線達到一個峰值,如圖 4-2(C)。圖 4-1(D)表示雷射鑷夾捕捉 力稍大於微粒子與線毛之間黏附力的瞬間,被捕捉的微粒子將與線毛分開,並回到雷射 鑷夾的捕捉中心,此時,訊號曲線也會回到平衡位置,如圖4-2(D)所示。由四象限光偵 測器反應的偏移量D 訊號曲線的峰值,我們可以得到此時所對應的雷射鑷夾對微粒子的 捕捉力,即為微粒子表面膠原蛋白與線毛之間的黏附力。 圖4-1 雷射鑷夾測量線毛與膠原蛋白間黏附力實驗示意圖
(A)
(B)
(C) (D)
E coli
Collagen-coated
Optical
Tweezeers
QPD
D = 0
PZT XYZ-Stage
E coli
Collagen-coated
Optical
Tweezeers
QPD
D>
0
PZT
E coli
Collagen-coated
Optical
Tweezeers
QPD
D
=0
PZT XYZ-Stage
QPD
E coli
Collagen-coated
Optical
Tweezeers
D
= 0
PZT XYZ-Stage
17 圖4-2 雷射鑷夾測量線毛與膠原蛋白間黏附力的訊號曲線 4-2 第三型線毛在應力下的機械特性 細菌培養 將載有完整 mrk 基因操縱子的質體,利用轉殖技術使克雷白氏肺炎桿菌的第三型線 毛表現在大腸桿菌 JM109(pmrkABCD)的表面上。主要因為大腸桿菌本身並不具有第三 型線毛,而利用這樣轉化方式我們可以分析不同mrk 基因操縱子的表現行為。在線毛量 測實驗之前,我們將帶有第三型的大腸桿菌培養在攝氏溫度37 度的 LB 培養基中搖晃約 20 小時,然後直接用於 TEM 分析。對於彈性實驗,細菌離心 9000 轉/1 分鐘,洗兩次 用磷酸鹽緩衝液(10 mM PBS , pH=7.4, 含有 137 mM 氯化鈉、2.7 mM 氯化鉀),然後懸 浮在緩衝液中待後續使用。 電子顯微鏡 細菌在 LB 培養基中於 37℃培養 16 小時後,以 4,000g 離心收集細胞,並以ㄧ倍磷 酸緩衝液清洗一次,回溶於等體積之ㄧ倍磷酸緩衝液。取在清洗後復溶於磷酸緩衝液的 菌液2.5 微公升,滴在銅網(Formvarcarbon-coatedglow-dischargedgrid---200mesh)上,等 待1 分鐘使細菌沉澱於銅網後,以濾紙沿著銅網邊緣小心地將多餘的菌液吸掉,用 3 滴 蒸餾水,沾有2 滴 2%(質量/體積)醋酸鈾染劑,等待 5 秒鐘後,再以濾紙沿著銅網將染 劑盡量吸乾,之後以電子顯微鏡觀察。
-5
-4
-1
0
1
2
100 200 300 400 500 600 700 800 900 偏 移 量 (A.U.)Signal
時間(a.u.)-2
-3
(A) (B) (C) (D)
18 乳膠珠製備 黏附於細菌表面線毛的 1 微米橡膠小珠,表面塗覆 Protein A (Inc.,Warrington,PA), 我們已以下方式處理:首先,我們以 A/G 的緩衝液(0.1MTris-HCl,0.15M 氯化鈉)清洗 橡膠小珠,與反MrkD 抗體在 4℃培養 1 小時。接著,將表面塗覆 MrkD 抗體的橡膠小 珠再次以A/G 的緩衝液清洗,然後懸浮在磷酸緩衝液 PBS,並保持在 4℃以備後用。 樣品製備 蓋玻片以去離子水徹底清洗,待完全乾燥後,將兩個蓋玻片之間以兩層雙面膠帶間 隔開約 170 微米厚度。接下來,40µl 新鮮懸浮在 PBS 大腸桿菌注射到兩個蓋玻片之間 的空間,靜置 1 分鐘後,用濾紙完全去除在空間的 PBS,然後離開 30 分鐘,以確保固 定細菌於玻片底部。隨後,將內含 1 微米表面塗覆 ProteinA 橡膠小珠的 PBS 溶液 40 微 升,注射到兩個蓋玻片之間。最後,將樣品放入光學顯微鏡樣品平台 1 小時,在彈性測 量實驗前,再次確保附著細菌的最底層蓋玻片。 雷射鑷夾 雷射鑷夾裝置圖如前面實驗所示。基本上,捕捉雷射光束和探測雷射光束被引導到 一個倒立式光學顯微鏡,然後通過一個高 NA 物鏡,在樣品垂直面上形成一上一下兩個 焦點。接著,利用捕捉雷射光束捕捉表面塗覆 ProteinA 的橡膠小珠。探測雷射光束則用 於偵測橡膠小珠與捕捉雷射中心的位移,用以分析捕捉雷射捕捉力。 為了量測線毛的彈性以及楊氏模數,我們必須讓細菌固定位置,而其線毛的一端從 細菌的外膜生長出來,另一端與微粒子緊密黏附。接著,我們使用雷射鑷夾將已黏附線 毛的微粒子捕捉搬運使之遠離大腸桿菌,讓線毛產生形變,進而量測其機械特性。 首先,我們將一隻大腸桿菌黏附固定於玻片底部,再利用雷射鑷夾捕捉一顆 1 微米 微粒子去黏大腸桿菌的線毛,微粒子表面塗附Nhydroxysuccinimide(NHS),NHS 對於生 物分子有極強的黏附性,因此微粒子與大腸桿菌的線毛將緊密黏附。接著,我們利用雷 射鑷夾拉著這顆黏牢線毛的微粒子使之遠離大腸桿菌,如圖4-3 所示。 當雷射鑷夾捕捉這顆已黏附線毛的微粒子並遠離大腸桿菌時,線毛從蜷曲而拉直最 後被拉長。如圖4-4 所示,在四象限光偵測器的訊號曲線中,一開始線毛蜷曲時的彈力 為零,訊號保持水平平衡位置﹔而線毛被拉直而拉長的過程,訊號開始偏移平衡位置, 此時線毛彈力增加,雷射鑷夾對微粒子的捕捉力也增加。最後,當雷射鑷夾對微粒子的 捕捉力不足以將微粒子拔離線毛時,微粒子將脫離雷射鑷夾的捕捉中心。
19 圖4-3 利用雷射鑷夾操控黏附線毛的微粒子示意圖 圖4-4 單一方向線毛彈力實驗的訊號曲線 圖 4-4 為同一根線毛經過數次的單一方向拉拔所得到的數據圖,每一種顏色曲線代 表一次拉拔數據,以上共有十次,並顯示出具有高度重複性,這代表此線毛在受力下, 仍是具有恢復力。 如圖 4-4(A)所示,我們利用雷射鑷夾捕捉一顆已黏附大腸桿菌線毛的微粒子向+X 軸方向移動,使之遠離大腸桿菌,直到微粒子脫離雷射鑷夾捕捉中心為止,其中不同的 顏色是同一顆微粒子的重複實驗數據﹔而圖4-4(B)則是我們利用雷射鑷夾捕捉同一顆微 粒子向+Y 軸方向移動的重複實驗。我們分別將 X 軸以及 Y 軸方向各九次重複實驗數據 重疊比對,發現位移對捕捉力的訊號曲線幾乎重疊在一起。這代表兩個訊息,第一個訊 息是證實我們的儀器有高度穩定度,而另一個則表示線毛在重複的拉扯過程中,並沒有 從微粒子上脫落,而線毛也保持相同的彈性。 Optical Tweezers QPD D<0 E coli
Signal D [X] Signal D [Y]
(A)
(B)
偏 移 量 (A.U.) 偏 移 量 (A.U.) 時間(A.U.) 時間(A.U.)20 接著,我們利用雷射鑷夾操控一顆微粒子去黏附大腸桿菌的線毛,在相對於大腸桿 菌的兩側作來回一次的拉扯實驗,最後回到原來位置。圖4-5(A)~(E)為雷射鑷夾操控一 顆已黏大腸桿菌線毛的微粒子,從大腸桿菌最左側(A),拉置最右側(E)。 圖4-5 線毛長度量測實驗示意圖 我們利用雷射鑷夾操控一顆已經黏附大腸桿菌線毛的微粒子,在相對於大腸桿菌 X 軸方向的左右作拉扯,可以發現四象限光偵測器的X 軸、Y 軸、Z 軸訊號出現有趣的變 化。如圖4-6(A)所示,在四象限光偵測器的訊號曲線中,我們發現訊號會隨著移動的方 向改變。 在與雷射鑷夾移動方向相同的 X 軸訊號,有相對於訊號平衡位置的上下對稱性﹔而 另兩維訊號Y 軸、Z 軸則是相對於紅色虛線左右對稱。當雷射鑷夾捕捉微粒子在細菌位 置 X 軸左方移動時,訊號的方向為正,亦即受力方向向右﹔而在 X 軸右方移動時,訊 號的方向為負,亦即受力方向向左。而在Y 軸、Z 軸方向,四象限光偵測器的訊號方向 不變,因此線毛受力方向不會隨著位移方向而改變。 QPD D>0 E coli QPD D>0 E coli Optical Tweezers Optical Tweezers Optical Tweezers QPD D<0 E coli QPD D=0 E coli QPD D<0 E coli
(A)
(B) (C)
(E) (F)
Optical Tweezers Optical Tweezers21 圖4-6 線毛彈性以及楊氏模數實驗數據及示意圖:(A)訊號變化;(B)線毛受力模型。 另外,在圖 4-6(A)中,我們發現位移對捕捉力的訊號曲線在位移方向的訊號有上下 對稱性,而另兩軸則是左右對稱。這些訊號對有助於分析線毛長度、線毛在細菌體表線 毛根部生長位置、以及線毛長度。另外國際上在與線毛有關的研究都是一維的生物力分 析,我們將發展三維的訊號分析,這樣一來將可以得到一些一維訊號所得不到的訊息。 由訊號方向與位移方向相反,而訊號方向即為線毛受力方向,因此我們得知雷射鑷夾捕 捉微粒子的位移方向與線毛受力方向相反﹔而另兩維對應的訊號有相對重複性,代表其 受力的方向相同。
X-signal
Y-signal
Z-signal
0 0.5 1 1.5 2 2.5 3 3.5 4 1 5 9 13 17 21 25 29 33 37 41 45 49(A)
(B)
A B C D ESignal D
時間(A.U.) 偏 移 量 (A.U.)Bacterium
Pilus
Coverslip
Optical Tweezers
Bead
22 我們找出訊號對稱中心位置以虛線 C 標示,在開始有受力訊號的地方以虛線 B 及 虛線D 標示﹔而受力訊號最大的位置以虛線 A 及虛線 E 標示。其中,訊號對稱中心位 置C 表示線毛根部在雷射鑷夾捕捉微粒子移動直線上的垂足,而 BC 以及 CD 則代表線 毛長度在移動直線上的投影長﹔而 AB 以及 DE 則是線毛受微粒子拉扯的兩端受力訊 號,如圖4-6(A)所示。 如圖 4-6(B)所示為線毛受力模型,利用雷射鑷夾將已黏附線毛的微粒子左右拖曳到 恰不能捕捉為止,我們發現在拖曳的過程中有一段線毛不受力的區間,以BD 表示,此 為線毛長度在移動直線上的投影長。而當線毛開始受微粒子拉扯的區間為AB 以及 DE, 此時線毛受外力而拉長,綠色箭頭是微粒子受線毛的張力方向。 利用 AB 及 DE 之間線毛受外力而拉長的受力伸展曲線,我們可以推算出線毛在受 力方向上的彈力。圖4-7 線毛受力而伸長的示意圖,我們假設線毛彈性係數Kt,而雷射 鑷夾捕捉力常數為Kt。線毛伸長量X 以及微粒子偏移雷射鑷夾捕捉中心的位移 d,兩P 者的和即為平台位移X,X = X + d。線毛伸長量可表示為P X = X - d P 因此線毛彈力可表示為Fp =Kp(X -d),雷射鑷夾捕捉力為Ft = Ktd 。 而我們由四象限光偵測器的訊號曲線可以求出力對位移訊號的等效係數K ,則訊號曲eqv 線等效的力F=KeqvX 。而線毛彈力、雷射鑷夾捕捉力以及訊號曲線等效的力都相等 t p F F F = = ,最後我們得到等效彈性係數、雷射鑷夾彈性係數、與線毛彈性係數之間的 關係Keqv−1 =Kt−1+Kp−1。 圖4-7 線毛受力而伸長的示意圖[33]
23 4-3 第三型線毛主要蛋白氨基酸序列突變的影響[42] 點突變與實驗設計[42] 第三型線毛骨幹是由主要蛋白 MrkA 組成。胺基酸是構成蛋白質的基本單位,賦予 蛋白質特定的分子結構形態,使他的分子具有生化活性。蛋白質是由由二十種氨基酸一 個個相連組成而在立體空間上摺疊產生複雜的構造,這些胺基酸又被稱為殘基。每一種 蛋白質都有自己獨特的胺基酸序列,如圖 4-8 為第三型線毛主要蛋白 MrkA 胺基酸組裝 示意圖。我們將第三型線毛主要蛋白特定胺基酸進行點突變,分析胺基酸點突變對於線 毛堆疊以及線毛功能的影響,進而推斷這些胺基酸在線毛組裝的過程中所扮演的角色。 圖 4-8 第三型線毛骨幹蛋白胺基酸序列示意圖 利用定點突變技術,找出線毛主要蛋白分子MrkA 上與線毛組裝有關的胺基酸位置 與區域進行十二個定點突變,再進一步探討不同點突變株對於MrkA 交互作用以及線毛 正常功能的影響,包括進行生物膜(Biofilm)形成能力以及血球凝集能力的測試,此樣品 為清華大學生命科學系張晃猷老師實驗室所提供。首先,將大腸桿菌野生株JM109 和十 二個氨基酸點突變體細菌,生長在LB 培養基 (Luria-Bertani)於 37℃,並依據需要加入 100 μg/ml 的青黴素(ampicillin)。 除了線毛不可或缺的 β-鏈組成結構之外,在其他種類線毛的主要主要蛋白中的幾個 保守序列,也在第三型線毛主要蛋白Mrk A 中發現。各種線毛的主要主要蛋白 MrkA,
24 致)。這意味著不同類型的線毛的序列差異可能造成一些明顯的性能相異,例如,力學 性能可能會有所不同。為了檢視線毛主要蛋白Mrk A 的重要殘基,我們選定 MrkA 序列 的12 個位置,以丙氨酸取代 MrkA 殘基作定點突變。 如圖 4-9 所示,從五種類型線毛骨幹主要蛋白比較中,基於兩個標準選擇,分別以 丙氨酸的替換的十二個殘基突變體(1)幫助骨幹主要蛋白間相互作用的重要殘基 G30, F34,C87,G189,Y197 和 I199;(2)從五種類型線毛骨幹主要蛋白比較中,找出屬性不 一致的六個重要殘基V38A,V45A,S46A,N192A,T196A,和 Q202A。本研究所使 用的菌株與質體之基因型與特性,本實驗所使用的引子如表4-1 所示。 生物膜形成能力測試 細菌在 LB 中於 37℃培養 16 小時後,以 LB 將菌液稀釋 10 倍,並調至相同濃度。 接著,分別取100 μl 的上述菌液加入 96 孔平盤內,於 37℃靜置培養 48 小時。待生物 膜生成後,加入染劑於室溫下10~15 分鐘,再以清水洗三次,接著加入酒精搖晃均勻後, 測定其吸收光值,此方式換算突變株與野生菌株生物膜形成能力的相對數值。
25
圖4-9 各種線毛的線毛單元體的胺基酸序列比較圖[42]
比較五大線毛骨幹主要蛋白序列。mrkA:克雷白氏肺炎桿菌第三型線毛骨幹主要蛋白;
fim A:大腸桿菌線毛第一型線毛主要蛋白;pap A:大腸桿菌 P 型線毛主要蛋白; lpfA:沙門氏傷寒桿菌 LPF 線毛主要蛋白;MrpA:奇異變形桿菌 PMF 線毛主要蛋白。
黑色垂直框表示相同的殘基和灰色的垂直框表示類似殘基。α-螺旋和 β-鏈被標在序列上
面的信號肽,並取代殘留的標有“A“部分。黑色代表相同的胺基酸;灰色代表保留性胺 基酸。訊息序列以( )作代表。序列上方二級結構( )表示 α-helix; ( )表示 β-strand。序列下方的字母與位置則表示置換成的胺基酸與置換位置。
26
Strains Primer Sequence 5′→3′
G30A SL1073 AATGTTGGCGGCGCCCAGGTTAATTTCTTCGGTA SL1074 TACCGAAGAAATTAACCTGGGCGCCGCCAACATT F34A SL1075 GCGGCGGCCAGGTTAATGCCTTCGGTAAAGTTAC SL1076 GTAACTTTACCGAAGGCATTAACCTGGCCGCCGC V38A SL1091 AATTTCTTCGGTAAAGCTACCGATGTATCCTGT SL1092 ACAGGATACATCGGTAGCTTTACCGAAGAAATT V45A SL1095 TGTATCCTGTACTGCTTCCGTTAACGGCCA SL1096 TGGCCGTTAACGGAAGCAGTACAGGATACA S46A SL1097 TGTATCCTGTACTGTAGCCGTTAACGGCCAGG SL1098 CCTGGCCGTTAACGGCTACAGTACAGGATACA C87A SL1079 ACCGAAATCTTTCACCGCTGACGTCTCTAACTGCCA SL1080 TGGCAGTTAGAGACGTCAGCGGTGAAAGATTTCGGT G189A SL1099 CACGGTAACCACCGCTGTGGTTAACAGC SL1100 AGCTGTTAACCACAGCGGTGGTTACCGTG N192A SL1101 CACCGGTGTGGTTGCCAGCTACGCGACTTA SL1102 TAAGTCGCGTAGCTGGCAACCACACCGGTG T196A SL1103 GTGGTTAACAGCTACGCGGCTTACGAAATTACTT SL1104 AAGTAATTTCGTAAGCCGCGTAGCTGTTAACCAC Y197A SL1081 TTAACAGCTACGCGACTGCCGAAATTACTTACCAG SL1082 CTGGTAAGTAATTTCGGCAGTCGCGTAGCTGTTAA I199A SL1105 GCTACGCGACTTACGAAGCTACTTACCAGTAATT SL1106 AATTACTGGTAAGTAGCTTCGTAAGTCGCGTAGC Q202A SL1085 GACTTACGAAATTACTTACGCGTAATTCCGTCAGAA SL1086 TTCTGACGGAATTACGCGTAAGTAATTTCGTAAGTC 表4-1 點突變所使用的引子[42]
27
第五章 結果與討論
5-1 第三型線毛與第四型膠原蛋白間黏附力 如圖 5-1,我們先以電子顯微鏡觀察 mrkD-v1、v2、v3 和 v4 四種帶有不同頂端黏附 蛋白線毛的大腸桿菌,,我們發現各種大腸桿菌的線毛生長濃密度以及長度並不相同。 細菌與宿主細胞之間的黏附力可能會因為線毛表現疏密度而不同,一般來說,單一隻細 菌的表面線毛分布較稀疏,對於細菌與宿主細胞的黏附力較小﹔而線毛表現較密集的細 菌,則黏附力提高。我們主要的目的是藉由雷射鑷夾測量不同頂端黏附蛋白線毛與膠原 蛋白之間的黏附力,再進一步分析單一線毛頂端黏附力。 圖5-1 四種大腸桿菌的線毛生長密度電子顯微鏡照片[43]: (A)mrkD-v1;(B)mrkD-v2;(C)mrkD-v3;(D)mrkD-v4。 我們利用雷射鑷夾分別量測只表現出單一種頂端黏附蛋白 mrkD-v1、v2、v3 和 v4 的四種大腸桿菌的黏附力各50 次。其中 mrkD-v1 線毛黏附力太小,無法用雷射鑷夾測 量出來。接著,我們將mrkD-v2、v3 和 v4 各 50 次的黏附力數據,作成數據統計長條圖, 我們發現mrkD-v3 線毛黏附力數據分佈最廣,而紅色虛線顯示黏力分佈有群組的現象, 如圖5-2。200 nm
(B) JM109 [mrkD-v2] (D) JM109[mrkD-v4] 200 nm (A) JM109[mrkD-v1] 500 nm (C) JM109[mrkD-v3] 200 nm 200 nm28 圖5-2 線毛黏附力的數據統計圖[34] 由上至下分別為:(A) mrkD-v2;(B) mrkD-v3;(C) mrkD-v4。 實驗的次數分別為 51,48,及 53 次。 我們假定每一根線毛的黏力相近但不相等,則黏附相同數量的線毛的黏力也相近。 黏力大小與黏附線毛的數量有關,因此,黏力大小應具有週期性且倍數關係的分佈,如 圖5-2 為線毛黏附力的數據統計圖(Histogram)。在黏力分佈統計圖中,每一群組黏力的 平均值,彼此之間也會有倍數關係。因此尋找適當的黏力分群將有助於分析單一根線毛 的黏附力。
29 我們進一步將 mrkD-v2、v3 和 v4 三種大腸桿菌線毛黏附力分布圖,再對力的出現 頻率作傅立葉轉換,可以得到力出現頻率的大小分佈圖,再將此分佈圖橫軸頻率取倒 數,即是力的群組週期分佈圖,如圖 5-3。力的群組週期分佈圖中,力的群組週期強度 的最大值對應的是2 pN、4 pN、以及 2.9 pN。我們以上述三組力的分群週期,作為線毛 黏力的分組依據,將黏力分佈統計圖重新分組,並取各組黏力的平均值,則為線毛黏附 數量不同的黏附力。 如圖 5-4 所示,我們接著將每一組線毛黏附力的平均值與其對應的線毛黏附數量作 圖。線毛黏附力對黏附線毛數量的斜率,即為單一線毛與膠原蛋白間黏附力的大小。我 們發現其中mrkD-v1 黏附力太小,無法用雷射鑷夾測量出來,而 mrkD-v2、mrkD-v3 和 mrkD-v4 單一線毛的黏附力分別是 2.03±0.03pN、3.79±0.12pN、以及 2.87±0.15pN。由此 可知mrkD-v3 是克雷白氏肺炎桿菌第三型線毛的四種頂端黏附蛋白之中黏附力最強。
30
圖5-3 黏附力的群組週期分佈圖[34]
31
圖5-4 線毛黏附數量與黏附力圖[34]:
(A) mrkD-v2 = 2.03±0.03 pN;(B) mrkD-v3 = 3.79±0.12 pN; (C) mrkD-v4 = 2.87±0.15 pN。
32 SinglePili (c) 20 nm LowDensity(mrkDV1) 1μm HighDensity(mrkDV3) Height (nm ) LowDensity(mrkDV1) 1μm (a) (b) 5-2 第三型線毛在應力下的機械特性 製備線毛與線毛結構確認 首先,我們完成線毛製備,並利用原子力顯微鏡及電子顯微鏡確認,大腸桿菌表現 第三型線毛的外型輪廓,其線毛直徑約 3~4 奈米、長度約 2~3 微米,且線毛生長密度分 佈可以控制在數根之內,如圖 5-5 所示。而第一型線毛或 P 型線毛的直徑約為 6.8 奈米、 長度約 1 微米,第三型線毛相對於第一型線毛與 P 型線毛較為纖細。 圖5-5 利用原子力顯微鏡(AFM)以及高解析度電子顯微鏡(HR-TEM) (清華大學材料科學研究所游萃蓉老師實驗室) 觀測線毛微結構,圖(a) & (b)為原子力顯微鏡影像, (a)高線毛密度的菌株(b)低線毛密度的菌株,其單一線毛直徑 ~ 3 nm; (c)則為高解析度電子顯微鏡(HR-TEM)影像。 如圖 5-5 所示,大腸桿菌表現 pmrkABCDv1F 之線毛的密度分佈,其菌體分別培養 在 LB 培養基(a)或者 GCAA 培養基(b)中。兩種線毛分佈分別可以觀察到在分佈密度上 明顯差異,其中培養在LB 培養基的菌體,其線毛密度較為稀疏,適合應用在單一線毛 量測實驗中。
33 第三型線毛的彈性 當我們利用雷射鑷夾技術,捕捉、搬移一顆黏附有線毛的乳膠珠後,我們發現線毛 在外力作用下將被伸長,且當外力消失之後,乳膠珠會受到線毛的彈性影響而被拉回原 來的位置附近。因此,當我們對同一顆黏有線毛的乳膠珠向某方向重複拉拔,擷取的訊 號具有高度的重複性。而當我們捕捉乳膠珠,並使乳膠珠以原平衡點對稱的兩個相反方 向上來回搬移,擷取的訊號具有高度對稱的訊號。 我們利用以上實驗數據結果,我們可以得到該線毛的原長約為 2 微米、線毛所受到 最大三維線性捕捉外力為F =16.5皮牛頓、線毛後來伸長長度約為 2.271 微米。最後換 得該線毛的彈性係數約為60.9 皮牛頓/奈米(k = /F ΔL,ΔL=2.271−2=0.271微米)。再 利用電子顯微鏡測量線毛的直徑,以及線毛長度等數據,線毛的截面積A= πφ2 / 4,我 們可進一步分析線毛的楊氏模數。 第三型線毛的楊氏模數 一個外力(F)施加在截面積為(A)、原長為(l)的物體,引發一個伸長量為(Δl),楊氏模 數可表示為 2 ) 2 / ( / / D l k l l A F E p π × = Δ = ,楊氏模數E(牛頓/平方公尺)適用於一維伸長之材料。 外力F (牛頓)施加在一根棒狀型的線毛上。其截面積 A (平方公尺) = π (D/2)2;直徑D (公 尺);l (公尺)為未伸長線毛之長度;Δl (公尺)為受力後之伸長量;kp (牛頓/公尺)線毛的彈 力常數。 在電子顯微鏡照片用一個高的解析度放大,可清晰的看到第三型線毛骨幹呈條紋 狀,如圖5-6。我們估計線毛的寬度大約 7.4 ~ 4.3 奈米。我們估計第三型線毛線毛直徑 寬度的變化,是因為線毛直徑為橢圓形扭轉造成,其結構有類似P型線毛或第一型線毛 般以主要蛋白為骨幹的的螺旋結構。
34 圖5-6 第三型線毛電子顯微鏡照片,線毛直徑 7.4 ~ 4.2 奈米[33] 利用電顯圖所獲得的截面積為A=πab平方奈米,初步估計該第三型線毛的截面積 為A=π×7.4 ×4.2 平方奈米,而楊氏模數約為 100 MPa。目前相關研究指出,第一型線 毛的直徑6.9 奈米,而楊氏模數約為 100 MPa(皮牛頓/微米2);而 P 型線毛的楊氏模數與 截面積的乘積約為 154 ± 20 皮牛頓,而線毛的直徑 8.2 奈米,可推算出楊氏模數約為 50-400 MPa(皮牛頓/微米2)[5]。因此,對於比較三種線毛的彈力特性,楊氏模數是可以 參考比較的指標。以上所估計第三型線毛的楊氏模數相當符合已發表的聚合物之彈性 [35],相關材料的楊氏模數歸奈在表 5-1,其中線毛在類彈簧伸長狀態時的彈性類似橡膠。 材料[36] 鐵 玻璃 木頭 骨骼 聚氯乙烯 橡膠 楊氏模數[MPa] 200000 65000 13000 9000 3000 1-10 表5-1 常見材料的楊氏模數 上述材料由左置右分別是鐵、玻璃、木頭、骨頭、聚氯乙烯,以及橡膠,所對應的楊氏 模數大小的程度由左至右遞減。由此可知,當材料之楊氏模數越大,所呈現出來的機械 特性會越堅硬。
35 第三型線毛的拆解力、特徵力 線毛(或稱線毛)是表現在細菌外膜的附屬物,他們協助細菌黏附宿主細胞並進行建 立感染。最近,一些研究聲稱P 型線毛與第一型線毛的機械特性能幫助維持細菌與宿主 細胞間的黏附,他們的伸長彈性已用雷射鑷夾或原子力顯微鏡證實[5-8]。此外,與 P 型 線毛、第一型線毛同屬第一類線毛的第三型線毛的機械特性,將在本論文中呈現。 受外力的伸長曲線分為三個階段:(1)第一階段類彈簧階段,(2)第二階段拆解階段,以 及(3)第三階段非線性階段,如圖 5-7 所示。除第一階段類彈簧階段,第三型線毛受外力 的伸長曲線的定性類似P 型線毛或第一型線毛。 圖5-7 單一根第三型線毛之典型受力伸長關係圖[33]: (1)第一階段類彈簧階段,(2)第二階段拆解階段,以及(3)最終非線性階段。 如圖 5-8(A)~(E)為雷射鑷夾操控一顆已黏附大腸桿菌線毛的微粒子,從大腸桿菌附 近(A),將微粒子逐步拉離大腸桿菌至雷射鑷夾捕捉力最大值(E)。其中圖(A)~(B)為線毛 受力第一階段類彈簧階段,此時線毛因受外力而有類似虎克彈簧的彈性伸長;圖(C)為 線毛受力第二階段拆解階段,此時線毛骨幹因受外力而逐一拆解伸長;圖(D)~(E)為線毛 受力第三階段非線性階段,此時線毛骨幹已完全拆解,接著線毛因受外力逐漸增加,其 骨幹分子與分子間的夾角隨機地拉開。
36 (A)- (B) (C)-1 (C)-2 (D)-1 (D)-2 (E) -圖5-8 典型的第三型線毛受外力的伸長示意圖 (A)彈簧原長 (B)類似虎克彈簧的彈性伸長(I) (C)線毛主要蛋白逐一拆解伸長(II) (D)主要蛋白分子間的夾角拉開(III) (E)主要蛋白間的夾角完全拉開(III) (A) (B) (C) (D) (E) Bead E.coli Adhesin
37 如圖5-9 所示,黑色實線顯示了一個典型的第三型線毛受外力的伸長行為。在第一 階段類彈簧階段,第三型線毛隨外力的增加而被拉伸。當外力增加至接近拆解階段,第 三型線毛的拉伸行為類似一個彈簧,此時線毛的最大彈性係數為 261.8 皮牛頓/微米 段。接下來,當外力達到拆解力,第三型線毛切換到拆解階段而大幅延長。在拆解階段, 線毛持續以一個穩定的拆解力 70.1 皮牛頓,將其螺旋結構拆解。在拆解階段後,線毛 受外力的伸長曲線有一個傾斜的S 形,此時有一個特徵力 116.4 皮牛頓。當外力超過特 徵力之後,拉伸力迅速增加超過幾百皮牛頓。在拆解階段,第三型線毛的拆解力以0.01 微米/秒或在 0.05 微米/秒的速度拉動,這表示,所有實驗均在穩定狀態下運行。 圖5-9 典型的第三型線毛受外力的伸長曲線[33] 此外,另一個典型的受外力的伸長曲線,顯示了線毛逐一拆解伸展至近 9 微米的 拆解階段,如圖5-10 所示。請注意,在伸展 3 微米的拆解階段,拆解力從 127.0 皮牛 頓下降到 67.1 皮牛頓,這表示微粒子黏附兩根線毛,當外力增加時,兩根線毛同時解 開,然後依次脫離微粒子,其黏附拆解過程如圖5-11。
38 圖5-10 兩根第三型線毛受外力的伸長曲線[33] 圖5-11 兩根第三型線毛受外力的伸長示意圖 我們將所有線毛受外力伸長曲線拆解力的測量結果繪製成頻率直方圖。圖 5-12(A) 分別為微粒子黏附一根線毛以及兩根線毛的兩組拆解力。線毛拆解力的平均值分別為 66.8 ± 3.6 皮牛頓(19 根線毛) 和 124.1 ± 8.8 皮牛頓(3 根線毛)。根據拆解力測量的最小 公倍數結果,在穩定狀態拆解力平均值為66.2 ± 4.0 皮牛頓(20 根線毛)。而線毛特徵力 發生在最後的非線性階段,如圖5-12(B)所示,線毛特徵力平均值為 101.9 ± 8.6 皮牛頓 (14 根線毛)。

![圖 4-9 各種線毛的線毛單元體的胺基酸序列比較圖[42]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8461539.183192/33.892.95.805.213.975/圖49各種線毛的線毛單元體的胺基酸序列比較圖42.webp)
![圖 5-3 黏附力的群組週期分佈圖[34] (A) mrkD-v2;(B) mrkD-v3;(C) mrkD-v4。](https://thumb-ap.123doks.com/thumbv2/9libinfo/8461539.183192/38.892.97.800.111.1011/圖53黏附力的群組週期分佈圖34AmrkDv2BmrkDv3CmrkDv4.webp)
![圖 5-4 線毛黏附數量與黏附力圖[34]:](https://thumb-ap.123doks.com/thumbv2/9libinfo/8461539.183192/39.892.99.798.147.1039/圖54線毛黏附數量與黏附力圖34.webp)


