利用單株抗體探討β-乳球蛋白之結構與功能
65
0
0
全文
(2) 利用單株抗體探討乳球蛋白的結構與功能 學生: 宋俊瑩. 指導教授: 毛仁淡 博士. 國立交通大學生物科技所碩士班 β-乳球蛋白在牛乳中是一種熱不穩定蛋白。因此β-乳球蛋白易因加熱而 導致結構之改變,在先前研究中我們製作出可辨識受熱變性後β-乳球蛋白之 專一性單株抗體。於本研究中將利用此抗體深入探討β-乳球蛋白之結構與功 能。在研究中發現當生乳加熱溫度達攝氏 70 到 80 度時,此抗體與β-乳球蛋 白之結合能力會大幅地增加,證明β-乳球蛋白於加熱至攝氏 70 到 80 度時結 構 會 明 顯 改 變 。 為 了 找 出 這 抗 體 於 β- 乳 球 蛋 白 所 辨 識 之 抗 原 決 定 位 (epitope),我們利用綜合策略之免疫分析法包含: 酵素及化學法分解蛋白質 (胰蛋白酵素及 CNBr),化學修飾法(acetylation 及 carboxymethylation),胜肽 矩陣 (peptide array)與水溶性胜肽。 其結果準確的分析出抗原決定位於β乳球蛋白之 D 股上(其胺基酸為 66-76 之位置)。其次於旋光雙級光譜(Circular dichroic spectral analysis)分析中,發現 carboxymethylation 之β-乳球蛋白不僅 完全喪失二級結構,且與此單株抗體結合力遠超出經加熱後變性之β-乳球蛋 白 10 倍以上,此證明β-乳球蛋白結構形成 disorder 時會被此抗體辨識。於 抗體決定位做單點突變,結果發現 Lys-69 和 glu-74 是抗原決定位中與抗體 結合之重要胺基酸。此外,我們更證明出結構變性後之β-乳球蛋白不能與疏 水性分子(retinol 和 palmitic acid)結合,而此抗原決定位扮演重要的角色,. i.
(3) 當此抗原決定位結構改變被單株抗體所辨識,則β-乳球蛋白不能與疏水性分 子結合。而當 pH 9~10 時β-乳球蛋白上之抗體決定位的 Lys 呈電中性,會造 成其免疫反應能力與疏水性小分子結合能力下降。綜合上述結果,β-乳球蛋 白上 D 股為熱變性之區域及參與疏水性分子結合之功能。. ii.
(4) Investigation of the structure and function of β-lactoglobulin by monoclonal antibody Student: Chun-Ying Song. Advisor: Dr. Simon J.T. Mao Institute of Biotechnology. National Chiao Tung University Abstract β-lactoglobulin (β-LG) is a bovine milk protein sensitive to thermal denaturation.. Previously, we demonstrated that such structural change can be. detected by a monoclonal antibody (mAb) specific to denatured β-LG. In the present study, we show a dramatic increase in β-LG immunoreactivity when heating raw milk between 70 °C and 80 °C.. To map out the specific epitope of. β-LG recognized by this mAb, we used a combined strategies including tryptic digestion and CNBr fragments, chemical modifications (acetylation and carboxymethylation), peptide array containing in-situ synthesized peptides, and a synthetic soluble peptide for immunoassays.. The antigenic determinant we. defined was exactly located within the D strand (residues 66-76) of β-LG. Circular dichroic spectral analysis shows that carboxymethylation on β-LG not only resulted in a substantial loss of β-configuration, but also exerted a 10 x increase in immunoreactivity as compared to heated β-LG. The result suggests iii.
(5) that a further disordered structure occurred in β-LG and thus rendered the mAb recognition.. Mutations on each charged residue (three Lys and one Glu). revealed that Lys-69 and Glu-74 were extremely essential in maintaining the antigenic structure.. We also show an inverse relationship between the. immunoreactivity in heated β-LG and its binding to retinol or palmitic acid. Interestingly, at pH 9-10, which neutralizes the Lys groups of β-LG, not only reduced its immunoreactivity but also its binding to palmitic acid implicating a role of Lys-69.. Taken together, we concluded that strand D of β-LG. participated in the thermal denaturation between the temperatures of 70 °C and 80 °C and the binding to retinol and palmitic acid.. The antigenic and. biochemical roles of mAb specific to D strand are discussed in detail.. iv.
(6) 誌謝 Acknowledgements 兩年碩士的生涯,讓我不僅在學術研究的領域有學習及發揮的空間,更 讓我感受到與人相處及團隊合作的重要性;我喜愛科學但我更熱愛這群一 起奮鬥的夥伴。 今天能夠拿到著碩士學位,要感謝的人真的太多了,飲水 之恩必當湧泉以報。 首先要感謝的是一路栽培我讀到碩士學位的父母:宋金廷先生及蔡秋霞 女士,沒有他們的辛勤工作,沒有今天傑出的我,我將這份榮耀光彩獻給 他們。 接著要感謝的是毛仁淡教授,毛博無論是在實驗上的教導及要求, 或是在論文寫作上的指導,我都會謹記在心; 除此之外毛博在人生的原則 及規劃上的忠告,我也會虛心接受。. 常言道:一日為師終生為父,在此. 獻上最誠摯的感謝。 除此之外也要感謝林淑端博士、潘榮隆博士、楊裕雄 老師、楊進木老師、廖光文老師在口試及論文上的建議與指正,許多寶貴 的意見讓學生獲益良多。 在實驗室裡首先要感謝財木學長、繼鋒學長、文亮學長及以祥學長在實 驗上的教導與生活上的協助,特別是文亮學長在我還是菜鳥時特別的"照 顧 ",也特別感謝他在我這篇論文裡的指導與生活上的協助,也要感謝怡 荏學姐、彰威學長、宗義學長、盛龍學長及惠君學姊在我碩一時候實驗上、 想法上、生活上的分享與幫助。. 特別是我的麻吉彰威謝謝你在各方面的. v.
(7) 幫助。 接著我的同學中,宏輝、珮真、韶禪、很高興能跟你們一起奮鬥, 我想不論是過程或結局都是令人滿意的也都令我難以忘懷。. 特別是宏. 輝,跟你共事真是愉快,也許有一天我們可以成立 Wolfgene 一起合作。 在 學弟妹中,真是長江後浪推前浪,每個都是超人和女超人,明誌、文昭、 立品、佩茹,很高興認識你們。 特別是明誌,感謝你在我論文中的貢獻, 所以提醒你煙少抽一點。. 還有文昭,我也要感謝你這個大幫手,和你一. 起做實驗很愉快。此外中曦、威延、小涵也很高興能跟你們在這實驗室裡 打拼。除此之外,程允、晉豪、文鴻也是我求學過程中很好的夥伴,最後 也給我女朋友雅清獻上最深的感謝。 科學不僅僅是知識的累積,對我而言一個成功的發現往往都是一個合作 無間的團隊努力奮鬥的結果,因此我很榮幸能夠加入這個團隊,也很感謝 所有跟我合作過的人,謝謝你們。. vi.
(8) Content. page. Abstract in Chinese---------------------------------------------------------------------- i Abstract----------------------------------------------------------------------------------- iii Acknowedgements----------------------------------------------------------------------- v Content----------------------------------------------------------------------------------- vii Figure content--------------------------------------------------------------------------- ix Abbreviations----------------------------------------------------------------------------x Chapter 1 Introduction------------------------------------------------------------------- 1 1.1 Structure of β-LG---------------------------------------------------------------- 1 1.2 Biochemical properties of β-LG---------------------------------------------- 2 1.3 Physicochemical properites of β-LG----------------------------------------- 3 Chapter 2 Materials and Methods------------------------------------------------------ 6 2.1 Materials-------------------------------------------------------------------------- 6 2.2 Preparation of monoclonal antibody specific to dry milk------------------ 6 2.3 Trypsin and CNBr fragmentation--------------------------------------------- 7 2.4 Acetylation and carboxymethylation of β-LG------------------------------- 8 2.5 CD spectrum--------------------------------------------------------------------- 9 2.6 Peptide array--------------------------------------------------------------------- 9 2.7 Competitive ELISA------------------------------------------------------------ 10 2.8 Effect of pH on β-LG binding to mAb--------------------------------------11 2.9 Retinol and pamitic acid binding to mAb-----------------------------------11 2.10 Three dimensional analysis of β-LG structure----------------------------12 2.11 Three dimensional analysis of β-LG and palmitic acid complex structure ------------------------------------------------------------------------12 Chapter 3 Results------------------------------------------------------------------------ 13 vii.
(9) 3.1 Characterization of the monoclonal antibody specific to dry milk------ 13 3.2 Mapping of antigenic determinant of denatured β-LG utilizing tryptic digestion and acetyl-modification-------------------------------------------- 14 3.3 Immunoreactivity of CNBr fragments of β-LG---------------------------- 14 3.4 Final antigenic mapping using a solid-phase peptide array--------------- 15 3.5 Carboxymethylated β-LG and CD spectrum------------------------------- 17 3.6 Immunoreactivity of a soluble synthetic peptide--------------------------- 17 3.7 Role of charged residues in epitope specificity---------------------------- 18 3.8 Effect of pH on β-LG binding to mAb-------------------------------------- 18 3.9 Effect of pH and heating on β-LG binding to retinol and palmitic acid 19 Chapter 4 Discussion--------------------------------------------------------------------21 Chapter 5 References------------------------------------------------------------------- 32. viii.
(10) Figure content. page. Figure 1 Amino acid sequence of β-LG--------------------------------------- 39 Figure 2 3D structure of β-LG ------------------------------------------------- 40 Figure 3 Immunoreactivity of β-LG in raw milk heated with different temperature over time--------------------------------------------------- 41 Figure 4 Effect of tryptic digestion on immunoreactivity of β-LG --------42 Figure 5 Effect of acetylation on immunoreactivity of β-LG ---------------42 Figure 6 Effect of CNBr cleavage on immunoreactivity of β-LG ----------43 Figure 7 Delineation of an epitope recognized by mAb ---------------------44 Figure 8 SDS-PAGE profile of carboxymethylated β-LG (CM-LG) -------45 Figure 9 Immunoreactivity of carboxymethylated β-LG (CM-LG) --------45 Figure 10 Circular dichroic spectra of native β-LG, heated β-LG, CM-LG, and synthetic peptide 67-76 ------------------------------------------- 46 Figure 11 Immunoreactivity of the synthestic peptide determined by an ELISA--------------------------------------------------------------------- 47 Figure 12 Role of charge residues in antigenic specificity determined by a peptide array-------------------------------------------------------------- 48 Figure 13 Effect of pH on β-LG binding to mAb ----------------------------49 Figure 14 Effect of pH and heating on β-LG binding to retinol and palmitic acid ------------------------------------------------------------------------ 51 Figure 15 Three dimensional view of β-LG complexed with palmitic acid 52. ix.
(11) Abbreviations. β-LG CD CM-LG ELISA HRP LDL mAb PB PBS PDB SDS-PAGE. β-Lactoglobulin Circular dichroism Carboxymethylated-lactoglobulin Enzyme link immunosorbent assay Horseradish peroxidase Low density lipoprotein Monoclonal antibody Phosphate buffer Phosphate buffered saline Protein data bank Sodium dodecyl sulfate-polyacrylamide electrophoresis. x. gel.
(12) INTRODUCTION 1.1 Structure of β-LG Bovine β-lactoglobulin (β-LG) is one of the major proteins in milk consisting of about 10-15% (Hambling et al, 1992). The protein comprises 162 amino-acid residues, with one free cysteine and two disulfide linkages (Fig. 1).. According to. the 3D crystallographic studies, β-LG is predominantly a β-sheet configuration containing nine antiparallel β-strands from A to I (Qin et al, 1998; Qin et al, 1999; Kuwata et al, 1999) (Fig. 2).. Topographically, strands A-D form one surface of. the barrel (calyx) while strands E-H form the other.. The only 3-turn α-helical. structure is at the COOH-terminus, which follows strand H lying on the outer surface of the calyx (Uhrinova et al, 2000). It is of interest, the EF loop acts as a gate over the calyx.. It forms closed configuration to inhibit the ligand binding. while pH < 6, whereas such configuration turns to open status allowing ligands to penetrate into the calyx while pH ≥ 6.. The “latch” for this gate is Glu89, the. residue implicated in the Tanford transition observed, although not identified, as having an abnormally high pKa (Tanford et al, 1959; Brownlow et al, 1997; Qin et al, 1998). The bovine β-LG is monomeric at pH values below 3, but exists in non-covalent bonding dimeric form at natural pH (Jameson et al., 2002), 1.
(13) 1.2 Biochemical Properties of β-LG Due to the thermally unstable and molten-globule nature, β-LG has been studied extensively for its physical and biochemical properties (Qi et al, 1995; Sawyer and Kontopidis, 2000; Kontopidis et al, 2004).. Spectroscopic studies. have demonstrated that irreversible modification of the β-LG structure occurs upon thermal treatment above 65-70 °C.. Thermodynamic analysis of the calorimetric. signal reveals that there are two domains unfolding independently while heating (Fessas et al, 2001). Heat treatment of β-LG at neutral pH causes the dimeric native protein to dissociate, partially unfold, denature, and aggregate; the rates and pathways are known to be dependent on the protein concentration, pH, temperature (Havea et al, 2004).. Two major aggregating features are related to hydrophobic. association and disulfide-bond interchange reactions (Havea et al, 2004). When a -LG is heated at neutral pH, the equilibrium between the native β-LG dimers and monomers is shifted toward the monomers.. At temperatures above 60 °C, the. β-LG monomer partially unfolds, with the loss in the helical structure allowing Cys121 to interact with the Cys106-Cys119 disulfide bond, to give a non-native Cys106-Cys121 disulfide bond and Cys119 (Croguennec et al, 2003). 2. While,.
(14) Cys160 is also likely to become involved in interprotein disulfide bonding in whey protein aggregates (Creamer et al, 2004).. Thus, the resultant activated monomers. lead to the formation of various intermediate aggregation products.. Involvement. of the free thiol group in aggregation has been shown in a number of studies, most notably by site-directed mutagenesis (Cho et al., 1994) where thermal stability was enhanced by incorporating the free thiol into a third disulphide bridge to the neighbouring helix.. Further, the porcine protein has no free thiol and does not. form gels under the same conditions as the cow (Gallagher et al., 1996).. However,. the exact regions involved in the thermal denaturation are still unclear.. Whether. the subtle unfolding changes can be detected by an immunochemical approach remains at question.. 1.3 Physicochemical Properties of β-LG A remarkable physicochemical property of the β-LG is its ability to bind in vitro small hydrophobic molecules such as retinol, fatty acids, vitamin D, and cholesterol (Narayan and Berliner, 1997; Qin et al, 1998; Wu et al, 1999; Kontopidis et al, 2002). β-LG has been postulated to serve as a carrier for retinol in neonates (Sawyer et al, 1985), and it has been proposed that the binding of fatty 3.
(15) acids to β-LG at neutral pH is related to a biological function of this protein in bovine milk (Perez et al, 1992; Perez and Calvo, 1995).. The binding of fatty acids. to β-LG has been reported to increase the resistance of the protein to proteolytic degradation (Puyol et al, 1993), thermal degradation (Puyol et al, 1994), and unfolding in urea solution (Creamer, 1995).. Regardless of intensive research, biological function of this protein has not yet been satisfactorily resolved.. Recently, we immunized the mice with commercially. prepared dry milk and produced a panel of monoclonal antibodies (mAb).. From. 900 hybridomas screened a clone specific to dry milk, but not to raw milk, has been selected. Characterization of this dry milk specific mAb reveals that this antibody recognizes thermally denatured β-LG (Chen et al, 2004). It suggests that a new antigenic epitope in β-LG is being exposed by a heating process used in the preparation of dry milk.. In the present study, we defined the immunoreactive site. that was recognized by this specific mAb and attempted to relate it to the thermal denaturation properties of β-LG.. The strategy for epitope mapping combined. several approaches including tryptic and CNBr fragments, chemical modifications (acetylation and carboxymethylation), peptide array containing in-situ synthesized 4.
(16) peptides (with overlapped regions), and a synthetic peptide in solution for immunoassays.. We demonstrate that the epitope was located exactly within the D. strand of β-LG (residues 66-76). The immunoreactivity as recognized by this mAb was correlated to the thermal denaturation and conversion of β-sheet to a disordered structure of β-LG.. Interestingly, the D strand is associated with the. A-C strands forming one domain at the opening of the calyx (Fig. 1).. For this. reason, we also studied the effect of heating and pH on β-LG binding to retinol and palmitic acid. Further epitope mapping shows that conversion of Glu-74 into either Ala or negatively charged Asp totally abolished its immunoreactivity. similar result was seen in Lys-69, but not the other Lys residues.. A. Finally, we. propose that strand D plays a provocative role in the molten-globule state of β-LG as probed by our mAb.. 5.
(17) MATERIALS and METHODS. 2.1 Materials β-LG. was. purified. from. fresh. raw. milk. using. 30%. saturated. ammonium-sulfate top fraction followed by a G-150 column chromatography as described previously (Chen et al, 2004; McCreath et al, 1997).. 2.2 Preparation of Monoclonal Antibody Specific to Dry Milk Monoclonal antibodies were produced according to the standard procedures previously described by us (Mao et al, 1988; Mao et al, 1990), in which dry milk (Nestle Australia Ltd, Sidney, Australia) was used for immunization (Chen et al, 2004).. In brief, the myeloma cell line (FO) was fused with spleen cells from. immunized Balb/c mice at a ratio of 1:5.. The culture medium (between days 14. and 21 after fusion) was assayed for the production of specific antibodies by a solid-phase ELISA using both raw and dry milk as a respective antigen. Each monoclonal was established by limiting dilutions at least 2 x (Mao et al, 1988; Mao et al, 1990).. 6.
(18) 2.3 Trypsin and CNBr Fragmentation For trypsin treatment, 50 µg of β-LG in 100 µl phosphate buffered saline (PBS) containing 0.02 M phosphate and 0.12 M NaCl, pH 7.4, were preheated at 100 °C for 10 min.. After which time, 1 µl trypsin (0.1 mg/ml) was added and. incubated at room temperature for 4 h (Chen et al, 2004).. Trypsinized LG was. analyzed on a SDS-PAGE (18% polyacrylamide) followed by a Western blot. For CNBr fragmentation (Mao et al, 1975; Mao et al, 1977), 5 mg of β-LG were first dissolved in 70% (v/v) TFA with the addition of 10 mg CNBr in dark for 24 h at room temperature. After 3 x evaporation in Speed Vac (CVE 200D, ELELA, Japan) with the addition of 5 x volume of de-ionized water, the dry material was dissolved in the 10 mM phosphate buffer (PB), pH 7.0. The immunoreactivity of CNBr fragments were then analyzed on an 18% SDS-PAGE, followed by a Western blot.. 7.
(19) 2.4 Acetylation and Carboxymethylation of β-LG Chemical modification of β-LG by acetylation was conducted by a modification of the procedure previously described by us (Mao et al, 1980). To 5 mg of β-LG in 2 ml 50 mM sodium bicarbonate (pH 8.0) containing 6 M urea, 5 µl of acetic anhydride were slowly added into the reaction mixture step by step, while maintaining the pH at 8.0 using 0.1 M NaOH.. After 3 h incubation at room. temperature, the acetylated protein was desalted on Bio-gel P-2 column eluted by 0.05 M ammonium bicarbonate and lyophilized.. For carboxymethylation (Mao et. al, 1980; Tseng et al, 2004), 5 mg of β-LG were first dissolved in 5 ml of 0.1 M Tris-HCl buffer (pH 8.6) containing 6 M ultra pure urea and 0.02 M dithiothreitol. Following flushing with nitrogen, 20 mg of iodoacetic acid were added into the reaction mixture, while maintaining the pH at 8.6 by the addition of 0.1 M NaOH and incubation for another 3 h.. Finally, carboxymethylated (CM) β-LG was. desalted on a Bio-Gel P2 column eluted by 0.05 M ammonium bicarbonate and lyophilized.. By amino acid analysis, the CM-β-LG contained 4.98 residues of. CM-cysteine per mole of β-LG.. 8.
(20) 2.5 CD Spectrum The secondary structure of native, heated or chemically modified β-LG was determined. using. spectropolarimeter.. a. computerized. Jasco. J-715. circular. dichroic. (CD). Each protein sample was dissolved in 10 mM phosphate. buffer at pH 7 with a final concentration of 0.2 mg/ml.. About 300 µl of the. protein solution were used for analyzation within a cuvette of 1-mm path length. The obtained spectra were accumulated for 25 times at a scanning rate of 50 nm/min.. All the data were shown as the mean residue molar ellipticity [θ]MRW. (Tseng et al, 2004; Chen et al, 1994).. 2.6 Peptide Array Twelve synthetic peptides in one nitrocellulose-array, each containing 15 amino-acid residues, were designed corresponding to the residues 25-107 of β-LG or to the residues 67-75 within strand D (Fig. 1). The synthetic peptides were prepared under a contract with a local biotechnology company (Genesis Biotech Inc., Taipei, ROC).. Briefly, the peptides were directly synthesized in situ on a. nitrocellulose (NC) paper according to the method described (Frank, 1992). The NC membrane in 0.01 M Tris buffered saline containing 0.05% (v/v) Tween-20 9.
(21) (TBST), was blocked with 5% (w/v) gelatin in TBST for 2 h at room temperature followed by 3 x washes.. After incubation with mAb for 2 h and 3 x washes, goat. anti-mouse IgG conjugated with horseradish peroxidase (HRP) in 5% gelatin/TBST was added and incubated.. Finally, following the washes, chemiluminescent. substrate (ECL™ Western Blotting System, Amersham) was added, washed, and immediately developed by exposing onto a film.. 2.7 Competitive ELISA In brief, heated β-LG (1 µg in 50 µl of PBS) was first immobilized onto microtiter wells followed by 3 x washes to remove unbound β-LG (Chen et al, 2004; Mao et al, 1989).. The wells were then blocked by 3% gelatin in PBS.. After 3 x. washes, 50 µl of the competitive protein (β-LG, heated β-LG, acetylated or carboxymethylated β-LG, or synthetic peptide residues 67-76) in PBS containing 3% gelatin were mixed with 50 µl of mAb and incubated at room temperature for 1 h.. Following washes and secondary antibody (goat anti-mouse IgG conjugated. with HRP) incubation, the microtiter plate was developed with 2, 2-Azino-bis (3-ethylbenz-thiazoline-6-sulfonic acid) and read at 415 nm.. 10.
(22) 2.8 Effect of pH on β-LG Binding to mAb Heated β-LG (1 µg/well) was immobilized onto a microtiter plate followed by blocking and washing at neutral pH.. The immobilized β-LG was then incubated. with mAb at various pH for 2 h at room temperature.. After removing the. unbound mAb, the plate was developed according to the standard ELISA procedures at neutral pH (Chen et al, 2004; Mao et al, 1989).. Since pH itself can. affect the antigen-antibody binding, a control experiment using mouse IgG as immobilized antigen was also conducted at various pH for a parallel comparison.. 2.9 Retinol and Palmitic Acid Binding to β-LG β-LG was reported to be a 1-to-1 binding ratio with retinol or palmitic acid as measured by fluorescence emission techniques (Yang et al, 2002).. In general,. binding of retinol to β-LG was measured by extrinsic fluorescence emission of retinol molecule at 470 nm using excitation at 287nm.. Whereas, binding of. palmitic acid to β-LG was measured by the fluorescence enhancement of Trp residues of β-LG at 332 nm using excitation at 287 nm.. For the effect of the pH. experiment, 5 uM or 20 uM of native β-LG was instantly incubated with 5 uM of retinol or 20 uM of palmitic acid, respectively, at various pH at 24 °C. 11. For the.
(23) effect of the heat experiment, β-LG was preheated at 80 °C or 100 °C for 5 min and then incubated with retinol or palmitic acid at pH 8.. Fluorescence spectra were. recorded at 24 °C with a fluorescence Spectrophotometer F-4500 (Hitachi High-Tech.Cor., Tokyo, Japan).. 2.10 Three Dimensional Analysis of β-LG Structure 3D structure of β-LG used in this context was provided by Protein Data Bank (PDB, http://www.rcsb.org/pdb/), code 1CJ5 (Kuwata et al, 1999), with the diagram created by PyMOL (DeLano, 2002).. 2.11 Three Dimensional Analysis of β-LG and Palmitic Acid Complex Structure 3D structure of Bovine β-LG complexed with palmitic acid used in this context was provided by Protein Data Bank (PDB, http://www.rcsb.org/pdb/), code 1GXA (Kontopidis et al, 2002), with the diagram created by Rasmol (Sayle et al, 1995).. 12.
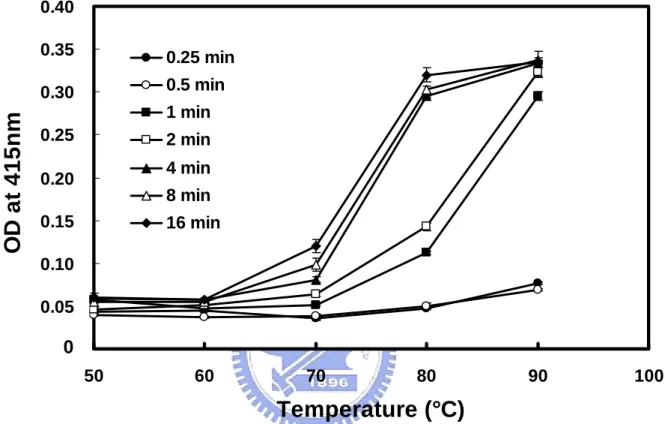
(24) RESULTS 3.1 Characterization of The Monoclonal Antibody Specific to Dry Milk Previous studies show that the monoclonal antibody (mAb) used in this report is specific to processed dry milk, but not to raw milk (Chen et al, 2004). recognizes β-LG, one of the major milk proteins.. It only. The mAb is apparently able to. discriminate the denatured β-LG from a given milk product (Chen et al, 2004). Since heating procedures are used to process the dry milk, the finding indicates that β-LG undergoes a conformational rearrangement, which facilitates the binding of this mAb.. In the present study, we show a dramatic and sharp increase in β-LG. immunoreactivity when raw milk was heated between 70 °C and 80 °C over time (Fig. 3). It was of interest that the increase in immunoreactivity was concomitant with the reported transition temperature for converting native to denatured β-LG (de Wit et al, 1980). The finding suggests that the immunoreactive site recognized by this mAb lied in the thermal denatured region of β-LG.. 13.
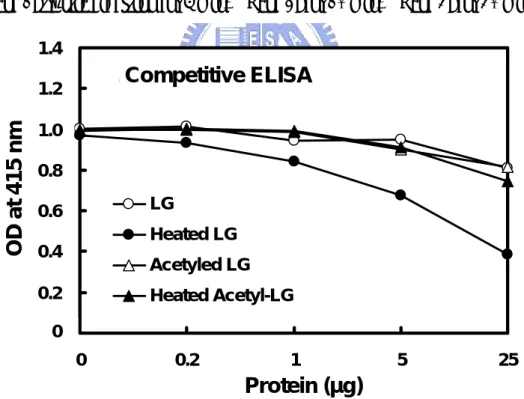
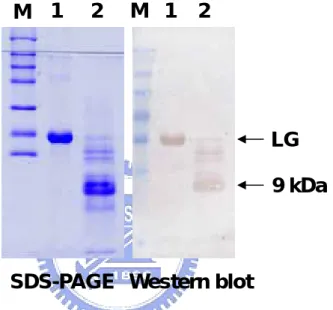
(25) 3.2 Mapping of Antigenic Determinant of Denatured β-LG Utilizing Tryptic Digestion and Acetyl-modification To initially map out the specific immunoreactive region, heated β-LG was limitedly digested by trypsin.. On Western blot, we demonstrated that the. immunoreactivity was totally abolished after the trypsin treatment (Chen et al, 2004) suggesting that Lys, Arg, or both residue(s) were probably involved in maintaining the antigenic structure for β-LG (Fig 4).. Chemical modification using acetylation. (pH 8.0) on positively charged residues (mostly Lys) attenuated the immunoreactivity of β-LG on a competitive ELISA (Fig. 5).. These two. experiments support the notion that positively charged amino acids of β-LG attributed for the mAb recognition.. 3.3 Immunoreactivity of CNBr Fragments of β-LG To further delineate the major antigenic domain, CNBr cleavage on β-LG was conducted.. Western blot analysis shows that there was a major immunoreactive. fragment corresponding to a molecular weight about 9 kDa (Fig. 6). As estimated from its Met cleavage site, this fragment was presumed a peptide containing residues 25-107 (Fig. 1).. Subsequently, the N-terminal sequence of this fragment 14.
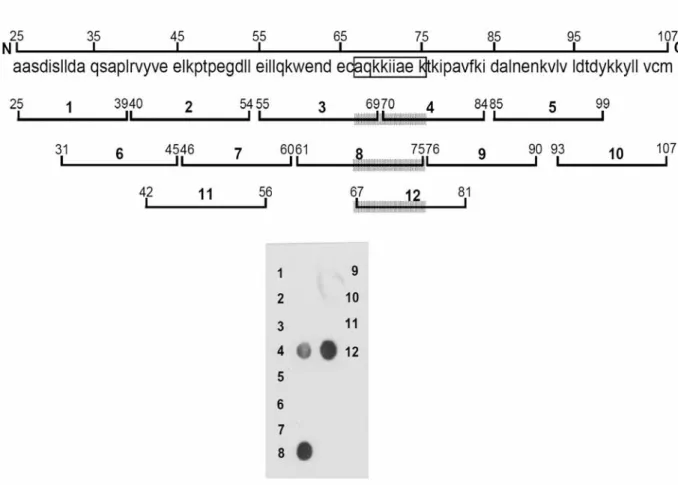
(26) was determined.. The first six amino-acid residues (AASDIS) confirmed that the. immunoreactive site was located between the residues 25-107 of β-LG (Fig. 1).. 3.4 Final Antigenic Mapping Using a Solid-phase Peptide Array As described above, Lys enriched-areas were assumed to participate in maintaining the antigenic structure.. Pro residues are also considered to be. involved as they are located at or near the antigenic determinant by forming a loop at the surface of a given protein.. Using an EMBOSS program for searching a. possible antigenic determinant within residues 25-107 of β-LG (Fig. 7), we predicted that two domains, namely residues 42-56 and 67-81, were most likely to be immunoreactive.. Accordingly, a solid-phase peptide array containing the. above predicted regions and ten other overlapped synthetic peptides (each with 15 residues) was prepared.. These peptides were directly synthesized on a. nitrocellulose membrane (Fig. 7). After binding of mAb and HRP-conjugated secondary antibody, the array was developed by a chemiluminescent agent.. We. show that only peptides 4 (residues 70-84), 8 (residues 61-75), and 12 (residue 67-81) were immunoreactive (Fig. 7).. Among which peptide 4 gave a partial. immunoreactivity suggesting that residues 67-70 were essentially involved in the 15.
(27) reactive site (Fig. 7).. Since the size of an epitope is relatively small usually. containing 6-9 amino-acid residues (Mao et al, 1990;. Mao et al, 1989; Bhatnagar. et al, 1983; Davies and Cohen, 1996; Atassi, 1984), it was possible to narrow down the reactive site from the immunoreactivity in overlapped peptides.. We proposed. that the reactive site was closely associated with AQKKIIAEK (or nine residues 67-75) (Fig. 7).. Notably, this region is highly positive in charges.. Observing. from the high resolution crystal structure of β-LG (Kuwata et al, 1999), it is fascinating that this proposed region is exactly located within the D strand of surfaced β sheet (residues 66-76) (Fig. 1 ). immunoreactive site as an epitope.. We therefore defined this. Another noticed point is that there is a. disulfide linkage between strand D (Cys-66) and carboxyl terminus (Cys-160). This disulfide linkage plays an important role to stabilize the β-structure by forming antiparallel sheets of β-LG.. The proposed epitope (67-75) in its native. state is rather ordered with a β-sheet span about 28 Å in length. As such, the orientation in native β-LG may prohibit the binding of our specific mAb.. We. hypothesized when the D strain underwent disordered structure, it would then allow the “denatured” mAb binding.. 16.
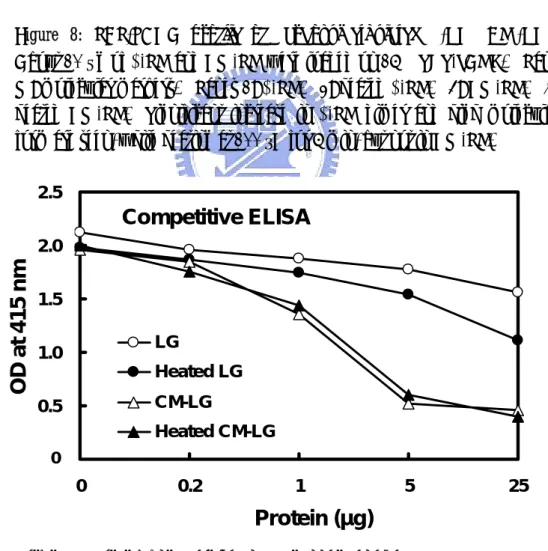
(28) 3.5 Carboxymethylated β-LG and CD Spectrum To test the above hypothesis by which the conformational change of the D strand would enhance the binding of our mAb, we chemically modified all the Cys residues to irreversibly block the disulfide linkages within the native β-LG (Fig. 8). Using a competitive ELISA with heated β-LG as a positive control, it revealed that carboxymethylation on β-LG resulted in a striking increase in its immunoreactivity. The increase was about 10 x greater than that of heated β-LG (Fig. 9). Meanwhile, analysis of CD spectra on carboxymethylated β-LG further confirmed a significant conformational change by converting β sheet (typically at 215 nm) to a more disordered structure than that of heated β-LG (Fig. 10).. 3.6 Immunoreactivity of a Soluble Synthetic Peptide Finally, a soluble peptide corresponding to the linear sequence of strand D (residues 67-76 or AQKKIIAEKT) was synthesized.. Figure 11 shows that this. linear sequence was able to completely inhibit mAb binding to heated β-LG on a competitive ELISA.. Furthermore, this synthetic peptide exhibited a typical. disordered structure rather than a β configuration (Fig. 10).. 17.
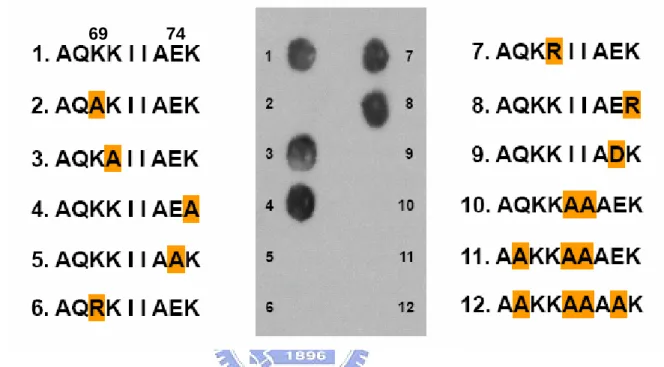
(29) 3.7 Role of Charged Residues in Epitope Specificity To determine which Lys(s) were responsible for the mAb recognition, mutation on each Lys (Lys-69, Lys-70, and Lys-75) with Ala was conducted.. As. shown in Fig. 8, only Lys-69 was extremely specific for the mAb binding. Replacement with positively charged Arg did not salvage the immunoreactivity. Glu-74 played a similar role, replacement with Ala or negatively charged Asp failed to show any immunoreactivity.. Ile-71 and Ile-72 also played an essential. hydrophobic role, although the exact residue has not been identified.. Meanwhile,. negative control peptides (peptides 11-12, Fig. 12) retaining all the Lys residues did not show any binding.. 3.8 Effect of pH on β-LG Binding to mAb Because the structural stability of β-LG is pH dependent (Casal et al, 1988), we tested whether changes of pH could also induce an increase in β-LG immunoreactivity. Fig. 13A shows that the CD structure of β-LG was stable at pH 2 with some changes between 3 and 7, while a transition to disorder was seen from 8 to 10. However, such disordered structure did not facilitate the mAb binding (Fig. 13B).. Since Lys-69 was essential (Fig. 12) and the overall positive charge of 18.
(30) this residue started to become neutralized under pH 8-10, the immunoreactivity was decreased (Fig. 13B).. A control experiment showing a typical pH dependent. antigen-antibody reaction was performed (Fig. 13C), and there was a slight decrease in immunoreactivity at pH 9-10.. 3.9 Effect of pH and Heating on β-LG Binding to Retinol and Palmitic Acid To explore the correlation between the structural change of β-LG (at various pH) and its retinol binding, we monitored the extrinsic fluorescent change of retinol upon the binding to β-LG.. The optimal binding for retinol appeared to be at pH. between 8 and 10 (Fig. 14A).. Heating β-LG at temperatures greater than 80 °C. almost completely abolished its binding for retinol (Fig. 14B).. The data support. the notion that the striking increase in immunoreactivity of the D strand at this temperature (Fig. 3) was negatively correlated to the retinol binding, which requires the integrity of a β-sheet structure of β-LG.. Interestingly, the binding to palmitic acid was decreased in some extent at pH 9-10 (Fig. 14C) correlated with the binding to mAb (Fig. 13B).. Since Lys-69. played an extremely essential role in the antigenic site (Fig. 12), such correlation 19.
(31) suggests that the protonated state of this residue might be involved in stabilizing both the mAb binding and β-LG-palmate complex formation (please see more detail in Discussion).. Similarly, heating on β-LG substantially reduced the. binding for palmitic acid.. 20.
(32) DISCUSSION Molten globules are thought to be general intermediates in protein folding and unfolding (Ptitsyn et al, 1990; Chang and Li, 2001).. β-LG, a major moiety of. bovine whey proteins, is one of the most investigated models for understanding the mechanism involved in protein stability upon heating. Although the 3D crystal structure of β-LG has been elucidated, the area involved in thermal denaturation remains unclear.. On the other hand the region responsible for Tanford transition. (Qin et al, 1998), occurring at pH from 6.5 to 8.0, is known to be within the residues 85-90 (E-F loop).. This region opens or blocks the entrance of the calyx. (Ragona et al, 2003).. The present study demonstrates that denatured strand D of β-LG was responsible for the binding of our thermally sensitive mAb (Chen et al, 2004). Several unique features of the binding are identified.. First, heating on native. β-LG resulted in a loss of β-sheet to more disordered structure (Fig. 10) in which the immunoreactivity was concomitantly increased (Fig. 9). Second, blocking the disulfide linkage between the D strand (Cys-66) and C-terminus (Cys-160) of β-LG by carboxymethylation not only produced a disordered structure (Fig. 10) but also 21.
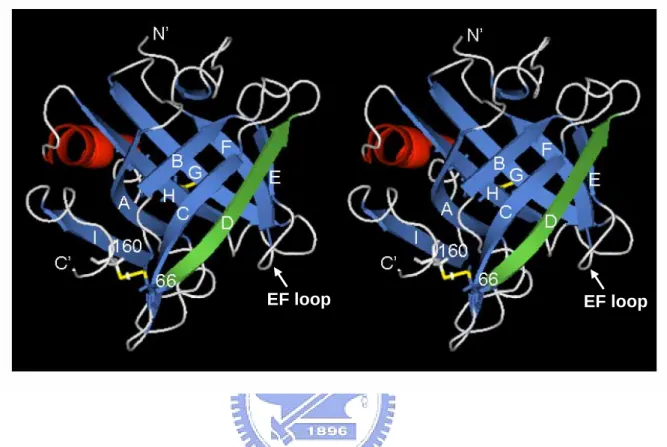
(33) markedly enhanced the mAb binding. β-LG did not give more binding.. Further heating on carboxymethylated. Such enhancement was even greater than heated. β-LG (10 x) on competitive ELISA (Fig. 9).. Presumably, this was due to the. augmented degree of freedom of the D strand without the disulfide linkage rendering more antibody binding.. It is of interest to point out that the secondary. structure of strand D alone, without including strand C, is predicted as 50% random coiled (residues 66-70) and 50% helical (residues 71-76) using the parameters from 3D-PSSM:. the. folding. (www.sbg.bio.ic.ac.uk/~3dpssm/). anti-parallel. orientation,. β-configuration.. the. recognition. server. at. the. ICRF. However, in the presence of strand C as an predicted. structure. of. strand. D. becomes. Obviously, the formation of an anti-parallel β-structure in native. β-LG molecule is stabilized through the help of a disulfide-linkage (Cys-66 and Cys-160) between strand D and the helical domain at COOH-terminus (Fig. 2). Thus, conformational change on strand D played a vital role for the mAb recognition.. It is worth mentioning that although severe heating might break the. disulfide linkage at Cys-66, it would be immediately “stabilized” via re-oxidation by forming high molecular or self-associated polymers as shown in our previous report (Chen et al, 2004). Thus, the immunoreactivity of heated β-LG was less 22.
(34) than that of carboxymethylated β-LG.. Third, the soluble synthetic-peptide. (residues 67-76) corresponding to strand D (without Cys-66) was able to completely inhibit the binding of mAb to β-LG.. Fourth, the D strand is. topographically located at the surface of β-LG (Fig. 2), which is agreeable with the general concept of a given antigenic epitope (Atassi, 1980).. Fifth, the buried side. chain of Lys-69 was exposed upon the heating and then recognized by the mAb (described below).. With respect to the exact size of the epitope that was recognized by our mAb, we excluded the possibility of Cys-66 as part of the epitope from the D strand. First, peptide 67-81 without Cys-66 gave an almost equal immunoreactivity to that of peptide 61-75 with Cys-66 in a peptide array assay (Fig. 7) suggesting that Cys-66. might. not. be. located. in. antigenic. determinant.. Second,. carboxymethylation on whole β-LG molecules with Cys-66 included in the modification markedly increased its immunoreactivity.. If Cys-66 was involved in. the antigenic site, introduction of such a bulky group (carboxymethyl) on this residue would have resulted in a significant loss of immunoreactivity (Atassi, 1984). It should be noted here that Cys-66 was only responsible for the conformational 23.
(35) restrain by crossly linking Cys-66 and Cys-160; as such it limited the binding for “denatured” mAb. Third, the length of a linear epitope can be as short as 6-7 residues as demonstrated by our previous work (Mao et al, 1990; Mao et al, 1989; Bhatnagar et al, 1983) and others (Davies and Cohen, 1996; Atassi, 1984).. The. present study shows the involvement of Glu-74 in the epitope (Fig. 12), which is eight residues apart from Cys-66.. Finally, our synthetic soluble peptide without. Cys-66 could completely inhibit the mAb binding (Fig. 11).. Regardless, our. antigenic mapping suggests that strand D located on the surface is involved in the molten globule and the unfolding structure of β-LG while heated.. Coincidently,. the transition temperature of native β-LG was between 70-80 °C which is in agreement with the increase in immunoreactivity for this mAb.. On the other hand,. the polyclonal antibody raised against the native β-LG did not exhibit such a unique property (Chen et al, 2004).. The antigenic mapping from this study also provides some interesting insight as to the specificity of antigen-antibody interaction.. Lys-69 and Glu-74 in the. epitope were found to be extremely essential in maintaining the antigenic structure (Fig. 12), substituting each with uncharged Ala diminished the mAb binding. 24.
(36) Replacing each with the same charged amino acid Arg and Asp, respectively, could not restore the immunoreactivity.. However, such point mutation in a given. protein between the same charged residues Lys/Arg or Glu/Asp is very common within or among the species, while still maintaining its biological function. Notably, fragment 70-84 (Fig. 4) without Lys-69 gave a partial immunoreactivity which differed from the mutation experiment (Fig. 12).. Although the mechanism. involved remains elusive, one possible explanation was the different solution property between the peptide with and without Lys-69.. We speculate that the. binding was more specific with the increase of chain length of the epitope, such as “lock and key”.. Another interesting feature is that the span between Lys-69 and. Glu-74 is six amino acids, which are sufficient to form an epitope as previously described by us using a mAb against fibrin (Mao et al, 1990).. It is consistent to. our observations for the interaction of protein antigens with antibodies, in which six determining residues involved in binding for the most part antigen (Mao et al, 1990; Davies and Cohen, 1996). We also demonstrate the importance of Ile-72 and Ile-73 within residues 69-73; substituting these two hydrophobic residues by Ala diminished its immunoreactivity.. Whereas, a further delineation using point. mutation to each residue is needed to draw a final conclusion. 25.
(37) Using mAb as a probe to study the structural and functional relationship of a given protein has been popular and reviewed (Miller and Cohen, 1991).. It. provides a powerful tool in defining the functional location within the molecule. Previously, we have shown mAb prepared against human hepatic lipase can distinguish between active and inactive forms of lipase (Mao et al, 1988).. Some. unique low- density-lipoprotein (LDL) mAb have been used for discriminating between patients with and without coronary artery disease (Patton et al, 1983; Mao, et al, 1983).. Those LDL mAb have also been utilized for probing the thermal. changes of human LDL (Mao et al, 1982), while the immunoreactivity was conversely correlated to the temperature with the optimal binding at 4 °C (Mao et al, 1983; Mao et al, 1982).. Since the 3D structure of lipase and LDL were lacking,. the exact mechanism involved remains elusive.. The epitope of β-LG we mapped. in this study, however, is known and provides a better understanding for its interaction with mAb.. We propose that due to highly ordered β-configuration of. D strain in its native state, the binding to our specific mAb is prohibited.. But the. D chain underwent a conformational change upon heating, which would then allow the “denatured” mAb binding.. 26.
(38) With respect to physiologic significance, the epitope region we defined is located at one of the critical domains forming a conical central calyx that is responsible for the binding of retinol and fatty acids (Wu et al, 1999; Kontopidis et al, 2002).. In vivo experiment shows that β-LG enhances the intestinal uptake of. retinol in preruminant calves (Kushibiki et al, 2001).. It has been suggested that. conformational changes in the calyx and the exposure of surface hydrophobic site of β-LG in molten globule state reduces the retinol affinity (Yang et al, 2002). The binding specificity may be determined by the dynamic motion of loops between the β strands (Ragona et al, 2003; Abduragimov et al, 2000).. It is not. known which β strand(s) is involved for the exposure of surface hydrophobic site. In this study we show a substantially decreased binding of retinol to β-LG that was preheated at transition temperature 80 °C (Fig. 14), while a significant increase in immunoreactivity (Fig. 3).. Notably, two hydrophobic residues Ile-71 (buried in. native state) and Ile-72 (exposed in native state) are located within the epitope and participate in the binding to mAb (Fig. 12).. One or both of these two residues. must be exposed to the surface during thermal transition for rendering mAb binding. Thus, it is conceivable that Ile-71 might have become exposed on the surface during heating.. Using cocrystallized β-LG with palmitic acid, the refined 27.
(39) structure (at 2.5-Å resolution) reveals that the carboxyl group of palmitic acid binds to both Lys-60 and Lys-69 (Qin et al, 1998; Wu et al, 1999).. Fig. 15 shows a. stereo view in which Lys-69 lies exactly within the antigenic epitope and orients at the entrance of the binding pocket, while the side chain of Ile-71 may interact with the fatty acyl chain.. For the retinol binding, there is no obvious contact between. the hydroxyl group of retinol and Lys-69 (Kontopidis et al, 2002), although Lys-69 is in a proximity oriented to the hydroxyl group (Cho et al, 1994).. Again, we. show a decreased binding of palmitic acid to β-LG that was preheated at 80 °C (Fig. 14), while a significant increase in immunoreactivity (Fig. 3).. Due to that β-LG and human retinol binding protein both belong to lipocalin superfamily and possess the retinol binding ability (Flower et al, 2000). By comparing their biochermical properties in all respects to realize whether or not we could apply our findings to the human retinol binding protein. To structure, their three dimensional structure are very similar and both have an eight-stranded β-barrel (caylx) accompanied by a short α-helix closing to C terminus (Fig. 2) (Greene et al, 2001 and Qin et al, 1998). However, their primary protein sequences show that there is no significant identity. To heating treatment, they both reveal the 28.
(40) similar thermal transition temperature (70-80 °C) (Muccio et al, 1992), but there is no corresponding thermal sensitive domain as we determined in the human retinol binding protein. Additionally, thermal denaturation is irreversible to β-LG, but not to human retinol binding protein (Greene et al, 2001and Muccio et al, 1992). To ligands binding ability, β-LG is able to bind retinol and fatty acid in the caylx binding pocket, nevertheless the retinol binding protein is able to specifically bind retionol derivatives. The retinol bindinf affinity of β-LG and human retinol binding protein are similar with Kd = 4.7x 10-8M and 7.5x 10-8M, respectively (Muccio et al, 1992 and Cho et al, 1994). Taken together, human retinol binding protein is similar to β-LG in some aspects, but our findings in β-LG can not totally fit in that of human retinol binding protein.. We demonstrate that the binding of retinol to β-LG was pH dependent with initial binding at pH 7, while reaching maximal pH between 8 and 10 (Fig. 14A). The data is consistent to the Tanford transition occurring at pH from 6.5 to 9.5 (Tanford et al, 1959).. Calyx opens at pH 7.1-8.2 (Qin et al, 1998) and closes at. pH 2.6 (Uhrinova et al, 2000).. Interestingly, the binding of palmitic acid was. attenuated at pH from 8 to 10 (Fig. 14C). 29. One of the possible explanations is that.
(41) the protonated state of buried side chain of Lys-69 was neutralized at pH above 8 and resulted in weakening the ionic interaction with the carboxyl group of palmate. Of course we could not rule out the other part of β-LG that might also undergo conformational changes at high pH affecting the interaction with palmate.. We. also speculate that the change of protonated state of Lys-69 contributed in the decreased immunoreactivity at pH 8-10 (Fig. 14C).. As mentioned above, the immunoreactivity assessed by this mAb was positively correlated to its molten globule state (between 70-80 °C in Fig. 3).. This. epitope in strand D was not only thermal sensitive but also negatively correlated to retinol and palmate binding.. Lys-69 in native state participates in the binding for. palmitic acid (Wu et al, 1999), but in heat denatured state it contributes to the binding for mAb.. Therefore, this mAb may be used as a probe to study the. thermal changes and the physiologic activity of β-LG, such as its binding to fatty acids and retinol.. Much is known about the physicochemical properties of β-LG (Sawyer and Kontopidis, 2000).. However, the biological function of this protein in addition to 30.
(42) the transport of retinol and fatty acids has not yet been satisfactorily resolved. Recent studies have shown that β-LG produces hypocholesterolemic (Nagaoka et al, 2001; Yamauchi et al, 2003) and antioxidant effects (Zommara et al, 1998; Chevalier et al, 2001) and may also serve as a growth factor for mammalian cells (Francis et al, 1995).. The protein is acid resistant in the gastro-intestinal tract. with a superior absorption capability via a receptor-mediated process (Guo et al, 1995; Papiz et al, 1986; Caillard and Tome, 1994).. Since β-LG is a major protein. consisting of about 10-15% of total milk proteins (Hambling et al, 1992) and is labile to heat treatment by forming large polymers with other milk proteins (Chen et al, 2004; Havea et al, 2001), it is conceivable that overheating should be avoided in order to maintain the physiologic role of β-LG.. Our mAb may, therefore, be. useful for monitoring the immunochemical and biochemical nature of β-LG in heat processed milks.. From a technological standpoint, the monoclonal antibody. prepared against the strand D region may be relevant to the design and operation of appropriate processes for thermal sanitation of milk and of other dairy products. Taken together, we conclude that our mAb was able to discriminate the dry milk from the raw milk.. The immunoreactivity of thermal denatured β-LG was. correlated to its molten globule state and structural changes from primarily β-sheet 31.
(43) to disordered conformation.. Epitope mapping reveals that Lys-69, Ile-71, Ile-72,. and Glu-74 in denatured D strand were directly involved in binding to mAb.. The. data suggest that the D strand plays a critical role in β-LG thermal denaturation, in which the buried side chains of Lys-69 and Ile-71 were exposed to the surface of β-LG calyx.. At thermal transition temperature 80 °C, the increased. immunoreactivity was associated with the decreased retinol and palmate binding. We propose the mAb produced in this study may be used as a probe to study the thermal changes and the physiologic activity of β-LG, such as its binding to fatty acids and retinol.. Whether the other β strands may also participate in this role. upon the heating between 70-80 °C, remains to be addressed.. 32.
(44) REFERENCES 1. Abduragimov, A. R., Gasymov, O. K., Yusifov, T. N., and Glasgow, B. (2000) J. Curr. Eye Res. 21, 824-32. 2. Atassi, M. Z. (1980) Mol. Cell Biochem. 32, 21-43. 3. Atassi, M. Z. (1984) Eur. J. Biochem. 145, 1-20. 4. Bhatnagar, P. K., Mao, S. J. T., Gotto, A. M. Jr., and Sparrow, J. T. (1983) Peptides 4, 343-9. 5. Brownlow, S., Cabral, J. H. M., Cooper, R., Flower, D. R., Yewdall, S. J., Polikarpov, I., North, A. C. T., and Sawyer, L. (1997) Structure 5, 481-495. 6. Caillard, I., and Tome, D. (1994) Am. J. Physiol. 266, 1053-9. 7. Casal, H. L., Kohler, U., and Mantsch, H.H. (1988) Biochim. Biophys. Acta. 957,11-20. 8. Chang, J. Y., and Li, L. (2001) J. Biol. Chem. 276, 9705-12. 9. Chen, L., Mao, S. J. T., McLean, L. R., Powers, R. W., and Larsen, W. J. (1994) J. Biol. Chem. 269, 28282-7. 10. Chen, W. L., Huang, M. T., Liu, H. C., Li, C. W., and Mao, S. J. T. (2004) J. 33.
(45) Dairy Sci. 87, 2720-9. 11. Chevalier, F., Chobert, J. M., Genot, C., and Haertle, T. (2001) J. Agric. Food Chem. 49, 5031-8. 12. Cho, Y., Batt, C. A., and Sawyer, L. (1994) J. Biol. Chem. 269, 11102 - 11107. 13. Cho, Y. J., Gu, W., Watkins, S., Lee, S. P., Kim, T. R., Brady, J. W., and Batt, C.A. (1994) Protein Eng. 7, 263-270. 14. Creamer, L. K. (1995) Biochemistry 34, 7170-7176. 15. Creamer, L. K., Bienvenue, A., Nilsson, H., Paulsson, M., van Wanroij, M., Lowe, E. K., Anema, S. G., Boland, M. J., and Jime´nez- Flores, R. (2004) J. Agric. Food Chem. 52, 7660-7668. 16. Croguennec, T., Bouhallab, S., Molle, D., O’Kennedy, B. T., and Mehra, R. (2003) Biochem. Biophys. Res. Commun. 301, 465-471. 17. Davies, D. R., and Cohen, G. H. (1996) Proc. Natl. Acad. Sci. U S A. 93, 7-12. 18. DeLano, W. L. The PyMOL Molecular Graphics System (2002) DeLano Scientific, San Carlos, CA, USA. 19. de Wit, J. N., and Swinkels, G. A. (1980) Biochim. Biophys. Acta. 624, 40-50. 20. Fessas, D., Iametti, S., Schiraldi, A., and Bonomi, F. (2001) Eur. J. Biochem. 268, 5439-48. 34.
(46) 21. Flower, D. R. (2000) Biochim. Biophys. Acta. 1482, 327-336. 22. Francis, G. L., Regester, G. O., Webb, H. A., and Ballard, F. J. (1995) J. Dairy Sci. 78, 1209-18. 23. Frank, R. (1992) Tetrahedron 48, 9217-9232. 24. Gallagher, D. P., Lynch, M. G., and Mulvihill, D. M. (1996) J. Dairy Res. 63, 479-482. 25. Greene, L. H., Chrysina, E. D., Irons, L. I., Papageorgiou, A. C., Acharya, K. R., and Berw, K. (2001) Protein Sci. 10, 2301-16. 26. Guo, M. R., Fox, P. F., Flynn, A., and Kindstedt, P. S. (1995) J. Dairy Sci. 78, 2336-44. 27. Hambling, S. G., MacAlpine, A. S., and Sawyer, L. (1992) in Advanced Dairy Chemistry (Fox, P. F., ed.) pp. 141–190, Elsevier, Amsterdam 28. Havea, P., Carr, A. J., and Creamer, L. K. (2004) J. Dairy Res. 71, 330-339. 29. Havea, P., Singh, H., and Creamer, L. K. (2001) J. Dairy Res. 68, 483-97. 30. Jameson, G. B., Adams, J. J., and Creamer, L. K. (2002) Int.Dairy J. 12, 319–329. 31. Kontopidis, G., Holt. C., and Sawyer, L. (2002) J. Mol. Biol. 318, 1043–1055 32. Kontopidis, G., Holt, C., and Sawyer, L. (2004) J Dairy Sci. 87, 785-96. 35.
(47) 33. Kushibiki, S., Hodate, K., Kurisaki, J., Shingu, H., Ueda, Y., Watanabe, A., and Shinoda, M. (2001) J. Dairy Res. 68, 579-86. 34. Kuwata, K., Hoshino, M., Forge, V., Era, S., Batt, C. A., and Goto, Y. (1999) Protein Sci. 8, 2541–2545 35. Mao, S. J. T., Gotto, A. M. Jr., and Jackson, R. L. (1975) Biochemistry 14, 4127-31. 36. Mao, S. J. T., Kazmar, R. E., Silverfield, J. C., Alley, M. C., Kluge, K., and Fathman, C. G. (1982) Biochim. Biophys. Acta. 713, 365-374 37. Mao, S. J. T., Patton, J. G., Badimon, J. J., Kottke, B. A., Alley, M. C., and Cardin, A. D. (1983) Clin. Chem. 29, 1890-7. 38. Mao, S. J. T., Rechtin, A. E., and Jackson, R. L. (1988) J. Lipid Res. 29, 1023-9. 39. Mao, S. J. T., Rechtin, A. E., Krstenansky, J. L., and Jackson, R. L. (1990) Thromb. Haemost. 63, 445-8. 40. Mao, S. J. T., Sparrow, J. T., Gilliam, E. B., Gotto, A. M. Jr., and Jackson, R. L. (1977) Biochemistry 16, 4150-6. 41. Mao, S. J. T., Sparrow, J. T., Gotto, A. M., and Jackson, R.L. (1980) Biochim. Biophys. Acta. 617, 245-253. 36.
(48) 42. Mao, S. J. T., Yates, M.T., Owen, T. J., and Krstenansky, J. L. (1989) J. Immunol. Methods 120, 45-50 43. McCreath, G. E., Owen, R. O., Nash, D. C., and Chase, H. A. (1997) J. Chromatogr. A. 773, 73-83. 44. Miller, E. J., and Cohen, A. B. (1991) Am. J. Physiol. 260, 1-12. 45. Muccio, D. D., Waterhous, D. V., Fish, F., and Brouillette, C. G. (1992) Biochemistry 31, 5560-5567. 46. Nagaoka, S., Futamura, Y., Miwa, K., Awano, T., Yamauchi, K., Kanamaru, Y., Tadashi, K., and Kuwata, T. (2001) Biochem. Biophys. Res. Commun. 281, 11-7. 47. Narayan, M., and Berliner, L. J. (1997) Biochemistry 36, 1906–1911 48. Papiz, M. Z, Sawyer, L., Eliopoulos, E. E., North, A. C., Findlay, J. B., Sivaprasadarao, R., Jones, T. A., Newcomer, M. E., and Kraulis, P. J. (1986) Nature 324, 383-5. 49. Patton, J. G., Badimon, J. J., and Mao, S. J. T. (1983) Clin. Chem. 29, 1898-903. 50. Perez, M. D., and Calvo, M. (1995) J. Dairy Sci. 78, 978-988. 51. Perez, M. D., Sanchez, L., Aranda, P., Ena, J. M., Oria, R., and Calvo, M. 37.
(49) (1992) Biochim.Biophys. Acta 1123, 151-155. 52. Ptitsyn, O. B., Pain, R. H., Semisotnov, G. V., Zerovnik, E., and Razgulyaev, O. I. (1990) FEBS Lett. 262, 20-4. 53. Puyol, P., Perez, M. D., Mata, L., Ena, J. M., and Calvo, M. (1993) Int. Dairy J. 3, 589-597. 54. Puyol, P., Perez, M. D., Peiro, J. M., and Calvo, M. J. Dairy Sci. 1994, 77, 1494- 1502. 55. Qi, X. L., Brownlow, S., Holt, C., and Sellers, P. (1995) Biochim. Biophys. Acta. 1248, 43-49. 56. Qin, B. Y., Bewley, M. C., Creamer, L. K., Baker, H. M., Baker, E. N., and Jameson, G. B. (1998) Biochemistry 37, 14014–14023 57. Qin, B. Y, Bewley, M. C., Creamer, L. K., Baker, E. N., and Jameson, G. B. (1999) Protein Sci. 8, 75-83. 58. Qin, B. Y., Creamer, L. K., Baker, E. N., and Jameson, G. B. (1998) FEBS Lett. 438, 272–278 59. Ragona, L., Fogolari, F., Catalano, M., Ugolini, R., Zetta, L., and Molinari, H. (2003) J. Biol. Chem. 278, 38840-6. 60. Sayle, R. A., and Milner-White, E. J. (1995) Trends Biochem. Sci. 20, 374. 38.
(50) 61. Sawyer, L., and Kontopidis, G. (2000) Biochim. Biophys. Acta. 1482, 136–148 62. Sawyer, L., Papiz, M. Z., North, A. C. T., and Eliopoulos, E. E. (1985) Biochem. Soc. Trans. 13, 265-266. 63. Tanford, C., Bunville, L. G., and Nozaki, Y. (1959) J. Am. Chem. Soc. 81, 4032-4036 64. Tseng, C. F., Lin, C. C., Huang, H. Y., Liu, H. C., and Mao, S. J. T. (2004) Proteomics 4, 2221-8. 65. Uhrinova, S., Smith, M. H., Jameson, G. B., Uhrin, D., Sawyer, L., and Barlow, P. N. (2000) Biochemistry 39, 3565-74. 66. Wu, S. Y., Perez, M. D., Puyol, P., and Sawyer, L. (1999) J. Biol. Chem. 274, 170-4. 67. Yamauchi, R., Ohinata, K., and Yoshikawa, M. (2003) Peptides 24, 1955-61. 68. Yang, J., Powers, J. R., Clark, S., Dunker, A. K., and Swanson, B. G. (2002) J. Agric. Food Chem. 50, 5207-14. 69. Zommara, M., Toubo, H., Sakono, M., and Imaizumi, K. (1998) Biosci. Biotechnol. Biochem. 62, 710-7.. 39.
(51) .. 40.
(52) 20. 1. A. NH2 livtqtmkgldiqkv agtwyslamaas disl l. d 40 C B a q ptpkleevyv e rlpas wkqllieldg e n 106 G F 100 80 E d 66 D e caqkkiiaekt kip avfki daln enkvlvldtdy k kyllvcmens a e I 140 121 H p COOH ihcqeelqtpn fslrih mplak lakdfkelaed dvept rvlcqcvls q e 60. 160. 119. SH. Figure 1: Amino acid sequence of β-LG. β-LG is consisted of 162 amino acids with nine β-sheet strands (A-I) and one α helix (shadow). There are two disulfide bonds located between strand D and carboxyl-terminus (Cys-66 and Cys-160) and between strands G and H (Cys-106 and Cys-119), while a free buried thio group is at Cys-121.. 41.
(53) EF loop. EF loop. Figure 2: 3D structure of β-LG. Crystal structure of β-LG determined by (6) and created by PyMOL (25) (PDB code 1CJ5), showing that strands A-D form one surface of the barrel while strands E-I form the other. The antigenic site recognized by the mAb is located within strand D (green) following thermal denaturation. The two disulfide linkages are also shown (yellow). Hydrophobic molecules such as retinol and palmate are bound in the pocket of calyx.. 42.
(54) OD at 415nm. 0.40 0.35. 0.25 min. 0.30. 0.5 min 1 min. 0.25. 2 min 4 min. 0.20. 8 min. 0.15. 16 min. 0.10 0.05. 0 0.00 50. 60. 70. 80. 90. 100. Temperature (°C) Figure 3: Immunoreactivity of β-LG in raw milk heated at different temperatures over time. Immunoreactivity was monitored using an ELISA on dry milk heated at various temperatures. The increase in immunoreactivity of β-LG assessed by dry milk specific mAb is correlated to the molten globule state of β-LG with a transition between 70-80 °C.. 43.
(55) 1. M. 2. 3. M 1. 2. 3. LG. SDS-PAGE. Western blot. Figure 4: Effect of tryptic digesition on immunoreactivity of β-LG. About 10 µg of tryptic digested LG were used with time for SDS-PAGE containing mercaptoethanol as a reducing agent (left) followed by a Western blot (right). Lane M: molecular markers. Lane 1: tryptic digestion for 5 min. Lane 2: for 10 min. Lane 3: for 30 min.. 1.4. A. Competitive ELISA. OD at 415 nm. 1.2 1.0 0.8. LG. 0.6. Heated LG. 0.4. Acetyled LG. 0.2. Heated Acetyl-LG. 0 0.0. 0. 0.2. 1. 5. 25. Protein (µg) Figure 5: Effect of acetylation on immunoreactivity of β-LG. Competitive ELISA using heated β-LG as an immobilized antigen, while competing with native, heated, acetylated, and heated acetylated β-LG.. Immunoreactivity of β-LG was. significantly increased upon heating at 100 °C for 5 min, but not of acetylated and heated acetylated β-LG. 44.
(56) M. 1. 2. M 1. 2. LG 9 kDa. SDS-PAGE Western blot Figure 6: Effect of CNBr cleavage on immunoreactivity of β-LG. About 10 µg of native and CNBr cleaved β-LG were used for SDS-PAGE containing mercaptoethanol as a reducing agent (left) followed by a Western blot (right). A major immunoreactive peptide with a molecular weight about 9 kDa was found to be in residues 25-107 as determined by an amino-terminal sequence analysis. In theory, fragments with molecular weight greater than 9 kDa represent those incompletely cleaved peptides (Fig. 1). Lane M: molecular markers. Lane 1: native β-LG. Lane 2: CNBr fragments of β-LG.. 45.
(57) Figure 7: Delineation of an epitope recognized by mAb. Twelve peptides corresponding to a 9 kDa CNBr fragment of β-LG (residues 25-107) were directly synthesized in situ on nitrocellulose membrane. Peptides 11 and 12 were prepared due to the presence of Pro residues thought to be potentially antigenic and Lys thought to be involved per our trypsin and acetylation experiments. The entire peptide array was commercially prepared under a contract for customer designing. Binding of antibody was conducted using HRP-labeled secondary antibody with chemiluminescent agent as a developer. The shed region represents the proposed epitope.. 46.
(58) M. kDa. 1. 2. 3. 4. 106.5 66.4 45 35 25 18.5. LG. 14.4. Figure 8: SDS-PAGE profile of carboxymethylated β-LG (CM-LG). About 10 µg of β-LG and CM-LG were loaded on 15% SDS-PAGE. Lane M: molecular markers. Lanes 1: β-LG. 2: heated β-LG. 3: CM-LG. 4: heated CM-LG. Significant increase in β-LG dimer and high molecular form are seen, while heating at 100 °C for 5 min, but not in CM-LG. 2.5. B. Competitive ELISA OD at 415 nm. 2.0 1.5 LG. 1.0. Heated LG CM-LG. 0.5. Heated CM-LG 0 0.0. 0. 0.2. 1. 5. 25. Protein (µg) Figure 9: immunoreactivity of carboxymethylated β-LG (CM-LG). Competitive ELISA of native, heated, CM, and heated CM β-LG. The plate was immobilized with heated β-LG. 47.
(59) Figure 10: Circular dichroic spectra of native β-LG, heated β-LG, carboxymethylated LG (CM-LG), and synthetic peptide 67-76. Samples at a final concentration of 0.2 mg/ml in 10 mM phosphate buffer, pH 7.4, were used for the study. A typical β-structure of native β-LG is shown at 215 nm. β-LG was heated at 100 °C for 5 min.. 48.
(60) 1.2. OD at 415 nm. 1.0 0.8 0.6 0.4. LG. 0.2. Synthetic peptide 67-76. 0 0.0. 0. 0.2. 1. 5. 25. 100. Protein (µg). Figure 11: Immunoreactivity of the synthetic peptide determined by an ELISA. A synthetic peptide corresponding to residues 67-76 (AQKKIIAEKT) in strand D was prepared as a soluble form. The plate was immobilized with heated β-LG.. 49.
(61) 69. 74. Figure 12: Role of charged residues in antigenic specificity determined by a peptide array. Peptide 1 represents the native sequence 67-75 of β-LG. Substitution of Lys-69 and Glu-74 by Ala or a same charged amino acid, respectively, resulted in a total loss of immunoreactivity (peptide 2-9).. Hydrophobic residues Ile-71 and Ile-71 were essential in maintaining the. antigenic structure, while peptides 11 and 12 retaining all the Lys residues were used as randomized negative controls.. 50.
(62) Figure 13 A: Effect of pH on β-LG binding to mAb. β-LG at pH from 2 to 10.. 51. CD spectra of.
(63) 1.6. OD at 415 nm. B. mAb anti-denatured LG 1.2. 0.8. 0.4. 0 2. 3. 4. 5. 6. 7. 8. 9. 10. pH. Figure 13B: Binding of mAb to β-LG at various pH determined by ELISA. The primary mAb binding was conducted at the pH indicated; all other reactions, including coating, blocking, washing, and secondary antibody binding, were performed at pH 7.4 according to standard procedures. The binding was significantly decreased at pH 8-10. 1.0. OD at 415 nm. C. Goat anti-mouse IgG 0.8 0.6 0.4 0.2 0.0 2. 3. 4. 5. 6. 7. 8. 9. 10. pH. Figure 13C: Control experiment evaluating the effect of pH on antigen-antibody interaction. This experiment was designed to study the pH effect on the binding of antigen and antibody in general. Antigen (mouse IgG) was coated on the plate and allowed the binding of HRP-labeled anti-IgG (goat) at various pH. The plate was then developed after removing unbound antibody using PBS at pH 7.4. The binding was slightly decreased at pH 9-10. 52.
(64) AA.. 20. Retinol binding. 15 10 5 0 -5 0. 22. 44. 66. 88. 10 10. 12. % Change of emission at 332 nm. Emission intensity at 470 nm. 25. 25. CC.. Palmitic acid. 20 15 10 5 0 -5. 0. 2. 4. pH. 8. 10. 12. pH pH 120. Relative enhancement (%). 120 Relative intensity (%). 6. BB. Retinol binding Retinol. 100 80 60 40 20 0. DD. 100. Palmitic acid binding. Palmitic acid. 80 60 40 20 0. native LG. native LG. 80℃. 80℃. 100℃. 100℃. native LG native LG. 80℃ 80℃. 100℃ 100℃. Figure 14: Effect of pH and heating on β-LG binding to retinol and palmitic acid. A: Fluorescence emission for binding of β-LG to retinol was measured at 470 nm with excitation at 287 nm. Binding was determined by the enhancement of extrinsic fluorescence of retinol at 24 °C. B: Effect of β-LG heating (5 min) on retinol binding. C: Fluorescence emission for binding of β-LG to palmitic acid was measured at 332 nm with excitation at 287 nm.. Binding. was determined by the enhancement of intrinsic fluorescence of β-LG at 24 °C. D: Effect of β-LG heating (5 min) on palmitic acid binding.. 53.
(65) Figure 15: Three dimensional view of β-LG complexed with palmitic acid. The diagram is constructed according to the RasMol wireframe model (26), showing the D strand (residues 66-76) and other residues (28-43, 60-65, 84-93, and 107-117) (light wireframe) nearby the calyx of β-LG.. Notably, palmitic acid (PA) is almost. perpendicularly oriented against the D strand, while Lys-69 is capable of interacting with the acidic group of palmate. A hydrophobic interaction between the side chain of Ile-71 and palmate is speculated. EF loop (85-89), which controls the open or close of calyx, is also seen. During the thermal denaturation, the conformational change of strand D results in the exposure of buried side chains of Lys-69 and Ile-71 and thereby attenuates and enhances the binding to palmitic acid and mAb, respectively.. 54.
(66)
數據

+7


相關文件
Mannocarolose, β-glucan, Amino acid nitrogen, Crude protein, Crude ash and
Woven fabrics of cotton, plain weave, containing less than 85% by weight of cotton, mixed mainly or solely with man-made fibres, weighing not more than 200 g/square meter,
Panty hose and tights, knitted or crocheted, of synthetic fibres, measuring per single yarn less than 67
Woven fabrics of cotton, plain weave, containing less than 85% by weight of cotton, mixed mainly or solely with man-made fibres, weighing not more than 200 g/square meter,
Stage l1c* Tumor either stage lla or llb but with tumor on the surface of one or both ovaries; or with capsule(s) ruptured; or with ascites present containing malignant cells or
關於萊克多巴胺下列說明何者錯誤?(A)為乙型受體素 (β 受體促進劑) (B)依台灣政 府訂定的
Woven fabrics of cotton, plain weave, containing less than 85% by weight of cotton, mixed mainly or solely with man-made fibres, weighing not more than 200 g/square meter,
Track suits, knitted or crocheted, of synthetic fibres 002.
