利用HeLa細胞探討Mst3誘導細胞凋亡之分子機制
56
0
0
全文
(2) 探討 Mst3 在 HeLa 細胞誘導細胞凋亡之分子機制 Molecular Mechanism study of Mst3-induced Apoptosis in HeLa cells. student : Pei-Ling Wang. 研究生 : 王珮玲. Advisors : Dr. Chiun-Jye Yuan. 指導老師 : 袁俊傑博士. 國立交通大學 生化工程所 碩士論文. A thesis Submitted to Institute of Biochemical Engineering National Chiao Tung University In Partial Fulfillment of Requirement For the Degree of Master of Science In Biochemical Engineering July 2004 Hsinchu, Taiwan, Republic of China. 中華民國九十三年七月 2.
(3) Mst3 誘導細胞凋亡機制之探討. 摘要. Mst3 為 52KDa 蛋白質,隸屬人類 STE20 相似蛋白激酵素 (human Ste20 like protein kinase)家族中的一員,在細胞凋亡過程中扮演重要角色。它具有 N 端的激酵素功能區域及 C 端的調控功能區域,之前研究指出 Mst3 一般存在於細胞質且會被 caspase 活化,且大量 表現 Mst3 野生株及截斷株,皆可誘發細胞凋亡之產生。 實驗室便利用廣泛性 caspase 抑制劑 (Z-VAD-fmk) 處理細胞,結果我們發現 Mst3 所導 致的細胞凋亡並不會被抑制,而且 Mst3 與 caspase-3 共同表現也不會促進細胞凋現象產生 亡,因而推測 Mst3 引起的細胞凋亡或許並非透過 caspase 所誘發或者兩者機制是並行存在 的。 另一造成細胞凋亡之訊息傳遞途徑則是透過粒腺體途徑 (mitochondrial pathway),之前 研究指出,當細胞遭遇細胞凋亡時,粒腺體內的膜電位 (membrane potential) 會改變、滲透 壓及鈣離子濃度亦會提高,進而導致部份粒腺體凋亡因子,如 cytochrome C、AIF 游離出來。 而這些凋亡因子則會受到一些粒腺體蛋白調控,我們研究以免疫染色發現 Cytochrome C 是 會與 Mst3 重疊,但 AIF 及 Endo G 則無此現象發生。而 Bcl-2 蛋白質會於 Mst3 存在時,細 胞質中的量也會相對減少。我們更意外發現:Mst3 不僅存在於細胞質中,也可在粒腺體中 發現。故便需進一步以釐清 Mst3 誘發的凋亡現象中,粒腺體所扮演的角色。. 3.
(4) Molecular Mechanism Studies of Mst3-induced Apoptosis in HeLa cell. Abstract Mst3, a novel human Ste20-like serine/threonine protein kinase, plays an important role in the process of cell apoptosis. It contains a kinase domain at its N-terminus and a regulatory domain at the C-terminus. Pervious studies have shown that overexpression of Mst3 or its truncated mutant induced apoptosis of HeLa and HEK293 cells. Further studies showed that the Mst3-indued apoptosis did not involve in any known MAPK cascades. Caspases are the central executioner of most classical apoptosis pathway, but its role in Mst3-induced apoptosis of HeLa cells is controversial. The result from this work showed that the universal caspase inhibitor, Z-VAD-fmk, could not supposed Mst3-induced HeLa cell apoptosis suggesting a caspase-independent pathway might exist. Mitochondria play an essential role in trigging caspase-dependent or independent apoptotic process. Many apoptotic factors are present in the mitochondrial intermembrane space. Once activated by apoptosis stimulus, these apoptotic factors may translocate from mitochondria to the cytoplasm or nucleus. The translocation of these apoptotic factors can be regulated by the Bcl-2 family proteins. Our results demonstrated that Mst3 is co-localized with cytochrome C in the mitochondria. This observation indicates that Mst3 may induce apoptosis by altering the cellular localization of cytochrome C. Interestingly the protein level of Bcl-2 was also reduced with the overexpression of Mst3. The subcellular redistribution of other apoptotic factors in response to the overexpression of Mst3 is unknown. Subsequent studies, we found that staurosporine may act differently to induce nuclear translocation of Mst3 as well as AIF proteins. Bcl-2 in nucleus was also increased with the stimulation of staurosporine. The role of Mst3 in mitochondria is unknown so far. The existence of Mst3 in mitochondria may provide a new aspect about the physiological roles of Mst3.. 4.
(5) Content Page. Chinese abstract. 3. English abstract. 4. Content. 5. Abbreviation. 6. Induction. 8. Materials and Methods. 14. Result. 21. Discussion. 26. Reference. 29. Appendix. 52. 5.
(6) Abbreviations. BSA. Bovine- serum albumin. Caspase. cystein aspartate-specific proteases. DAB. 3,3’Daminobenzidine. DMSO. Dimethyl sulfate. DMEM. Dulbecco’s modified eagle medium. DTT. Dithiothreitol. DIABLO. direct IAP binding protein with low pI. ER. endoplasmic reticulum. EDTA. Ethylene diamine tetra-acetic acid. EGTA. Ethlyene glycol-bis-(β-aminoethylether) N, N-tetra acetic acid. GFP. Green fluorescent protein. GCK. Germinal center kinase. Mst. Mammalian Ste20-like protein kinaes. PTP. permeability transition pore. PAGE. polyacryamide gel electrophoresis. PBS. phosphate-buffered saline. PMSF. phenylmethylsulfonyl fluoride. PVDF. polyvinylidine difluroide. SDS. sodium dodecyl sulphate. Smac. second mitochondrial activator of caspases. TNF. tumor necrosis factor. TdT. Terminal deoxynucleotidyl transferease. Tris. 2-amino-2-(hydroxymethyl)-1,3-propanediol 6.
(7) TUNEL. TdT-Utp Nick End Labeling. z-VAD-fmk. z-Val-Ala-Asp-fluormethylketone. 7.
(8) Introduction Apoptosis (or programmed cell death) is a controlled type of cell death that can be induced by a variety of physiologic and pharmacological agents (Kerr et al., 1972). It is an essential physiological process that plays a critical role in controlling the number of cells in embryonic development, differentiation, tissue homeostasis (Hengartner, 2000), and defense against pathogens (Zimmermann et al., 2001). The dysregulation of the apoptotic cell death may be involved in the pathogenesis of a variety of human diseases, including cancers, autoimmune diseases, neurodegenerative disorders and viral infections. The cell undergoing apoptosis exhibits cellular shrinkage, nucleus condensation, DNA fragmentation and cytoplasm vacuolization. The center of the apoptotic executive machinery is a family of cysteinyl aspartate-specific proteases, termed caspases (Nicholson and Thornberry, 1997). Thus far, the caspase gene family contains 14 mammalian members, which have an unusual and absolute requirement for cleavage at the bond right after aspartic acid (Asp) residues (Stennicke and Deveraux, 1998). Caspases can be divided in two groups according to their roles in apoptosis, the apoptotic initiators (caspase 8, 9 and 10), functioning to activate downstream effector caspases, and the apoptotic effectors (caspase 3, 6 and 7) (Ashkenazi and Cixit, 1998). Effector caspases are responsible for dismantling many cellular components, including proteins important for structure maintainance, cell survival, and for chromatin and nucleus integrity. (Thornbery et al., 1998, Blatt and Glick, 2001). For example, the cleavage of the ICAD (inhibitor of caspase-activated deoxyribonuclease) by caspase 3 is responsible for DNA fragmentation during cell apoptosis (Enari et al., 1998). Caspases also induce the disruption of cell structure by cleaving several 8.
(9) cytoskeletal proteins and their regulatory proteins (Kothakota et al., 1997), focal ashesion kinase (FAK) (Wen. 1997) and p21-activated protein kinase 2 (PAK2) (Rudel and Bokoch, 1997). Notably the activity of caspases are regulated by the Bcl-2 family proteins. The Bcl-2 family proteins can be classified into two sub-groups based on their roles in apoptosis. The anti-apoptotic Bcl-2-like proteins, including Bcl-2, Bcl-xL, Bcl-xS, Ced-9, Bcl-w, and Mcl-1, inhibit the apoptosis by preventing cytochrome C from exiting mitochondria and revoking the apoptosis. The pro-apoptotic Bcl-2-like proteins, such as Bcl-2 homology (BH) 1-3, Bax like proteins, Bak and Bok (consisting only the BH3 region) and Bad like proteins, Bid, Bik, Bim and Hrk, induce the apoptotic cell death. The Bax and Bax-like proteins, promote. of. cytochrome. C. from. mitochondria. which. activates. the. caspase-9-dependent apoptosis cascade. Notably the initiation of apoptosis depends on the ratio of Bcl-2 and Bax proteins in the cells. The predominance of pro-survival Bcl-2 proteins over pro-apoptotic Bcl-2-like proteins in the cells may block the initiation of apoptosis, whereas the predominance of the Bax protein in the cytoplasm promotes the implementation of death signals. AIF has a mitochondrial localization signal at its N-terminal and is confined to the mitochondria in the absence of apoptotic signal. Once imported into the mitochondrial intermembrane space, the mitochondrial localization signal of AIF is cleaved off to generate a mature 57 kDa AIF (Wu et al., 2002). Mature AIF translocates to the nucleus in response to apoptotic stimuli and causes DNA fragmentation (Susin et al., 1999). Endonuclease G, similar to AIF, can translocate to the nucleus and facilitate the DNA fragmentation (Li et al., 2001). Another mitochondrial factor, Smac (also called DIABLO), involves in apoptotic process by inactivating the inhibitor of apoptosis (IAP) (Mackenzie et al., 2000). All of these factors induce apoptotic cell death in response to apoptotic stimuli 9.
(10) through a caspase-independent pathway as implicated by the observation that the apoptosis induced by these factors cannot be suppressed by the pan-caspase inhibitor Z-VAD-fmk (Wu, 2002; Penninger and Kroemer, 2003). The cell death receptors, including TNF-α receptor (TNF-R) and Fas-ligands receptor Fas/APO-1/CD95 conduct apoptotic signals by converting pro-caspase 8 to the active one, which in turn activates Bcl and Bax. The activation of DEDs, DED1 and DED2, trigger a signaling cascade that modify and activate proteases ICE/CED-3. The first step is the conversion of the non active pro-caspase-8 into the active caspase-8. The caspase-8 then activates caspase-3 and Bid. The release of cytochrome C from mitochondria can be induced by the formation of channel-like structure of activated Bid and Bax complex on the surface of mitochondrial outer membrane. Cytochrome C then complexes with Apaf-1 and recruits caspase-9, forming the apoptosome (Zou H et al., 1997; Li P et al., 1997). The activated caspase-9 in turn actives caspases 3, 6, and 7 induce cell apoptosis. (Ravagnan et al., 2002). The endoplasmic reticulum (ER) is an essential organelle that. regulates. protein synthesis, lipid synthesis, cellular responses to stress, and calcium (Ca2+) storage (Berridge, 1995; Rapport, 1992). A variety of stimuli, including Ca2+ ionophore (A23187), inhibitor of glycosylation, chemical toxicants and oxidative stress, can induce stress on ER and ultimately lead to cell death (Liu et al., 1998; McCormick et al., 1997). Caspase 12 is thought to associate with ER (Nakagawa et al., 2000) and involve in the apoptosis-induction by the stress on ER (Nakagawa et al., 2000; Yoneda et al., 2001). The mechanism underlying caspase 12 activation is unclear so far. However, two possible mechanisms are proposed. One mechanism may involve the binding of caspase 7 and caspase 12 to GRP78 on ER surface resulting in autoactivation of csapase 12 (Yoneda et al., 2001). Alternatively, the 10.
(11) activation of casapse 12 may be mediated by calpain (Nakagawa et al., 2000). Apoptosis can be induced in several ways, some clearly involving Ca2 . This, +. coupled to the accepted role of proteolysis in the degradation phenomena associated with the death of cells has led to frequent suggestions of possible role for calpains. More significantly, dexamethasone was found to result in calpain activation, as determined by the increase in Ca2+–dependent proteolysis of added substract and, especially, by the disappearance of the 80 kDa form of calpain in favour of the activated autolytic products (Ernesto and Maurizio, 1998). Msts (mammalian Ste20-like protein kinase) are human counter parts of the budding yeast Ste20 protein kinase that involve in the pheromone response. Notably, Ste20 serves as a link between the G protein and MAPK cascade (Ramer et al., 1993). At least 30 members of mammalian Ste20-like protein kinases have been identified and cloned. They can be divided into two subfamilies based on the locations of their kinase domains: the p21-activated kinase (PAK) subfamily and germinal center kinase (GCK) subfamily. The PAKs consist of a C-terminal kinase domain and an N-terminal regulatory domain, with a small GTPase Rac1/Cdc42 binding region. GCKs possess an N-terminal kinase domain and a C-terminal regulatory domain with unknown function. Several lines of evidence have shown that some Ste20-like protein kinases function as potential upstream activations of MAPK pathways (Shi et al., 2000). Other than this ability to induce cell proliferation,. mitogen-activated. protein. kinases,. including. extracellular. signal-related kinase (ERK1/2), JNK/SAPK and p38 MAPK, were implicated to play role in the progression of apoptosis. (Kyriakis et al., 1996; Fanger et al., 1997). Four Mst family members have been identified, including Mst1/Krs2 (56 kDa), 11.
(12) Mst2/Krs1 (56.5 kDa), Mst3 (47.5 kDa) and Mst4/MASK (46 kDa). They can be classified into two GCK subfamilies, GCKII (Mst1 and Mst2) and GCKIII (Mst3 and Mst4), based on the sequence homology (Dab et al., 2001). Mst1, Mst2 and Mst3 have been shown to be involved in the process of cell apoptosis, while the role of Mst4 is controversial (Lin et al., 2001). Mst1 and Mst2 can be activated by apoptotic agents, including staurosporine, Fas-ligand, okadaic acid, synthetic apoptotic peptide MT-21, anti-cancer drugs and environmental stresses, such as heat shock and high concentrations of sodium arsenite (Graves et al., 1998; Reszka et al., 1999). With the stimulation of apoptotic signals, Mst1 can be prototypically activated by caspases, which can be specifically blocked by the selective caspase inhibitor (Graves et al., 1998; Lee et al., 2001). Upon activation Mst1 may transmit the apoptotic signal through either MEKK1-based MAPK pathway (Graves et al., 2001) or the JNK/SAPK pathway. Mst4, a member of GCKIII subfamily, was found to facilitate cell growth and transformation via MEK/ERK pathway (Lin et al., 2001). However, recent studies in our laboratory have shown that Mst4 may also play a role in triggering the apoptosis of HEK293 and MCF-7 cells (Chen, P. H. 2003). Mst3, a novel human Ste20-like serine/threonine protein kinase, shares 88% sequence similarity to the kinase domain of SOK1 and Mst4. Pervious studies have shown that overexpression of full-length or truncated Mst3 induced apoptosis of HeLa and HEK293 cells. Subsequently, a caspase-cleavage site in Mst3 was identified at AETD313 (Huang et al., 2002). After truncation, Mst3 translocates from cytoplasm to nucleus. Further works were carried out to elucidate that Mst3 contained NLS (nucleus localization signal) and NES (nucleus exporting sequence) signals at the C-terminus of its kinase domain (278-292) and in the C-terminal regulatory domain, respectively (Huang et al., 2002; Wang et al., 2004). 12.
(13) Furthermore Mst3 does not activate any known MAPK pathway (Schinkmann et al., 1997; Huang et al., 2002). Although the molecular mechanism underlying the Mst3-induced cell apoptosis is unclear so far, caspases were shown to play a role in activating Mst3 in Jurkat cells (Huang et al., 2000). Caspases 3, 7 and 8 were suggested to activate Mst3 by removing it C-terminal regulatory domain. The cleavage of the C-terminal regulatory domain triggered the nuclear translocation of truncated Mst3WT. △314. .. Hence, it will be interesting to understand the roles of caspases in Mst3-induced cell apoptosis and nucleocytoplasmic translocation of Mst3. Apoptosis can be initiated by disrupting the integrity of mitochondria, activating the cell surface death receptor and by endoplasmic reticulum (ER) stress-mediated cell death (Rammohan, 2001). Mitochondria play an important role in trigging apoptotic process. At the onset of apoptosis, the mitochondrial membrane potential is altered and the apoptotic factors relaesed in the intermembrane space. The factors include cytochrome C, apoptosis inducing factor (AIF), endonuclease G (EndoG) and Smac/DIABO. Since mitochondria play a central role in the pathways of caspase-dependent and caspase-independent cell apoptosis. We like to find out the role of mitochondria and the mitochondrial apoptotic factors in the Mst3-induced cell apoptosis.. 13.
(14) Materials and Methods Cell Line and Cell Culture HeLa (Human cervical cancer cell line) cell line was purchased from Food Industrial Research and Development Institute, Hsin-Chu, Taiwan. HeLa cells were grown in the DMEM medium containing 10% FBS and 1% each of penicillin and streptomycin. Cells were incubated at 37 ℃ in a humidified incubation chamber containing 95% air and 5% CO2. Cells were sub-cultured every 2-3 days.. Chemicals and Reagents Caspase inhibitor (z-vad-fmk) was the product of Merck; Inhibitors for p38-MAPK (SB220025) and a PD98059 inhibitor for Jun-N-terminal kinase (JNK), were purchased from Biosource. The TdT-FragEL DNA Fragmentation Detection Kit was obtained from Oncogene. Specific antibody for Bax was purchased from Biosource. Antibody for Bcl2 was obtained from Serotec. Antibody for AIF was brought from Santa Cruz. The antibody for cytochrome c oxidase subunit IV (COXIV) was purchased from Molecular Probes. The antibodies for Bcl-XL, Bad, Bid and EndoG and Cytochrome C Releasing Apoptosis Assay Kit were brought from BioVision, Rhodamine (TRITC) was purchased from Santa Cruz. MitoTracker Red dye was purchased from Molecular Probes. Fetal bovine serum (FBS) and Dulbecco’s modified egale medium (DMEM) were purchased Hyclone. The trypsin-EDTA, penicillin and streptomycin and lipofectamine reagent were brought from GIBCO BRL. G418 was product of Calbiochem. NP-40 was pruchased from PIERCE. Other chemicals and reagents used were reagent grade.. 14.
(15) Exogenous β- galactosidase Activity Assay For the exogenous β-galactosidase Activity assay, HeLa cells were cultured in the 35-mm culture dishes to about 70% confluence. The expression plasmid pcDNA 3/Mst3WT or pcDNA 3/Mst3KR (0.7 µg each) was then co-transfected with pSV-Laz vector (0.7 µg) into the cells by using lipofectamine (GIBCO). Twenty-four hours following transfection, cells were lysed by incubating with 200 µL cell lysis buffer (20 mM PIPES, PH 7.2, 100 mM NaCl, 1 mM EDTA, 0.1% CHAPS and 10% sucrose) at room temperature for 15 min. The β-galactosidase activity assay was carried out by mixing 100 µL cell lysate with 200 µL PM2 buffer (0.1 M Na3PO4, 1 mM MgSO4, 0.2 mM MnSO4), and 200 µL O-nitrophenyl-β-D-galactopyranoside (ONPG; 4 mM/mL). Reaction mixture was then incubated at 37 ℃ for 6 h or until a yellow color developed. Stop the reaction by adding 0.5 mL 1M Tris-HCl, PH 7.0. The activity of β-galactosidase of each sample was determined by meaning the absorbance at 420nm.. TUNEL (TdT-Utp Nick End Labeling) Assay DNA fragmentation induced by Mst3 was determined using TdT-FragEL kit (Oncogene) according to manufacture’s instruction. HeLa cells were cultured on the cover-glass slide in a 35-mm cultural dish to about 70% confluence before subjecting to transient transfection and transfected with pcDNA3-Mst3 or its mutants. After recovering for 16 h, cells were washed twice with TBS buffer ( 20 mM Tris-HCl, PH 7.6, 140 mM NaCl) and fixed with 4% formaldehyde in 1× TBS at 4 ℃ for 30 min. After fixation, cells were rinsed once with TBS and permeabilized by adding 2 mL TBST solution (20 mM Tris-HCl, PH 7.6, 140 mM NaCl and 0.05% Tween 20) on the top of cells at room temperature for 5 min. After 15.
(16) permeabilization, 100 µL 3% H2O2/methanol were added and incubated at room temperature for 5 min to deprive the endogenous peroxidase. Cells were washed twice with TBS and incubated with 100 µL 1× TdT equilibration Buffer (1M sodium cacodylate, 0.15M Tris, 1.5 mg/ml BSA, 3.75 mM CoCl2, pH 6.6) and incubated at room temperature for 20 min. Following incubation, carefully remove the TdT equilibration buffer from the glass slide, and immediately applied 60 µL TdT- labeling reaction mixture (3 vials containing a mixture of labeled and unlabeled deoxynucleotides at a ratio optimized for DNA Fragment End Labeling with TdT) onto the cells. The labeling reaction was performed by placing the slide in a 37 ℃ incubator for 90 min. After incubation, the reaction was terminated by adding 100 µL stop buffer (0.5 M EDTA, pH 8.0) at room temperature for 5 min. Rinse the entire specimen once with TBS and incubate with 100 µL blocking buffer (4 % BSA in PBS) at room temperature for 10 min. Remove the blocking buffer, immediately add 100 µL of 1× peroxidase-conjugated streptavidin solution to the cells at room temperature for 30 min. The HRP activity was then detected by incubating 100 µL DAB solution at room temperature for 10 min. After reaction, cells were used with demonized H2O once, followed by adding 100 µL methyl green (0.3 % methyl green) counter stain solution at room temperature for 3 min. Cells were then washed with 100% ethanol and dried the specimen under the room temperature. Finally, observe the cells under the light microscopy and count the number of positively stained cells.. Cell Transfection The plasmid transfection into cells was performed using lipofectamine reagent (GIBCO). Briefly, HeLa cells (2.5×105/well) were grown in a 6-well culture plate 16.
(17) at 37 ℃ to about 70% confluence. The transfection mixture was freshly prepared by mixing solution I (1 µg DNA in 100 µL Opti-MEM) solution II (4 µL lipofectamine in 100 µL Opti-MEM). The resulting mixture was then incubated at room temperature for 30-45 min. After incubation, add 800 µL Opti-MEM into the transfection mixture and mix well. Remove the old culture medium from each well and smoothly lay 1 mL transfection mixture on the top of the cells. The incubation was carried out at 37 ℃ for 5 hr in a CO2 incubator. Following incubation, replace the transfection mixture with 2 mL fresh DMEM containing 10% FBS. The cells were then recovered at 37 ℃ for about 14 h, before the subsequent treatments. The translocation. of. mitochondrial. proteins. and. were. performed,. using. immunofluorescent staining. The z-VAD-fmk (50 µM), JNK inhibitor (20 µM) and ERK inhibitor were used to treat the cells. The treatment was processed at 37 ℃ incubator for 24 hr.. Immunofluorescent Staining The translocation of various mitochondrial proteins in cells was monitored by immunofluorescent staining HeLa cells (2.5×105/well) were cultured in a 35-mm culture dish containing a piece of 2×2cm cover-glass to about 70-80% confluence. Cells were then transfected with pEGFP-Mst3WT (1 µg) or pEGFP-Mst3KR (1 µg). After transfection, Cells were recovered in the DMEM containing 10% FBS for 14 hours. The cells on the cover slip were washed once with 1× PBS and fixed with 4% paraformaldehy in PBS containing 50mM NaHCO3 at room temperature for 20 min. Cells were then blocked by 5% FBS at room temperature for 1 h, followed by incubating with first antibody (1: 200 dilution for AIF antibody, 1:100 dilution for EndoG and cytochrome C antibodies) at room temperature for 1-3 h in a humidified chamber. After incubation, cells were washed three times with PBST 17.
(18) (20 mM phosphate, PH 7.6, 140 mM NaCl and 0.05 % Tween 20) followed by incubating with the secondary antibody (1:200 dilution for Rhodamine-conjugated secondary antibody and 1:400 dilution for FITC-conjugated secondary antibody) at room temperature for 30 min. After incubation, cells were the nucleus of further stained with 200 ng/mL 4’6’-diamidino-2-phenylindole (DAPI) in the dark, at room temperature for 15-30 min. After staining, cells were washed three times with PBST. The fluorescent images were acquired by the confocal laser scanning microscope ( FV500, OLYMPUS).. Isolation of Cytoplasmic and Nuclear fractions HeLa cells were cultured in a 100-mm cultural dish to about 70-80% confluence. Cells were then harvested by scrapping and collected via centrifugation. Cells were re-suspended in 200 µL buffer I (10 mM Tris-HCl, pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA and 1 mM DTT) and incubated on ice for 15 min. After incubation, 10% NP40 (12.5 µL) was added into the cell suspension and vigorously vortexed for 10 second under the room temperature. Centrifuge the tubes at 13200 rpm for 30 second to collect the nuclei. Transfer the upper cytoplasmic fraction to a new 1.5 mL microfuge tube. To the nuclei pellet buffer II (20 mM Tris-HCl, pH 7.9, 40 mM NaCl, 1 mM EDTA, 1 mM EGTA and 1 mM DTT) were added, followed by a vigorous shaking in the cold for 15 min. After incubation, mixture was centrifuged at 13200 rpm in the cold for 30 second. Transfer the upper nuclear fraction to a new 1.5 mL tube and stored at –80℃ or subjected to experiment directly.. Isolation of Mitochondrial proteins The isolation of mitochondrial proteins was performed using Cytochrome C 18.
(19) releasing Apoptosis Assay kit following manufacture’s protocol. Briefly, cells (5×107) were collected by scrapping in 1 mL PBS, followed by centrifugation at 1400 rpm for 5 min at 4 ℃. The pellet of cells were re-suspended 1 mL 1× Cytosol Extraction buffer mix containing 1mM DTT and protease inhibitor and incubated on ice for 10 min. Transfer homogenate to a 1.5 mL microfuge tube and centrifuge at 2000 rpm at 4 ℃ for 10 min. Transfer supernatant to a 1.5 mL microfuge tube and centrifuge at 13200 rpm at 4 ℃ for 30 min. Collect the supernatant as cytoplasmic fraction. Re-suspend the pellet in 100 µL of 1× Mitochondria Extraction Buffer containing DTT and protease inhibitor. Vortex for 10 second and save as mitochondria proteins.. Western Blotting HeLa cells or pcDNA3.0 and pcDNA/Mst3KR stably transfected cells were cultured in a 100-mm cultural dish to about 70-80% confluence. The cell lysate was prepared by incubating cells with cell lysis buffer (50 mM Tris pH 7.4, 1 mM NaCl, 1 mM EGTA, 1mM EDTA, 1% Triton X-100, 1% Sodium deoxycholate, 0.1% SDS, 1 mM PMSF, 10 µg/ml Aprotinin and 10 µg/ml Leupeptin) on ice for 30 min. Total protein (60 µg) of each cell lysate was loaded onto a 12% SDS-polyacrylamide electrophoresis gel (SDS-PAGE). After electrophoresis at 150 Volt for 1.5 h, the separated protein bands were transferred to a polyvinylidine difluroide (PVDF) membrane on a BIO-LAB Mini-Trans Blot Electrophoretic Transfer Cell soaked in a transfer buffer (12.5 mM Tris, 96 mM glycin, 20% methanl and 0.1% SDS). After transblotting, PVDF membrane was blocked with 5% skim milk in PBST at room temperature for 1-2 h. The membrane was then incubated with first antibody (1: 5000 dilution for β-actin, 1: 1000 dilution for 19.
(20) Mst3 antibody, 1: 2000 dilution for AIF antibody, 1: 500 dilution for cytochrome C antibody, 1: 500 dilution for EndoG antibody, 1:1000 dilution for Bcl2 antibody, 1:1000 dilution for Bax antibody and 1: 500 dilution for Bcl-XL) at room temperature for 1h or at 4℃ for overnight. After incubation, the membrane was washed three times with PBST, followed by incubating with secondary antibody (1: 1000 dilution for host rabbit, mouse and goat) at room temperature for 1 h. After washing three times with PBST and once with PBS, 1 mL ECL was spreaded on the PVDF membrane for chemiluminescent reaction. The specific protein bands were visualized by exposing on a X-ray film (Hyperfilm). Protein concentration for SDS-PAGE was determined by Braford reagent (Bio-Rad).. 20.
(21) Results Mst3 Induced Apoptosis of HeLa cells It has been demonstrated that Mst3 could induce the major characteristics apoptosis in HeLa cells (Huang et al., 2002). Overexpression of Mst3 induced DNA fragmentation, a major characteristic of cell apoptosis of HeLa cells. The TdT-FragEL KIT was further used to determine whether Mst3-induced DNA fragmentation in HeLa cells. The Terminal deoxynucleotidyl transferase (TdT) catalyzes the labeling of exposed 3’-OH ends due to DNA fragmentation with biotin-conjugated nucleotide. The HRP-conjugated streptavidin was then used to recognize the biotin- conjugated newly exposed 3’-end. Diaminobenzidine (DAB) reacts with the labeled sample to generate the insoluble dye deposit (brownish red) at the site of DNA fragmentation. Counterstaining of cells with methyl green could aid in the morphological evaluation and characterization of normal and apoptotic cells. The result shows that DNA fragmentation could be induced in HeLa cells by transiently transfected either with Mst3WT (Fig. 1D) or truncated Mst3. △314. (Fig. 1E).. In consist, HeLa cells either untreated, or transfected with vector alone or Mst3 KR showed no sign of DNA fragmentation (Figs. 1A, 1B and 1C).. Mst3 Induces a Caspase-independent Apotosis Caspases are the central executioners of most classical apoptotic pathways and are suggested to involve in Mst3-mediated apoptosis in Jurkat cells (Huang et al., 2002). We want to examine whether Mst3-induced HeLa cell apoptosis of HeLa cells is a caspase–dependent process. Accordingly, an exogenous β-galactosidase assay was performed by co-transfecting pSV-LacZ plasmid with Mst3WT or its muntants into the HeLa cells. Six hours after transfection, a universal-caspase 21.
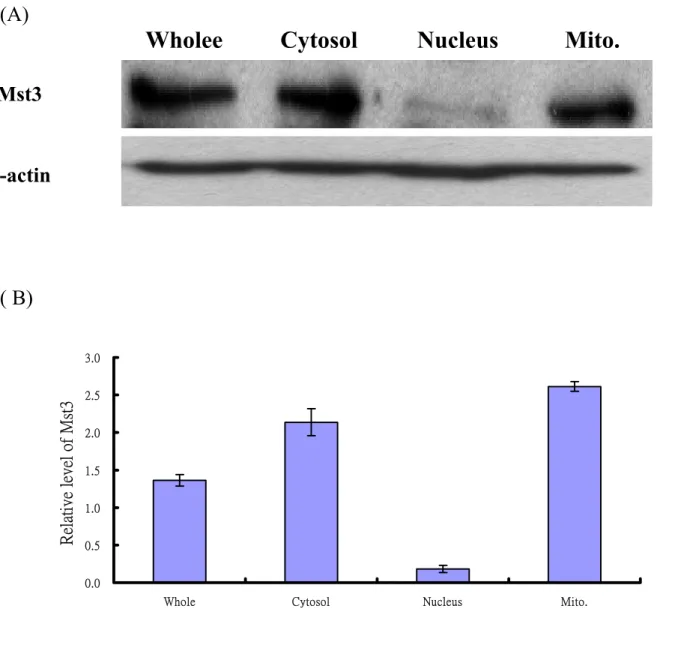
(22) inhibitor zVAD-fmk (50 µM) was added into in the culture medium. Notably, Mst3WT-induced cell death could not be inhibited by 50 µM zVAD-fmk (Fig. 2). Both pcDNA3.0 vector and Mst3KR showed no significant effect on HeLa cells and were not affected by caspase inhibitor. The result suggests that Mst3 may induce apoptosis of HeLa cells in a caspase-independent manner.. Effect of JNK nor p38/MAPK inhibitor in Mst3-induced apoptosis Mst1 or Mst2 has been shown to induce apoptosis through the JNK/SAPK and p38/MAPK pathways (Lin et al., 2001). Early works in our laboratory also indicated that Mst4-induced apoptosis was a JNK-dependent process. Although Mst3 was shown to induce cell apoptosis through a pathway independent of any known MAPK pathway (Huang et al., 2002), we want to further confirm this observation using exogenous β-galactosidase assay in the presence of. SP600125,. an inhibitor for JNK or SB220025, an inhibitor for p38/MAPK. As expected Mst3-induced apoptosis of HeLa cells was not affected by JNK inhibitor, SP600125 (Fig. 3) and p38/MAPK inhibitor, SB220025 (Fig. 4), confirming the previous observations (Huang et al., 2002).. Localization of endogenous Mst3 in HeLa Cells Pervious study reveals that full length Mst3 was mainly present in the cytoplasm, whereas Mst3WT∆314 accumalceted in the nucleus. The nucleus localization of Mst3 WT∆314 was suggested to play a role in promoting apoptosis of cells. However, this result can not explain the observation that overexpression of full-length Mst3WT can also induce apoptosis (Huang et al., 2002). Interestingly, Mst4, a closely related isoform of Mst3, was shown to distribute at Golgi apparatus, especially at cis-Golgi network. This observation promotes us to check the 22.
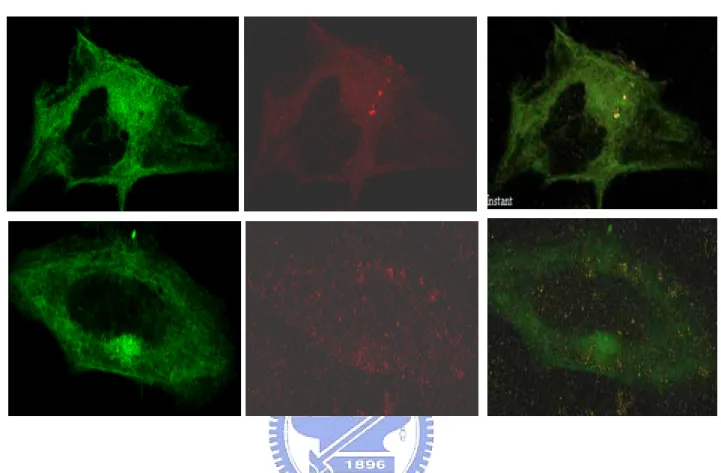
(23) subcellular distribution of Mst3 in HeLa cells by western blotting. The cytoplasmic, nuclear and mitochondrial fractions were first prepared as indicated in Materials and Methods. The existence of endogenous Mst3 in various fractions was determined by Western blotting. To our surprise, we found that endogenous Mst3 was not just only present in the cytoplasm but also existed in the mitochondria (Fig. 5). Consistent with previous observation, only little Mst3 is present in the nucleus. This observation was further demonstrated by monitoring the fluorescent images of mitochondria, mitochondrial marker (Molecular Probe) in EGFP-Mst3WT and EGFP-Mst3KR transiently transfected HeLa cells. Fig. 6 shows that EGFP-Mst3 WT or EGFP-Mst3KR (green) co-localizes with the mitotracker (red), indicating that Mst3 is present in mitochondria. The reason for the presence of Mst3 in mitochondria is, however, remain elucidated.. Locatization of apoptotic factors in HeLa Cells Several lines of evidence have shown that many apoptotic factors, including cytochrome C, apoptosis inducing factor (AIF) and endonuclease G (Endo G), are present in the mitochondria. Cytochrome C was in the intermembrane space (IMS) (Grup et al., 2003; Yu et al., 2002; Ohsato et al., 2002). EndoG and AIF were also confined to the mitochondrion but the exact location is not clear so far. Thus, it will be very interesting to know if any one of these apoptotic factors can associate with Mst3, which is just indicated to exist in mitochondria. The existence of apoptotic factors in mitochondria was first demonstrated by double immunofluorescent staining. Mitochondria were stained with a specific fluorescent conjugated mitochondrial marker, mitotracker red (Molecular Probe). Fluorencent images of apoptotic factor and mitotracker were observed under a confocal laser scanning microscope (Fig. 7). The result clearly shown that the apoptotic factor proteins 23.
(24) studied in this work are resided in the mitochondria. We next try to find out whether Mst3 associates with any of the above apoptotic factors. HeLa cells were transiently tranfected with EGFP-Mst3WT or EGFP-Mst3KR plasmid prior to the immunofluorescent staining of apoptotic factors. Sixteen hours after transfection, cells were fixed and detected by the antibody specifically against one of above apoptotic factors. The localization of apoptotic factors was visualized using Rhodamine-conjugated secondary antibody. Interestingly, we found that the fluorescent images of cytochrome C and EGFP-Mst3 and EGFP-Mst3KR were colocalize (Fig. 8), presumably in the mitochondria. The fluorescent image of AIF was also observed to overlap with that of EGFP-Mst3KR (Fig. 9). EndoG, on the contrary, did not show any sign of overlapping with EGFP-Mst3 (Fig. 10).. Effect of exogenously expressed Mst3WT and Mst3KR on the Bcl-2 protein level Members of Bcl-2 family play important roles in the regulation of cell apoptosis. These proteins can be classified into two subfamilies based on their role in the regulation of apoptosis. The anti-apoptotic Bcl-2 subfamily, including Bcl-2, Bcl-XL and Bcl-W, promotes cell survival; the pro-apoptotic subfamily, including, Bax, Bid and Bak, induces cell death. Among Bcl-2 homologs, Bcl-2, Bcl-XL and Bax represent the most well characterized members. Bcl-2 can promote cell survival against various cytotoxic insults (Biose et al., 1995) and Bcl-XL could inhibit apoptosis induced by free radical, UV light and chemicals. Bax, on the other hand, can cause the release of cytochrome C from mitochondria and activates caspase 3-dependent cell death. Hence, it is worth to know the influence of Mst3 in the expression of Bcl-2-like proteins in HeLa cells. HeLa cells transfected with pcDNA-Mst3WT and pcDNA-Mst3KR were used as the models to find out the effects of Mst3WT and Mst3KR to the protein level of Bcl-2, Bcl-XL, and Bax. The 24.
(25) result reveals that the protein level of Bax and Bcl-XL was unchanged. The protein level of Bcl-2 was reduced with the overexpression of Mst3WT (Fig. 11). In the presence of Mst3KR, however, Bcl-2 level was almost unchanged. This result suggests that Mst3-induced cell death may be proceed via changing the availability of anti-apoptotic Bcl-2.. Staurosporine induces nuclear localization of Mst3 Earlier work has shown that the association of Mst3 with cytochrome C may involve the regulation of cell apoptosis. Interestingly, the apoptosis induced by some anti-cancer drugs including, staurosporine, etopside and camptothecin involved in the interference of mitochondrial functions. For example, release of cytochrome C can be induced by etoposide (Yang et al., 1997; Sun et al., 1999). Staurosporine can induce the proteolytic cleavage of Mst3 and hence the apoptotic cell death of Jurkat cells (Huang et al., 2002). The effect of staurosporine on Mst3 in HeLa cells, however, has not been studied. The HeLa cells were treated with 0.5 µM staurosporine at 37 ℃ for 3 h. Subsequently, cells were harvested and fractionated to prepare cytoplasmic, nuclear and mitochondrial fractions. Cells treated with 0.05 % DMSO was used as control. Surprisingly, we found that staurosporine could induce a nuclear translocation of full-length Mst3 in HeLa cells. The subcellular redistribution of Mst3 and other apoptosis regulating factors, such as AIF, EndoG and Bcl-2 was studied. The nuclear localization of AIF also induced by the treatment of 0.5 µM stauroaporine (Fig. 14).The protein level of Bcl-2 in nucleus was also increased with the stimulation of staurosporine.. 25.
(26) Discussion Pervious studies have revealed that overexpression of Mst3 might induce apoptosis of HeLa and HEK293 cells (Huang et al., 2002). To further confirm the role of Mst3 in apoptosis DNA fragmentation, one of the hallmarks of apoptosis, the cell singles were analyzed by TUNEL assay. I found that Mst3 could induce DNA fragmentation in HeLa cells as demonstrated by the brownish staining in nuclei of Mst3WT and Mst3 WT. Δ314. transfected cells (Fig 1).. GCK II family protein kinases, such as Mst1 and Mst2, have been shown to activate either JNK/SPAK or p38/MAPK signaling pathway upon overexpression (Dan et al., 2001; Widmann et al., 1999). Mst1, sharing 56% sequence homologous with Mst3, is involved in the JNK and p38 pathways (Taylor et al., 1996). It was also found that Mst4 could induce apoptosis of MCF cells via a JNK-dependent pathway. Notably, Mst3 was suggested to induce an unique signaling pathway without the involvement of any known MAPK cascades. This work further confirmed this observation by finding that Mst3-indued HeLa cell apoptosis was not affected by JNK or p38 inhibitors. Caspases are the central executives of most classical apoptosis pathways. Although caspase3 is shown to cleave and activate various Ste20 group kinases including Mst1, Mst2 or PAK2 (Dan et al., 2001), Mst3-induced apoptosis of HeLa cells can not be inhibited by its inhibitor, Z-VAD-fmk. This result suggests that Mst3 may induce cell death via a caspase-independent pathway in HeLa cells (Daugas et al., 2000). Mitochondrion is one of the important organelles in cells playing multiple roles to supply energy needed for cellular functions as well as to mediate ageing, oxidative stress and cell apoptosis. Mitochondria mediate cell apoptosis via several ways, including activating caspase activators, changing redox status of the electron 26.
(27) transport chain, disrupting the mitochondria transmembrane potential, and cooperating with anti- and pro-apoptotic Bcl-2 family proteins (Adams and Cory, 1998). Release of cytochorome C from mitochondria induces the activity of caspase 9, and subsequently, activates caspase 3 and following apoptotic processes, termed caspase-dependent pathway. Numerous studies have demonstrated that apoptosis induced by some apoptotic stimuli cannot be inhibited by the pan-caspase. inhibitor,. Z-VAD-fmk,. suggesting. the. existence. of. caspase-independent apoptotic pathway (Wu, 2000; Okuno et al., 1998). Normally, theseapoptotic factor proteins are present in the mitochondrial intermembrane space. Once activated by any apoptosis stimulus, these apoptotic factors may translocate to the cytoplasm or the nucleus. Surprisingly, we found that cytochrome C is co-localized with mst3 in the mitochondria. This result indicated that Mst3 might induce apoptosis by altering the cellular localization of cytochrome C. The role of Mst3 in mitochondria is unknown so far. The existence of Mst3 may provide a new physiological aspect about Mst3 and showed the important for its role in regulating cell apoptosis. The translocation of these apoptotic factors are regulated by pro-survival Bcl-2 family members, such as Bcl-2 and BclXL, and pro-apoptotic Bcl-2 family members, such as Bax, Bad and Bim (Adams and Cory, 1998). The ratio of pro-survival and pro-apoptotic Bcl-2 family members determines the susceptibility of cells to a certain apoptotic signal. This postulation is partially supported by the finding that the protein level of Bcl-2 was reduced with overexpression of Mst3 in HeLa cells. The subcellular distribution of other apoptotic factors in response to the overexpression of Mst3 is unknown. Since Mst3-induced apoptosis is caspase-independent, the distribution of apoptotic factors altered by the activation of Mst3 is expected and will be studied in the future. 27.
(28) Staurosporine has been known to induce the proteolytic activation of Mst3 in Jurkat cells (Huang et al., 2002). In this we found that staurosporine may act differently by inducing the nuclear translocation of Mst3 and AIF. Bcl-2 in nucleus was also increased with the stimulation of staurosporine. There is mounting evidence shows that some forms of cell death can be mediated by calcium release from ER stores, perhaps accompanied by calcium entry from the extracellular milieu (Baffy et al., 1993; Pu and Chang, 2001; Abeele et al., 2002). This redistribution of calcium leads finally to a decisive induction of Pore. Transition. (PT). through. mitochondrial. calcium. overload.. Other. calcium-dependent mechanisms may or may not involve mitochondria, such as calpain activation. The interorganelle flow of calcium is facilitated by direct or indirect connections between mitochondria and the ER. Bcl-2 is known to regulate the redistribution of calcium, particularly reducing the amount of calcium releasable from the ER, thus preventing mitochondrial calcium overload and ensuring cell survival (Abeele et al., 2002). Bax, on the other hand, produce the opposite effect, making more calcium available for inducing PT in mitochondria (Nutt et al., 2002). Although regulation of Bcl-2 family involved many different cell death pathway, the results of this study indicates that Mst3 may play an essential role to regulate the release of caspase activator or other apoptotic factors with the cooperation of Bcl-2 family members.. 28.
(29) References Adams, J. M. and Cory, S. (1998) The Bcl-2 protein family: arbiters of cell survival. Science, 281: 1322-1326. Adams, J. M. and Cory, S. (2001) Life-or-death decisions by the Bcl-2 protein family. Trends Biochem Sci. 26: 61-66. Askenazi, A and Dixit, V. M. (1998) Death receptors: signaling and modulation. Science, 281: 1305-1308 Green, D. R. and Reed, J. C., (1998) Mitochondria and Apoptosis. Science, 28: 1308-1312. Huang, C. Y., Wu Y. M., Hsu, C. Y., Lee, W. S., Lai, M. D., Lu, T. J., Huang, C. L., Leu, T. H., Shih, H. M., Fang, H. I., Robinson, D. R., Kung, H. J. and Yuan, C.J., (2002) Caspase activation of mammalian sterile 20-like kinase 3 (Mst3). Nuclear translocation and induction of apoptosis. J. Biol. Chem. 277: 34367-34374. Blatt, N. B. and Glick, D. G. (2001) Signaling pathways and effector mechanism Pre-Programmed Cell Death. Bioorganic and Med. Chem. 9: 1371-1384. Cande, C., Cohen, I., Daugas, E., Ravagnan, L., Larochette, N., Zamzami, N. and Kroemer,. G.. (2002). Apoptosis-inducing. factor. (AIF):. a. novel. caxpase-independent death effector released from mitochondria. Biochimie. 84: 215-222. Chang, C. H. (2001) Exploring the induction of apoptosis mediated by the mammalian Ste20-like kinase, Mst3. Institute of Biological Science and Technology National Chiao Tung University In Partial Fulfillment of the Requirements for the Degree of Master of Science. Dan, I., Watanabe, M. N., and Kusumi, A. (2001) The Ste20 group kinases as regulators of MAP kinase cascades. Trend in Cell Biol. 11(5): 220-23. 29.
(30) Daugas, E., Nochy, D., Ravagnan, L., Loer, M., Susin, S. A., Zamzami, N. and Kroemer, G. (2000) Apoptosis-inducing factor (AIF): a ubiquitous mitochondrial oxidoreductase involved in apoptosis. FEBS Letters, 476:118-123. Enari, M., H. Sakahira, H. Yokoyama, K., Okawa, A., Iwamatsu and Nagata, S. (1998) A caspase-activated Dnase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature, 391: 43-50. Ernesto, C. and Maurizio, M. (1998) Calpain: A Protease in Search of a Function? Biochem. And Biophysical Research Communications, 247: 193-203. "Graves, J. D., Draves, K. E., Ambeose, D., Han, D.K.M., Wright, M., Chernoff, J., Clark, E.A and Krebs, E.G. (1998) Caspase-mediated activation and induction of apoptosis by the mammalian STE20-like kinase Mst1. EMBO J. 17: 2224-2234. Graves, J. D., Draves, K. E., Gotoh, Y., Krebs, E. G., Clark, E. A. (2001) Both Phosphorylation and Caspase-mediated Cleavage Contribute to Regulation of the Ste20-like Protein Kinase Mst1 during CD95/Fas-induced Apoptosis. J. Biol. Chem. 276: 14909-14915. Grup, M. V., Festjens, N., Loo G. V., Saelens X., Vandenabeele P. (2003) Mitochondrial intermembrane proteins in cell death. BBRC. 304: 487-497. Henartner, M.O. (2000) The biochemistry of apoptosis. Nature, 407: 770-776. Hou, Q., Cymbalyuk E., Hsu, S.C., Xu, M., Hsu, Y. T. (2003) Apoptosis modulatory activaties of transiently expressed Bcl2 : Roles in cytochrome C release and Bax regulation. Apoptosi, 8: 617-629. Huseyin Mehmet. (2000) Caspases find a new place to hide. Nature, 403: 29-30. Ippeita Dan, Noeinobu M., Watanabe and Akihiro Kusumi. (2001) The Ste20 group kinases as regulateors of MAP kinase cascades. TRENDS in Cell Biology. 11-5: 220-230.. 30.
(31) Jerry, M. Admas and Suzanne Cory. (1998) The Bcl-2Proyein Eamily: Arbiters of Cell Survival. Science, 28: 1322-1326. Kerr, J. F., Wyllie, A. H., Currie, A. R. (1972) Apoptosis: a basic biological phenomenon with wide-ranging implication in tissue kinetics. Br J Cancer. 26(4): 237-257. Kim, C. H. and King, T. E. (1983) A mitochondrial Protein Essentail for the Formation of rhe Cytochrome c1-c Complex. J.B.C. 258: 13543-13551. Lee, K. K., Murakawa, M., Nishida, E., Tsubuki, S., Kaawashima, S., Saakamaki, K., and Yonehara, S. (1998) Proteolytic activation of MST/krs, STE20-related protein kinase, by caspase during apoptosis. Oncogen, 16: 3029-3037. "Lee, K. K., Ohyama, T., Yajima, N., Tsubuki, S., Yonehara, S. (2001) MST, a physiological caspase substrate, highly sensitizes apoptosis both upstream and downstream of caspase activation. J. Biol. Chem. 276: 19276-19285. Lei, Kui and Davis, R. J. (2003) JNK phosphorylation of Bim-related members of the Bcl2 family induces Bax-inpendent apoptosis. PNAS. 100: 2432-2437. Li, L. Y., Luo, X. and Wang, X. (2001) Endonuclease G is an apoptotic Dnase when released from mitochondria. Nature, 412: 95-99. Lin, J. L., Chen, H. C., Fang, H. I., Robinson, D., Kung, H. J., Shih, H. M. (2001) MST4, a new Ste20-related kinase that mediates cell growth and transformation via modulating ERK pathway. Oncogene, 20: 6559-6569. Lockshin, R. A. and Zakeri, Z. (2002) Caspase-independent cell deaths. Curr. Opin Cell Biol. 14: 727-733. Loeffler, M., Daugas E, Susin, S. A., Zamzami, N., Brothers, G., Penninger, J. M., Kroemer, A. G. (2001) Dominant cell death inductuin by extramitochondrially targeted apoptosis-inducing factor. The FASEB Journal. 15: 758-767.. 31.
(32) Lorenzo, H. K. and Susin, S. A. (2004) Mitochondrial effector in caspase-independent cell death. FEBS. 557: 14-20. "Nicholson, D. W., and Thornberry, N. A. (1997) Caspases: killer proteases. Trends Biochem. Sci. 22: 299-306. MacKenzie, A. and LaCasse, E. (2000) Inhibition of IAP’s protection by Diablo/Smac: new therapeutic opportunities? Cell Death Differ. 7(10): 866-867. Nakagawa, T., Zhu, H., Morishima, N., Li, E., Xu, J., Yankner, B. A. and Yuan, J. (2000) Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-bata. Nature, 403: 98-103. Nicholson, D. W., and Thornberry, N. A. (1997) Caspases : killer proteases. Trends Biochem. Sci. 22: 299-306. Nylandsted, J., Rohde, M., Brand, K., Bastholm, L., Elling, F and Jaattela, M. (2000) Selective depletion of heat shock protein 70 (HSP70) activates a turmor-specific death program that is independent of caspases and bypasses Bcl-2. PNAS. 97: 7871-7876. "Okuno, S., Shimizu, S., Ito, T., Nomurea, M., Hamada, E., Tsujimoto, Y., and Matsuda, H. (1998) Bcl-2 Prevents Caspase-independent Cell Death. J. Biol. Chem. 273: 34272-34277. Penninger, J. M. and Kroemer, G. (2003) Mitochondria, AIF and caspaes-rivaling for cell death execution. Nat Cell Biol. 5: 97-99. Reszka, A. A., Halasy-Nagy, J. M., Masarachia, P. J. and Rodan, G. A. (1999) Bisphosphonates act directly on the osteoclast to induce caspase cleavage of Mst1 kinase during apoptosis. J. Biol. Chem. 274: 34967-34973. Scarlett, J. L., Sheard, P. W., Hughes G., Ledgerwood, E. C., Ku, H. H., Murphy, M. P.. (2000). Changes. in. mitochondrial. membrane. potential. during. Staurosporine-induced apoptosis in Jurkat cells. FEBS Letters, 475: 267-272. 32.
(33) Scorrano, L, Oakes, S. A., Opferman, J. T., Cheng, E. H., Sorcinelli, M. D., Pozzan T., Korsmeyer S. J. (2003) Bax and Bak Regulation of Endoplasmic Reticulum Ca2+ : A Control Point for Apoptosis. Scienc, 300: 135-139. Springs, S. L., Diavolitsis, V. M., Goodhouse, J., McLendon, G. L. (2002) The Kinetics of Translocation of Smac/DIABLO from the Mitochondria to the Cytosol in HeLa Cells. J.B.C. 29: 45715-45718. Stennicke, H. R. and Deveraux, G. S. (1998) Properties of the caspases. Biochim Biophys Acta, 1387: 13-17. Susin, S. A., Lorenzo, H. K., Zamzami, N., Marzo, I., Snow, B. E., Brothers, G. M., Mangion, J., Jacotot, E., Costantini, P., Loeffler, M., Larochette, N., Goodlett, D. R., Siderovski, D. P., Penninger, J. M. and Kroemer, G. (1999) Molecular characterization of mitochondrial apoptosis-inducing factor. Nature, 397: 441-446. Tang Damu, Lahti J. M., Kidd V. J. (2000) Casapse-8 Activation and Bid Cleavage Controbute to MCF7 Cellular Execution in a Caspase-3-dependent Manner during Staurosporine-mediated Apoptosis. J.B.C. 275: 9303-9307. Thornberry, N. A. and Lazebnik, Y. (1998) Caspase : Enemies Within. Science, 281: 1312-1316. Tyalor, L. K., Wang, H. C. R. and Erikson, R. L. (1996) Newly identified stress-responsive protein kinases, Krs-1 and Krs-2. Proc. Natl. Acad. Sci. U.S.A. 93: 10099-10104. Wu, M., Xu, L. G., Li, X., Zhai, Z., and Shu, H. B. (2002) AMID, an Apoptosis-inducing Factor-homologous Mitochondrion associated Protein, Induces Caspase-independent Apoptosis. J. Biol. Chem. 277: 25617-25623. Xiang, J., Chao, D. T. and Korsmeyer, S. J. (1996) Bax-induced cell death may not require interleukin 1β-converting enzyme-like protease. Pro. Natl. Acad. Sci. 33.
(34) USA, 93:14559-14563. "Xue, L., Fletcher, G. C. and Tolkovshy, A. M. (2001) Mitochondria are selectively eliminated from eukaryotic cells after blockade of caspase during apoptosis. Curr. Biol. 11: 361-365. Zimmermann, K. C., Bonzon, C. and Green, D. R. (2001), The machinery of programmed cell death. Pharmcol. & Thre. 92: 57-70. Lee, W. S. (2003) The role of caspases in Mst3-induced cell Apoptosis. 國立交通 大學 碩士論文 Du, J. L. (2003) Mechanisms of Germanium Oxide-induced Apoptosis in HL-60 Cells. 國立清華大學 碩士論文 Chen, P. H. (2003) The research on the biological function of a novel human Ste20-like protein kinase Mst4. 國立交通大學 碩士論文. 34.
(35) (A) Non-transfection. (B) pcDNA3.0. (C) pcDNA/Mst3KR. (D) pcDNA/Mst3WT. (E) PcDNA/Mst3WT∆314. Fig. 1 The TUNEL assay of HeLa cells. HeLa cells were transfected without (A) or with pcDNA3.0, (B) pcDNA/Mst3KR, (C) pcDNA-Mst3WT, (D) and PcDNA/Mst3WT∆ 314 (E). Cells were recovered for 18 h prior to the TUNEL assay. Cells transfected with pcDNA3.0 and pcDNA/Mst3KR were used as negative controls. Cells underwent DNA fragmentation were stained brownish red in the nucleus, whereas the viable cells exhibited green in the picture.. 35.
(36) 1.2. Relative Cell Vizbility. 1. 0.8. 0.6. 0.4. 0.2. 0. Relative activits. pcDNA 3.0. pcDNA+ZVAD-fmk. Mst3WT. Mst3WT+ZVAD-fmk. Mst3KR. Mst3KR+ZVAD-fmk. 1. 1. 0.49. 0.47. 0.88. 0.93. Fig. 2 Effect of the pan-inhibitor Z-VAD-fmk on Mst3 induced apoptosis. HeLa cells were co-transfected with pSV-LacZ and pcDNA3/Mst3WTor pcDNA3/Mst3KRplasmids. Six hours after transfection , cells were left untreated or treated with Z-VAD-fmk (50 µM) at 37℃ for 24 h after treatment, cells harvested and assay for β-galactosidase activity. Results shown are mean ± S.E of three independent experiments. The β-galactosidase activity of pcDNA3.0-transfected cells was used as 1.0.. 36.
(37) 1.2. Relative Cell Vizbility. 1. 0.8. 0.6. 0.4. 0.2. 0. Relative activits. pcDNA 3.0. pcDNA+S P60 0125. Mst3WT. Mst3WT+S P 600125. Mst3KR. Mst3KR+S P6 00125. 1. 1. 0.53. 0.52. 0.94. 0.91. Fig. 3 Effect of the JNK inhibitor (SP600125) on Mst3 induced apoptosis. HeLa cells were co-transfected with pSV-LacZ and pcDNA3/Mst3WTor pcDNA3/Mst3KR vectors. Six hours after transfection, cells were left untreated or treated with JNK inhibitor SP600125 (0.5 µM) at 37℃ for 24 h. After incubation, cells were harvested and assay for β-galactosidase activity. Results shown are mean ± S.D of three independent experiments. The β-galactosidase activity of pcDNA3.0-transfected cells was used as 1.0.. 37.
(38) 1.2. Relative Cell Vizbility. 1. 0.8. 0.6. 0.4. 0.2. 0. Relative activits. pcDNA 3.0. pcDNA+S B22 0025. Mst3WT. Mst3WT+S B 220025. Mst3KR. Mst3KR+S B2 20025. 1. 1. 0.61. 0.59. 0.83. 0.87. Fig. 4 Effect of the p38 MAPK inhibitor (SB220025) on Mst3 induced apoptosis. HeLa cells were co-transfected with pSV-LacZ and pcDNA3/Mst3WTor pcDNA3/Mst3KR vectors. Six hours after transfection, cells were left untreated or treated with p38 MAPK inhibitor SB220025 (0.5 µM) at 37℃ for 24 h. Cells were then harvested and assay for β-galactosidase activity. Results shown are mean ± S.D of three independent experiments. The β-galactosidase activity of pcDNA3.0-transfected cells was used as 1.0.. 38.
(39) (A). Wholee. Cytosol. Nucleus. Mito.. Mst3. β-actin. ( B). Relative level of Mst3. 3.0 2.5 2.0 1.5 1.0 0.5 0.0 Whole. Cytosol. Nucleus. Mito.. Fig. 5 Localization of endogenous Mst3 in HeLa cell. (A) Western blot analysis of the endogenous Mst3 in HeLa cells, Cells were grown to about 80-90% confluences and then the whole cell lysate, cytoplasmic, nuclear and mitochondrial fractions were isolated as described in Materials and Methods. Sample (60 µg each) was loaded and separated on a 12% SDS-PAGE. β-actin was used as internal control. Mst3 is mainly present in cytoplasm and mitochondria. (B) The relative amount of Mst3 of different subcellular fractions in HeLa cell. The density of β-actin bend in each lane was viewed as 1 to normalize the density of Mst3 in each fraction. 39.
(40) EGFP. Mitotracker. Merge. (A) EGFP. (B) EGFPMst3WT. (C) EGFPMst3KR. Fig. 6 Localization of EGFP-Mst3 in mitochondria. HeLa cells were transiently transfected with (A) EGFP vector only, (B) EGFP-Mst3WT or (C) EGFP-Mst3KR. Sixteen hours after transfection, cells were fixed and stained with the mitochondrial marker, mitotracker (Molecular Probe). Fluorescent images were monitored under a confocal laser scanning microscope (FV500, OLYMPUS). The yellow color indicates the co-localization of EGFP- Mst3WT (green) and mitotracker (red).. 40.
(41) FITC. Mitotracker. Merge. (A) cytochrome C. (B) AIF. (C) EndoG. Fig. 7 Localization of cytochrome C, AIF and Endo G in mitochondria. The apoptotic factors in HeLa cells were immunofluorescent stained using mouse-anti cytochrome C, mouse-anti AIF and rabbit-anti EndoG antibodies, followed by staining with a FITC-conjugated secondary antibody ageist mouse or rabbit IgG. Mitochondria in HeLa cells were stained with a specific marker, mitotracker red (Molecular Probe). Fluorescent images were observed under a confocal laser scanning microscope (FV500, OLYMPUS). The co-localization of apoptotic factors (green) and mitochondria (red) was visualized as yellow color in the merged images. 41.
(42) EGFP. Cytochrome C. Merge. (A) EGFPMst3WT. (B) EGFPMst3KR. Fig. 8 Co-localization of Mst3 and cytochrome C in mitochondria. HeLa cells were transiently transfected with EGFP-Mst3WT (A) or EGFP-Mst3KR (B). Sixteen hours after transfection, immunofluorescent staining of cytochrome C was performed using mouse anti cytochrome C antibody and visualized by a Rhodamine-conjugated secondary antibody anti-mouse IgG. Fluorescent images were observed under a confocal laser scanning microscope (FV500, OLYMPUS). The co-localization of EGFP-Mst3WT or EGFP-Mst3KR (green color) and cytochrome C (red) was demonstrated as the yellow color in the merged images.. 42.
(43) EGFP. AIF. Merge. (A) EGFPMst3WT. (B) EGFPMst3KR. Fig. 9 Immunofluorescent Staining in HeLa cells. HeLa cells were transiently transfected with EGFP-Mst3WT (A) or EGFP-Mst3KR (B). Sixteen hours after transfection, AIF was recognized mouse using anti-AIF antibody and visualized by a Rhodamine-conjugated secondary anti-mouse antibody. Fluorescent images were observed under a confocal laser scanning microscope (FV500, OLYMPUS). The co-localization of EGFP-Mst3KR (green) and AIF (red) was demonstrated as the yellow color in the merged images.. 43.
(44) EGFP. Endo G. Merge. (A) EGFPMst3WT. (B) EGFPMst3KR. Fig. 10 Immunofluorescent Staining in HeLa cells. HeLa cells were transiently transfected with EGFP-Mst3WT (A) or EGFP-Mst3KR (B). Sixteen hours after transfection, EndoG was recognized using rabbit anti-Endo G antibody and visualized by a Rhodamine-conjugated secondary anti-rabbit antibody. Fluorescent images were observed under a confocal laser scanning microscope (FV500, OLYMPUS).. 44.
(45) Control. pcDNAMst3WT. pcDNAMst3KR. Bax. Bcl-2. Bcl-xL. actin. Fig. 11 Expression level of Bcl-2 in the presence of Mst3WT and Mst3KR. Whole cell extracts (50 µg) from pcDAN-Mst3WT and pcDAN-Mst3KR-transfected HeLa cells were separated on a 12% SDS-PAGE, followed by probing the protein bends of Bax, Bcl-2 and Bcl-xL using specific antibodies. Actin was used as internal control.. 45.
(46) (A). Whole. Cytosol. Nucleus. Mito.. Bcl-2. β-actin. (B). Relative level of Bcl-2. 1.5. 1.0. 0.5. 0.0 Whole. Fig. 12. Cytosol. Nucleus. Mito.. Subcellular distribution of Bcl-2 in HeLa cells. Endogenous Bcl-2 was. determined in whole cell extract, cytosolic fraction, nuclear fraction and mitochondrial fractions by Western blotting. Samples (60 µg each) various fractions samples were separated on a 12% SDS-PAGE, followed by the western blotting analysis. (B) The relative amount of Bcl-2 of different cell fractions of HeLa cell. The density of β-actin in each lane was viewed as 1 to normalize the density of Mst3 in each fraction.. 46.
(47) 120. C e ll s u rv iv a l (% ). 100. 80. 60. 40. 20. 0. DMSO. Staurosporine. 0. 1. 2. 3. 4. 5. 6. 7. 95.632. 93.408. 86.18. 85.07. 82.29. 79.51. 78.39. 78.39. 100. 89.52. 77.84. 73.39. 50.59. 44.48. 30.02. 24.46. Time. Fig. 13 Time-dependent effect of sturosporine on HeLa cells. HeLa cells treated with DMSO or 0.5 µM staurosporine at 37℃ for different time. The cells were harvested and assay for MTT activity Assy. Results shown are mean ± S.D of three independent experiments.. 47.
(48) staurosporine. DMSO. Whole. Cytosol Nucleus. Mito.. Whole Cytosol. Nucleus. Mito.. Mst3 AIF EndoG Bcl-2 β-actin. Fig. 14 The effect of staurosporine on the subcellular dietribution of Mst3 and apoptotic regulation factors in HeLa cell. HeLa cells treated with DMSO or 0.5 µM staurosporine at 37℃ for 3 hours were fractionated to isolated the whole protein, cytoplasmic, nuclear and mitochondrial fractions Various sample (40 µg) were separated on a 12% SDS-PAGE. The resulting protein bands were then transferred to PVDF membrane and subjected to Western blotting. Antibodies against Mst3, AIF, EndoG, Bcl-2 and β-actin were used to detected the coverpouding proteins on the PVDF membr 48.
(49) Apoptosis pathway. 49.
(50) Proposed pathway in Jurkat cells. 50.
(51) Proposed pathway in HeLa cells. 51.
(52) Appendix I Nucleotide and amino acid sequence of Mst3.. The putative start codon is indicated in bold. The kinase domain is underlined. Numbering of nucleotides and amino acids is indicated on the right.. 52.
(53) Appendix II. The plasmid map of pEGFP-C2 vector. Restriction Map and Multiple Cloning Site (MCS) of pEGFP-C2. (Unique restriction sites are in bold). The EagI site is not unique. The XbaI site cannot be used for fusions since it contains an in-frame stop codon. The XbaI and Bcl I sites(*) are methylated in the DNA provided by CLONTECH. If you wish to digest the vector with these enzymes, you will need to transform the vector into a dam- host and make fresh DNA. 53.
(54) Appendix III. The plasmid map of pcDNA3 vector. CTC TCT GGC TAA CTA GAG AAC CCA CTG CTT ACT GGC TTA TCG AAA TTA HA tag --> ATA CGA CTC ACT ATA GGG AGA CCC AAG CTT GGT ACC ATG GCT TAC CCT HindIII. KpnI NcoI. TAT GAT GTG CCA GAT TAT GCC TCC CGG GAT CCA CTA GTA ACG GCC GCC SmaI BamHI AGT GTG CTG GAA TTC TGC AGA TAT CCA TCA CAC TGG CGG CCG CTC GAG EcoRI. EcoRV. NotI. XhoI. CAT GCA TCT AGA GGG CCC TAT TCT ATA GTG TCA CCT AAA TGC TAG AGC XbaI. ApaI. TCG CTG ATC AGC CTC GAC TGT GCC TTC TAG TTG CCA GCC ATC TGT TG 54.
(55) Appendix IV The plasmid map pSV-β-Galactosidase Vector. Restriction Map of pSV-β-Galactosidase Vector.. The SV40 early promoter and enhancer drive transcription of the bacterial lacZ gene, which in turn, is translated into the β-galactosidase enzyme. β-galactosidase is an excellent reporter enzyme, that can be assayed quickly and directly in cell extracts using spectrophotometric, fluorescent or chemiluminescent assays.. 55.
(56) 56.
(57)
數據

相關文件
The purpose of this research is to study a tiling problem: Given an m × n chessboard, how many ways are there to tile the chessboard with 1 × 2 dominoes and also ”diagonal”
You are given the wavelength and total energy of a light pulse and asked to find the number of photons it
Reading Task 6: Genre Structure and Language Features. • Now let’s look at how language features (e.g. sentence patterns) are connected to the structure
This is especially important if the play incorporates the use of (a) flashbacks to an earlier time in the history of the characters (not the main focus of the play, but perhaps the
• 是細胞不正常增生,且這些增生的細胞可
incapable to extract any quantities from QCD, nor to tackle the most interesting physics, namely, the spontaneously chiral symmetry breaking and the color confinement..
Basing on the observation and assessment results, this study analyzes and discusses the effects and problems of learning the polynomial derivatives on different level students
This paper will explore the spirit of Humanistic Buddhism as found in Ox-Head Chan through analysis of six different versions of the Treatise on the Ceasing of Notions, an
