INTRODUCTION
Recent studies have suggested that the continental shelves, although comprising <10% of the world ocean, may be very important in affecting global car-bon cycling due to their large standing stocks of organic carbon as well as high rates of primary
pro-duction (Mantoura et al. 1991, Biscaye et al. 1994, Jick-ells 1998, Wong et al. 2000). Heterotrophic bacterio-plankton are believed to play important roles in regu-lating accumulation, export, re-mineralization and transformation of the largest organic carbon pool (dis-solved organic carbon) in aquatic ecosystems over small (local) and large (global) scales (Cole et al. 1988, Michaels et al. 1994, Hansell & Carlson 1998, Carlson et al. 1999). In order to understand organic carbon © Inter-Research 2001
*E-mail: [email protected]
Differential coupling of bacterial and primary
production in mesotrophic and oligotrophic
systems of the East China Sea
Fuh-Kwo Shiah
1,*, Tzong-Yueh Chen
1, Gwo-Ching Gong
2, Chung-Chi Chen
3,
Kuo-Ping Chiang
4, Jia-Jang Hung
51Institute of Oceanography, National Taiwan University, Taipei, Taiwan, ROC 2Department of Oceanography, National Taiwan Ocean University, Keeloung, Taiwan, ROC
3National Center for Ocean Research, National Taiwan University, Taipei, Taiwan, ROC 4Department of Fishery Science, National Taiwan Ocean University, Keeloung, Taiwan, ROC 5Institute of Marine Geology and Chemistry, National Sun Yat-Sen University, Kaohsiung, Taiwan, ROC
ABSTRACT: Spatial patterns of integrated bacterial production (IBP; 9 to 179 mg C m–2d–1), bacter-ial biomass (IBB; 125 to 658 mg C m–2), bacterial turnover rates (Bµ = IBP/IBB; 0.03 to 0.37 d–1), pri-mary production (IPP; 18 to 2079 mg C m–2d–1) and hydrographical variables were measured in the continental shelf of the East China Sea (25.4 to 31.6° N, 120.5 to 127.0° E) during summer and autumn 1998. Sea surface temperatures were > 21°C in both seasons. The slopes for Log10IBP vs Log10IPP (summer 0.44 ± 0.09, autumn 0.52 ± 0.06) and Log10Bµ vs Log10IPP (summer 0.52 ± 0.06, autumn 0.50 ± 0.06) were significant with no seasonal difference. IBB showed no trend with IPP. When data were analyzed by separating the study area into the inner- (surface NO3 > 0.25 µM; mesotrophic) and outer- (surface NO3< 0.25 µM; oligotrophic) shelf systems, the slope values of Log10IBP vs Log10IPP derived from the inner-shelf (summer 0.48 ± 0.25, 12 stations; autumn 0.52 ± 0.09, 16 stations) data sets were significantly higher than those of the outer-shelf (summer 0.27 ± 0.13, 20 stations; autumn 0.36 ± 0.11, 26 stations); there was no difference for the slopes between seasons within each system. Mechanisms causing such differential coupling are not clear since this issue has seldom been addressed before. The results implied that care should be taken when analyzing cross-system or large spatial scale data sets. Our study also revealed that the IBP:IPP ratios (4 to 57%) of both seasons showed negative relationships with IPP. Phytoplankton effects on these ratios were dominated con-comitantly by algal turnover rate and biomass on the inner-shelf in summer and then shifted to algal biomass dominated over the whole shelf in autumn. Ecological and biogeochemical implications of our findings are discussed.
KEY WORDS: Bottom-up control · Continental shelf · Chlorophyll · East China Sea · Heterotrophic bacterioplankton · Inorganic nutrients · Kuroshio · Primary production
cycling and the microbial loop (Pomeroy 1974, Will-iams 1981, Azam et al. 1983), more detailed studies of which environmental factors regulate bacterial growth and, thus, carbon consumption in different marine provinces are needed.
In aquatic systems, the sources of substrate supply for bacteria may come from phytoplankton exudation, zooplankton sloppy feeding, excretion from other planktoners, release from dead particles and virus-induced lysis (see Ducklow & Carlson 1992, Fuhrman 1992 for review). In addition, substrates from alloch-thonous inputs such as river runoff and sediment resuspension also support substantial bacterial growth, particularly in the near-shore systems. Over longer time (seasonal) or spatial (across systems) scales, it has been suggested that primary production is the ‘ulti-mate’ factor in controlling bacterial growth (e.g. Cole et al. 1988, Conan et al. 1999 and citations therein), pri-marily due to the theory that phytoplankton exudation has been considered the most important source of organic substrate in supporting bacterial production. However, organic substrate supply rate from the non-algal sources mentioned above also co-vary with pri-mary production, indicating that bacteria-phytoplank-ton coupling might not necessarily be a ‘direct’ causal relationship. Moreover, up to present, we are still hav-ing difficulty quantifyhav-ing the relative importance of non-biogenic and biogenic components in contributing organic substrate for bacterial production in different ecosystems. The question that needs to be addressed is the complexity of the sources of organic substrate sup-ply in relation to the coupling of bacterial and primary production. Bacteria-phytoplankton coupling might occur due to external controlling factors that affect both bacteria and algae in a similar way (co-variation).
The East China Sea (ECS) has one of the largest shelf systems of the world. Coastal runoff, particularly from the Yantze River (aka The Chiangjiang) plume, has been recognized as having a significant seasonal im-pact on the ECS shelf, in terms of physical and chemi-cal hydrography as well as sources of inorganic nutri-ents and dissolved organic matter (Wong et al. 2000 and citations therein). The Three-Gorges Dam is pres-ently under construction in the mid-stream of the Yantze River, and its possible impacts on the biogeo-chemical cycles in the ECS shelf after it begins to oper-ate in 2009 have been of great concern. Any environ-mental data collected before its completion in this area may be very useful in evaluating its impacts after-wards. Four seasonal large area field surveys in the ECS shelf have been conducted since December 1997 by Taiwan’s Kuroshio Edge Exchange Process project (KEEP; Wong et al. 2000). Here we report on the data collected from the last 2 cruises conducted in summer and autumn 1998.
Several bacterial studies in the ECS have been previ-ously performed, but were limited to cold seasons. Shiah et al. (1999, 2000a,b) suggested that during cold seasons (winter and spring), bacterial growth on the in-ner-shelf was not limited by substrate supply but by temperature, while on the outer-shelf, the situation was the reverse. Intuitively, one may expect that bacterial growth during warm seasons should be predominately controlled by organic substrate supply over the whole shelf and a positive correlation between bacterial pro-duction (and turnover rate) and primary propro-duction should be observed. On the other hand, since the runoff of the Yantze River varies dramatically with season, a comparison of the bacteria-phytoplankton relationship during warm periods but with contrasting plume pat-terns caused by the Yantze River might provide some insight into the study of aquatic microbial ecology. As predicted above, here we show that bacterial produc-tion was highly coupled with primary producproduc-tion dur-ing warm periods. However, although the magnitude of coupling varied between mesotrophic (the inner-shelf) and oligotrophic (the outer-shelf) systems, the magni-tude of coupling seemed to be unchanged between summer and autumn within each system.
MATERIAL AND METHODS
Study site. Data were collected from the 2 cruises conducted in the continental shelf of the ECS north of Taiwan (Fig. 1) in July (34 stations) and November (32 stations) 1998. Seawater was collected from a SeaBird CTD-General Oceanic Rosette assembly with 20 l Go-Flo bottles. Light intensity was measured with a light meter (QSP200L; Biospherical). The depth of the euphotic zone (Ze) was defined as 1% of the surface light level. Surface nitrate concentrations (NO3) were used as the boundary of the inner- and outer-shelf sta-tions. For the outer-shelf stations, the surface NO3 con-centrations were all < 0.25 µM, which was the detec-tion limit of our nitrate analysis method (see below). In other words, the inner- and outer-shelf stations were located in eutrophic to mesotrophic and oligotrophic ecosystems, respectively. All of the measurements listed below were taken from the same depth within the Zefrom the same cast.
Bacterial production, abundance and turn-over rates. Bacterial abundance was measured by the acri-dine orange epifluorescence microscopy (Hobbie et al. 1977) method. Biomass was calculated using a carbon conversion factor of 2 ×10–14g cell–1(Lancelot & Billen 1984). Bacterial production was estimated by the method of 3H-thymidine (Fuhrman & Azam 1982) incorporation with a conversion factor of 1.18 × 1018 cells mol thymidine–1(Cho & Azam 1988). For details,
see also Shiah et al. (1999). For each station, triplicate samples were taken from 7 to 11 depths within the Ze. The integrated bacterial biomass (and pro-duction) was obtained by integrating (trapezoidal method) over Ze. Bacterial turnover rate (Bµ) was calculated by dividing integrated bacterial produc-tion (IBP) with integrated bacterial bio-mass (IBB). Note that the conversion factors were not determined empiri-cally and we assumed that they did not change with water mass and season.
Substrate enrichment experiments.
Experiments were performed on the
July cruise at the 3 stations (Fig. 1a) that represented the inner-, mid- and outer-shelf waters (Table 1). At each station, whole surface water samples were pre-incubated at in situ (Table 1) and 5 other temperatures (5 to 30°C) in 24, 2.0 l opaque polycarbonate bottles for 1 h, which was sufficient to allow adjustment to the new temperature. A dissolved free amino acid mixture (DFAA; 15 amino acids, final conc., 0.5 µM; Shiah et al. 2000b) was added into 12 bottles with duplicates for each temperature treatment after pre-incubation; the remaining 12 unenriched bottles were used as controls. Bacterial production and cell abundance were sampled every 2 to 6 h. The end of the lag period was defined as the time whenever the bacterial production readings in the DFAA treatments were statistically (usually > 20%) higher than those of the controls.
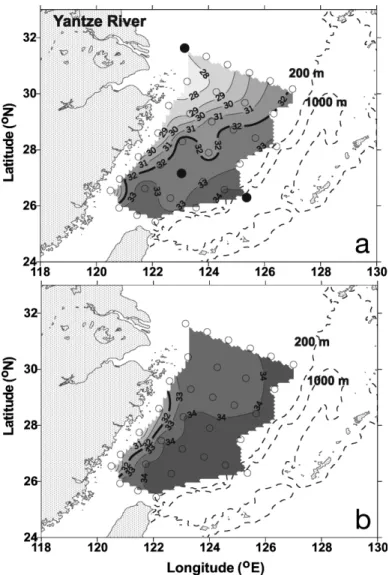
Primary production. Primary production was mea-sured by the 14C assimilation method (Parsons et al. 1984). In brief, 2 light and 1 dark 250 ml, acid cleaned polycarbonate bottles were filled with water samples taken from the Ze. After inoculation with H14CO3(final conc., 10 µCi ml–1), samples were incubated for 2 to 4 h in a self-designed tank with an artificial light source. Water temperature was maintained with running sea-water. To simulate light intensity, incubation bottles were wrapped with neutral density filters (LEE filters). Following retrieval, the water samples were immedi-ately filtered through Whatman 25 mm GF/F filters un-der low light and low pressure (<100 mm Hg). The fil-ters were then placed in scintillation vials, and 0.5 ml of 0.5 N HCl was added to remove residual HCO3. Radio-activity was measured in a liquid scintillation counter (Packard 1600) after the addition of 10 ml scintillation cocktail (Ultima Gold, Packard) into the vials. Phyto-plankton turnover rate was calculated as depth inte-grated primary production divided by depth-inteinte-grated chlorophyll concentrations with a conversion factor of 58 g C g–1chlorophyll a (chl a) (Eppley et al. 1992). Fig. 1. Contours of surface salinity with sampling stations (s)
in the East China Sea of (a) summer and (b) autumn 1998. (d) Substrate enrichment experiment stations are also shown. These experiments were performed on the summer-98 cruise only. Bold lines indicate the isohalines of 32.0 psu. Dashed
lines indicate bottom depth in meters
Item Units Inner-shelf Mid-shelf Outer-shelf
waters mixed waters waters
Temperature °C 21 27 29
Salinity psu 28.03 32.57 34.21
Nitrate µM 12.4 0.30 < 0.25a
Euphotic zone depth m 42 85 95
Chlorophyll mg chl m– 3 3.93 0.21 0.11 Primary mg C m– 3d–1 160.7 6.5 3.2 production Bacterial mg C m– 3d–1 5.86 1.95 0.83 production aDetection limit
Table 1. List of the in situ values of measured variables for samples used in sub-strate enrichment experiments in summer 1998
Chl a and nitrate concentrations and data analysis. Chl a and nitrate concentrations were measured fol-lowing the methods of Parsons et al. (1984). Water sam-ples for nutrient analyses were frozen immediately with liquid nitrogen in clean 100 ml polypropylene bot-tles. Nitrate was analyzed with a self-designed flow injection analyzer (Gong et al. 1995) and was reduced to nitrite with a cadmium wire, which was activated with a copper sulfate solution. The detection limit of this method was 0.25 µM. For chl a, 1 to 2 l of seawater were filtered through 25 mm Whatman GF/F filters, which were then immediately stored at –20°C. Back at the laboratory, the filters were ground in 10 ml 90% acetone followed by extraction in a 4°C shaking incu-bator for 2 h. After centrifugation at 1000 rpm (~200 × g) for 5 min, the concentrations of chlorophyll in the supernatant were measured on a Turner fluorometer (model 10-AU-005). The StatView IITM in Macintosh was used for statistical analysis including linear regression (model 2 for the field data; Sokal & Rohlf 1969), analysis of variance (ANOVA), analysis of covariance (ANCOVA) and multiple comparisons.
RESULTS
Seasonal changes in the Yantze River plume (salinity < 32.0 psu) could be identified by the patterns of
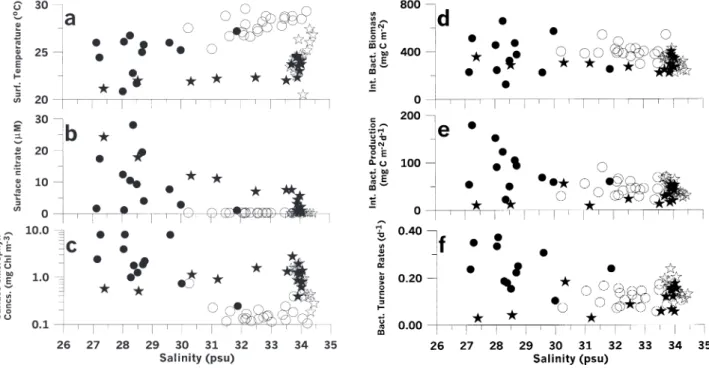
sur-face salinity (Fig. 1). In summer, the plume extended to the mid-shelf but in autumn the plume was observed only at a few stations adjacent to the China coast. In both seasons, the ECS shelf was covered with warm seawater with surface temperatures ranging from 21 to 30 and 21 to 27°C, respectively; sea surface tempera-tures were colder on the inner-shelf and increased off-shore (Fig. 2a). The depths of the euphotic zone were shallower than 10 m on the inner-shelf and extended down to 90 m on the outer-shelf. Surface nitrate con-centrations (NO3) in summer (< 0.2 to 28.1 µM) and autumn (< 0.2 to 24.3 µM) were high on the inner-shelf and then decreased seawards (Fig. 2b). Note that sur-face NO3 was depleted at a salinity of 30.0 psu during summer, while that of autumn was still detectable at a salinity of 34.0 psu. Surface chl a concentrations (Schl a) in summer (0.2 to 8.0 mg chl m– 3; Fig. 2c) varied about 40-fold, with most of the high values within the inner-shelf. Values of Schl a in autumn dropped to 0.2–2.8 mg chl m– 3but with the 1.0 mg chl m– 3isoline extending more toward the mid-shelf (area with salin-ity of 34 psu; Fig. 2c).
The euphotic zone integrated primary production (IPP; < 20 to 2080 mg C m–2d–1) basically followed the patterns of Schl a, with higher values (>1000 mg C m–2 d–1) recorded on the inner-shelf during summer. Autumn IPP values over the whole shelf were low, with values similar to the IPP recorded on the outer-shelf in
sum-Fig. 2. Scatter plots of surface values of (a) temperature, (b) nitrate, (c) chlorophyll a, and (d –f) bacterial measurements vs surface
salinity for the data derived from summer inner-shelf (d), summer outer-shelf ( ), autumn inner-shelf (
Q
) and autumn outer-shelf ( ) data. Panel (c) is in log10scale for better presentationmer. The values of the euphotic zone IBB (Fig. 2d) and IBP (Fig. 2e) in summer (125 to 658 mg C m–2and 22 to 179 mg C m–2d–1) were at least 50% higher than those of autumn (222 to 426 mg C m–2 and 9 to 70 mg C m–2d–1). For both seasons, IBB values were higher and formed dome-shaped patterns outside the Yantze River, and then decreased seawards. For both cruises, there were no significant relationships for IBB vs euphotic zone integrated chlorophyll concentrations (Ichl a; 14 to 68 mg chl m–2; p > 0.05) or IBB vs IPP (Fig. 3a).
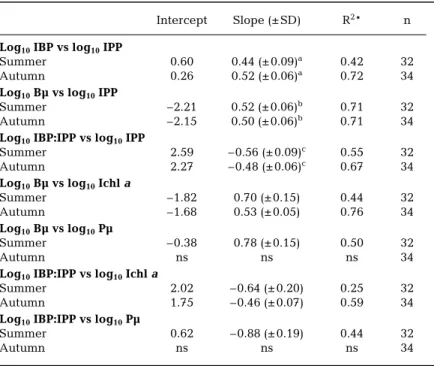
IBP of summer and autumn showed distinct spatial patterns (Fig. 2e). In summer, IBP was high on the in-ner-shelf and then decreased seawards; autumn IBP peaked in the mid-shelf (area of ~34.0 psu) with lower values at both ends. Spatial distribution of Bµ(Fig. 2f) in summer (0.06 to 0.37 d–1) and autumn (0.03 to 0.24 d–1) followed the same patterns of their IBP respectively. Both IBP and Bµ were positively correlated with IPP (Fig. 3b,c) with R2(coefficient of determination) values > 0.42 (Table 2). Note particularly that the slopes for log10IBP vs log10IPP and log10Bµ vs log10IPP calculated from the whole summer and autumn data sets were not significantly different from each other (Table 2). The ranges and averages of IBP:IPP ratio for summer and autumn were 4 to 36 and 4 to 57% as well as 15 ± 8 and 15 ± 10%, respectively. These ratios showed a negative trend with IPP (Fig. 3d) in both seasons. Further analy-sis showed that summer Bµ and IBP:IPP ratio were neg-ative functions of Ichl a and Pµ (algal turnover rates)
simultaneously, while those of autumn were negatively correlated with Ichl a only (Table 2).
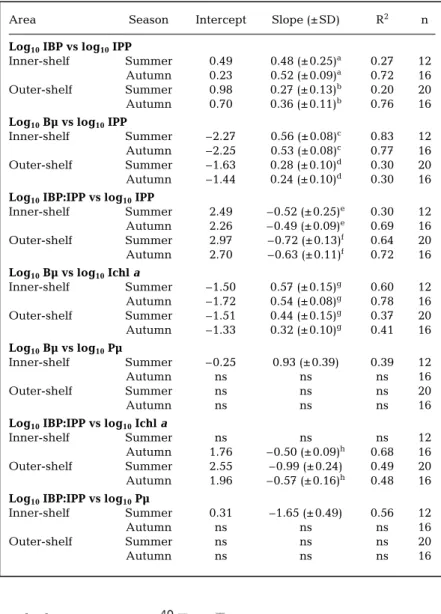
The relationships of IBP, Bµ and IBP:IPP ratio vs IPP revealed more interesting information when regional (inner- and outer-shelf systems) differences were con-sidered. For the inner-shelf, the slope values of IBP vs IPP of summer (0.48 ± 0.25) and autumn (0.52 ± 0.09) were about 2× those derived from the outer-shelf (0.27 ± 0.13 and 0.36 ± 0.13). Furthermore, in neither the inner- nor the outer-shelf was there a difference between seasonal slopes (Table 3). The relationships for Bµ vs IPP and IBP:IPP ratio vs IPP were the same as those described above except that the slopes of the latter were negative and higher on the outer-shelf (–0.72 ± 0.13 and –0.63 ± 0.11) than those of inner-shelf (–0.52 ± 0.25 and –0.49 ± 0.09) for both seasons.
The in situ conditions for the DFAA enrichment
ex-periments performed at the 3 stations on the summer cruise are listed in Table 1. Dramatic contrasts could be observed among experimental stations, both in terms of chemical and biological measurements. When incu-bated at in situ temperatures, bacteria responded to
the addition of DFAA within 2 h for the mid- and outer-shelf samples, while it took a further 2 h to occur in the inner-shelf samples (Fig. 4). As incubation
tempera-tures decreased (< 20°C), the lag periods (required for bacteria to respond to the added DFAA) increased in all experiments, indicating that the enrichment effect required a longer time to occur at low temperatures (5 to < 20°C).
DISCUSSION AND CONCLUSION
The continental shelf of the ECS during the warm periods showed strong gradients of inorganic nutri-ents, algal biomass and primary production. For both seasons, NO3concentrations were always high on the inner-shelf area (Fig. 2b), but values of Schl a (Fig. 2c)
and IPP (Fig. 3a) in autumn were only 50% of those recorded in summer. Several studies have shown that the Yantze River and the other 7 Chinese rivers that
Fig. 3. Scatter plots of (a) depth-integrated bacterial biomass (IBB), (b) production (IBP), (c) turnover rate (Bµ = IBP/IBB) and (d) the ratio of IBP:IPP vs depth-integrated primary production (IPP) for the data derived from summer inner-shelf (d), summer outer-shelf ( ), autumn inner-shelf (
Q
) and autumndischarge into the ECS shelf are characterized by a high inorganic N/P ratio ((NO3+ NO2)/PO4; > 40 mol N mol–1 P; Wong et al. 2000 and citations therein) and that such imbalances in the N/P ratio might result in PO4-limitation of the growth of phytoplankton (Gong et al. 1996, Wong et al. 1998). Higher Schl a
concentra-tions and IPP observed on the inner-shelf during sum-mer might be due to coastal upwelling induced by the prevalence of the southwest monsoon that increases PO4 availability from the bottom water (Gong et al. 2001). Surface PO4 values on the inner-shelf during summer were above the detection limit (0.03 µM), with a mean concentration and N/P ratio of 0.14 ± 0.14 µM and 75 ± 52 mol N mol–1P, respectively. On the other hand, surface PO4and N/P ratio of the inner-shelf dur-ing autumn were 0.46 ± 0.37 µM and 17 ± 8 mol N mol–1 P, respectively. Higher surface PO
4 concentra-tions on the inner-shelf during autumn might be due to lower values of Schl a and IPP (lower phytoplankton
uptake) resulting from low level sunlight intensity (Gong et al. 2001). Note also that the N/P ratios were similar to the Redfield ratio (N/P = 16) during autumn while those for the summer were much higher (N/P ranged from 22 to 167). For the oligotrophic outer-shelf, NO3concentrations were low and its availability
has been suspected to be the major limiting factor for algal growth (Chen et al. 1999, Chen 2000, Gong et al. 2000, 2001) since most of the surface PO4 values were still detectable with N/P ratios lower (N/P < 11) than the Redfield ratio.
These results indicate that the inner-and outer-shelf were more akin to 2 dis-tinct ecosystems dominated by new and regenerated production, respectively (Chen et al. 1999). Such dissimilarity might result in different phytoplankton species composition with mainly large phytoplankton (diatoms) on the inner-shelf and small phytoplankton (flagel-lates) on the outer-shelf (Chiang et al. 1997, Huang et al. 1999, Chen 2000). This implies that care should be taken when analyzing the relationship be-tween bacterial and primary production with large spatial scale data sets. More specifically, the bacteria-phytoplankton relationship might be system-depen-dent (see discussion below).
Seawater temperatures over the study area were all > 20°C, which were (or near to) the optimal temperature for the growth of non-hyperthermo-philic bacteria (c.f. Wiebe et al. 1992, Shiah et al. 2000b and citations therein). These systems offered a good opportunity to verify the relationship between bacterial growth (particularly turnover rate) and primary production when temperature was not limiting. Although the ranges and spatial patterns of the Yantze River freshwater plume of these 2 cruises were quite distinct, the coupling between bacterial rate parameters (IBP and Bµ) and IPP were significant with similar slopes and intercepts when the whole data sets were used in the analysis (Table 2).
Table 3 indicates that after further analysis of our data sets, the magnitude of coupling, as judged from the slope values, could be quite different on the inner- than on the outer-shelf, which represent meso- and oligotrophic systems, respectively. The results revealed that the algal effects on the spatial variability of IBP (and Bµ) on the inner-shelf were larger (i.e. 2×) than those on the outer-shelf. Such a phenomenon consistently appeared in sum-mer and autumn, although IPP (sumsum-mer 950 ± 600, autumn 292 ± 231 mg C m–2d–1) and Pµ (summer 0.39 ± 0.10, autumn 0.19 ± 0.03 d–1) on the inner-shelf changed dramatically within these 2 seasons. Conan et al. (1999, their Table 3) summarized the IBP-IPP relationship from their research, and the research of many others, and found that on an areal basis, the slope values of log10IBP
Intercept Slope (± SD) R2* n Log10IBP vs log10IPP
Summer 0.60 0.44 (± 0.09)a 0.42 32 Autumn 0.26 0.52 (± 0.06)a 0.72 34 Log10Bµ vs log10IPP
Summer –2.21 0.52 (± 0.06)b 0.71 32 Autumn –2.15 0.50 (± 0.06)b 0.71 34 Log10IBP:IPP vs log10IPP
Summer 2.59 –0.56 (± 0.09)c 0.55 32 Autumn 2.27 –0.48 (± 0.06)c 0.67 34 Log10Bµ vs log10Ichla
Summer –1.82 0.70 (± 0.15) 0.44 32
Autumn –1.68 0.53 (± 0.05) 0.76 34
Log10Bµ vs log10Pµ
Summer –0.38 0.78 (± 0.15) 0.50 32
Autumn ns ns ns 34
Log10IBP:IPP vs log10Ichla
Summer 2.02 –0.64 (± 0.20) 0.25 32
Autumn 1.75 –0.46 (± 0.07) 0.59 34
Log10IBP:IPP vs log10Pµ
Summer 0.62 –0.88 (± 0.19) 0.44 32
Autumn ns ns ns 34
Table 2. List of linear regression analysis (model 2) of euphotic zone integrated bacterial measurements (dependent variables) on euphotic zone integrated phy-toplankton variable (independent variable). SD: standard deviation; R2: coeffi-cient of determination; and n: sampling size. Superscripts (a–c) indicate no dif-ference at p = 0.05 by ANCOVA. IBP: depth-integrated bacterial production; Bµ: bacterial turnover rate; IPP: depth-integrated primary production; Ichl a,
depth-integrated chl a; and Pµ: phytoplankton turnover rates; ns: non-significant at
vs log10IPP were in the range of 0.37 to 0.75. Our slopes (0.27 to 0.52, Table 3) were within but at the lower end of the reported values. The most unique find-ing of our study that has not been fully addressed previously by many other re-searchers was that the coupling be-tween bacterial and primary production might be system-dependent.
Several recent studies have demon-strated that phytoplankton species composition, algal exudation capac-ity (Huang et al. 1999), bacterivory (Sanders et al. 1992) and bacterial community structure might change with different inorganic nutrient sta-tus (Lebaron et al. 1999) and seasons (Pinhassi & Hagrstrom 2000 and cita-tions therein). Vrede (1999) showed that algal species composition could affect bacterial growth. Therefore, the relationship (the slopes) of IBP vs IPP might be expected to deviate substan-tially by the changes of any of the factors on the inner- and outer-shelf systems. Unfortunately, we have not found any plausible explanation due to the lack of planktonic species com-position data.
It is well known that bacterial growth also may be limited by inor-ganic nutrients such as ammonium (NH4) and/or PO4 (Thingstad et al. 1999, Touratier et al. 1999 and cita-tions therein) when the C/N, C/P or N/P ratios of the organic substrate were higher than those of bacteria. A more intriguing idea is that the
differ-ential coupling between bacteria and phytoplankton on the inner- and outer-shelf might be regulated by mechanisms independent of the organic substrate con-trol scenario proposed by Cole et al. (1988) and others. That is, bacterial and algal activities might be driven by external sources in a similar manner (co-variation). In enrichment experiments, Shiah (1999) demonstrated that bacterial growth in the surface water samples taken from the outer-shelf (the Kuroshio waters) dur-ing summer was not limited by organic carbon (glu-cose) but by NH4and/or PO4. His experiments, unfor-tunately, did not further differentiate which inorganic nutrient was the limiting one. With his and our DFAA experiment results, we suspect that bacterial growth on the outer-shelf was more likely to be limited by nitrogen instead of PO4 for one important reason. If PO4 were the limiting nutrient, then the addition of
Area Season Intercept Slope (± SD) R2 n Log10IBP vs log10IPP
Inner-shelf Summer 0.49 0.48 (± 0.25)a 0.27 12 Autumn 0.23 0.52 (± 0.09)a 0.72 16 Outer-shelf Summer 0.98 0.27 (± 0.13)b 0.20 20 Autumn 0.70 0.36 (± 0.11)b 0.76 16 Log10Bµ vs log10IPP
Inner-shelf Summer –2.27 0.56 (± 0.08)c 0.83 12 Autumn –2.25 0.53 (± 0.08)c 0.77 16 Outer-shelf Summer –1.63 0.28 (± 0.10)d 0.30 20 Autumn –1.44 0.24 (± 0.10)d 0.30 16 Log10IBP:IPP vs log10IPP
Inner-shelf Summer 2.49 –0.52 (± 0.25)e 0.30 12 Autumn 2.26 –0.49 (± 0.09)e 0.69 16 Outer-shelf Summer 2.97 –0.72 (± 0.13)f 0.64 20 Autumn 2.70 –0.63 (± 0.11)f 0.72 16 Log10Bµ vs log10Ichl a
Inner-shelf Summer –1.50 0.57 (± 0.15)g 0.60 12 Autumn –1.72 0.54 (± 0.08)g 0.78 16 Outer-shelf Summer –1.51 0.44 (± 0.15)g 0.37 20 Autumn –1.33 0.32 (± 0.10)g 0.41 16 Log10Bµ vs log10Pµ Inner-shelf Summer –0.25 0.93 (± 0.39) 0.39 12 Autumn ns ns ns 16 Outer-shelf Summer ns ns ns 20 Autumn ns ns ns 16
Log10IBP:IPP vs log10Ichla
Inner-shelf Summer ns ns ns 12
Autumn 1.76 –0.50 (± 0.09)h 0.68 16 Outer-shelf Summer 2.55 –0.99 (± 0.24) 0.49 20 Autumn 1.96 –0.57 (± 0.16)h 0.48 16 Log10IBP:IPP vs log10Pµ
Inner-shelf Summer 0.31 –1.65 (± 0.49) 0.56 12
Autumn ns ns ns 16
Outer-shelf Summer ns ns ns 20
Autumn ns ns ns 16
Table 3. As in Table 2 but for data sets that were separated into inner- and outer-shelf areas. Slopes with the same superscripts (a–i) indicate no difference at p =
0.05 by ANCOVA
Fig. 4. Lag period required for bacteria to respond to the enrichment of dissolved free amino acids at different temper-atures. Experiments were performed in summer 1998 using samples taken from the inner- ( ), the mid- (■) and the
DFAA (which contained no P) would not enhance bac-terial growth. The low inorganic N/P ratios observed on the outer-shelf suggested that algal growth might be limited by inorganic nitrogen. It is very likely that inorganic nitrogen, particularly NH4 availability, was the limiting factor in regulating the spatial variability of bacteria and phytoplankton growth simultaneously on the outer-shelf. The system becomes more olig-otrophic as one moves from the mid- to the outer-shelf, and both IBP and IPP decrease due to reducing NH4 availability, which results in a positive relationship of IBP and IPP. Meanwhile, bacteria are more competi-tive for NH4uptake due to their much larger surface to volume ratio than phytoplankton (Bratbak & Thingstad 1985), and this might lead to a higher IBP:IPP ratio in a lower IPP area.
On the other hand, inorganic nutrient limitation on bacterial growth on the inner-shelf seemed to be less likely due to copious NH4(0.34 to 1.31 µM; Liean 1999)
and PO4 (summer 0.14 ± 0.14 µM; autumn 0.46 ±
0.37 µM) in the surface waters. Dissolved organic car-bon (DOC) concentrations in this area were high (85 to 120 µM) in the Yantze River plume and decreased sea-wards to concentrations of < 70 µM (Hung et al. 2001). DOC degradation rate for the coastal waters was about 0.05 d–1, suggesting that some portions of riverine DOC were labile to semi-labile (Hung et al. 2001). The DFAA enhancement effect on the coastal waters might be due to the addition of organic nitrogen instead of organic carbon. In these experiments, the lag periods of the in-ner-shelf samples (Fig. 4) were about 2×(4 h) longer than those of the mid- and outer-shelf samples, while the lag periods of the latter 2 samples showed no differ-ence from each other (t-test, p > 0.05, n = 6). This
im-plied that the strength of bottom-up control probably was weaker in the inner-shelf (Sanders et al. 1992), where surface chlorophyll (3.9 mg chl m– 3) and primary production (161 mg C m– 3d–1) were at least 10×higher than those of the other 2 areas (Table 1). Another possi-bility, as stated above, was that the in situ temperature
of the inner-shelf sample (21°C) was lower than those of the mid- and outer-shelf (27 to 29°C) samples.
As for bacterial biomass, no correlation was ob-served for IBB vs IPP. This suggests that processes other than substrate supply, such as bacterivory (Fuhrman & McManus 1984, Sherr et al. 1987, Sanders et al. 1992) and viral lysis (Proctor & Fuhrman 1990, 1991), could be important in regulating the spatial pattern of IBB. This is consistent with the conclusion of Ducklow (1999) that bacterial biomass and produc-tion might be independently regulated by different processes. Fig. 2d shows that in the low salinity (< 33.5 psu) area, summer bacteria biomass (370 ± 164 mg C m–2) was slightly higher than that of autumn (306 ± 52 mg C m–2), indicating that there might be
more riverine bacteria imported into the shelf by the Yantze River plume during summer. Note that the IBP recorded on the inner-shelf during summer (88 ± 46 mg C m–2d–1) was about 2.7×that of autumn (33 ± 18 mg C m–2d–1) while IBB of the former was only 21% higher than that of the latter. The abundance of het-erotrophic ciliates (Lin 2000) recorded on the inner-shelf during summer (40 to 60 × 104cells m– 3) was at least 4×higher than that of the autumn (5 to 15 × 104 cells m– 3), indicating the possibility of higher ciliate grazing pressure on IBB during summer. In addition, abundant ciliate numbers imply that there might be higher organic substrate flux from ciliates in support-ing higher Bµ (Nagata & Kirchman 1992) on the inner-shelf during summer than in autumn.
Although the slope values Bµ vs IPP (= Ichl a× Pµ) were similar between summer and autumn within each system, Table 3 reveals more subtle information with regard to their relationship. The spatial variation of Bµ derived from different systems and seasons were al-ways related to the changes of Ichl a instead of Pµ, with
1 exception. Note that Pµ on the inner-shelf during summer (0.39 ± 0.10) was about 2×higher than those recorded on the inner-shelf during autumn (0.19 ± 0.03 d–1), the outer-shelf during summer (0.23 ± 0.07 d–1) and the outer-shelf during autumn (0.18 ± 0.04 d–1). This indicates that Pµ effects on Bµ occurred only in the system when Pµ was high (see also below).
Ratios of IBP:IPP varied 4 to 57% for both cruises, with an overall average of 15 ± 9%. These values were somewhat lower than, but still within the range of, a review paper of oceanic IBP:IPP ratio (25%) presented by Ducklow (1999). In a study conducted in the NW Mediterranean Sea, Conan et al. (1999) found that the relationship between IBP:IPP ratio and chlorophyll normalized primary production (PE = IPP/Ichl a, an
index of Pµ) could be expressed as a power function of (IBP/IPP) ratio = 25.1 ×PE– 0.68. In addition, for a large range of PE >1.0 g C g–1chl h–1(=0.21 d–1of Pµ of this study), the ratio remained low (< 25%), but increased dramatically for lower values of PE. Our summer inner-shelf data coincided with their observation but further showed that Ichl a also plays a role in affecting the
variation of IBP:IPP ratio (Table 3). This suggests that phytoplankton effects on IBP:IPP ratio might be medi-ated through either algal biomass, turnover rate con-trols or both, and that such concon-trols might shift be-tween seasons and systems. As mentioned above, Pµ on the inner-shelf during summer were high (0.23 to 0.57 d–1) while most of (40 out of 51) the Pµ values derived from the other seasons and systems were < 0.23 d–1. It was suspected that algal turnover rate control on IBP:IPP ratio might occur only when Pµ was high, such as in the case of Conan’s et al. (1999) study and our summer inner-shelf data. On the other hand,
when Pµ values were low, algal biomass control on the spatial variation of IBP:IPP ratio probably would be more important. The same kind of argument could probably also be applied to the relationships for Ichl a
and Pµ on Bµ (Table 3).
The negative relationship between IBP:IPP ratios and IPP has been observed in the cold Southern Ocean (< 3°C; 47 to 60° S, Lochte et al. 1997, their Fig. 8b). Such a phenomenon was ascribed to the uncoupling or time lag between phytoplankton and bacteria development. Lochte et al. (1997) proposed several possible scenarios as an explanation, which included extremely low exu-dation rate of DOC by phytoplankton, low bacterial up-take or conversion of organic material, and repression of bacterial metabolism due to low temperatures or high grazing pressure. In our study, the low temperature sce-nario seemed not to be the case since water tempera-tures were all > 21°C during study periods and no corre-lation was observed for summer (n = 34) and autumn (n = 32) IBP:IPP ratios vs water temperatures or salinity (p > 0.05). Nevertheless, our data could not verify any of the other explanations but do document comparative find-ings in a very distinct ecosystem (i.e. temperate-sub-tropical). In fact, uncoupling (between phytoplankton and bacteria) is a phenomenon that has been observed through time-series study only (e.g. Ducklow 1999, his Figs. 2, 3 & 5). Our investigations were conducted in a way to address spatial variation. Nevertheless, from the organic carbon cycling point of view, as suggested by Conan et al. (1999), a high IBP:IPP ratio may lead to a consequence that there may be less material available for higher trophic level and/or for export to the deep ocean and sediments.
In summary, inorganic nutrient, phytoplankton bio-mass and production showed strong gradients in the continental shelf of the ECS during warm seasons with high seasonal contrast in the spatial patterns of the Yantze River plume. High seawater temperature (> 21°C) seemed to be the ultimate factor driving bac-terial growth, which was strongly coupled to primary production over the ECS shelf. However, our major finding that has not been fully addressed before sug-gests that algal effects on bacterial rate parameters might be system-dependent, as indicated by the differ-ent slopes for IBP (and Bµ) vs IPP from the inner- and outer-shelf systems. This implies that care should be taken in analyzing cross-system data sets derived from large spatial scale studies. Our study further indicates that phytoplankton effects on bacterial turnover rate might be concomitantly via algal biomass and turnover rate control on the inner-shelf in summer, and via algal biomass control on the outer-shelf in summer and over the whole shelf in autumn. The phenomena derived from the inner-shelf during warm seasons are quite different from previous studies (Shiah et al. 1999,
2000b) performed in the same area during cold sea-sons, stating that temperature was the major limiting factor in controlling bacterial spatial variation. This suggests that temperature, organic substrate and inor-ganic nutrient controls on bacterial growth in the ECS shelf might switch among locations within season and among seasons. The IBP:IPP ratios (15 ± 9%) recorded from this study are in the range of reported values of the world ocean. The negative correlation between IBP:IPP ratios and IPP has been reported in several ecosystems, but its ecological implications are not very clear and must be further explored.
Acknowledgements. Support for this research was provided by
the National Science Council, Taiwan. Cruise assistance from the crew of the RV ‘Ocean Researcher I’ and the valuable com-ments from 2 anonymous reviewers are deeply appreciated.
LITERATURE CITED
Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F (1983) The ecological role of water-column microbes in the sea. Mar Ecol Prog Ser 10:257–263 Biscaye PE, Flagg CN, Falkowski PG (1994) The shelf-edge
exchanges processes experiment, SEEP-II: an introduction to hypotheses, results and conclusions. Deep-Sea Res 41(2/3):231–252
Bratbak G, Thingstad TF (1985) Phytoplankton-bacteria inter-actions: an apparent paradox? Analysis of a model system with both competition and commensalism. Mar Ecol Prog Ser 25:23–30
Carlson CA, Bates NR, Hansell DA, Ducklow HW (1999) Esti-mation of bacterial respiration and growth efficiency in the Ross Sea, Antarctica. Aquat Microb Ecol 19:229–244 Chen LYL (2000) Comparisons of primary productivity and
phytoplankton size structure in the marginal regions of southern East China Sea. Cont Shelf Res 20(4/5):437–458 Chen LYL, Lu HB, Shiah FK, Gong GC, Liu KK, Kanda J (1999) New production and F-ratio on the continental shelf of the East China Sea: comparisons between nitrate inputs from the subsurface Kuroshio Current and the Chiang-jiang River. Estuar Coast Shelf Sci 48:59–75
Chiang KP, Shiah FK, Gong GC (1997) Distribution of summer diatom assemblages in and around a local upwelling in the East China Sea northeast of Taiwan. Bot Bull Acad Sin 38:121–129
Cho BC, Azam F (1988) Major role of bacteria in biogeochem-ical fluxes in the ocean’s interior. Nature 332:441–443 Cole JJ, Findlay S, Pace ML (1988) Bacterial production in
fresh and saltwater ecosystems: a cross-system overview. Mar Ecol Prog Ser 43:1–10
Conan P, Turley C, Stutt E, Pujo-Pay M, Van Wambeke F (1999) Relationship between phytoplankton efficiency and the pro-portion of bacterial production to primary production in the Mediterranean Sea. Aquat Microb Ecol 17: 131–144 Ducklow HW (1999) The bacterial content of the ocean
euphotic zone. FEMS Microbiol Ecol 30:1–10
Ducklow HW, Carlson CA (1992) Oceanic bacterial produc-tion. In: Marshall KC (ed) Advance in microbial ecology. Plenum Press, New York, p 113–181
Eppley RW, Chavez FP, Barber RT (1992) Standing stocks of particulate carbon and nitrogen in the equatorial Pacific at 150oW. J Geophys Res 97(C1):655–661
Fuhrman JA (1992) Bacterioplankton roles in cycling of organic matter: the microbial food web. In: Falkowski PG, Woodhead AD (eds) Primary productivity and biogeo-chemical cycles in the sea. Plenum Press, New York, p 361–383
Fuhrman JA, Azam F (1982) Thymidine incorporation as a measurement of heterotrophic bacterioplankton produc-tion in marine surface waters: evaluaproduc-tion and field results. Mar Biol 66:109–120
Fuhrman JA, McManus GB (1984) Do bacteria-sized marine eukaryotes consume significant bacterial production? Sci-ence 224:1257–1260
Gong GC, Liu KK, Pai SJ (1995) Prediction of nitrate concen-tration from two end member mixing in the Southern East China Sea. Cont Shelf Res 15:827–842
Gong GC, Chen LYL, Liu KK (1996) Chemical hydrography and chlorophyll distribution in the East China Sea during summer: implications in nutrient dynamics. Cont Shelf Res 16:1561–1590
Gong GC, Shiah FK, Liu KK, Wen YH, Liang MH (2000) Spatial and temporal variation of chlorophyll a, primary productivity and chemical hydrography in the southern East China Sea during summer: implications in nutrient dynamics. Cont Shelf Res 20(4/5):411–436
Gong GC, Wen YH, Wang BW, Liu GJ (2001) Seasonal varia-tion of chlorophyll a concentravaria-tion, primary producvaria-tion and environmental conditions in the subtropical East China Sea: prior to the operation of the Three-Gorges Dam. Deep-Sea Res (in press)
Hansell DA, Carlson CA (1998) Deep-ocean gradients in the concentration of dissolved organic carbon. Nature 395: 263–266
Hobbie JE, Daley RJ, Jasper S (1977) Use of nuclepore filters for counting bacteria by fluorescence microscopy. Appl Environ Microbiol 33(5):1225–1228
Huang B, Hong H, Wang H (1999) Size fractionated primary productivity and the phytoplankton-bacteria relationship in the Taiwan Strait. Mar Ecol Prog Ser 183:29–38 Hung JJ, Chen CH, Gong GC, Sheu DD, Shiah FK (2001)
Dis-tribution, stoichiometric patterns of dissolved organic mat-ter in the East China Sea and their cross-shelf export. Deep-Sea Res (in press)
Jickells TD (1998) Nutrient biogeochemistry in the coastal zone. Science 281:217–222
Lancelot C, Billen G (1984) Activity of heterotrophic bacteria and its coupling to primary production during the spring phytoplankton bloom in the south bight of the North Sea. Limnol Oceanogr 29(4):721–730
Lebaron P, Servais P, Troussellier M, Courties C, Vives-Rego J, Muyzer G, Benard L, Guindulain T, Schafer H, Stacke-brandt E (1999) Changes in bacterial community structure in seawater mesocosms differing in their nutrient status. Aquat Microb Ecol 19:255–267
Liean CH (1999) Temporal and spatial variability of ammo-nium concentrations and its uptake by different size phytoplankton in the East China Sea. Master thesis, Insti-tute of Marine Biology, Nationall Sun Yat-Sen University Lin CY (2000) Studies on temporal and spatial variations of
the abundance of oligotrich ciliates in the East China Sea. Master thesis, Dept Fishery Science, National Taiwan Ocean University
Lochte K, Bjonrsen PK, Giesenhagen H, Weber A (1997) Bac-terial standing stock and production and their relationship to phytoplankton in the Southern Ocean. Deep-Sea Res II 44(1-2):321–340
Mantoura RFC, Martin JM, Wollast R (1991) Ocean processes in global change. Wiley & Sons, New York
Michaels AF, Bates NR, Buesseler KO, Carlson CA, Knap AH (1994) Carbon-cycle imbalance in the Sargasso Sea. Nature 372:537–539
Nagata T, Kirchman DL (1992) Release of organic matter by heterotrophic protozoa: implication for microbial food-webs. Arch Hydrobiol 35:99–109
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis. Pergamon, New York
Pinhassi J, Hagrstrom K (2000) Seasonal sucession of marine bacterioplankton. Aquat Microb Ecol 21(3):245–256 Pomeroy LR (1974) The ocean’s foodweb, a changing
para-digm. BioScience 24(7):499–504
Proctor LM, Fuhrman JA (1990) Viral mortality of marine bac-teria and cyanobacbac-teria. Nature 343(4):60–62
Proctor LM, Fuhrman JA (1991) Roles of viral infection in organic particle flux. Mar Ecol Prog Ser 69:133–142 Sanders RW, Caron DA, Berninger U (1992) Relationships
between bacteria and heterotrophic nanoplankton in marine and fresh waters: an inter-ecosystem comparison. Mar Ecol Prog Ser 86:1–14
Sherr EB, Sherr BF, Albright LJ (1987) Bacteria: link or sink. Science 235:88–89
Shiah FK (1999) Diel cycles of heterotrophic bacterioplankton abundance and production in the ocean surface waters. Aquat Microb Ecol 17(3):239–246
Shiah FK, Gong GC, Liu KK (1999) Temperature vs substrate limitation of heterotrophic bacterioplankton production across trophic and temperature gradient in the East China Sea. Aquat Microb Ecol 17(3):247–254
Shiah FK, Liu KK, Kao SJ, Gong GC (2000a) The coupling of bacterial production and hydrography in the southern East China Sea. Cont Shelf Res 20(4/5):459–477
Shiah FK, Gong GC, Chen TY, Chen CC (2000b) Temperature dependence of bacterial specific growth rates on the con-tinental shelf of the East China Sea and its potential appli-cation in estimating bacterial production. Aquat Microb Ecol 22(2):155–162
Sokal RR, Rohlf FJ (1969) Biometry: the application and prac-tice of statistics in biological research. Freeman & Com-pany, San Francisco
Thingstad TF, Perez M, Pelegri S, Dolan J, Rassoulzadegan F (1999) Trophic control of bacterial growth in microcosms containing a natural community from northwest Mediter-rean surface waters. Aquat Microb Ecol 18:145–156 Touratier F, Legendre L Vezina A (1999) Model of bacterial
growth influenced by substrate C:N ratio and concentra-tions. Aquat Microb Ecol 19:105–118
Vrede K (1999) Effects of inorganic nutrients and zooplankton on the growth of heterotrophic bacterioplankton — enclo-sure experiments in an oligotrophic clear water lake. Aquat Microb Ecol 18(2):133–134
Wiebe WJ, Sheldon WM, Pomeroy LR (1992) Bacterial growth in the cold: evidence for an enhanced substrate require-ment. Appl Environ Microbiol 58(1):359–364
Williams PJLeB (1981) Incorporation of microheterotrophic processes into the classical paradigm of the planktonic food web. Kiel Meeresforsch 5:1–28
Wong GTF, Gong GC, Liu KK, Pai SJ (1998) ‘Excess Nitrate’ in the East China Sea. Estuar Coast Shelf Sci 46:411–418 Wong GTF, Chao SY, Li YH, Shiah FK (2000) The Kuroshio
Exchange Processes (KEEP) — an introduction to hypothe-ses and highlights. Cont Shelf Res 20(4/5):335–347
Editorial responsibility: Patricia Glibert, Cambridge, Maryland, USA
Submitted: August 1, 2000; Accepted: December 20, 2000 Proofs received from author(s): February 9, 2001