運動治療在糖尿病對心臟細胞 PI3K-Akt 存活途徑之療效探討
Effect of exercise therapy on Cardiac apoptosis and PI3K-Akt
survival pathway in diabetes
(CMU97-219)
Effect of Exercise Training on Cardiac survival pathway in streptozotocin-induced diabetic rats
Background. Cardiac apoptosis was found in diabetes but very limited information regarding the influence of chronic exercise on cardiac pro-survival and anti-apoptosis in diabetes was available. The purpose of this study was to evaluate the influence of chronic exercise on cardiac pro-survival and anti-apoptotic relative pathways. Methods. Twelve Wistar rats (Control) and nine Streptozotocin-induced diabetic rats (DM) at 4 month of age were served as negative and positive control and eleven Streptozotocin-induced diabetic rats underwent running exercise on treadmill 1 hour daily, 5 sections per week, for 8 weeks (DM-EX). After chronic exercise or sedentary status, the excised hearts were measured by H&E stain, TUNEL assays and Western Blotting. Results. Citrate synthase activity in skeletal muscle in DM-EX is significantly increased compared with sedentary Control and DM group. Levels of cardiac IGF1-R/PI3K/AKT survival pathway related proteins, including IGF1, IGF
receptor, p-PI3K, p-Akt, and pro-survival Bcl-2 family, including Bcl-2, Bcl-xL and p-BAD were decreased in DM compared with control. In DM-EX group, those survival related proteins in cardiac IGF1-R/PI3K/AKT survival pathway and
pro-survival Bcl-2 family were increased and apoptotic activities were decreased (TUNEL and active caspase-3) compared with DM.
Conclusions. Chronic exercise enhance cardiac IGF1-R/PI3K/AKT survival pathway
and pro-survival Bcl-2 family associated pathway in diabetes and attenuate the diabetes-associated cardiac apoptotic activity. Chronic exercise may be one of possible therapeutic approaches for preventing cardiac apoptosis in diabetes.
Key words. Diabetes, heart, IGF1-R/PI3K/AKT survival pathway, pro-survival Bcl-2
Introduction:
Diabetes mellitus is a worldwide epidemic whose incidence and prevalence have significantly increased in recent decades[1, 2]. Diabetes is a chronic metabolic disorder associated with secondary complications in the cardiovascular system and autonomic control in humans and animals. It is recognized as an independent risk factor for cardiovascular morbidity and mortality[3]. Several reports have described early changes of cardiac structure and function in diabetic patients. And more unclearly mechanisms relating diabetes-associated cardiac pathophysiology include the direct metabolic consequences of this disease on the myocardium[3, 4]. Diabetic cardiomyopathy is a seriously complications, according to the obtained results it could be concluded that the changing rate of diabetic cardiomyopathy was in direct correlation with the quality of diabetes control, the duration of diabetes, and the presence of complications in other organs[5]. In human and animal models of diabetes, heart disease is associated with excessive apoptotic cell death[6, 7]. It is possible that this loss of cells progressively leads to interstitial fibrosis, myocardial hypertrophy, contractile impairment, and eventual heart disease[8, 9]. However very limited information related to cardiac pro-survival pathway was available in diabetes.
It is well known that exercise affects heart rate and cardiac function in humans significantly. Physical activity and exercise has been regarded as a therapeutic approach in the treatment of diabetes [9-11]. And many studies already proved that the exercise training improved myocardial function in diabetes rats [11, 12]. Effect of exercise on cardiac pro-survival pathway in diabetes were not yet been investigated.
The insulin-like growth factor (IGF) system efficiently signals to cells to grow, differentiate, and survive. IGF-1 is a major survival factor in serum and prevents apoptosis in a number of cell types [13, 14]. When it activates phosphatidylinositol
3-kinase (PI3K) and serine/threonine kinase Akt pathway, it will attenuate the
apoptotic activity [15]. Akt is a cell survival downstream growth factors, it enhances the survival of cells by blocking the function of proapoptotic proteins and processes. Akt negatively regulates the function or expression of several Bcl-2 family, which exert anti-apoptotic effects by binding to and inactivating pro-survival Bcl-2 family members. For instance, Akt directly phosphorylates and inhibits the BH3-only protein BAD [16, 17]. Survival factors stimulate Akt-mediated phosphorylation of BAD on S136 ( p-BAD), which triggers release of BAD from its target proteins [18]. p-BAD is an anti-apoptotic protein, prevents cytochrome c release and apoptotic activity. Pro-apoptotic and anti-apoptotic Bcl2 family members can homodimerize or heterodimerize to each other, and appear to interact with and neutralize each other, so that the relative balance of these effectors strongly influences cytochrome c release. Bcl-2 and Bcl-xL, anti-apoptotic proteins, may stabilize the mitochondrial membrane and prevent the activation of downstream apoptotic signaling [19].
Apoptosis, a physiological program of cellular death, may contribute to many cardiac disorder[20, 21]. The occurrence of apoptosis has been reported to contribute to the loss of cardiomyocytes in cardiomyopathies, and is recognized as a predictor of adverse outcomes in subjects with cardiac diseases or heart failure [22]. Widely believed that caspase-3, a cysteine protease, leads to nucleosomal fragmentation of DNA [19, 23].
The therapeutic effect of exercise training on cardiac pro-survival pathway in diabetes is not understood. In the current study, we investigated whether exercise training in diabetes are associated with induced the survival signaling transduction. The heart weight index, myocardial morphology, and key components of IGF1, IGF receptor, p-PI3K, p-Akt, p-BAD, Bcl-2 and Bcl-xL and apoptotic activity in three
groups, Wistar rats (Control) and STZ-induced diabetes rats (DM) under sedentary status or after exercise training(DM-EX), were determined by heart weighing, H&E stain, TUNEL assay and Western blotting. We hypothesized that exercise training may enhance cardiac pro-survival pathway and prevent cardiac apoptotic activity in diabetes.
Materials and Methods
Animal model
This study was approved by Institutional Animal Care and Use Committee of National Cheng Kung University. All procedures followed the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (NIH). Male Wistar rats (8-week old), purchased from Laboratory Animal Center of National Cheng Kung University were randomly assigned to one of three groups. All animals were housed in a temperature-controlled room (22-25℃) with a 12 h dark-light cycle at Laboratory Animal Center of National Cheng Kung University. No statistical difference in body weight and blood glucose level was found in the beginning of the experiment. To induce type 1 diabetes mellitus, rats from the DM and DM+Exgroups were fasted for three days before induction. Rats were generally anaesthetized by an intraperitoneal injection of sodium pentobarbital in the concentration of 45 mg/kg of body weight. After anaesthetization, femoral artery pulse was palpated to identify the position of femoral vein (femoral artery and vein are parallels nearby). Then the skin superficial to the femoral vein was cut into an incision but also the superficial connective tissues were removed. When the femoral vein was exposed, Streptozotocin (65 mg/kg of body weight), freshly prepared by dissolving in 50 mM citrate buffer (pH 4.5) with the ratio of 1:65, was injected immediately into femoral vein[25]. The incision then sewed up and disinfected. To confirm the induction of type 1 diabetes, fasting blood glucose level (fasting overnight) was determined one week after STZ injection by glucose test of tail vein blood using glucometer. STZ-induced rats failed to exhibit an elevated blood glucose level greater than 200 mg/dL were excluded from the study.
Exercise training
Chen et al [26]. Before the beginning of exercise training, rats from the SHR-Ex group had run on a horizontal treadmill (Model T510E, Diagnostic and Research Instruments Co., Taiwan) at the speed of 12 m/min for the familiarization. After one week of familiarization, all rats had run on the treadmill 60 minutes per session, 5 sessions per week, for 8 weeks totally. During the training period, the running speed was gradually increased 3 m/min every two weeks, from 15 m/min to 24 m/min finally. In contrast, rats from sedentary groups were placed on the treadmill without running for 15 minutes each session. To avoid acute effect of exercise, all animals were sacrificed 48 hours after exercise training. Body weight and fasting blood glucose level were measured before sacrificing.
Cardiac characteristics
The hearts of wistar rats and diabetes rats were excised and cleaned with PBS (Phosphate buffered saline). The left ventricle were separated and weighed. The right tibias were also separated and tibia lengths were measured by the electronic digital venire caliper to adjust the whole heart weight. The ratios of the total heart weight to body weight, the left ventricle weight to body weight, the left ventricle weight to the whole heart weight, the whole heart weight to tibia length, and the left ventricle weight to tibia length were calculated.
Tissue Extraction
Cardiac tissue extracts were obtained by homogenizing the left ventricle samples in a lysis buffer at a ratio of 100 mg tissue/1ml buffer for 1 min. The homogenates were placed on ice and then centrifuged at 12,000 g for 40 min. The supernatant was collected and stored at -80°C for further experiments.
Assay of citrate synthase activity
studies have proven that citrate synthase activity increased significantly after chronic exercise. Therefore an increase in citrate synthesis activity is commonly used as a surrogate index for the effectiveness of exercise training. Soleus muscles (weight ~0.1g) isolated from four groups were immediately stored at -80℃ until analyzed. When measuring, the defrosted muscle samples were homogenized by homogenizer (Model 398, Biospec Products Inc, Mexico) with quintuple volumes of Tris buffer (0.1 mol/L) containing Triton X-100 (0.1%). After homogenizing, the samples were centrifuged by centrifuge (Kendro Laboratory Products, Langenselbold, Germany) at the speed of 13000 rpm for 15 minutes, and then the supernatants were collected. The 50 μlofsupernatantwasadded into cuvette which had already contained 100 μlof DTNB (1mM),30μlofacetyl-CoA (10mM), and 770μlofH2O.Theabsorption at 412 nm of wave length was measured by spectrophotometer (Beckman Instruments, Inc., DU640B, CA, USA) for 10 minutes to measure the possible acetyl-CoA deacylase activity. With the addition of 50μl ofoxaloacetate (10mM), the measurement of enzymatic activity was begun also by spectrophotometeric assay for 5 minutes. Every sample tested twice and averaged. The result was expressed as micromoles of substrate utilized per minute per gram of wet tissue.
Electrophoresis and Western Blot
Protein concentration of cardiac tissue extracts was determined by the Lowry protein assay. Protein samples (40μg/lane) were separated on a 10% SDS polyacrylamide gel electrophoresis (SDS-PAGE) with a constant voltage of 75 V. Electrophoresed proteins were transferred to polyvinylidene difluoride (PVDF) membrane(Millipore,Bedford,MA,0.45 μm pore size) with a transfer apparatus (Bio-red). PVDF membranes were incubated in 5% milk in TBS buffer. Primary antibodies including IGF1, IGF receptor, p-PI3K, PI3K, p-Akt, Akt, p-BAD, Bcl-2,
Bcl-xL and α-tubulin (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were diluted to 1:500 in antibody binding buffer overnight at 4°C. The immunoblots were washed three times in TBS buffer for 10 min and then immersed in the second antibody solution containing goat anti-mouse IgG-HRP, goat anti-rabbit IgG-HRP, or donkey anti goat IgG-HRP (Santa Cruz) for 1 hour and diluted 500-fold in TBS buffer. The immunoblots were then washed in TBS buffer for 10 min three times. The immunoblotted proteins were visualized using an enhanced chemiluminescence ECL western Blotting luminal Reagent (Santa Cruz, CA, USA) and quantified using a Fujifilm LAS-3000 chemiluminescence detection system (Tokyo, Japan).
Hematoxylin-eosin staining (H&E staining) and Terminal Deoxynucleotide Transferase-mediated dUTP Nick End Labeling (TUNEL)
After the hearts were excised, the hearts were soaked in formalin, dehydrated through graded alcohols, and embedded in paraffin wax. In heart tissues, the 0.2-µm thick paraffin sections were cut from paraffin-embedded tissue blocks. The tissues sections were deparaffinized by immersing in xylene, and rehydrated. For Hematoxylin-eosin staining, the slices were then dyed with hematoxylin and eosin. After gently rinsing with water, each slide was dehydrated through graded alcohols. Finally, they were soaked in xylene twice. Photomicrographs were obtained using Zeiss Axiophot microscopes. For TUNEL assay, the sections were incubated with proteinase K, washed in phosphate-buffered saline, incubated with permeabilisation solution, blocking buffer, and then washed two times with PBS. The terminal deoxynucleotidyl transferase and fluorescein isothiocyanate-dUTP for 60 min at 37 °C from an apoptosis detection kit (Roche Applied Science, Indianapolis, IN, USA) was used for detection. Then added the DAPI (4,6–diamidino-2-phenylindole) 5mins and the nucleus position were fluoresced by blue light at 340 / 380 nm. TUNEL-positive
nuclei (fragmented DNA) were fluoresced by bright green light at 450-500 nm. The mean number of TUNEL-positive cells were counted for at least 5-6 separate fields x 2 slices x 3 regions of the left ventricle (upper, middle, lower) excised from six rat hearts in each group. All counts were performed by at least two independent individuals in a blinded manner.
Statistical Analysis
The all data of weight index, blood sugar, protein levels, and the percentage of TUNEL positive cells were compared among the control, DM, and DM-EX groups using one-way analysis of variance (ANOVA) with pre-planned contrast comparison. In all cases, P<0.05 was considered significant.
Results
Body weight and cardiac characteristics
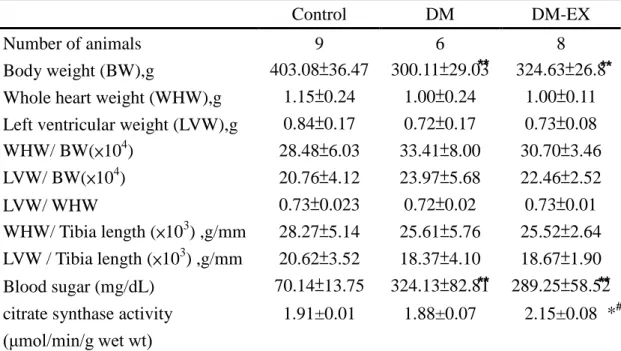
DM and DM-EX weighed were less than the Control. The other index of whole heart weight (WHW), left ventricular weight (LVW), WHW/body weight, LVW/body weight, LVW/WHW, WHW/tibia and LVW/tibia length showed no different amount three groups, Control, DM, DM-EX. And the blood sugar increased in DM and decreased in DM-EX (Table 1).
Assay of citrate synthase activity
Exercise training for 8 weeks significantly increased the citrate synthase activity of soleus muscle in the DM-Ex group as compared with the Control, DM groups (P<0.05), as shown in Table 1. These results indicated that our exercise program was effective.
Hematoxylin-eosin staining (H&E staining)
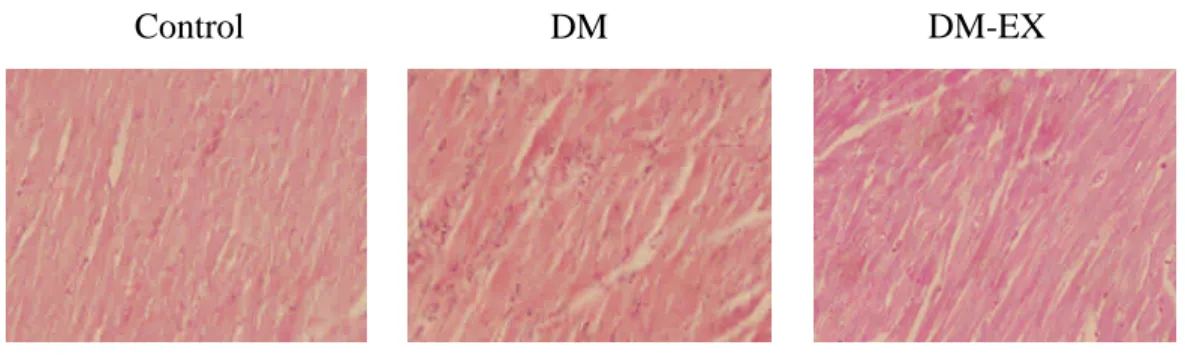
To investigate whether there were changes in cardiac architecture, we did a histopathological analysis of ventricular tissue stained with hematoxylin and eosin. After viewing 400 X magnified images, we found that the ventricular myocardium in the Control group showed normal architecture with normal interstitial space, but the abnormal myocardial architecture and the increased interstitial space were observed in the DM group. These myocardial architecture abnormalities in the DM-EX group became normal than those in the DM group (Fig 1).
Cardiac IGF1-R/PI3K/AKT survival pathway
To investigate the components of Cardiac IGF1-R/PI3K/AKT survival pathway in diabetic rats after exercise training, we measured the pro-survival related protein levels of in hearts excised from control, DM, and DM-EX groups. Compared with the control group, the protein levels of IGF 1, IGF receptor (Fig 2), p-PI3K and p-Akt
(Fig 3) were significantly decreased in the DM group. The protein levels of IGF 1, IGF receptor (Fig 2), p-PI3K and p-Akt (Fig 3), in the DM-EX group were significantly higher than those in the DM group.
Cardiac pro-survival Bcl-2 family associated pathway
In order to identify the components of Cardiac pro-survival Bcl-2 family associated pathway were measured by Western blotting in the hearts excised from the control, DM, and DM-EX groups. Compared to the control group, the protein products of p-BAD, Bcl-2 and Bcl-xL were decreased in the DM groups (Fig 4). The protein levels in DM-EX group were increased compared with those in the DM group (Fig 4).
TUNEL-positive apoptotic cells of cardiac tissues
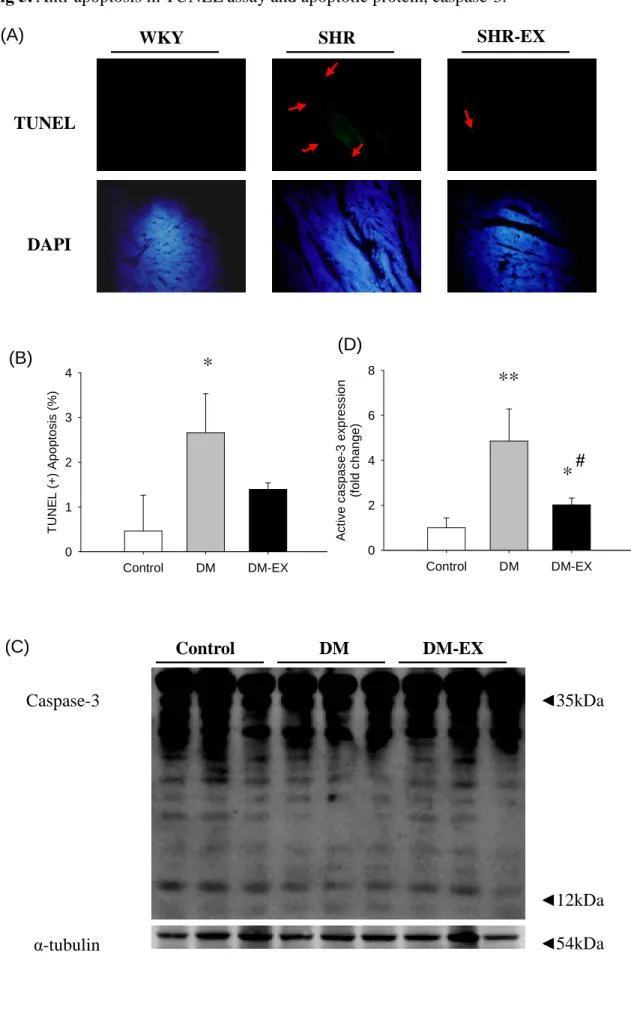
In order to view the apoptotic activity in cardiac tissues, the apoptotic cells and total cells were measured by TUNEL assay and DAPI staining respectively in the hearts excised from the control, DM, and DM-EX groups. Viewing images magnified 400 X, we observed that the left ventricles of the SHR groups stained with TUNEL assay had a greater number of TUNEL-positive cardiac cells than those in the control; group whereas the number of TUNEL-positive cardiac cells was similar between control and DM-EX group. Decreases in number of TUNEL-positive cardiac cells were found in the DM-EX group, compared with DM group (Fig 5).
Discussion
Major findings
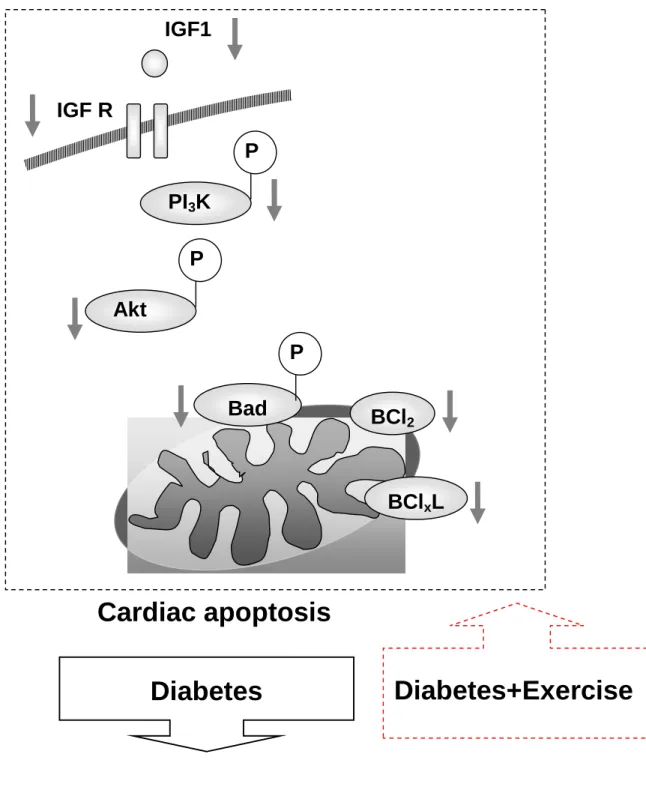
Our main findings can be summarized as follows: Our main findings can be summarized as follows: (1) the myocardial architecture abnormalities in diabetic rats after exercise training became normal than those in the diabetic rats sowed in body weight and blood sugar. And whole heart weight, left ventricular weight, whole heart weight normalized by tibia length and body weight, left ventricular weight normalized by tibia length and body weight in both groups were no difference. (2) The activity of the cardiac IGF1-R/PI3K/AKT survival pathway and pro-survival Bcl-2 family associated pathway in diabetic rats were significantly increased after exercise training, the evidence for which is based on decreases in IGF1, IGF receptor, p-PI3K, p-Akt, p-BAD, Bcl-2 and Bcl-xL, compared with the sedentary hypertensive group. (3) After integrating our current findings into improved pro-survival theories, we draw a hypothesized diagram in Fig 6, which suggest that cardiac pro-survival pathways were decreased in diabetes and stimuli by exercise training. Our findings imply that exercise therapy could be one of possible therapeutic approaches to prevent delirious cardiac apoptosis in diabetes mellitus.
Experimental limitation
The benefit of exercise may be involved in multiple factors such as anti-hypertension, weight loss, anti-diabetes, and normalized lipid profiles [27]. Exercise improved heart function successfully and further attenuated age-induced elevation in BAX/Bcl-2 ratio, and inhibit apoptosis in rat heart[24]. Therefore, the underlying mechanism how exercise prevent cardiac apoptosis in diabetes remain unknown. Any therapeutic effect of exercise training on cardiomyopathic changes and cardiac apoptosis noted in the current study cannot be isolated and attributed to any
specific factor, such as weight loss, blood pressure changes or other unclear factors. However, the current study can differentiate therapeutic effects of exercise training on cardiomyopathic changes and cardiac apoptosis.
Cardiac changes
While in the recently studies, exercise training has been shown to protect the myocardium dysfunction[28, 29]. However, the current study can differentiate therapeutic effects of sedentary diabetes and diabetes with chronic exercise training. The current study showed that exercise training improved myocardial structure. The balance between cell death and cell survival is a tightly controlled process, especially in terminally differentiated cells, such as the cardiomyocytes. Therefore, prevention of cardiac apoptosis and cardiac cell death in heart health is very critical issues in the diabetes-associated heart diseases.
Cardiac IGF1-R/PI3K/AKT survival pathway and pro-survival Bcl-2 family associated pathway
This study also represents the first to report the prevention from a decrease in the cardiac survival pathway, in diabetes after exercise training. The IGF1, IGF receptor, p-PI3K, p-Akt, p-BAD, Bcl-2 and Bcl-xL, which were suppress cardiac myocyte apoptosis. In the current study, exercise training was found to significantly prevent cardiac apoptosis observed in sedentary diabetes, as evidenced by decreases in cardiac IGF1, IGF receptor, p-PI3K, p-Akt, p-BAD, Bcl-2 and Bcl-xL, protein expression levels in diabetes rats hearts and increase after exercise training. Therefore, our findings strongly suggest that exercise training in diabetes could prevent cardiac apoptosis by affect survival pathway.
Hypothesized clinical application
Diabetes is considered as a major risk factor for the development of heart failure, even when other known risk factors for heart failure are excluded. Since cardiac tissues are difficult to be extracted from diabetic human hearts, the current diabetic animal model under exercise training should provide an important explanation how clinical exercise prescription prevents heart failure or apoptosis-related cardiac diseases in diabetic humans. Since diabetes will enhance cardiac apoptosis and exercise training can prevent this progression, diabetes should be highly aware of the possibility of progressive development in cardiac abnormality as well as the importance of exercise therapy and modified lifestyle. Based on the current evidence from animal study showing that exercise training did effect the survival pathway and prevent diabetes associated cardiac disease, we might further hypothesize exercise is a medicine of cardiac apoptosis in diabetes mellitus. Besides, further questions is raised what underlying mechanism stimuli cardiac survival pathway when exercise training undergo as well as what exercise intensity, exercise duration, and exercise frequency are minimal requirement to prevent cardiac apoptosis in the certain degree of diabetes mellitus. Of course, further studies are required to elucidate specific mechanisms responsible for this therapeutic effect in diabetes and further clinical studies are required to clarify the possible therapeutic application in diabetic human.
References
1. Raddino, R., et al., Diabetes and ischemic heart disease: specific treatment. G Ital Cardiol (Rome), 2008. 9(10 Suppl 1): p. 52S-55S.
2. Gonzalez, A., et al., Apoptosis in hypertensive heart disease: a clinical
approach. Curr Opin Cardiol, 2006. 21(4): p. 288-94.
3. Sack, M.N., Type 2 diabetes, mitochondrial biology and the heart. J Mol Cell Cardiol, 2009.
4. Morrish, N.J., et al., Mortality and causes of death in the WHO Multinational
Study of Vascular Disease in Diabetes. Diabetologia, 2001. 44 Suppl 2: p.
S14-21.
5. Perisic, Z., et al., [Clinical and echocardiographic characteristics of patients
with diabetic cardiomyopathy]. Vojnosanit Pregl, 2004. 61(3): p. 259-66.
6. Frustaci, A., et al., Myocardial cell death in human diabetes. Circ Res, 2000. 87(12): p. 1123-32.
7. Fiordaliso, F., et al., Myocyte death in streptozotocin-induced diabetes in rats
in angiotensin II- dependent. Lab Invest, 2000. 80(4): p. 513-27.
8. Ghosh, S., et al., Brief episode of STZ-induced hyperglycemia produces
cardiac abnormalities in rats fed a diet rich in n-6 PUFA. Am J Physiol Heart
Circ Physiol, 2004. 287(6): p. H2518-27.
9. Ghosh, S., et al., Role of dietary fatty acids and acute hyperglycemia in
modulating cardiac cell death. Nutrition, 2004. 20(10): p. 916-23.
10. Colberg, S.R., Physical activity, insulin action, and diabetes prevention and
control. Curr Diabetes Rev, 2007. 3(3): p. 176-84.
11. De Angelis, K.L., et al., Effects of exercise training on autonomic and
myocardial dysfunction in streptozotocin-diabetic rats. Braz J Med Biol Res,
2000. 33(6): p. 635-41.
12. Thompson, P.D., Exercise and physical activity in the prevention and
treatment of atherosclerotic cardiovascular disease. Arterioscler Thromb Vasc
Biol, 2003. 23(8): p. 1319-21.
13. Hong, F., et al., Insulin-like growth factor-1 protects H9c2 cardiac myoblasts
from oxidative stress-induced apoptosis via phosphatidylinositol 3-kinase and extracellular signal-regulated kinase pathways. Life Sci, 2001. 68(10): p.
1095-105.
14. Vincent, A.M. and E.L. Feldman, Control of cell survival by IGF signaling
pathways. Growth Horm IGF Res, 2002. 12(4): p. 193-7.
15. Hedhli, N., M. Pelat, and C. Depre, Protein turnover in cardiac cell growth
and survival. Cardiovasc Res, 2005. 68(2): p. 186-96.
cell-intrinsic death machinery. Cell, 1997. 91(2): p. 231-41.
17. del Peso, L., et al., Interleukin-3-induced phosphorylation of BAD through the
protein kinase Akt. Science, 1997. 278(5338): p. 687-9.
18. Datta, S.R., et al., Survival factor-mediated BAD phosphorylation raises the
mitochondrial threshold for apoptosis. Dev Cell, 2002. 3(5): p. 631-43.
19. Wang, L., et al., Regulation of cardiomyocyte apoptotic signaling by
insulin-like growth factor I. Circ Res, 1998. 83(5): p. 516-22.
20. Haunstetter, A. and S. Izumo, Apoptosis: basic mechanisms and implications
for cardiovascular disease. Circ Res, 1998. 82(11): p. 1111-29.
21. Lee, S.D., et al., Roles of insulin-like growth factor II in cardiomyoblast
apoptosis and in hypertensive rat heart with abdominal aorta ligation. Am J
Physiol Endocrinol Metab, 2006. 291(2): p. E306-14.
22. Narula, J., et al., Apoptosis in heart failure: release of cytochrome c from
mitochondria and activation of caspase-3 in human cardiomyopathy. Proc
Natl Acad Sci USA, 1999. 96(14): p. 8144-9.
23. Nicholson, D.W., et al., Identification and inhibition of the ICE/CED-3
protease necessary for mammalian apoptosis. Nature, 1995. 376(6535): p.
37-43.
24. Kwak, H.B., W. Song, and J.M. Lawler, Exercise training attenuates
age-induced elevation in Bax/Bcl-2 ratio, apoptosis, and remodeling in the rat heart. Faseb J, 2006. 20(6): p. 791-3.
25. Angulo, J., et al., Impairment of nitric oxide-mediated relaxations in
anaesthetized autoperfused streptozotocin-induced diabetic rats. Naunyn
Schmiedebergs Arch Pharmacol, 1998. 358(5): p. 529-37.
26. Chen Hi, H., I.P. Chiang, and C.J. Jen, Exercise Training Increases
Acetylcholine-Stimulated Endothelium-Derived Nitric Oxide Release in Spontaneously Hypertensive Rats. J Biomed Sci, 1996. 3(6): p. 454-460.
27. Elwood PC, Y.J., Pickering J, Fehily AM,O’Brien JR,Exercise, Wbrinogen, and other risk factors for ischemic heart disease. Br Heart J, 1993.
28. Starnes, J.W. and D.K. Bowles, Role of exercise in the cause and prevention of
cardiac dysfunction. Exerc Sport Sci Rev, 1995. 23: p. 349-73.
29. Powers, S.K., et al., Exercise and cardioprotection. Curr Opin Cardiol, 2002. 17(5): p. 495-502.
Table 1. Cardiac characteristics of Control, DM group and DM with exercise training
Control DM DM-EX
Number of animals 9 6 8
Body weight (BW),g 403.08±36.47 300.11±29.03 324.63±26.8 Whole heart weight (WHW),g 1.15±0.24 1.00±0.24 1.00±0.11 Left ventricular weight (LVW),g 0.84±0.17 0.72±0.17 0.73±0.08
WHW/ BW(×104) 28.48±6.03 33.41±8.00 30.70±3.46 LVW/ BW(×104) 20.76±4.12 23.97±5.68 22.46±2.52 LVW/ WHW 0.73±0.023 0.72±0.02 0.73±0.01 WHW/ Tibia length (×103) ,g/mm 28.27±5.14 25.61±5.76 25.52±2.64 LVW / Tibia length (×103) ,g/mm 20.62±3.52 18.37±4.10 18.67±1.90 Blood sugar (mg/dL) 70.14±13.75 324.13±82.81 289.25±58.52 citrate synthase activity
(μmol/min/g wetwt)
1.91±0.01 1.88±0.07 2.15±0.08
Values are means SD among Wistar rats (Control), streptozotocin-induced diabetic rats (DM) and diabetic rats with exercise training (DM-EX). *P<0.05, ** P<0.01
significant differences between Control and DM or Control and DM-EX group.
#
P<0.05 significant differences between DM group and DM-EX group.
** **
** **
Fig 1. H&E
Fig 1. Representive histopathological analysis of cardiac sections from left ventricles
was performed with Hematoxylin and eosin (H&E) staining in Wistar rats (Control),
streptozotocin-induced diabetic rats (DM) and diabetic rats with exercise training
(DM-EX). The images of cardiac architecture were magnified by 400 times. (n=3~4
in each group).
Fig 2.
Fig 1. (A) The representive protein products of insulin-like growth factor one (IGF1) and IGF receptor extracted from the left ventricles of excised hearts in 3 Wistar rats (Control), streptozotocin-induced diabetic rats (DM) and diabetic rats with exercise training (DM-EX) were measured by Western Blotting analysis. α-tubulin was use as a internal control. (B) (C) Bars represent the relative fold changes of protein quantification relative to Control group in IGF 1/α-tubulin and IGF receptor /α-tubulin and mean values±SD (n=6 in each group). *P<0.05, **P<0.01, significant differences from Control group. ##P<0.01 significant differences between DM group
and DM-EX group. IGF 1 IGF receptor α-tubulin ◄17kDa ◄155kDa ◄54kDa (B) (C) DM-EX DM Control (A) Control DM DM-EX IG F R e c e p to r e x p re s s io n (f o ld c h a n g e ) 0.0 0.5 1.0 1.5
**
##**
Control DM DM-EX IG F 1 e x p re s s io n (f o ld c h a n g e ) 0.0 0.5 1.0 1.5 2.0*
##Fig 3.
Fig 1. (A) The representive protein products of phosphates Phosphoinositide-3 kinase (p-PI3K) and p-Akt extracted from the left ventricles of excised hearts in 3 Wistar rats
(Control), streptozotocin-induced diabetic rats (DM) and diabetic rats with exercise training (DM-EX) were measured by Western Blotting analysis. α-tubulin was use as a internal control. (B) (C) Bars represent the relative fold changes of protein quantification relative to Control group in p-PI3K /α-tubulin and p-Akt /α-tubulin and
mean values±SD (n=6 in each group). **P<0.01, significant differences from Control group. ##P<0.01 significant differences between DM group and DM-EX group.
p-PI3K p-Akt α-tubulin ◄80kDa ◄60kDa ◄54kDa (B) (C) Control DM DM-EX p -P I3 K e x p re s s io n (f o ld c h a n g e ) 0.0 0.5 1.0 1.5
**
## DM-EX DM Control (A) Control DM DM-EX p -A k t e x p re s s io n (f o ld c h a n g e ) 0 1 2 3**
##Fig 4.
Fig 3. (A) The representive protein products of phosphates Bcl-2-associated death promoter (p-BAD), B-cell lymphoma 2 gene (Bcl-2) and B-cell lymphoma-extra large (Bcl-xL), extracted from the left ventricles of excised hearts in 3 Wistar rats (Control), streptozotocin-induced diabetic rats (DM) and diabetic rats with exercise training (DM-EX) were measured by Western Blotting analysis. α-tubulin was use as a internal control. (B) (C) (D) Bars represent the relative fold changes of protein quantification relative to Control group in p-BAD /α-tubulin and Bcl-2/α-tubulin and Bcl-xL /α-tubulin and mean values±SD (n=6 in each group). **P<0.01, significant differences from Control group. #P<0.05, ##P<0.01 significant differences between
DM group and DM-EX group.
Control DM DM-EX B c l-x L e x p re s s io n (f o ld c h a n g e ) 0.0 0.5 1.0 1.5
**
## Control DM DM-EX B c l-2 e x p re s s io n (f o ld c h a n g e ) 0.0 0.5 1.0 1.5**
## Control DM DM-EX p -B A D e x p re s s io n (f o ld c h a n g e ) 0.0 0.5 1.0 1.5 2.0 ** # Bcl-2 Bcl-xL α-tubulin ◄25kDa ◄30kDa ◄25kDa (B) (C) DM-EX DM Control (A) ◄54kDa p-BAD (D)Fig 5. Anti-apoptosis in TUNEL assay and apoptotic protein, caspase-3. SHR-EX SHR WKY (A) TUNEL DAPI (B) Control DM DM-EX T U N E L (+ ) A p o p to s is (% ) 0 1 2 3 4
*
DM-EX DM Control (C) Caspase-3 α-tubulin ◄12kDa ◄54kDa ◄35kDa Control DM DM-EX A c ti v e c a s p a s e -3 e x p re s s io n (f o ld c h a n g e ) 0 2 4 6 8**
*
# (D)Fig 5. (A) Representive stained apoptotic cells of cardiac sections from left ventricle
in Wistar rats (Control), streptozotocin-induced diabetic rats (DM) and diabetic rats
with exercise training (DM-EX) were measured by staining with TUNEL assay with
dark background (right panels, green spots). The relative background tissues were
shown in left panels. The images were magnified by 400 times. (B) Bars present the
percentage of TUNEL positive cells relative to total cells. (C) Protein expression level
of caspase-3. α-tubulin was use as a internal control. (D) Bars present the statistic
result. *P<0.05, **P<0.01, significant differences from Control group. #P<0.05
Fig 6.