Induction of a distinct CD8 Tnc17 subset by transforming
growth factor-
 and interleukin-6
Shih-Jen Liu,*
,1Jy-Ping Tsai,*
,1Chia-Rui Shen,
†Yuh-Pyng Sher,
‡Chia-Ling Hsieh,
§Yu-Ching Yeh,* Ai-Hsiang Chou,* Shu-Rung Chang,* Kuang-Nan Hsiao,* Feng-Wei Yu,
†and Hsin-Wei Chen*
,2*Vaccine Research and Development Center, National Health Research Institutes, Miaoli, Taiwan,
China;
†Graduate Institute of Medical Biotechnology, Chang Gung University, Tao-Yuan,
Taiwan, China;
‡Center for Molecular Medicine, China Medical University Hospital,
Taichung, Taiwan, China; and
§Graduate Institute of Life Sciences, National Defense Medical Center, Taipei,
Taiwan, China
Abstract:
Cross-talk between TGF-
 and IL-6
has been shown to direct the differentiation of
CD4
ⴙcells into special IL-17-secreting cells,
which are termed Th17 cells. In this study, we
demonstrated that TGF-
 and IL-6 could
stimu-late CD8
ⴙcells to differentiate into
noncyto-toxic, 17-producing cells in MLC. These
IL-17-producing CD8
ⴙcells exhibit a unique
gran-zyme B
–IFN-
␥
–IL-10
–phenotype. The mRNA
level of Th2/T cytotoxic 2 (Tc2) transcription
factors GATA3 and Th1/Tc1 transcription
fac-tors T-box expressed in T cell (T-bet) as well as
its target H2
䡠O-like homeobox (Hlx) is decreased
in CD8
ⴙcells from TGF-
- and IL-6-treated
MLC. In addition, these CD8
ⴙcells display a
marked up-regulation of retinoic acid-related
or-phan receptor-
␥t, a key IL-17 transcription
fac-tor. These results demonstrate that the existence
of an IL-17-producing CD8
ⴙsubset belongs to
neither the Tc1 nor the Tc2 subset and can be
categorized as a T noncytotoxic 17 (Tnc17)
subset. J. Leukoc. Biol. 82: 354 –360; 2007.
Key Words:
IL-17
䡠mixed lymphocyte culture
䡠cytotoxic T
lym-phocyte
INTRODUCTION
Cytokines play key roles in regulating the development of
immune effector cells and possess direct effector functions
in fighting diseases. TGF-
 has been found at the site of
most tumors [1], and it inhibits the proliferation and
func-tional differentiation of T lymphocytes [2, 3] and other
immune cells [4, 5]. Further, TGF-
 production by tumor
cells prevented activation of CTL function [3]. This effect
was presumably a result of the inhibition by TGF-
 of the
expression and function of IL-2 and IL-2Rs [6, 7] and
cytolytic gene products [8], or it is the result of inducing T
regulatory cells (Tregs) [9 –13]. It has also been
demon-strated that TGF-
 plays a synergistic role with IL-10 to
polarize tumor-infiltrating lymphocytes to predominantly Th2/T
cytotoxic 2 (Tc2) phenotypes [14]. As CTL and Th1-associated
cytokine production are important for achieving effective,
im-mune-mediated tumor eradication, suppression of these functions
by TGF-
 would effectively subvert a proper immune response.
IL-6 may also play a pivotal role in cancer development.
It is known that IL-6 can be a differentiation inducer in lung
adenocarcinoma cells [5, 6] and other tumors [7, 15]. It is
not clear whether IL-6 is secreted by cancer cells or by the
immune system in response to the tumor or both. However,
several recent reports have highlighted the nature of
cross-talk between IL-6 and TGF-
 [16, 17]. IL-6 was shown to be
able to antagonize tumor-derived TGF-
 and restore
lym-phokine-activated killing activity [9] and plays a key role in
T cell activation by overcoming the suppressive effect of
CD4
⫹CD25
⫹Tregs [17], indicating that IL-6 may interfere
with TGF-
 in initiating the induction of CD4
⫹CD25
⫹Tregs. During the past year, a new subset of CD4
⫹cells,
Th17, has been identified and is characterized by
produc-tion of IL-17. Th17 cell differentiaproduc-tion is initiated by TGF-

and IL-6 [18 –20]. It has shown recently that the retinoic
acid-related orphan receptor (ROR)
␥t is the key
transcrip-tion factor that orchestrates the differentiatranscrip-tion of Th17 [21].
However, the role and effects of TGF-
 and IL-6 on CD8
⫹cell differentiation remain relatively undefined. We have
found recently that high levels of TGF-
 and IL-6 are
present in malignant effusion of cancer patients [22]. We
hypothesize that the cross-talk between TGF-
 and IL-6
may modulate the CD8
⫹CTL to kill tumor cells.
In this study, we showed that IL-6 acted cooperatively with
TGF-
 to elicit a high frequency of IL-17-secreting CD8
⫹cells
with a noncytotoxic phenotype. IFN-
␥ as well as IL-10 was not
expressed in these IL-17-secreting CD8
⫹cells. It leads to the
fact that IL-17-producing CD8
⫹cells may represent a distinct
subset of CD8
⫹cells, T noncytotoxic 17 (Tnc17).
1These authors contributed equally to this work.
2Correspondence: Vaccine Research and Development Center, National
Health Research Institutes, No. 35, Keyan Road, Zhunan Town, Miaoli County 350, Taiwan, China. E-mail: [email protected]
Received February 13, 2007; revised March 30, 2007; accepted April 27, 2007.
MATERIALS AND METHODS
Mice and cell lines
Female, 6- to 8-week-old BALB/c (H-2d) and C57BL/6 (H-2b) mice were
purchased from the National Laboratory Animal Breeding and Research Center (Taipei, Taiwan, ROC). All mice were housed at the Laboratory Animal Center of the National Health Research Institutes (NHRI; Taiwan, ROC). All of the animal studies were approved by the Animal Committee of the NHRI and performed according to their guidelines. The cell lines used in the study include the thymoma EL-4 of H-2band plasmacytoma P815 of H-2d.
MLC
Splenocytes were obtained from BALB/c (H-2d) and C57BL/6 (H-2b) mice, and
single cell suspensions were made. BALB/c splenocytes were used as respond-ers (R), and 2000 rads X-irradiated C57BL/6 splenocytes were used as stimulators (S) at a R:S ratio of 3:1. Cells were resuspended at 3.3⫻ 106
cells/ml in RPMI-1640 medium containing 5% FBS, penicillin and strepto-mycin, gentamicin, HEPES, and 2-ME. Cells were seeded in 24-well tissue-culture plates at 2 mL per well. Recombinant TGF- (1 ng/mL) and/or IL-6 (100 ng/mL; R&D Systems, Minneapolis, MN, USA) were added in parallel cultures as indicated. After 4 –5 days of culture, proliferation responses, cell-mediated cytotoxicity, and cytokine profiles were determined.
Determination of proliferation responses
The cell density of BALB/c (H-2d) splenocytes was adjusted to 3⫻ 107/mL
and labeled with 10 uM CellTracker Green 5-chloromethylfluorescein diac-etate (CMFDA; Molecular Probes, Eugene, OR, USA) at 37°C for 10 min. Cells were washed with culture medium once and used as responders. The MLC was set as described above. Recombinant TGF- (1 ng/mL) and/or IL-6 (100 ng/mL) were added as indicated. After 4 days of culture, the cells were harvested for staining. Nonspecific binding was blocked by incubation with rat anti-mouse CD16/CD32 antibody (BD Bioscience, San Jose, CA, USA) in PBS for 10 min at 4°C. Cells were stained with anti-CD8 antibodies conjugated with PE (BD Biosciences). After washing, cells were acquired and analyzed on a FACSCalibur flow cytometer with CellQuest software. Proliferative CD8⫹cells were determined by fluorescence intensity of CMFDA dilution.
Determination of cytolytic activity
EL-4 (H-2b) or P815 (H-2d) was labeled with 0.5M CMFDA at 37°C for 20
min. Cells were washed with culture medium once and used as target cells. Effector cells were prepared from H-2d(BALB/c) against H-2b(C57/BL6) MLC
on Day 5. Target cells were cocultured with effector cells at the indicated E:T ratios. After 1.5 h incubation at 37°C, cells were stained with Annexin V conjugated with PE (R&D Systems) according to the manufacturer’s instruc-tions. CMFDA-positive cells (10,000) were harvested and analyzed on a FACSCalibur flow cytometer with CellQuest software. Cytolytic activities were determined by the percentages of Annexin V-positive cells within the CMFDA-positive gate. Less than 5% of target cells were Annexin V-CMFDA-positive when cultured with medium only in each experiment.
Intracellular staining
The MLC was established as described above. On Day 5, cells were restimu-lated with anti-CD3 antibody (2C11) and Brefeldin A (eBiosciences, San Diego, CA, USA) for 4 h. Cells were first stained with PE-Cy5-conjugated anti-CD8 antibody (eBiosciences) and then treated with fixation and perme-abilization buffer (eBiosciences) according to the manufacturer’s directions. Intracellular staining was performed using FITC- or PE-conjugated antibodies to IFN-␥, IL-10, and Granzyme B (eBiosciences). The PE-conjugated IL-17 antibody was purchased from BD Biosciences. Isotype-matched anti-bodies conjugated with FITC or PE were used as negative controls. CD8⫹cells were further gated for evaluation of cytokine expression profiles.
Real-time quantitative PCR (qPCR) analysis of
gene expression in CD8
⫹cells
The MLC was set as described above and supplemented with or without recombinant TGF- (1 ng/mL) and IL-6 (100 ng/mL). After 4 days of culture,
CD8⫹ cells were purified using anti-CD8 magnetic microbeads and MACS columns (Miltenyi Biotec, Bergishc Galdbach, Germany) according to the manufacturer’s instruction (purity was⬎95%). Total RNA of isolated cells was extracted using the RNeasy mini kit (Qiagen, Valencia, CA, USA) following the manufacturer’s instructions. RNA (0.5–1g) was reverse-transcribed to cDNA with an oligo-dT primer in a 20-l vol by using SuperScript III RT (Invitrogen, Carlsbad, CA, USA). The mouse Universal Probe Library (UPL) set (Roche, Mannheim, Germany) was used to perform the real-time qPCR assay for gene expression in isolated cell populations. The specific primers and UPL number were as follows: hypoxanthine guanine phosphoribosyl transferase (HPRT), 5⬘-ggagcggtagcacctcct-3⬘ (forward) and 5⬘-ctggttcatcatcgctaatcac-3⬘ (reverse) with UPL#69; T-box expressed in T cell (T-bet), 5⬘-tcaaccagcaccagacagag-3⬘ (forward) and 5⬘-aaacatcctgtaatggcttgtg-3⬘ (reverse) with UPL#19; H2䡠O-like homeobox (Hlx), 5⬘-aagccagaccgaaagcag-3⬘ (forward) and 5⬘-tgcgcctcctta-gagtgc-3⬘ (reverse) with UPL#88; GATA3, 5⬘-cttatcaagcccaagcgaag-3⬘ (for-ward) and 5⬘-cccattagcgttcctcctc-3⬘ (reverse) with UPL#77; ROR␥t, 5⬘-ttcac-cccacctccactg-3⬘ (forward) and 5⬘-caagggatcacttcaatttgtg-3⬘ (reverse) with UPL#56. The reaction mixture contained 5 ng cDNA, 0.2M primers, and LightCycler 480 Probe Master (Roche) and was performed in a LightCycler 480 system (Roche). All qPCRs were carried out with an initial denaturation at 95°C for 10 min, followed by 45 cycles of 95°C for 10 s, 60°C for 20 s, and 72°C for 2 s. Target gene expression was calculated using the comparative method for relative quantity upon normalization to HPRT gene expression.
Statistical analysis
The statistical significance of differential findings between experimental groups was determined by an unpaired Student’s t-test. Data were considered statistically significant if Pⱕ 0.05.
RESULTS
IL-6 abrogated the inhibition by TGF-
 of CD8
⫹cell proliferation in MLC
A murine MLC model was used in this study to investigate the
role of TGF-
 and IL-6 in regulating the CD8-mediated
im-mune responses. Cell proliferation was analyzed by
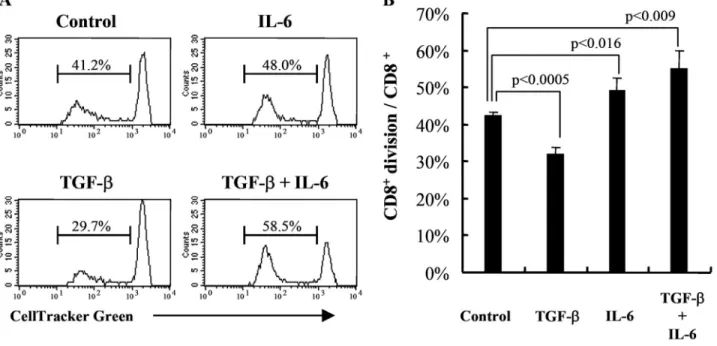
determin-ing the CMFDA intensity. Results are shown in Figure 1.
Approximately 42% of total CD8
⫹cells showed diluted
CM-FDA in unmodified MLC (control-MLC). However, in the
pres-ence of TGF-
 (TGF--MLC), the percentages of divided
CD8
⫹cells in total CD8
⫹cells were reduced significantly to
32% (P
⬍0.0005, compared with control-MLC). In contrast,
when the MLC was supplemented with IL-6 (IL-6-MLC), the
percentage of proliferating cells was elevated significantly to
49% (P
⬍0.016, compared with control-MLC) of the whole
CD8
⫹population. It is most interesting that the presence of
IL-6 plus TGF-
 (TI-MLC) strongly stimulated CD8
⫹cell
proliferation. Close to 55% of CD8
⫹cells showed diluted
CMFDA (P
⬍0.009, compared with control-MLC).
IL-6 was unable to restore the cytotoxic
response inhibited by TGF-

The above result demonstrated that IL-6 was able to abrogate
the suppressive effect of TGF-
 on CD8
⫹cell proliferation.
Here, we investigated whether the cytotoxic response was
modulated by TGF-
 and/or IL-6. The effect of TGF- and
IL-6 on allo-specific CTL activity is shown in Figure 2.
Specific CTL activity in MLC was obtained against the
haplo-type-matched, allogeneic target cells (EL4 of H-2
b) and not for
the syngeneic target cells (P815 of H-2
d). In TGF-
-MLC, the
cytotoxic activity was abolished; in contrast, the killing activity
was enhanced in IL-6-MLC. However, IL-6 was unable to
restore the allo-specific cytolytic activity fully in the TI-MLC.
The experiments have been repeated three times, and the
results were reproducible. These results were quite different
from the proliferation data.
TGF-
 and IL-6 induce IL-17-producing CD8
⫹cells
The CD8
⫹cells in TI-MLC showed the highest level of
pro-liferative ability but had a low level of cytotoxicity (Figs. 1 and
2). However, CD8
⫹cells in MLC were shown to play the major
role of performing cytotoxic activity (data not shown). These
results indicate that the effect of TGF-
 and IL-6 on
prolifer-ation of CD8
⫹cells is different from that on the cytotoxicity of
CD8
⫹cells. To address this issue, the cytokine production
profile of CD8
⫹cells was evaluated by analyzing the
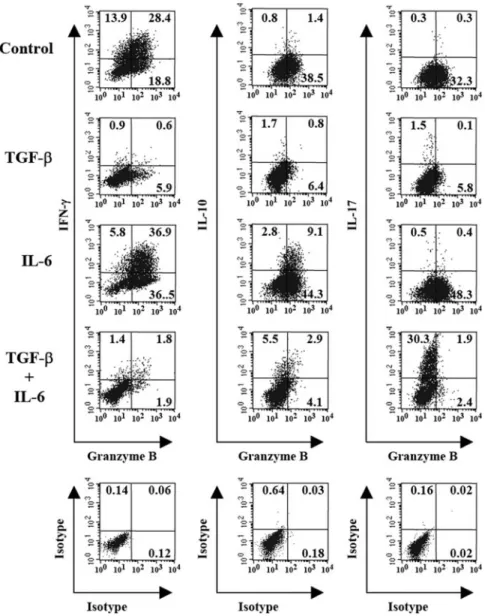
intracel-lular staining. As shown in Figure 3,
⬃44% and 32% of
CD8
⫹cells contributed to IFN-
␥ production in control- and
IL-6-MLC, respectively. TGF-
 induced profound suppression
of IFN-
␥-secreting CD8
⫹cells. Less than 3% of CD8
⫹cells
expressed IFN-
␥ in TGF-- and TI-MLC. Approximately 10%
of CD8
⫹T cells were able to secrete IL-10 in IL-6-MLC. The
frequency of IL-10-secreting CD8
⫹T cells decreased slightly
to 6% in TI-MLC. No significant numbers of IL-10-producing
Fig. 1. IL-6 restores the inhibitory effect of TGF- on CD8⫹cell proliferation successfully. MLC, consisting of 2000 rads X-irradiated C57BL/6 splenocytes and BALB/c splenocytes, prelabeled with CMFDA, were established with the addition of TGF- (1 ng/mL) and/or IL-6 (100 ng/mL) as indicated. (A) CD8⫹cell proliferation was assessed by determining CMFDA dilution in cells positively stained for CD8 on Day 4. A representative experiment is shown. (B) The percentages of CD8⫹division over total CD8⫹cells were plotted. The means withSDfrom three independent experiments are shown.Fig. 2. Modulating effects of IL-6 and TGF- on allocytotoxicity. BALB/c (H-2d) against C57BL/6 (H-2b) MLC was established with the addition of TGF- (1
ng/mL) and/or IL-6 (100 ng/mL) as mentioned. Cultured cells were harvested on Day 5 as effector cells and incubated with CMFDA-labeled target cells EL4 (H-2b)
or P815 (H-2d) for 1.5 h at E:T ratios at 20:1 or 5:1. The cytotoxicity was then determined by labeling cells with PE-conjugated Annexin V, and the CMFDA-positive
CD8
⫹cells were detected in control- and TGF-
-MLC. It is
striking that more than 30% of CD8
⫹T cells expressed IL-17
in TI-MLC, and low percentages of the CD8
⫹cell population
expressed IL-17 in control-, TGF-
-, and IL-6-MLC. All
ex-periments have been repeated three times, and the results were
reproducible.
Distinct IL-17
⫹granzyme B
–and IL-10
⫹granzyme
B
–CD8
⫹T cells were induced by TGF-

and IL-6
Granzyme B is known to play a pivotal role in CTL function to
eliminate virus-infected cells or tumor cells. Next, we have
further assessed whether the expression of granzyme B in these
IL-17-secreting CD8 T cells was shown to bear good
prolifer-ative but poor cytotoxic ability. From Figure 4, we found that
in control- and IL-6-MLC, 28% and 37% of CD8
⫹cells were
IFN-
␥
⫹granzyme B
⫹phenotypes, respectively. However, less
than 2% of CD8
⫹cells had an IFN-
␥
⫹granzyme B
⫹phenotype
in TGF-
- and TI-MLC. Regarding CD8
⫹cells with the
IL-10
⫹granzyme B
⫹phenotype, only in IL-6- and TI-MLC, there
were significant numbers of CD8
⫹cells to produce IL-10.
However, the IL-6-MLC produced a higher frequency of the
IL-10
⫹granzyme B
⫹phenotype in CD8
⫹cells than the
TI-MLC (9.1% vs. 2.9%). Few IL-17
⫹granzyme B
⫹CD8
⫹cells
could be found in control-, TGF-
-, and IL-6-MLC. Less than
2% of CD8
⫹cells were the IL-17
⫹granzyme B
⫹phenotype in
TI-MLC. Therefore, it appears that the majority of
IL-17-producing CD8
⫹cells does not express granzyme B. The
experiments were performed three times, and the results were
reproducible. In fact, granzyme B-expressing CD8
⫹cells are
diminished by the addition of TGF-
. In contrast, IL-6
en-hances the expression of granzyme B in CD8 cells. However,
IL-6 was unable to reverse the suppressive effect of TGF-
 on
granzyme B production. It can be concluded that the profile of
granzyme B expression in CD8
⫹cells correlated with the
results of their cytotoxic activity as shown in Figure 2.
To verify whether IL-17-expressing CD8
⫹cells in TI-MLC
expressed IFN-
␥ or IL-10, several experiments were set up,
and the results are shown in Figure 5. More than 26% of
IL-17-expressing CD8
⫹cells do not produce IFN-
␥ or IL-10.
There were merely 6% IL-17
–IFN-
␥
⫹and 3% IL-17
–IL-10
⫹phenotype CD8
⫹cells. Less than 2% of CD8
⫹cells were the
IL-17
⫹IFN-
␥
⫹or IL-17
⫹IL-10
⫹phenotype. The experiments
have been repeated twice, and the results were reproducible.
These results demonstrate that most of IL-17-secreting CD8
⫹cells were the IFN-
␥
–IL-10
–phenotype.
Moreover, we have also examined the mRNA expression
levels of cytokines and transcription factors of CD8
⫹cells in
TI-MLC by real-time PCR. The expression of the IFN-
␥ mRNA
level in TI-MLC was sevenfold less than control-MLC. By
contrast, IL-10 and IL-17 mRNA expression was 80- and
507-fold, respectively, higher than control-MLC (data not
shown). These profiles were in agreement with the intracellular
cytokine data (Fig. 3). Finally, analysis of the expression of
Th1/Th2 signature transcription factors showed a substantial
down-regulation of T-bet, Hlx, and GATA3 mRNA (Fig. 6,
A–C
).
It has been demonstrated recently that ROR
␥t is a key
transcription factor directing the differentiation of Th17 [21]. It
is notable that the mRNA expression level of ROR
␥t was
increased 30 fold (Fig. 6D). Taken together, our data suggest
that the IL-17-producing CD8
⫹cells represent a distinct T
effector lineage, neither Tc1 nor Tc2.
DISCUSSION
Tumor growth and survival are affected by the cytokines
present in the tumor microenvironment. We reported that a
variety of cytokines was found at elevated levels in the
malig-nant effusions of cancer patients [22]. The most prominent
feature was the presence of high levels of TGF-
 and IL-6 in
all the effusion samples tested [22]. Recent studies have
dem-onstrated that TGF-
 and IL-6 promote the development of
IL-17-producing CD4
⫹cells, which have been called Th17
[20]. IL-17 has been shown to increase the IL-6 production
[23–26]. Therefore, in such, the environmental condition will
facilitate IL-17 production. Expression of IL-17 mRNA was
detected at the tumor sites, including cervical carcinoma [27]
and ovarian cancer [28]. However, the effect of IL-17 on tumor
growth is paradoxical. Benchetrit et al. [29] reported that IL-17
is able to inhibit tumor growth by means of a T cell-dependent
manner. By contrast, Tartour et al. [27] and others [30, 31]
demonstrated that IL-17 promotes tumor growth via
potentia-tion of tumor angiogenesis. Our results also demonstrated that
Fig. 3. IFN-␥, IL-10, and IL-17 expression profiles of CD8⫹T cells inIL-6-and/or TGF--treated MLC, which when consisting of C57BL/6 and BALB/c splenocytes, were established with the addition of TGF- (1 ng/mL) and/or IL-6 (100 ng/mL) as mentioned. The cultures were harvested after 5 days and stained with anti-CD8 followed by intracellular staining for IFN-␥, IL-10, or IL-17. CD8⫹cells were gated, and expression of cytokine profiles was plotted. The numbers in the histograms indicate the percentage of positive cells. Data shown are representative of three independent experiments.
IL-17-producing CD8
⫹cells, Tnc17, are deficient in cytolytic
activity (Figs. 2 and 4). These findings provide the possible
mechanism that tumor-infiltrating T cells regulate angiogenesis
via elaboration of IL-17 when TGF-
 and IL-6 are present at
the tumor microenvironment. The consequence is tumor
growth.
TGF-
 is a pleiotropic cytokine with multiple regulatory
functions in the immune system [32]. One regulatory function
of TGF-
 is the inhibition of naı¨ve T cell differentiation into
effector cells. Our initial experiments supported the notion that
TGF-
 inhibited CD8
⫹cell proliferation (Fig. 1) and
allo-specific cytotoxic activity (Fig. 2) in MLC. IL-6 was found to
play an opposing role to TGF-
 in several biological functions
[7, 16, 33]. In contrast to the suppressive effect of TGF-
 on
CD8
⫹cell proliferation and cytotoxicity (TGF-
-MLC), IL-6
enhanced CD8
⫹cell division and allo-specific cytotoxicity,
and the proliferative potential of the CD8
⫹cell was enhanced
further in TI-MLC when TGF-
 and IL-6 are present. However,
the generation of CD8
⫹CTL remained to be suppressed in
TI-MLC (Figs. 1 and 2). To confirm that the cytotoxic response
was restricted to CD8
⫹cells, anti-CD4, anti-CD8, and isotype
control antibodies were applied in the cytotoxic assay. The
cytotoxicity remained unchanged by adding anti-CD4 or
iso-type control antibodies but was abrogated completely by
anti-CD8 antibody (data not shown). These results indicate that
Fig. 4. Granzyme B is expressed by IFN-␥ and IL-10but not IL-17-producing CD8⫹ T cells in TGF- -and/or IL-6-supplemented MLC. TGF- (1 ng/mL) and/or IL-6 (100 ng/mL) were added to MLC as de-scribed above. Cells were harvested on Day 5 and stained with anti-CD8. Intracellular staining for gran-zyme B and IFN-␥, IL-10, or IL-17 was performed. CD8 cells were gated and displayed as granzyme B versus IFN-␥, IL-10, or IL-17. The isotype controls were shown in the bottom panel. Numbers in quadrants indicate percent-positive cells. Data shown are repre-sentative of three independent experiments.
Fig. 5. IL-17-producing CD8⫹T cells did not express IFN-␥ or IL-10 in IL-6-and TGF--treated MLC. BALB/c (H-2d) against C57BL/6 (H-2b) MLC were
established in the presence of TGF- (1 ng/mL) and IL-6 (100 ng/mL). After 5 days in culture, cells were harvested and stained with anti-CD8. Intracellular staining for IL-17 with IFN-␥ or IL-10 was performed. CD8⫹cells were gated and plotted as IL-17 versus IFN-␥ or IL-10. Numbers in quadrants indicate percent-positive cells in each quadrant. Data shown are representative of two independent experiments.
cytotoxic activity was mediated mainly by the CD8
⫹subset.
Thus, there is discordance between the proliferative potential
and the generation of CTL of the CD8
⫹cells in TI-MLC. These
findings uncover a novel role of the cross-regulation by TGF-

and IL-6 of the activation of CD8
⫹cells.
Several lines of evidence have indicated that T cells
mod-ulate immune responses by producing Type 1 or 2 cytokines in
vivo [34 –39]. After activation, CD4
⫹and CD8
⫹cells
differ-entiate into effector cells, which are specialized in terms of the
cytokines that they produce. Naı¨ve CD8
⫹cells are able to
differentiate into a Tc1 subset, which secretes IFN-
␥ and IL-2
predominately, and a Tc2 subset, which secretes IL-4, IL-5,
and IL-10 preferentially [40 – 42]. We examine the functional
characteristics of CD8
⫹cells in MLC by investigating their
cytokine and granzyme B expression. There were substantial
numbers of IFN-
␥-producing CD8
⫹cells in control-MLC and
IL-6-MLC (Fig. 3). Most of the IFN-
␥
⫹CD8
⫹cells also
ex-pressed high levels of granzyme B. In control-MLC, 67% of
IFN-
␥
⫹CD8
⫹cells were granzyme B-positive cells. More than
86% of IFN-
␥
⫹CD8
⫹cells were granzyme B-positive in
IL-6-MLC (Fig. 4). Naı¨ve CD8
⫹cells were blocked by TGF-
 from
differentiating into Tc1 cells. Significant numbers of
IL-10-secreting CD8
⫹cells were obtained in IL-6-MLC and TI-MLC
(Fig. 3). In IL-6-MLC,
⬃77% of IL-10
⫹CD8
⫹cells were the
granzyme B
highphenotype. Less than 35% of IL-10
⫹CD8
⫹cells expressed high levels of granzyme B when TGF-
 was
also present (Fig. 4). These results indicate that IL-6 skews
naı¨ve CD8
⫹cells toward the IL-10
⫹phenotype. In the
pres-ence of TGF-
 and IL-6, expression of granzyme B is inhibited
in the majority of IL-10
⫹CD8
⫹cells.
It has been shown that TGF-
 and IL-6 facilitate Th17
differentiation in CD4
⫹cells [20]. We examined further
whether CD8
⫹cells would produce IL-17 in the presence of
these two cytokines. Indeed, IL-17-producing CD8
⫹cells were
induced in TI-MLC but not in control-, TGF-
-, or IL-6-MLC
(Fig. 3). Furthermore, up to 94% of IL-17
⫹CD8
⫹cells were the
granzyme B
lowphenotype (Fig. 4). These observations explain
clearly why proliferation of CD8
⫹cells is enhanced in the
presence of IL-6 and TGF-
, but they lack the cytolytic
activity.
Like other differential processes, the naı¨ve T cell
develop-ment is mediated by lineage-specific transcription
mecha-nisms. It was reported recently that ROR
␥t plays the key role
in Th17 differentiation [21], which resembles the roles of T-bet
[43], Hlx [44], and GATA3 [45, 46] in the development of Th1
and Th2 cells. Our data indicate that CD8
⫹cells from TI-MLC
down-regulate expression of GATA3 and T-bet, as well as its
downstream Hlx (Fig. 6, A–C). Conversely, expression of
ROR
␥t is elevated significantly (Fig. 6D). We also showed that
most of the IL-17
⫹CD8
⫹cells do not produce IFN-
␥ or IL-10
(Fig. 5). These data indicate clearly that differentiation of naı¨ve
CD8
⫹cells into IL-17-producing cells, Tnc17, is independent
of Tc1 and Tc2 cell-development programs. In this study, we
provide compelling evidence to show that a previously
unde-fined, IL-17-expressing CD8 subset is elicited in the presence
of TGF-
 and IL-6. The precise role of Tnc17 in tumor
development will be clarified in our further studies.
ACKNOWLEDGMENTS
This study was supported partly by a grant (VC-094-PP-02)
from the NHRI and grants CMRP1225 and 140041 from Chang
Gung Memorial Hospital, Taiwan. We thank Dr. Chou-Chik
Ting for critical review of the manuscript.
REFERENCES
1. Pasche, B. (2001) Role of transforming growth factor in cancer. J. Cell.
Physiol. 186, 153–168.
2. Cheng, M-L., Chen, H-W., Tsai, J-P., Lee, Y-P., Shih, Y-C., Chang, C-M., Ting, C-C. (2006) Clonal restriction of the expansion of antigen-specific CD8⫹memory T cells by transforming growth factor-. J. Leukoc. Biol.
79,1033–1042.
3. Gorelik, L., Constant, S., Flavell, R. A. (2002) Mechanism of transforming growth factor-induced inhibition of T helper type 1 differentiation. J.
Exp. Med. 195, 1499 –1505.
Fig. 6. IL-17-producing CD8⫹cells are distinct from Th1 and Th2 subsets. MLC was established in the presence or absence of TGF- (1 ng/mL) and IL-6 (100 ng/mL). CD8⫹cells were purified after 4 days of culture. The mRNA levels of (A) T-bet, (B) Hlx, (C) GATA3, and (D) ROR␥t were determined by real-time qPCR.
4. Gorelik, L., Flavell, R. A. (2001) Immune-mediated eradication of tumors through the blockade of transforming growth factor- signaling in T cells.
Nat. Med. 7, 1118 –1122.
5. McCormick, C., Freshney, R. I. (2000) Activity of growth factors in the IL-6 group in the differentiation of human lung adenocarcinoma. Br. J.
Cancer 82, 881– 890.
6. McCormick, C., Freshney, R. I., Speirs, V. (1995) Activity of interferon␣, interleukin 6 and insulin in the regulation of differentiation in A549 alveolar carcinoma cells. Br. J. Cancer 71, 232–239.
7. Walia, B., Wang, L., Merlin, D., Sitaraman, S. V. (2003) TGF- down-regulates IL-6 signaling in intestinal epithelial cells: critical role of SMAD-2. FASEB J. 17, 2130 –2132.
8. Thomas, D. A., Massague, J. (2005) TGF- directly targets cytotoxic T cell functions during tumor evasion of immune surveillance. Cancer Cell 8, 369 –380.
9. Chen, W., Jin, W., Hardegen, N., Lei, K-j., Li, L., Marinos, N., McGrady, G., Wahl, S. M. (2003) Conversion of peripheral CD4⫹CD25–naive T cells
to CD4⫹CD25⫹regulatory T cells by TGF- induction of transcription factor Foxp3. J. Exp. Med. 198, 1875–1886.
10. Fantini, M. C., Becker, C., Monteleone, G., Pallone, F., Galle, P. R., Neurath, M. F. (2004) Cutting edge: TGF- induces a regulatory pheno-type in CD4⫹CD25–T cells through Foxp3 induction and down-regulation
of Smad7. J. Immunol. 172, 5149 –5153.
11. Park, H-B., Paik, D-J., Jang, E., Hong, S., Youn, J. (2004) Acquisition of anergic and suppressive activities in transforming growth factor- -co-stimulated CD4⫹CD25–T cells. Int. Immunol. 16, 1203–1213.
12. Schramm, C., Huber, S., Protschka, M., Czochra, P., Burg, J., Schmitt, E., Lohse, A. W., Galle, P. R., Blessing, M. (2004) TGF regulates the CD4⫹CD25⫹T-cell pool and the expression of Foxp3 in vivo. Int.
Immu-nol. 16, 1241–1249.
13. Zheng, S. G., Wang, J. H., Gray, J. D., Soucier, H., Horwitz, D. A. (2004) Natural and induced CD4⫹CD25⫹ cells educate CD4⫹CD25–cells to
develop suppressive activity: the role of IL-2, TGF-, and IL-10. J.
Im-munol. 172, 5213–5221.
14. Sheu, B-C., Lin, R-H., Lien, H-C., Ho, H-N., Hsu, S-M., Huang, S-C. (2001) Predominant Th2/Tc2 polarity of tumor-infiltrating lymphocytes in human cervical cancer. J. Immunol. 167, 2972–2978.
15. Zhang, X. L., Topley, N., Ito, T., Phillips, A. (2005) Interleukin-6 regu-lation of transforming growth factor (TGF)- receptor compartmentaliza-tion and turnover enhances TGF-1 signaling. J. Biol. Chem. 280, 12239 –12245.
16. Hsiao, Y-W., Liao, K-W., Hung, S-W., Chu, R-M. (2004) Tumor-infiltrat-ing lymphocyte secretion of IL-6 antagonizes tumor-derived TGF-1 and restores the lymphokine-activated killing activity. J. Immunol. 172, 1508 –1514.
17. Pasare, C., Medzhitov, R. (2003) Toll pathway-dependent blockade of CD4⫹CD25⫹ T cell-mediated suppression by dendritic cells. Science
299,1033–1036.
18. Bettelli, E., Carrier, Y., Gao, W., Korn, T., Strom, T. B., Oukka, M., Weiner, H. L., Kuchroo, V. K. (2006) Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells.
Nature 441, 235–238.
19. Mangan, P. R., Harrington, L. E., O’Quinn, D. B., Helms, W. S., Bullard, D. C., Elson, C. O., Hatton, R. D., Wahl, S. M., Schoeb, T. R., Weaver, C. T. (2006) Transforming growth factor- induces development of the TH17 lineage. Nature 441, 231–234.
20. Veldhoen, M., Hocking, R. J., Atkins, C. J., Locksley, R. M., Stockinger, B. (2006) TGF in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179 – 189.
21. Ivanov, I. I., McKenzie, B. S., Zhou, L., Tadokoro, C. E., Lepelley, A., Lafaille, J. J., Cua, D. J., Littman, D. R. (2006) The orphan nuclear receptor ROR␥t directs the differentiation program of proinflammatory IL-17⫹T helper cells. Cell 126, 1121–1133.
22. Tsai, J-P., Chen, H-W., Cheng, M-L., Liu, H-K., Lee, Y-P., Ling Hsieh, C., Luh, K-T., Wu, C-W., Hsu, L-H., Chao, T-Y. (2005) Analysis of host versus tumor interaction in cancer patients: opposing role of transforming growth factor-1 and interleukin-6 in the development of in situ tumor immunity. Immunobiology 210, 661– 671.
23. Chabaud, M., Fossiez, F., Taupin, J-L., Miossec, P. (1998) Enhancing effect of IL-17 on IL-1-induced IL-6 and leukemia inhibitory factor production by rheumatoid arthritis synoviocytes and its regulation by Th2 cytokines. J. Immunol. 161, 409 – 414.
24. Fossiez, F., Djossou, O., Chomarat, P., Flores-Romo, L., Ait-Yahia, S., Maat, C., Pin, J. J., Garrone, P., Garcia, E., Saeland, S., Blanchard, D., Gaillard, C., Das Mahapatra, B., Rouvier, E., Golstein, P., Banchereau, J., Lebecque, S. (1996) T cell interleukin-17 induces stromal cells to produce
proinflammatory and hematopoietic cytokines. J. Exp. Med. 183, 2593– 2603.
25. Jovanovic, D. V., Di Battista, J. A., Martel-Pelletier, J., Jolicoeur, F. C., He, Y., Zhang, M., Mineau, F., Pelletier, J-P. (1998) IL-17 stimulates the production and expression of proinflammatory cytokines, IL- and TNF-␣, by human macrophages. J. Immunol. 160, 3513–3521.
26. Shalom-Barak, T., Quach, J., Lotz, M. (1998) Interleukin-17-induced gene expression in articular chondrocytes is associated with activation of mi-togen-activated protein kinases and NF- B. J. Biol. Chem. 273, 27467– 27473.
27. Tartour, E., Fossiez, F., Joyeux, I., Galinha, A., Gey, A., Claret, E., Sastre-Garau, X., Couturier, J., Mosseri, V., Vives, V., Banchereau, J., Fridman, W. H., Wijdenes, J., Lebecque, S., Sautees-Fridman, C. (1999) Interleukin 17, a T-cell-derived cytokine, promotes tumorigenicity of human cervical tumors in nude mice. Cancer Res. 59, 3698 –3704. 28. Kato, T., Furumoto, H., Ogura, T., Onishi, Y., Irahara, M., Yamano, S.,
Kamada, M., Aono, T. (2001) Expression of IL-17 mRNA in ovarian cancer. Biochem. Biophys. Res. Commun. 282, 735–738.
29. Benchetrit, F., Ciree, A., Vives, V., Warnier, G., Gey, A., Sautes-Fridman, C., Fossiez, F., Haicheur, N., Fridman, W. H., Tartour, E. (2002) Inter-leukin-17 inhibits tumor cell growth by means of a T-cell-dependent mechanism. Blood 99, 2114 –2121.
30. Numasaki, M., Fukushi, J-i., Ono, M., Narula, S. K., Zavodny, P. J., Kudo, T., Robbins, P. D., Tahara, H., Lotze, M. T. (2003) Interleukin-17 pro-motes angiogenesis and tumor growth. Blood 101, 2620 –2627. 31. Numasaki, M., Watanabe, M., Suzuki, T., Takahashi, H., Nakamura, A.,
McAllister, F., Hishinuma, T., Goto, J., Lotze, M. T., Kolls, J. K., Sasaki, H. (2005) IL-17 enhances the net angiogenic activity and in vivo growth of human non-small cell lung cancer in SCID mice through promoting CXCR-2-dependent angiogenesis. J. Immunol. 175, 6177– 6189. 32. Letterio, J. J., Roberts, A. B. (1998) Regulation of immune responses by
TGF-. Annu. Rev. Immunol. 16, 137–161.
33. Ohta, K., Yamagami, S., Taylor, A. W., Streilein, J. W. (2000) IL-6 antagonizes TGF- and abolishes immune privilege in eyes with endo-toxin-induced uveitis. Invest. Ophthalmol. Vis. Sci. 41, 2591–2599. 34. Carter, L. L., Dutton, R. W. (1995) Relative perforin- and Fas-mediated
lysis in T1 and T2 CD8 effector populations. J. Immunol. 155, 1028 – 1031.
35. Cerwenka, A., Carter, L. L., Reome, J. B., Swain, S. L., Dutton, R. W. (1998) In vivo persistence of CD8 polarized T cell subsets producing type 1 or type 2 cytokines. J. Immunol. 161, 97–105.
36. Cerwenka, A., Morgan, T. M., Harmsen, A. G., Dutton, R. W. (1999) Migration kinetics and final destination of type 1 and type 2 CD8 effector cells predict protection against pulmonary virus infection. J. Exp. Med.
189,423– 434.
37. Li, L., Sad, S., Kagi, D., Mosmann, T. R. (1997) CD8Tc1 and Tc2 cells secrete distinct cytokine patterns in vitro and in vivo but induce similar inflammatory reactions. J. Immunol. 158, 4152– 4161.
38. Jung, U., Foley, J. E., Erdmann, A. A., Eckhaus, M. A., Fowler, D. H. (2003) CD3/CD28-costimulated T1 and T2 subsets: differential in vivo allosensitization generates distinct GVT and GVHD effects. Blood 102, 3439 –3446.
39. Dobrzanski, M. J., Reome, J. B., Hollenbaugh, J. A., Dutton, R. W. (2004) Tc1 and Tc2 effector cell therapy elicit long-term tumor immunity by contrasting mechanisms that result in complementary endogenous type 1 antitumor responses. J. Immunol. 172, 1380 –1390.
40. Carter, L. L., Dutton, R. W. (1996) Type 1 and type 2: a fundamental dichotomy for all T-cell subsets. Curr. Opin. Immunol. 8, 336 –342. 41. Mosmann, T. R., Li, L., Sad, S. (1997) Functions of CD8 T-cell subsets
secreting different cytokine patterns. Semin. Immunol. 9, 87–92. 42. Sad, S., Marcotte, R., Mosmann, T. R. (1995) Cytokine-induced
differen-tiation of precursor mouse CD8⫹ T cells into cytotoxic CD8⫹ T cells secreting Th1 or Th2 cytokines. Immunity 2, 271–279.
43. Szabo, S. J., Kim, S. T., Costa, G. L., Zhang, X., Fathman, C. G., Glimcher, L. H. (2000) A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 100, 655– 669.
44. Mullen, A. C., Hutchins, A. S., High, F. A., Lee, H. W., Sykes, K. J., Chodosh, L. A., Reiner, S. L. (2002) Hlx is induced by and genetically interacts with T-bet to promote heritable TH1 gene induction. Nat.
Im-munol. 3, 652– 658.
45. Zhang, D-H., Cohn, L., Ray, P., Bottomly, K., Ray, A. (1997) Transcrip-tion factor GATA-3 is differentially expressed in murine Th1 and Th2 cells and controls Th2-specific expression of the interleukin-5 gene.
J. Biol. Chem. 272, 21597–21603.
46. Zheng, W-p., Flavell, R. A. (1997) The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells.