International Journal of Radiation Biology, May 2013; 89(5): 346–355 © 2013 Informa UK, Ltd.
ISSN 0955-3002 print / ISSN 1362-3095 online DOI: 10.3109/09553002.2013.762136
Inhibitory effects of Rhenium-188-labeled Herceptin on prostate
cancer cell growth: A possible radioimmunotherapy to prostate
carcinoma
Hsin-Yi Wang
1,2, Wan-Yu Lin
2, Mei-Chih Chen
1, Teh Lin
9, Chih-Hao Chao
3, Fu-Ning Hsu
1, Eugene
Lin
1,4, Chih-Yang Huang
5,11, Tsai-Yueh Luo
6& Ho Lin
1,7,8,101Department of Life Sciences, National Chung Hsing University, Taichung, Taiwan, 2Department of Nuclear Medicine, Taichung Veterans General Hospital, Taichung, Taiwan, 3Department of Internal Medicine, Chang Bing Show Chwan Memorial Hospital, Changhwa, Taiwan, 4Department of Urology, Chang Bing Show Chwan Memorial Hospital, Changhwa, Taiwan,
5Graduate Institute of Basic Medical Science, China Medical University, Taichung, Taiwan, 6Institute of Nuclear Energy Research, Taoyuan, Taiwan, 7Graduate Institute of Rehabilitation Science, China Medical University, Taichung, Taiwan, 8Agricultural Biotechnology Center, National Chung Hsing University, Taichung, Taiwan, 9Department of Radiation Oncology, Fox Chase Cancer Center, Philadelphia, Pennsylvania, USA, 10Department of Urology, University of Texas Southwestern Medical Center, Dallas, Texas, USA, and 11Department of Health and Nutrition Biotechnology, Asia University, Taichung, Taiwan
Abstract
Purpose: Herceptin is widely used in treating Her2-overexpressing breast cancer. However, the application of Herceptin in prostate cancer is still controversial. Our previous results have indicated the relevance of Her2 in the transition of the androgen require- ment in prostate cancer cells. In this study, the effects of radioim- munotherapy against Her2 in prostate cancer were investigated. Materials and methods: DU145, an androgen receptor-negative prostate cancer cell line, was used in vitro and in vivo to evaluate the effects of Herceptin labeled with a beta emitter, Rhenium- 188 (Re-188). Its effects on cell growth, extent of apoptosis, the bio-distribution of Re-188 labeled Herceptin (Re-H), and protein levels were determined.
Results: Treatments with Re-188 and Re-H reduced the prolif-eration of DU145 cells in dose- and time-dependent manners compared to the Herceptin-treated group. Growth inhibition and apoptosis were induced after Re-H treatment; growth inhi-bition was more distinct in cells with high Her2/p-Her2 levels. Our in vivo xenograft studies revealed that Re-H treatment significantly retarded tumor growth and altered the levels of apoptosis-related proteins. The bio-distribution of Re-H in mice demonstrated a tissue-specific pattern. Importantly, the levels of p35 protein, which is related to cancer cell survival and inva-sion, dramatically decreased after Re-H treatment.
Conclusions: Our data demonstrate that Re-188-labeled Herceptin effectively inhibited the growth of DU145 cells compared to the Herceptin- and Re-188-treated cohorts. This implies that targeting Her2 by both radio- and immuno-therapy might be a potential strategy for treating patients with androgen-independent prostate cancer.
Introduction
Prostate cancer is the most common malignancy in males (Jemal et al. 2007). Hormone therapy is the primary treatment for locally advanced and metastatic prostate can- cers (Henry and O’Mahony 1999). However, these tumors tend to convert to a metastatic hormone-refractory state which is currently incurable (Stavridi et al. 2010). Human epidermal growth factor receptor 2 (Her2) is a transmem- brane receptor tyrosine kinase normally involved in cell growth, differentiation, and transformation (Dougall et al. 1994). The Her2 expression rate is approximately 9–30% in primary prostate cancers; it increases after hormone treatment to 59–68% in localized tumors and 78–80% in cas- trated metastatic tumors (Signoretti et al. 2000, Osman et al. 2001, Shi et al. 2001, Morris et al. 2002, Calvo et al. 2003, Hernes et al. 2004). It has been reported that PTEN (phosphatase and tensin homolog) stratification with high levels of Her2/3 expression is important to the prognosis of prostate cancer patients (Ahmad et al. 2011). Impor-tantly, the elevated expression of Her2 is associated with exposure to hormone therapy and the androgen indepen-dence of prostate cancer (Shi et al. 2001).
Herceptin (trastuzumab), a humanized anti-Her2 monoclonal antibody, is widely used for the treatment of patients with Her2-overexpressing breast cancer (Albanell et al. 2003). Although applied in prostate cancer, the therapeutic effect of Herceptin is not as encouraging as that in breast cancer (Ziada et al. 2004). In xenograft models, Herceptin demonstrates growth inhibition in
dependent tumors but not in androgen-independent Keywords: Herceptin, Rhenium-188, prostate cancer, Her2,
Cdk5 tumors (Agus et al. 1999). Several lines of evidence indicate
Correspondence: Prof. Ho Lin, PhD, Department of Life Sciences, National Chung Hsing University, Taichung, Taiwan. Tel: + 886 4228 40416 ext. 310.
In t J R ad ia t B io l D ow nl oa de d fr om in fo rm ah ea lth ca re .c om b y T ai pe i V et er an s G en er al H os pi ta l o n 05 /0 1/ 13 F or p er so na l u se o nl y.
Fax: + 886 4228 74740. E-mail: hlin@d r agon.nc h u.edu.tw
(Received 9 July 2012; revised 2 November 2012; accepted 14 December 2012)
Rhenium-188-labeled Herceptin and prostate cancer 347
that combination therapy of Herceptin with other drugs (such as docetaxel or gefitinib) would be beneficial to treat prostate cancer (Formento et al. 2005, Legrier et al. 2007). Radio-immunotherapy refers to the treatment of a disease by using radiopharmaceuticals which are composed of an antibody labeled with a therapeutic radionuclide (Sharkey and Goldenberg 2011). The antibody targets the disease site through its specificity for a disease-associated antigen and the radionuclide delivers a lethal dose of therapeutic radiation to the disease site. Radiolabeling of Herceptin has been used to develop a therapeutic radiopharmaceuti- cal for Her2-overexpressing tumors (Rasaneh et al. 2010).
Rhenium-188 (Re-188), a β-emitting radionuclide, is widely utilized in radio-immunotherapy due to its optimal radia-tion energy (maximum of 2.12 MeV for β ), suitable half-life (16.9 hours), and convenient production (Chen et al. 2009, Luo et al. 2009). Apart from its application to breast cancer and nasopharyngeal carcinoma, the relevance of Re-188-labeled Herceptin in treating prostate cancer has not yet been studied.
Cyclin-dependent kinase 5 (Cdk5) is primarily found in bovine brain and responsible for nervous system develop- ment (Lee et al. 1996, Dhavan and Tsai 2001). Cdk5 is regu- lated by p35 protein, not cyclins, is expressed in neurons and contributes to neuronal differentiation (Tsai et al. 1994). In recent years, Cdk5 overactivation in the brain has been associated with neurodegenerative diseases, such as Alzheimer’s disease (Patrick et al. 1999). Our previous stud- ies in a neurodegenerative model also indicated that Cdk5 can be overactivated by an upstream kinase (Lin et al. 2007a). With regard to cancer cells, Cdk5 and p35 may con- trol the growth of thyroid cancer cells under Her2 regula-tion (Lin et al. 2007b). Hyperactivaregula-tion of Cdk5 also determines the fate of prostate cancer cells after drug treat- ment (Lin et al. 2004). Our recent findings have indicated that p35/Cdk5 activation regulates protein stability of the androgen receptor and the tumor growth of prostate cancer cells (Hsu et al. 2011b). Moreover, we have demonstrated that Her2 is required for stability of the androgen receptor during the transition of androgen requirement of prostate cancer cells (Hsu et al. 2011a). In addition, Strock et al. (2006) found that Cdk5 controls the motility and metastatic potential of prostate cancer cells. These findings suggest that the protein levels of p35/Cdk5 and Cdk5 kinase activity in cancer cells might be correlated to their functions. There- fore, Cdk5 and p35 might be potential indicators in our study.
To the best of our knowledge, the inhibitory effect of Re-188-labeled Herceptin in prostate cancer is not known. The primary aim of this study was to determine these effect by treating Her2-positive/androgen receptor-negative DU145 prostate cancer cell lines in vitro and in vivo with Re-188- labeled Herceptin and compare them to cohorts treated with Herceptin alone and Re-188 alone. The results indicated that Re-188 labeling enhanced the effectiveness of Herceptin treatment on the growth inhibition and apoptosis of Her2- positive prostate cancer cells. Therefore, Re-188-labeled Herceptin is a
potential therapeutic for the treatment of Her2-positive/androgen receptor-negative prostate cancer.
In t J R ad ia t B io l D o w n lo ad e d f ro m in fo rm ah e al th ca re .c o m b y Ta ip e i V et e ra n s G e n e ra l H o sp it al o n 0 5 /0 1 /1 3 F o r p er so n al u se o n ly .
348 H.-Y. Wang et al.
Materials
and methods
C e l l c u l t u r eFive human prostate cancer cell lines, DU145 (BCRC-60348), LNCaP (BCRC-60088), PC3 (60122), CWR22Rv1 (22Rv1, BCRC-60545), and BT-474 (BCRC-60359) were pur-chased from Food Industry Research and Development Institute, Taiwan. LNCaP, CWR22Rv1, and PC3 cells were cultured in RPMI-1640 (Sigma, St Louis, MO, USA), and DU145 cells were cultured in DMEM (Sigma) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Invitrogen, Carlsbad, CA, USA), 1% penicillin and streptomy- cin (Sigma, St Louis, MO, USA). HPEpiC is a normal prostate epithelial cell line cultured in a specific commercial condi- tioned medium (purchased from ScienCell Research Laboratories, Carlsbad, CA, USA). All cell lines were
main-tained in a humidified incubator at 37°C in an atmosphere contain ing 5% CO2. Preparation of Re-188 labeled Herceptin
Generation and labeling of Rhenium-188 (Re-188) onto Herceptin was performed in the Institute of Nuclear Energy Research (INER), Taoyuan, Taiwan (Luo et al. 2009). The anti-Her2/neu antibody, Herceptin (trastuzumab), was purchased from Genentech (San Francisco, CA, USA).
Evaluation of cell proliferation inhibition induced by Re-188-labeled Herceptin
Doses of Herceptin (H) and Rhenium-188 (Re-188) comparable to those used for Re-188-labeled Herceptin (Re-H) were administered to DU145 cells. DU145 cells were cultured in 24-well plates (20,000 cells/24-well) 8 h before administering the therapeutic agents. Re-H: The radioactivity of Re-H was measured by a dose calibrator (Atomlab 100,
Biodex). Seven serial doses (1, 5, 10, 20, 40, 80, and 160 μCi) of Re-H were administered in triplicate. H: Seven serial concentrations of Herceptin that matched the Herceptin concentrations in Re-H (0.3125, 0.625, 1.25, 2.5, 5, 10 and 20 mg/ml) were administered in triplicate. Re-188: Seven
serial doses of Re-188 (1, 5, 10, 20, 40, 80, and 160 μCi) were administered in triplicate. Control: Each cell line was treated with PBS (phosphate buffered saline) solution instead of the therapeutic agent in triplicate and used as control. Three hours later, the supernatants were removed. The wells were washed gently with PBS to remove non-binding agents. The culture medium was then added into each well, and the
plates were incubated at 37°C in an atmosphere containing
5% CO2. The cells were cultured for 4 and 6 days before evaluating proliferation by MTT assay (Sigma, St Louis, MO,
USA) as described previously (Hsu et al. 2011b).
LNCaP, DU145, and PC3 cells, expressing different levels of Her2, were cultured as mentioned above and treated in tripli- cate with comparable doses of H and Re-188 (75, 150, and
300 μCi) as well as with a comparable volume of PBS as the
control for 3 h. Three hours after drug administration, the cells
were washed with PBS, the appropriate culture medium was added, and the cells were then cultured for 3 days. Cell prolif- eration was evaluated by MTT assay 3 days after treatment and presented as the percentage of the PBS-treated group.
TUNEL assay
Cell apoptosis was characterized using a TUNEL assay with an In Situ Cell Death Detection Kit from Roche Applied Science (Mannheim, Germany) according to the manufac- turer’s instructions. Briefly, cells were grown on cover glasses and fixed with 4% paraformaldehyde in PBS for 1 h at room temperature. After washing three times with PBS, the cells were permeabilized with 0.1% Triton X-100 (Sigma, St Louis, MO, USA) in 0.1% sodium citrate for 10 min on ice. The cells were then stained for the TUNEL assay, and green fluorescent apoptotic bodies were visualized under a fluo- rescence microscope (Zeiss Axioplan2 fluorescence micro- scope, Germany). DAPI was used for counterstaining. The experiments were performed in duplicate and repeated twice (Lin et al. 2007a).
Evaluation of tumor growth inhibition induced by Re-188 labeled Herceptin
Male severe combined immunodeficiency (SCID) mice, 4– 6 weeks old, were maintained in a pathogen-free environment. The mice were inoculated subcutaneously in the left thigh with 1 × 107 DU145 cells mixed with
Matrigel (BD Biosciences, Bedford, MA, USA) and used for experi- ments after the tumor size was larger than 20 mm3. The mice were assigned randomly into four groups.
Three therapeutic groups were treated via tail vein with
Re-188-labeled Herceptin (10 μCi/g body weight, number of mice [n] = 8), Re-188 (10 μCi/g body weight, n = 9), and Herceptin
(2.5 μg/g body weight, Herceptin concentration comparable
to Re-H, n = 8). The mice (n = 8) in the control group
were treated with 0.05 ml of normal saline via tail vein. The body weight and two perpendicular diameters of the tumor were measured twice per week with vernier calipers. Tumor volumes were calculated as mentioned previously
(π/6 × larger diameter × smaller diameter2). Tumor
growth
inhibition (TGI) was calculated as the ratio of the tumor volume in the treated group to that of the control group at a given time × 100. The mice were ethically killed when their tumors reached a volume of 2,000 mm3. The
animal use protocol has been reviewed and approved by the Institu- tional Animal Care and Use Committee in National Chung Hsing University, 96–91.
Bio-distribution of Re-188 labeled Herceptin
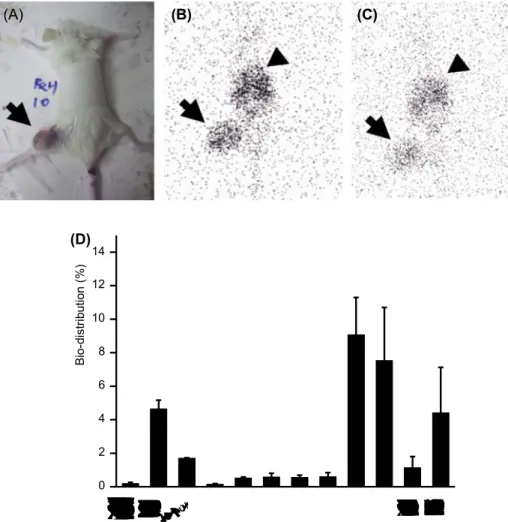
Gamma scintigraphy was used for the sequential, non-invasive evaluation of the in vivo tissue distribution of Re-188-labeled Herceptin in tumor-bearing mice. Six mice with a DU-145 xenograft on the left thigh were admin-istered 11.1 MBq of Re-H via the tail vein and sedated for scintigraphic evaluation 17 and 24 h later. The static images were acquired with a gamma scintillation camera equipped with a pinhole collimator (E-cam, Siemens,
Knoxville, TN, USA) for 5,000 counts. The matrix size was 128 × 128 with zoom 2. Photopeak was at 155 keV with 10% width. The images were evaluated by an experienced nuclear medicine physician. Evaluation by tissue counting: The six mice bearing DU-145 tumors on the left thigh used for scintigraphic evaluation were sacrificed 24 h after being injected with 11.1 MBq of Re-H , immediately
In t J R ad ia t B io l D ow nl oa de d fr om in fo rm ah ea lth ca re .c om b y T ai pe i V et er an s G en er al H os pi ta l o n 05 /0 1/ 13 F or p er so na l u se o nl y.
following completion of scintigraphic acquisition. The main organs, such as the liver, lung, spleen, kidney, testis, muscle, bone, blood, and tumor, were weighed and counted by gamma counter. The injected dose of Re-188-labeled Herceptin was counted as well. The bio-distribution of Re-188-labeled Herceptin is expressed as the percentage of the injected dose per gram of tissue (% injected dose/g).
West ern blotti ng
Protein expression in cell lysates was analyzed by Western blotting (30 μg/lane) as described previously (Hsu et al.
2011b). Antibodies against Her2, phospho-Her2, Cdk5, p35, PARP, Bax, Bcl2, tubulin (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and actin (Chemicon, Temecula, CA, USA) were used. Peroxidase-conjugated anti-mouse
or anti-rabbit antibodies (Jackson
ImmunoResearch Laboratory, West Grove, PA, USA) were used as the secondary antibody. ECL detection reagent (Perkin Elmer, Shelton, CT, USA) was used to visualize the immunoreactive proteins on PVDF membranes (Perkin Elmer, Shelton, CT, USA) after transfer using a Trans-Blot SD (Bio-Rad, Hercules, CA, USA).
S t a t i s t i c s
The data are presented as the means 士 standard deviation. For the apoptosis study, Student’s t-test was used to evaluate the differences between the tests and controls. Results with P values less than 0.05 were considered statistically significant.
R
e
s
u
l
t
s
Her2 levels in different cell linesThe protein levels of Her2 and phospho-Her2 in different prostate cell lines were surveyed; BT-474, a breast cancer cell line, was used as a positive control (Figure 1A). The normal prostate epithelial cell line, HPEpiC, showed low Her2/phospho-Her2 expression; BT-474 cell line showed high Her2/phospho-Her2 expression; DU145, an androgen- independent prostate cancer cell line without androgen receptor (AR) expression, showed high Her2/phospho-Her2 expression; LNCaP, an androgen-dependent prostate cancer cell line with AR
expression, showed high Her2/ phospho-Her2
expression; CWR22Rv1, an androgen- independent prostate cancer cell line with AR expression, showed moderate Her2/phospho-Her2 expression; and PC3, an androgen-independent prostate cancer cell line without AR expression, showed low Her2/phospho- Her2 expression. Previous studies from our lab indicate that Her2 is important to the growth of prostate cancer cells as the cells transition from an androgen-dependent to an androgen-independent state (Hsu et al. 2011a). Because DU145 is an androgen refractory cell line and has high Her2 expression, and a high phospho-Her2/Her2 ratio, it was suitable for this study.
Inhibitory effects of Re-188-labeled Herceptin on DU145 proliferation
Serial doses of Re-188-labeled Herceptin (Re-H), Rhenium-188 (Re-Rhenium-188), and Herceptin (H) were used to treat DU145
Figure 1. The effects of Herceptin (H), Re-188, and Re-188-labeled Herceptin (Re-H) on the proliferation of Her2-expressing prostate cancer cell lines. (A) Protein levels of Her2 and phospho-Her2 (Y1221/1222) in HPEpiC cells (a normal prostate epithelial cell line), four prostate cancer cell lines (DU145, PC3, LNCaP, CWR22Rv1), and BT-474 (a breast cancer cell line) were evaluated by Western blotting. Tubulin served as
an internal control. DU145 cells cultured in 24-well plates (20,000 cells/well) were treated with Re-H (1, 5, 10, 20, 40, 80, and 160 μCi), H (0.3125,
0.625, 1.25, 2.5, 5, 10 and 20 mg/ml), Re-188 (1, 5, 10, 20, 40, 80, and 160 μCi), and the control in triplicate. Three hours later, the supernatant
was removed. The cells were gently washed with PBS to remove non-binding agents. The appropriate medium was then added into each well, and the cells were incubated at 37°C for additional four days (B) and 6 days (C) before evaluating proliferation with the MTT assay. LNCaP (D), DU145 (E), and PC3 (F) were treated with Re-188 or Re-H (75, 150, and 300 μCi) as well as the control for 3 hours, and the cells were incubated for three additional days before measuring proliferation as described above. The data are presented as the percentages of viable cells to the control group. + and + +, p < 0.05 and p < 0.01 versus H group; ∗ and ∗∗, p < 0.05 and p < 0.01 vs. the group of concentration = 0; #, p < 0.05
versus Re-188 group.
cells for 3 h. Unbound drugs were then washed out, and the cells were cultured in fresh medium for an additional 4 or 6 days. Proliferation of the DU145 cells was evaluated by MTT assay. A significant inhibition of cell proliferation induced by Re-H after culturing for 4 days was observed at 160 μCi, while there was no significant inhibition after treat-ment with Re-188 or H (Figure 1B). The Re-H-triggered growth inhibition was more significant after 6 days of treatment and was dose-dependent (Figure 1C). Under
these conditions, the Re-188 group showed significant inhibition after high dosage treatments; however, the inhibi- tion triggered by Re-188 was still less than that of the Re-H
In t J R ad ia t B io l D o w n lo ad e d f ro m in fo rm ah e al th ca re .c o m b y Ta ip e i V et e ra n s G e n e ra l H o sp it al o n 0 5 /0 1 /1 3 F o r p er so n al u se o n ly .
treatments (at a dose of 160 μCi, the survival fraction was
83.9% vs. 70.9% on the 4th day and 22.8% vs. 19.9% on the
6th day, respectively). The H group demonstrated no effect on cell proliferation after the six-day treatment (Figure 1C).
Analysis of the effect of Re-188-labeled Herceptin on different cell lines
To differentiate the potential inhibitory effects of Re-H and Re-188 treatments on different prostate cancer cell lines, DU145, LNCaP, PC3, and their control counterparts were analyzed. The doses of Re-H and Re-188 administered
were 0, 75, 150, and 300 μCi. Herceptin treatment was not
350 H.-Y. Wang et al.
included because the data mentioned earlier demonstrated no correlation between the H group and cell proliferation (Figure 1B and 1C), in accordance with previous investiga-tions (Agus et al. 1999). The experimental procedure was similar to those in Figures 1B and 1C, except that the cells were now incubated for 3 additional days after treatment. The data are presented as the percentage of the control group and indicates that both Re-H and Re-188 showed dose-dependent inhibition of proliferation in the three cell lines (Figure 1D–1F). Additionally, the differences in growth inhibition between the Re-H and Re-188 treatments were greater in the DU145 and LNCaP cells (p < 0.01) than in the
PC3 cells (p < 0.05) at 300 μCi, which implies the possible
correlation between Her2/phospho-Her2 expression and drug response in prostate cancer
cells.
Re-188-labeled Herceptin induces apoptosis in DU145 cells Because the growth inhibition induced by Re-H was significant (Figure 1), we investigated the role of apoptosis during Re-H treatment. TUNEL staining and measurement of apoptosis-related proteins were performed to evaluate the effects of the four DU145 groups on apoptosis. Re-H (160 μCi), Re (160 μCi), H (20 mg/ml), and Control (c) were
treated in culture medium for 3 h and then replaced by fresh
medium and cultured for an additional 3 days; the cells were observed under a fluorescent microscope, and a green signal indicated apoptotic cells. The cell lysates were collected and analyzed by Western blotting. As seen in Figure 2A, Re-H treatment significantly increased apoptosis when compared to the control group and the Re and H treatment groups (Figure 2A). Apoptosis-related proteins, including cleaved PARP and Bax, increased after Re-H treatment, while the
anti-apoptotic protein Bcl-2 dramatically decreased (Figure 2B). This is the first demonstration indicating that Re-188-labeled Herceptin can directly cause apoptosis of cancer cells.
Effects of Re-188-labeled Herceptin on tumor growth in xenografts
Tumor growth in the xenograft model in the four treatment groups (Re-H, Re-188, H and C) was determined as described in Materials and methods. The tumor growth curves are shown in Figure 3A. A significant decrease in tumor growth was observed 13 days after Re-H injection when compared to the Re-188 and H groups, and the Re-188 and H groups showed a moderate but not significant decrease in tumor growth compared to the control group (Figure 3A) while the average body weights among four groups of animals showed no significant different (data not shown). Tumor growth inhibition (TGI) calculated as the ratio of tumor vol- ume (treatments/control) at a given time is shown in Table I. In contrast, the levels of apoptosis-related proteins in tumor samples from individual animals of the four groups were also monitored. The data indicate that Re-H treatment can increase the protein levels of cleaved PARP and Bax, and decrease Bcl-2 levels in tumors when compared to other groups (Figure 3B). Bax protein levels slightly increased after Re-188 treatment. Thus, our protein expression analysis sug- gests that Re-H treatment might cause apoptosis in tumor xenografts. Taken together, in vivo treatment of Re-H could effectively retard tumor growth, potentially through apoptosis.
Bio-distribution of Re-188-labeled Herceptin in mice
Since the in vivo treatments have systemic effects, bio-distribution of the isotope in the whole body is important
(A) TUNEL DAPI Control Re-H Re H (B) cleaved PARP Bax Bcl-2 In t J R ad ia t B io l D ow nl oa de d fr om in fo rm ah ea lth ca re .c om b y T ai pe i V et er an s G en er al H os pi ta l o n 05 /0 1/ 13 F or p er so na l u se o nl y.
Rhenium-188-labeled Herceptin and prostate cancer 3 5 4
Actin
Figure 2. 188-labeled Herceptin induces apoptosis in DU145 cells. DU145 cells were treated with 188-labeled Herceptin (H, 160 μCi), Re-188 (160 μCi), Herceptin (H, 20 mg/ml), or Control (c) for 3 hours. TUNEL staining (A) and Western blotting (B) were performed after three days of incubation. Apoptosis-related proteins were then detected. The green-fluorescent signal represents cells that underwent apoptosis. DAPI was used as a nuclear stain. Actin served as the internal control. This Figure is reproduced in color in the online version of International Journal of Radiation Biology.
(A) (B) C H Re ReH Mice ID: 1 2 3 5 6 7 1 2 3 4 5 6 1 2 3 4 6 7 8 1 2 3 6 cleaved PARP Bax Bcl-2 Actin
Figure 3. Tumor growth of the DU145 xenografts in SCID mice was inhibited exclusively by Re-188-labeled Herceptin treatment. SCID mice were treated with Herceptin (H, 2.5 μg/g body weight), Re-188 (10 μCi/g body weight), Re-188-labeled Herceptin (Re-H, 10 μCi/g body weight), and normal saline, used as the control (Ctrl), on day 34. (A) The tumor volume was recorded twice a week by calculating the tumor diameter. ∗ and
∗∗, p < 0.05 and p < 0.01 vs. the control group. (B) Apoptosis-related proteins in tumor samples from individual animals of four groups (indicated
by numbers) were detected by Western blotting. Actin served as the internal control.
to understand potential side-effects. Figure 4A showed the posture of the mouse while scintigraphic images were recorded. The site of the tumor xenograft is indicated by an arrow. The scintigraphic images of Re-H distribution 17 h and 24 h after injection were shown in Figure 4B and 4C, respectively. Radioactivity at the tumor site on the left thigh was significantly higher, revealing the specific binding of Re-H at the tumors (indicated by the arrows). In addition, the spleen and liver were determined to be the dose-limiting organs as observed by the accumulation of the tracer, indi- cated by the arrow heads. After image acquisition, the mice were sacrificed for isotope counting of individual organs. The bio-distribution of Re-H 24 h after intravenous injection is Table I. Tumor growth inhibition (TGI) induced by the treatments of Herceptin (H), Re-188 (Re), and Re-188 labeled Herceptin (Re-H) in xenografted model. The TGI was calculated as the ratio of relative volume of the xenograft in the treated to control group at a given time.
Days H Re Re-H 23 100.0% 100.0% 100.0% 27 76.7% 93.7% 103.5% 30 82.7% 77.7% 92.5% 34 140.1% 100.6% 131.2% 37 103.9% 70.6% 95.4% 41 120.2% 91.9% 110.4% 43 83.5% 61.2% 74.6% 48 85.9% 64.4% 55.1% 51 90.3% 70.1% 37.8% 55 96.7% 87.4% 36.6%
H, Herceptin; Re, Re-188; Re-H, Re-188 Herceptin.
In t J R ad ia t B io l D o w n lo ad e d f ro m in fo rm ah e al th ca re .c o m b y Ta ip e i V et e ra n s G e n e ra l H o sp it al o n 0 5 /0 1 /1 3 F o r p er so n al u se o n ly . Tu m o r si ze ( cm 3)
shown in Figure 4D. The relative high tumor uptake of iso- tope indicated specific binding at the tumor site. However, the dose-limiting organs, spleen and liver, were highly radioactive, as shown in Figure 4B and 4C, respectively. In addition, higher activities in kidney and urine resulted from the excretion of isotope via the urinary system.
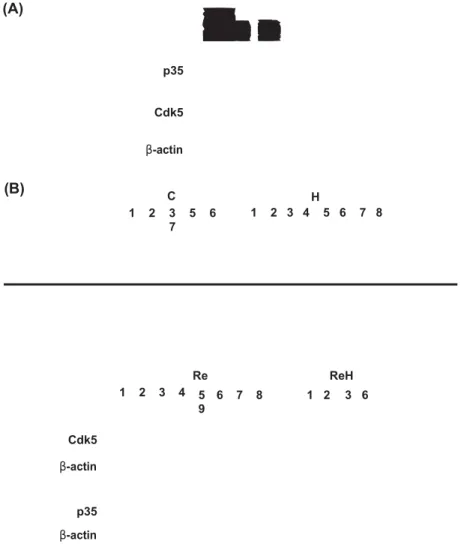
Effects of Re-188-labeled Herceptin on p35/Cdk5 proteins Cdk5 has been reported to be involved in the response of cells to radiation (Bolin et al. 2012). Previous studies from our laboratory have indicated that Cdk5 is important to the survival of cancer cells (Lin et al. 2004, 2007b, Hsu et al. 2011b). Therefore, we wanted to test whether Cdk5 mediated the decrease in the survival of prostate cancer cells after Re-H treatment. We monitored the protein levels of Cdk5 and its regulator, p35, both in vitro in DU145 cells and in vivo in tumor xenografts. Our analysis revealed a gradual decrease in the levels of Cdk5 and p35 proteins with
increasing doses of Re-H (Low: 80 μCi and High: 160 μCi for
3 h) (Figure 5A). In tumor xenografts, although the levels of
Cdk5 protein in tumors were not significantly different between control and treated groups, the levels of p35 protein in the Re-H-treated group dramatically decreased. Notably, the level of p35 protein in the Re-188-treated group also slightly decreased when compared to the control and H groups (Figure 5B). The data suggests that activation of p35/Cdk5 might mediate growth inhibition as a result of Re-188-labeled Herceptin treatment in prostate cancer cells.
352 H.-Y. Wang et al. (A) (B) (C) (D) 14 12 10 8 6 4 2 0
Figure 4. Immuno-specific binding of Re-188-labeled Herceptin to DU145 xenografts was determined by scintigraphy. (A) The image shows the posture of SCID mice during acquisition and the site of the DU145 xenograft on the left thigh. Scintigraphic images with a gamma scintillation
camera equipped with pinhole collimator (E-cam, Siemens) for 5,000 counts, matrix size of 128 × 128 with zoom 2, and a photopeak at 155 keV with
10% width were acquired 17 h (B) and 24 h (C) after 11.1 MBq of Re-188 Herceptin was injected via the tail vein. The arrows indicate the significant accumulation of radioactivity at the site of tumor xenograft on the left thigh, which confirmed the specific targeting of Re-188 Herceptin to the Her2-expressing tumor. The arrowheads indicate high tracer uptake at the dose-limiting organs, namely, the spleen and liver. (D) The bio- distribution of Re-188-labeled Herceptin in SCID mice with DU145 xenografts was evaluated at 24 h after injection. The organs and tumors were weighed and counted by gamma counter. The injected dose of Re-188-labeled Herceptin was counted as well. The values of bio-distribution of Re-188-labeled Herceptin were expressed as the percentage of the injected dose per gram of tissue (% injected dose/g). This Figure is reproduced in color in the online version of International Journal of Radiation Biology.
Discussion
Her2, a tyrosine kinase receptor, plays an essential role in the survival of several types of cells. Overexpression or mutation of Her2 can transform cells into cancer cells (Neve et al. 2001). In prostate carcinomas, elevated Her2 expression has been noted, especially in hormone-refractory prostate cancers (59–80% vs. 9–30%) (Shi et al. 2001). Refractory pros- tate carcinomas are also frequently metastatic with a poor prognosis. Therefore, novel treatments for prostate cancer such as targeted therapy are being developed. Here, we report Re-188-labeled Herceptin treatment as a potential alternative strategy for treating patients with refractory prostate cancer with Her2-targeted radiotherapy.
DU145, an androgen receptor-negative prostate cancer cell line with high Her2 expression (Figure 1A), was treated with different doses of radioactive Herceptin to determine the effective doses. Effective growth inhibition
is observed at high doses of treatment or after a long incubation time (Figure 1B and 1C). In addition to direct
In t J R ad ia t B io l D ow nl oa de d fr om in fo rm ah ea lth ca re .c om b y T ai pe i V et er an s G en er al H os pi ta l o n 05 /0 1/ 13 F or p er so na l u se o nl y. B io -d is tr ib ut io n ( % )
Rhenium-188-labeled Herceptin and prostate cancer 3 5 3
effects, this may be a result of the indirect cytotoxic effects of radiation inducing the production of free radicals (Chen et al. 2009, Luo et al. 2009). Since the first effect of radiation is to damage DNA and arrest cell proliferation, only cells with irreparable damage would die. This explains why the survival difference between the treated and control cells becomes more significant after longer incubation time even though the duration of the treatment is identical (Figure 1B and 1C). A previous report indicates that Herceptin is more effective in androgen-dependent than androgen-inandrogen-dependent cells (Agus et al. 1999). Here, we found that Herceptin treatment showed no significant growth inhibition on DU145 cells in contrast to the effects of Re-188-labeled Herceptin (Figure 1B and 1C). In addi- tion, the growth of DU145 cells in vitro was also affected by Re-188 treatment (Figure 1C). This might be due to the limited reaction environment in vitro; thus, beta rays emitted from the free Re-188 could damage adjacent cells without specifically binding to them. Our in vivo tumor studies support this hypothesis because the growth
(A) p35 Cdk5 β-actin (B) C 1 2 3 5 6 7 H 1 2 3 4 5 6 7 8 Re ReH 1 2 3 4 5 6 7 8 9 1 2 3 6 Cdk5 β-actin p35 β-actin
Figure 5. Cdk5 and p35 proteins in DU145 cells were altered by treatment with Re-188-labeled Herceptin both in vitro and in vivo. (A) DU-145 cells were treated for 3 hours as follows: control, low dose (80 μCi) of labeled Herceptin (low), and high dose (160 μCi) of Re-188-labeled
Herceptin (High). The cells were washed off the drugs and were incubated for three additional days before analyzing the protein levels by Western blotting. (B) The protein samples from the same experiment in Figure 3 were detected by Western blotting. Actin served as the internal control.
inhibitions of Re-188 and Herceptin treatments were com-parable, but significantly less than the decreased prolifera-tion observed with Re-188-labelled Herceptin treatment (Figure 3).
To further elucidate the correlation between Herceptin-mediated Her2 targeting and growth inhibition, Re-188 and Re-188-labeled Herceptin were administered to three pros-tate cancer cell lines (DU145, PC3, and LNCaP) exhibiting different Her2 expression levels. Expectedly, the growth dif-ferences of LNCaP and DU145 cells (with high levels of Her2 expression) between treatments of Re-188-labeled Hercep- tin and Re-188 were more significant when compared to the treatments in PC3 cells (with low Her2 expression) when
exposed to radiations below 300 μCi (Figures 1D–1F). In addition to Her2 expression levels, the levels of other ErbB members, such as epidermal growth factor receptor (EGFR), might also affect the efficiency of Her2 targeting because they are dimerization partners. EGFR itself can also be a treatment target in PC3, LNCaP, and DU145 (Sgambato et al. 2004, Telliez et al. 2007). Therefore, EGFR expression levels and the protein
interaction between EGFR and Her2 in different cells affect the efficiency of Herceptin inhibition. This may explain why we did not observe a Her2 dose- dependent effect for growth inhibition in different cell lines.
In t J R ad ia t B io l D o w n lo ad e d f ro m in fo rm ah e al th ca re .c o m b y Ta ip e i V et e ra n s G e n e ra l H o sp it al o n 0 5 /0 1 /1 3 F o r p er so n al u se o n ly .
In addition to Her2 expression levels, androgen depen- dency might also play a role in growth inhibition by Herceptin- targeted treatment. A previous report indicates that Her2 may modulate AR function through its
DNA binding and protein stability
(Mellinghoff et al. 2004). Our previous findings also suggest that Her2 is important in stabilizing AR during the transition from androgen dependence to independence in prostate cancer cells (Hsu et al. 2011a). Therefore, prostate cancer cells with androgen dependency, such as LNCaP, were more susceptible to Herceptin, whereas androgen- independent cells such as DU145 and PC3 were not. The high growth inhibition of LNCaP after Re-188-labeled Herceptin treatment might result from the high
Herceptin susceptibility to androgen
dependency and the fact that high Her2 expression increases the conjugation of Herceptin with lethal radioisotopes. For the androgen-independent cell line DU145, after treatment with Herceptin labeled with Re-188, there was an increase in growth inhibition although Hercep- tin alone was ineffective (Figure 1B and 1C). Radio-induced growth inhibition was also noted in the PC3 cell line, which is another androgen-independent prostate cancer cell line thought to be unsusceptible to Herceptin (Figure 1F). These data imply that Re-188-labeled Herceptin might be a poten- tial treatment for androgen-independent prostate cancer.
Previous studies have demonstrated that the major con-tribution of radiation-induced tumor shrinkage is apoptosis (Stephens et al. 1991, Meyn 1997). TUNEL staining and analysis of apoptosis-related proteins showed significant apoptosis of DU145 cells after Re-188-labeled Herceptin treatment compared to the control group (Figure 2). In addi- tion, in vivo results also indicated that the apoptosis-related proteins were significantly affected by Re-188-labeled Her- ceptin while Herceptin and Re-188 treatments alone did not show similar effects (Figure 3). Taken together with previous findings, our data suggests that the decrease of tumor size
might be induced by β radiation emitted from Re-188
conju-gated with
Herceptin.
We also explored the molecular mechanisms of Re-188-labeled Herceptin treatment to elucidate its inhibitory effects. Several lines of evidence have recently demonstrated that Cdk5 and its regulator, p35, play important roles in can- cer biology (Lin et al. 2007b, Eggers et al. 2011, Demelash et al. 2012). Our report indicates that hyper-activation of Cdk5 is important to determine the fate of prostate cancer cells (Lin et al. 2004). Subsequently, Cdk5 was found to have a significant role in controlling cell motility and metas- tasis in prostate cancer (Strock et al. 2006). Our recent study further illustrated that Cdk5 may regulate the androgen receptor through phosphorylation (Hsu et al. 2011b). In this study, we found that Re-188-labeled Herceptin treatment significantly decreased the levels of Cdk5 and its activator, p35, in DU145 cells. In addition, the inhibitory effects of Re-188-labeled Herceptin on p35 protein levels became more significant in tumor xenografts. Indeed, knock-down of Cdk5 or p35 resulted in a significant decrease of DU145 cell growth (data not shown). These data suggest that the decrease of p35/Cdk5 protein and its activity might mediate decreased prostate cancer cell proliferation upon treatment with Re- 188-labeled Herceptin.
In conclusion, the inhibitory effects in Her2-expressing and androgen-independent prostate cancer cells suggest that Re-188-labeled Herceptin may be therapeutically bene- ficial for Her2-expressing prostate cancer patients in the future.
Acknowledgements
The authors thank Dr S.L. Hsu and Ms M.C. Liu (Taichung Veterans General Hospital, Taiwan); Dr H.C. Chen, Dr T.H. Lee, Dr Y.M. Liou, Dr J.W. Chen and Dr C.M. Cheng (National Chung Hsing University, Taiwan) for technical support.
Declaration
of
interest
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
The work was supported in part by grants from the Taichung Veterans General Hospital Research Program (TCVGH-986703B), Taichung Veterans General Hospital/ National Chung Hsing University Joint Research Program (TCVGH-NCHU-1017607 to H. Lin), Taiwan National Science Council (NSC97-2320-B-005-002-MY3 to H. Lin), and Taiwan Ministry of Education under the ATU plan.
In t J R ad ia t B io l D o w n lo ad e d f ro m in fo rm ah e al th ca re .c o m b y Ta ip e i V et e ra n s G e n e ra l H o sp it al o n 0 5 /0 1 /1 3 F o r p er so n al u se o n ly .
References
Agus DB, Scher HI, Higgins B, Fox WD, Heller G, Fazzari M, Cordon-Cardo C, Golde DW. 1999. Response of prostate cancer to anti-Her-2/neu antibody in androgen-dependent and -independent human xenograft models. Cancer Research 59:4761–4764.
Ahmad I, Patel R, Singh LB, Nixon C, Seywright M, Barnetson RJ, Brunton VG, Muller WJ, Edwards J, Sansom OJ, Leung HY. 2011. HER2 overcomes PTEN (loss)-induced senescence to cause aggres-sive prostate cancer. Proceedings of the National Academy of Sciences of the USA 108:16392–16397. Albanell J, Codony J, Rovira A, Mellado B, Gascon P.
2003. Mechanism of action of anti-HER2 monoclonal antibodies: Scientific update on trastuzumab and 2C4. Advances in Experimental Medicine and Biology 532:253–268.
Bolin C, Boudra MT, Fernet M, Vaslin L, Pennaneach V, Zaremba T, Biard D, Cordelieres FP, Favaudon V, Megnin-Chanet F, Hall J. 2012. The impact of cyclin-dependent kinase 5 depletion on poly(ADP-ribose) polymerase activity and responses to radiation. Cellular and Molecular Life Sciences 69:951–962. Calvo BF, Levine AM, Marcos M, Collins QF, Iacocca
MV, Caskey LS, Gregory CW, Lin Y, Whang YE, Earp HS, Mohler JL. 2003. Human epidermal receptor-2 expression in prostate cancer. Clinical Cancer Research 9:1087–1097.
Chen KT, Lee TW, Lo JM. 2009. In vivo examination of (188)Re(I)- tricarbonyl-labeled trastuzumab to target HER2-overexpressing breast cancer. Nuclear Medicine and Biology 36:355–361.
Demelash A, Rudrabhatla P, Pant HC, Wang X, Amin ND, McWhite CD, Naizhen X, Linnoila RI. 2012. Achaete-Scute Homolog-1 (ASH1) stimulates migration of lung cancer cells through Cdk5/p35 pathway. Molecular Biology of the Cell 23:2856– 2866.
Dhavan R, Tsai LH. 2001. A decade of CDK5. Nature Reviews Molecular Cell Biology 2:749–759.
Dougall WC, Qian X, Peterson NC, Miller MJ, Samanta A, Greene MI. 1994. The neu-oncogene: Signal transduction pathways, transforma- tion mechanisms and evolving therapies. Oncogene 9:2109–2123. Eggers JP, Grandgenett PM, Collisson EC, Lewallen
ME, Tremayne J, Singh PK, Swanson BJ, Andersen JM, Caffrey TC, High RR, Ouellette M, Hollingsworth MA. 2011. Cyclin-dependent kinase 5 is amplified and overexpressed in pancreatic cancer and activated by mutant K-Ras. Clinical Cancer Research 17:6140–6150.
Formento P, Hannoun-Levi JM, Gerard F, Mazeau C, Fischel JL, Etienne-Grimaldi MC, Gugenheim J, Milano G. 2005. Gefitinib- trastuzumab combination on hormone-refractory prostate cancer xenograft. European Journal of Cancer 41:1467–1473.
Henry RY, O’Mahony D. 1999. Treatment of prostate cancer. Journal of Clinical Pharmacy and Therapeutics 24:93–102.
Hernes E, Fossa SD, Berner A, Otnes B, Nesland JM. 2004. Expression of the epidermal growth factor receptor family in prostate carci- noma before and during androgen-independence. British Journal of Cancer 90:449–454.
Hsu FN, Yang MS, Lin E, Tseng CF, Lin H. 2011a. The significance of Her2 on androgen receptor protein stability in the transition of androgen requirement in prostate cancer cells. American Journal of Physiology, Endocrinology and Metabolism 300:E902–908.
Hsu FN, Chen MC, Chiang MC, Lin E, Lee YT, Huang PH, Lee GS, Lin H. 2011b. Regulation of androgen receptor and prostate cancer growth by cyclin-dependent kinase 5. Journal of Biological Chemis- try 286:33141–33149.
Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ. 2007. Cancer
statistics, 2007. CA: A Cancer Journal for Clinicians 57:43–66. Lee KY, Rosales JL, Tang D, Wang JH. 1996. Interaction of
cyclin- dependent kinase 5 (Cdk5) and neuronal Cdk5 activator in bovine brain. Journal of Biological Chemistry 271:1538–1543. Legrier ME, Oudard S, Judde JG, Guyader C, de Pinieux G, Boye
K, de Cremoux P, Dutrillaux B, Poupon MF. 2007. Potentiation of antitumour activity of docetaxel by combination with trastuzumab in a human prostate cancer xenograft model and underlying mecha-nisms. British Journal of Cancer 96:269–276.
Lin H, Juang JL, Wang PS. 2004. Involvement of Cdk5/p25 in digoxin-triggered prostate cancer cell apoptosis. Journal of Biological Chemistry 279:29302–29307.
Lin H, Lin TY, Juang JL. 2007a. Abl deregulates Cdk5 kinase activity and subcellular localization in Drosophila neurodegeneration. Cell
Death and Differentiation 14:607–615.
Lin H, Chen MC, Chiu CY, Song YM, Lin SY. 2007b. Cdk5 regulates STAT3 activation and cell proliferation in medullary thyroid carci- noma cells. Journal of Biological Chemistry 282:2776–2784.
Luo TY, Tang IC, Wu YL, Hsu KL, Liu SW, Kung HC, Lai PS, Lin WJ. 2009. Evaluating the potential of 188Re-SOCTA-trastuzumab as a new radioimmunoagent for breast cancer treatment. Nuclear Medicine and Biology 36:81–88.
Mellinghoff IK, Vivanco I, Kwon A, Tran C, Wongvipat J, Sawyers CL. 2004. HER2/neu kinase-dependent modulation of androgen recep- tor function through effects on DNA binding and stability. Cancer Cell 6:517–527.
Meyn RE. 1997. Apoptosis and response to radiation: Implications for radiation therapy. Oncology (Williston Park) 11:349–356; discussion 356,361,365.
Morris MJ, Reuter VE, Kelly WK, Slovin SF, Kenneson K, Verbel D, Osman I, Scher HI. 2002. HER-2 profiling and targeting in prostate carcinoma. Cancer 94:980–986.
Neve RM, Lane HA, Hynes NE. 2001. The role of overexpressed HER2 in transformation. Annals of Oncology 12:S9–13.
Osman I, Scher HI, Drobnjak M, Verbel D, Morris M, Agus D, Ross JS, Cordon-Cardo C. 2001. HER-2/neu (p185neu) protein expression in the natural or treated history of prostate cancer. Clinical Cancer Research 7:2643–2647.
Patrick GN, Zukerberg L, Nikolic M, de la Monte S, Dikkes P, Tsai LH. 1999. Conversion of p35 to p25 deregulates Cdk5 activity and pro- motes neurodegeneration. Nature 402:615–622. Rasaneh S, Rajabi H, Babaei MH, Daha FJ. 2010. 177Lu labeling
of Herceptin and preclinical validation as a new radiopharmaceutical for radioimmunotherapy of breast cancer. Nuclear Medicine and Biology 37:949–955.
Sgambato A, Camerini A, Faraglia B, Ardito R, Bianchino G, Spada D, Boninsegna A, Valentini V, Cittadini A. 2004. Targeted inhibition of the epidermal growth factor receptor-tyrosine kinase by ZD1839 (‘Iressa’) induces cell-cycle arrest and inhibits prolif- eration in prostate cancer cells. Journal of Cellular Physiology 201: 97–105.
Sharkey RM, Goldenberg DM. 2011. Cancer radioimmunotherapy. Immunotherapy 3:349–370.
Shi Y, Brands FH, Chatterjee S, Feng AC, Groshen S, Schewe J, Lieskovsky G, Cote RJ. 2001. Her-2/neu expression in prostate can-cer: High level of expression associated with exposure to hormone therapy and androgen independent disease. Journal of Urology 166:1514–1519.
Signoretti S, Montironi R, Manola J, Altimari A, Tam C, Bubley G, Balk S, Thomas G, Kaplan I, Hlatky L, Hahnfeldt P, Kantoff P, Loda M. 2000. Her-2-neu expression and progression toward androgen inde- pendence in human prostate cancer. Journal of the National Cancer Institute 92:1918–1925.
Stavridi F, Karapanagiotou EM, Syrigos KN. 2010. Targeted therapeu- tic approaches for hormone-refractory prostate cancer. Cancer Treatment Reviews 36:122–130.
Stephens LC, Ang KK, Schultheiss TE, Milas L, Meyn RE. 1991. Apoptosis in irradiated murine tumors. Radiation Research 127:308–316.
Strock CJ, Park JI, Nakakura EK, Bova GS, Isaacs JT, Ball DW, Nelkin BD. 2006. Cyclin-dependent kinase 5 activity controls cell motility and metastatic potential of prostate cancer cells. Cancer Research 66:7509–7515.
Telliez A, Desroses M, Pommery N, Briand O, Farce A, Laconde G, Lemoine A, Depreux P, Henichart JP. 2007. Derivatives of Iressa, a specific epidermal growth factor receptor inhibitor, are powerful apoptosis inducers in PC3 prostatic cancer cells. ChemMedChem 2:318–332.
Tsai LH, Delalle I, Caviness VS Jr, Chae T, Harlow E. 1994. p35 is a neural-specific regulatory subunit of cyclin-dependent kinase 5. Nature 371:419–423.
Ziada A, Barqawi A, Glode LM, Varella-Garcia M, Crighton F, Majeski S, Rosenblum M, Kane M, Chen L, Crawford ED. 2004. The use of trastuzumab in the treatment of hormone refractory prostate cancer; phase II trial. Prostate 60:332–337.
In t J R ad ia t B io l D ow nl oa de d fr om in fo rm ah ea lth ca re .c om b y T ai pe i V et er an s G en er al H os pi ta l o n 05 /0 1/ 13 F or p er so na l u se o nl y.