Biochimica e~ Biophysica Acto, 955 (1988) 1-9 Elsevier
BBA 33139
Biochemical compariison of lens crystalHns from t~ee reptilian species
Shyh-Horng Ch/ou, Wan-F~n Chang and Chhh-Hcrng Lo
Institute of Biochemical Sciences, National Taiwan University and Institute of Biological C~emistry, Academia Sinica, P.O. Box 23-195, Taipei (Taiwan, China)
(Received 4 January 1988)
Key words: Crystallin; Circular dichro~sm; Subunit structure; Amino acid composition: Sequence homology: (Reptile lens)
Lens cryst~|lins were isolated from the homogenates of reptilian eye lenses ~]efived from t h e e different species by gel-permeation chromatography and character~ed by gel electrophoresis, amino-acid analysis, N-terminal s~nence analysis and circular dichroism. Four fractions corresponding to a-, 8 / e / p - , p- and v-crysta~ns were obtained for the crys~lins from caiman lenses, whereas B- and V-crys~|in fraction were present in lesser amounts or missing in the turtle and snake lenses, respectively. The native molecu|ar masses for these purified fractions and their po|ypeptide compositions were determ|ned by gel filtration a~ad SDS-ge| electrophoresis, respectively, re~Tea|ing the typical s~bunit compositions for each c|assified crysta|- iin. The spectra of circular dichroism indicate a predominant fl-sbeet structure in a-, p- and v-crys~ilins, and a major contribution of a-he|ical s ~ c t u r e in 8/e-crystalfin fraction, which bears a resemb|ance to the secondary structm~e of &crystal|in from file chicken lenses. Comparison of the amino-acid contents of each orthologous class of repti|ian crystal|ins with those of evolutionary distant species sti|l exhibited simi|arity ~¢L~ their amino-acid compositions. N-terminal sequence analysis of the crysta||in fractions revealed that a|| fractions except that of V-crystal|in are N.terminal|y blocked. Extensive sequence simi|arity between the repfi|ian ¥-crystaliin po|ypepfides and those from other vertebra~te species were found, which establish the close relatedness of V-crystal|ins amongst the major classes of vertebrates.
Introduction
The lens crystallins of vertebrates comprise a complex group of conserved structural prc, teins with distant evolutionary relationships [1-3]. We have recently characterized the crystallins from several different species encompassing one class of invertebrates [4] and all five major classes of vertebrates [5-10] with the aim of searching for their evolutionary relatedness and phylogenetic
Correspondence: S.-H. Chiou, Institute of Biological Chera- istry, Academia Sin|ca, P.O. Box 23-195, Taipei, Taiwan, China.
relationships on the basis of their protein and gene structures. The different classes of crystallins not only vary between species, but are also dif- ferentially expressed during lens development. In addition, the recent reports of sequence similarity between a minor e-crystallin of the duck lens and lactate dehydrogenase [11,12] plus the relatedness between p-crystallin of the amphibian lens and aldose reductase/aldehyde reductase family [13] have added some interesting aspects of crystallin evolution in relation to the general topics of pro- tein evolution. It is imp,:rative to compare crystal- lins of various species from different phylogenetie levels in order to shed ~ome light on the mecha- 016%4838/88/$03.50 © 1988 Elsevier Science Publishers B.V. (Biomedical Division)
nism u~derl)4ng the process of crystallin diversifi- cation and evolution. The molecular approach to this important problem will lie in the extensive characterization of crystallins at both the protein and gene levels from the available species in the animal kingdom. In this report we have furnished some basic information on the physicochemical properties of reptilian crystallins from three differ- ent species, which should form a useful basis for the genomic comparison between reptilian and other vertebrate crystallins in the future.
Materials and Methods
Isolation of lens crystallins
Caiman (Caiman crocodylus apaporiensis), snake
(Trimeresurus mucrosquamatus) and river turtle
(Amyda sinensis) lenses were obtained from local reptile farms. The decapsulated lenses were ho- mogenized in 10-20 ml 0.05 M Tris-sodium bi- sulfite buffer (pH 7.5) containing 5 mM EDTA as described before [14,15]. The supernatant from centrifugation at 27 000 × g was adjusted to give a concentration of about 20-30 mg/ml, and the 5.0 ml aliquot was applied to Fractogel TSK HW-55 (Superfine Grade, Merck). It offered good and well-defined resolution, similar to that found in high-performance liquid chromatography (HPLC). Native molecular masses of the eluted fractions were determined on the same column (2.5 × 115 cm) using the following standard proteins: thyroglobulin (670 kDa), catalase (240 kDa), transferrin (80 kDa), ovalbumin (45 kDa) and soybean trypsin inhibitor (20 kDa).
The dialyzed and lyophilized 7-crystallin frac- tion from the gel-filtration column was further separated into its subfractions on a TSK CM-650 (S) cation-exchange column with a linear gradient of 0.05-0.25 M ammonium acetate (pH 5.9). The separated subfractions and the unfractionated 7- crystallin were taken for amino-acid analysis and circular dichroic studies without detecting sub- stanfial differences.
Gel electrophoresis
Isoelectric focusing in 5~ polyacrylarnide gel containing 2.8~ (v/v) carrier ampholytes of pH 3.5-10 was carried out on a slab gel with the incorporation of 0.1~ 2-mercaptoethanol and 6 M
urea. The gel was fixed in 12.5~ trichloroacetic acid, washed several times with 20~ methanol/7~ acetic acid and stained with Coomassie blue. SDS-polyacrylamide slab gel (5~ stacking/14~ resolving gel) was as described [16] with some modifications.
Amino-acid analysis
The amino-acid compositions were determined with the LKB-4150 amino-acid analyzer using a single-column system. The dialyzed and lyophi- lized protein samples were hydrolyzed at 150°C on a dry heating block in Pierce re-usable vacuum hydrolysis tubes with constant-boiling 6 M HCI (Pierce Chemicals, U.S.A.) for 1.5 h. We have shown the applicability of high temperature (150°C) and shortened time (1.5 h) in achieving comparable amino-acid composition data similar to thc~e obtained by the tedious conventional 110°C/24 h protocol (Ref. 17 and unpublished results).
The half-cystine content was determined sep- arately after performic acid oxidation. Tryptophan was not determined. The procedure of Marchalonis and Weltman [18] was used to analyze and com- pare the relatedness of amino-acid compositions using the equation:
S~Q-- ~:(xij - xkj) 2
where the subscripts i and k identify the particu- lar protein pairs being compared, Xj is the mole content of a given amino acid of type j. The summation is carried out over the 17 types of amino acids typically determined on the 6 M HCI hydrolysates of crystallin samples.
N-terminal sequence analysis
The N-terminal sequences of the four major fractions from the gel-permeation column and the fractionated y-crystallin subfractions from TSK CM-650 (S) cation-exchange chromatography were carried out by automated Edman degradation with a microsequencing sequenator (Model 477A, Ap- plied Biosystems). The lyophilized crystallin sam- ples each containing about 1-5 nmol of protein were dissolved in 200 ill of 0.1% trifluoroacetic acid or 0.1~ SDS/0.1~; trifluoroacetic acid (1 : 1, v/v) and 10/tl each for sequence determinations.
Circular dichroism
The circular dichroic spectra for each crystallin were obtained on a Jasco J-41C spectropolarime- ter under constam nitrogen flush and the sample was placed in a cell thermostated at 20 o C. The instrument was calibrated with an aqueous solu- tion of (+)-10-camphorsulfonic acid. The crystal- lins were dissolved in 0.05 M "Iris buffer (pH 7.8) at a concentration of 0.5-1.2 mg/ml, depending on the samples. All protein solutions were filtered through an Acrodise (Gelman) membrane (0.2 #m) before spectra~ were taken. The ellipticity data were converted to mean-residue-weight ellipticity using a mean residue weight of 115 for all crystal- fins. Analysis of CD in terms of the fractions of the structural elements, i.e., helix,/3.,sheet,/~-turn and unordered l~orm, was carried out according to the procedure of Chang et al. [19]. A nonlinear least-~quares curve f~tting in the 205-240 nm re- gion at 2 nm intervals of CD-spectra was used to find the best estimate for the percent contribution of each structural element in the studied crystal- [ins.
Protein concentration determinations
The protein concentrations of crystallin solu- tions were determined by amino-acid analysis. The absorption coefficient (1 mg/ml, at 280 nm) was then determined for each purified crystallin. The protein contents of crystallin samples for the CD study were estimated by their absorbances at 280 nm.
Results and Discussion
There have been some reports on the compara- tive studies of a-, /~- [20-22] and 8-crystallins [23-24] of the submammalian species. However, the systematic characterization and comparison on the crystallin fractions from different species be- longing to the reptilian class are still lacking. We have previously detected similar amino-acid com- positions for the crystailins of differeut vertebrate species, which suggested the existence of the se- quence similarity in their primary structures [3,5-9]. A more extensive chzracterization of the crystallins from the species of the the same class of vertebrates may provide some insight into the phenomenon of sp.~des diversification and the
<
A;
40 6.~ : 0 I I t ~ I I I I l I 10 ~ 12~ I~C Fraction NumberFig. 1. Gel-permeation chromatography of reptilian c.~3,~talhns on Fractogel TSK HW-55(S) (2.5 x 115 cm colal,~m). The ~,ol° umn eluates (3.5 ml/tube per 4.1 min) were monitored for absorbance at 280 nm. The peak fractions were collected and used for amino-acid analyses (Table I) and SDS-polyacryl- amide gel electrophoresis in Fig. 2. The absorbances at 280 nm are relative concentrations in arbitrary units. The small broad peak at the end of elution pattern ef river turtle lens is nonprotein components of low molecular masses. The arrow indicates the elution position of 8-crystallin (Sk2) for the snake
lens.
accompanying molecular origin of crystallin evolution within the vertebrate.
Characterization of crystallins of three reptilian species from the gel-permeation chromatography
Fig. 1 shows elution patterns of lens extracts from several reptile species. Four peaks were ob- tained for all three species in contrast to three for the fish and five for the mammal [3,8]. The identi- fication of each crystallin fraction from the coi-
umn was based on the subunit analysis by SDS-gel electrophoresis of Fig. 2. The clean and character° istic pattern of doublet band for a-crystallin is clearly shown in the first peaks of all three species. In contrast to the case of frog crystalfins [9], the reptilian a-crystallin fraction is well separated from r - and other classes of crystallins. The native molecular mass was estimated to be about 600 kDa, which is slightly lower than that of bovine a-crystallin [14].
Re-chromatography of the peaks of gel permea- tion on the same column to remove some cross- contaminating fractions is sufficient to obtain over 90~ pure proteins as indicated in the SDS gels, except that of peak 2 which consistently showed cross-contamination of ~- and e-crystallins in the major fraction of 8-crystallin as revealed by SDS- gel electrophoresis.
It is of interest to note that 8-crystallin seemed to be absent from the fiver turtle lens [10] when comparing the subunit compositions of the three reptilian species (lanes 2 of Fig. 2A-C). The possi- bility that it aggregated as a high-molecular species as indicated by the band in the top region of the gel with molecular mass greater than 120 kDa (see peak 2 of Fig. 2C) can be ruled out since the electrophoresis was run in the presence of 5 mM dithiothreitol and 0.1% SDS. However, Williams et all. [25] reported the prominent presence of &crystallin in the lenses of the red-eared turtles
(Pseudemys scripta),
with a subunit molecular weight of about 48000. The reason for the dis- crepancy as to 8-crystallin in the two species ot turtles is not clear. However, the fiver turtle used in this study belongs to the order of Amphiche- lydia and the sea turtle that of Cryptodira (both were evolved from the order of Anapsida). The absence of 8-crystallin in the fiver turtle may just reflect the diversification between the two species of turtles. This should constitute an interesting topic for the future phylogenetic comparison of lens crystallins in these two groups of modem turtles from the viewpoint of protein evolution and species diversification.Another interesting aspect in the comparison of the three species of reptiles lies in the apparent lack of ~-crystallin in the snake as judged from the subunit composition of snake crystallins (Fig. 213). The amino-acid compositions of these crystal-
A
]3
C
8 4
~ wS
W W W1
a 6
8 8 4 S
$ 1 2 8 4
• , W w. a
p v
Fig. 2. Gel electrophoresis of the fractionated reptilian crystal- fins under denaturing conditions (SDS-polyacrylamide gel elec- trophoresis) in the presence of 5 mM dithiothreitol. Lanes S, standard proteins used as molecular mass markers (in kDa): phosphorylase b (94), bovine serum albumin (66), ovalbumin (45), carbonic anhydrase (30), soybean trypsin inhibitor (20.1) and iysozyme (14.5). Lanes 1-4 correspond to the four peak fractions of of Fig. 1. The gels were stained with Coomassie blue in (A) caiman and (C) river turtle and silver staining for (B) snake crystallins. The unambiguous assignment of fractionated crystallins to each class of crystallins has been indicated below each lane. Arrow and double-head arrow in (A) represent the electrophoretic positions of 8-crystallin and
TABLE I
COMPARISON OF AMINO-ACID COMPOSITIONS OF CRYSTALLINS
Amino acid a Cml Skl Rtl Sk2 Crn,3 Rt3 Sk4 Cm,~ Rt4 H ~ f C y s 0.5 0.5 0.6 0 0.6 0.7 1.4 3.2 2.6 Asx 9.7 10.0 9.4 12.0 9.5 9.9 8.1 9.2, 9.9 Thr 4.1 4.2 4.0 7.5 3.2 3.3 2.9 3.0 2.3 Ser 12.2 12.0 10.7 8.1 8.6 7.7 !0.6 5.9 6.8 G ~ 9.6 10.5 9.9 10.7 15.8 15.3 14.6 14.0 12.7 Pro 7.5 7.6 8.7 3.3 6.6 5.8 7.5 3.6 5.2 Gly 5.3 5.2 5.6 6.5 10.1 10.8 9 4 7.8 7.6 Ala 3.2 3.1 4.1 7.0 3.3 4.5 5.5 2.5 1.9 Val 5.4 5.5 4.7 6.1 6.2 7.4 5.7 3.5 4.5 Met 1.9 1.3 1.2 1.4 1.6 0.8 1.8 2.9 3.0 Ile 6.1 4.7 5.2 6.2 5.8 4.3 4.2 4.7 4.4 Leu 9.6 8.8 8.6 11.7 5.5 5.8 5.8 10.6 9.0 Tyr 3.6 2.6 2.1 3.6 4.7 3.5 3.8 9.3 8.1 Phe 6.0 7.8 7.5 3.5 5.2 5.2 4.6 5.0 5.7 His 4.6 4.7 5.0 1.7 2.7 2.2 2.5 " 2.1 3.2 Lys 3.9 3.7 5.2 5.8 5.3 7.5 5.2 2.3 2.1 Arg 6.8 8.0 7.8 5.1 5.7 5.8 6.0 10.5 10.8 T ~ n.d. b n.d. n.d. n.d. n.d. n.d. n.d. n.d. n.d. SAQ c 10 8 115 133 15 28 142 14
a Data of amino-acid analyses are expressed as mol~, and the abbreviations Cm, Sk and Rt are for caiman, snaky and river turtle, respectively, with the numbers indicating the peak fractions of gel filtration chromatography in Fig. 1.
b n.d., not determined.
c SAQ represents the pail'wise comparison of amino-acid contents of adjacent crystallins as described in 'Materials and Methods'.
TABLE II
PHYSICOCHEMICAL PARAMETERS OF REPTILIAN CRYSTALLINS
Caiman (Cm) Snake (Sk) River turtle (Rt) a , B , 8, ~, V a, & B a , B , V, 8(7) 1. classes of crystallins 2. Native molecular mass (kDa) 3. Subunit molecular mass (kDa) 4. Major secondary structure (a-helix/B-sheet) Cml (6OO) Skl (580) Rtl (~10) Cm2 (190) Sk2 (190) Rt2 (160) Cm3 (120) Sk3 (140) Rt3 (120) Cm4 (23) Sk4 (60) Rt4 (25) Cml (20, 22) Skl (21, 22) Rtl (20, 22) Cm2 (50, :~7, 26) Sk2 (50) Rt2 (150, 32, 26, 23) Cm3 (26, 24) Sk3 (50, 26, 23) Rt3 (26, 23, 22) Cm4 (19-2.;) Sk4 (23-26) Rt4 (20-23) C m l (fl) Skl (fl) Rtl (/~) Cm2 (a) Sk2 (a) Rt2 (fl) Cm3 (fl) Sk3 ( f l / a ) Rt3 (fl) Cm4 (B) Sk4 (B) Rt4 (B)
5. % Secondary structure for caiman crystallins
Cml Cm2 Cm3 Cm4
a-helix 9 + 5 5 2 + 6 15+ 4 12± 3
B-sheet 35 + 8 17 + 5 41 + 6 38 ± 6
B-turn 23 4-10 20 + 6 24 + 10 21 + 12
Unordered form 33 + 6 114- 8 20 + 8 29 :t: 5
The methods used for determination of these parameters are described in 'Materials and Methods'. The values shown for secondary structure represent the estimates from nonlinear least-squares curve fitting of CD spectra + (estimated uncertainty in the fitting procedure).
lins (Table 1) also corroborate that none of the fractions of the snake crystallins corresponded to the composition of y-crystallin, and the peak 4 fraction of snake lens (Sk4) indeed belonged to /~-crystallin family. Since the crystallins isolated from the caiman lens [15] comprise both 8- and ~/-crystallins which are missing in the river turtle and snake lenses, respectively, the detailed bio- chemical comparison has been carried out based on the characterization of caiman crystallins.
W~ysicochemical parameters of reptilian crystallins
Table II summarizes the physicochemical prop- e~'~ies of reptilian crystallins characterized in this study. The native molecular weight and the sub- umt structures of the orthologous classes of or-,/]- ancl y-crystallins are in fair agreement with other vertebrate species characterized before [26]. De- spite the apparent homogeneity of each fraction as
I 2 3 4
Fig. 3. l.umlecmc focusing of caiman crystallins under denatur- ing conditions. Lanes 1-4 correspond to Cml-Cm4 of Fig. 1. The slab gel contained 6 M urea and 0.1~ 2-mercaptoethanol and the electrophoreMs was run at an initial voltage of 200 V for 6 h until a f'mal current of less than 0.5 mA was reached. The approximate ~soelectric points of various ¢rystallin poly- peptides were estimated from a p I calibration kit (range 4.7-10.6, BDH Chemicals). The p l values of 4.8-5.9, 5.0-7.1, 6.2-7.1 and 6 . 8 - 8 . 6 were obtained for Cml to Cm4,
respectively. 0 -2 -4 ~ " 6 " ._E - 8 - 0 -o -10 - E -12 " o >¢ (D -4 - 6 - 8 - - I I I I I A
~
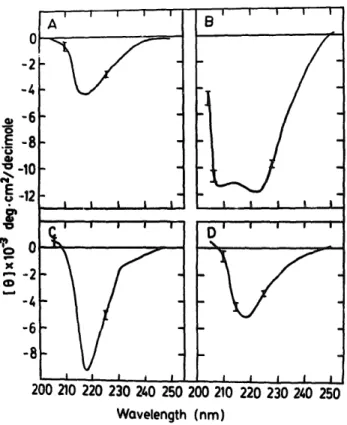
1 I I I I i B i , l 5 i t D I I I I I . B i I l i I i .I I I I 200 210 Z20 230 240 250 200 210 220 230 240 250 Wovelength (nrn)Fig. 4. CD spectra of caiman crystalfins in the far-ultraviolet region. Conditions are as described in 'Materials and Methods'. [ 01 is the mean residue ellipticity in degrees, cm 2 per decimole based on a mean residue weight of 115 for all crystallins. (A) Cml (B) Cm2 (C) Cm3 and (D) Cm4 correspond to the four re-chromatographed peaks of Fig. 1. The error bars represent
the noise levels of the instrument.
shown by the well-defined subunit compositions similar to those reported for mammalian crystal- lins [26,27], the charge heterogeneity can be de- tected by isoelectric focusing under denaturing conditions as indicated in Fig. 3. Caiman y (Cm4, lane 4 of Fig. 3) shows more than four charge-iso- meric forms (pI 6.8-8.6) similar to carp and bovine ~,-crystallin [7,8,29]. a-Crystallin (lane 1 of Fig. 3) shows four distinct bands with low p I correspond- ing to well-characterized a-A and a-B chains [27]. Peak 2 (Cm2, lane 2 of Fig. 3) exhibited the highest heterogeneity among all crystallin frac- tions. Williams and Piatigorsky [30] have shown five major and nine minor O-crystallin bands from the mallard lens by isoelectric focusing. Ap- parendy, the subunits of O-crystallin possess an extensive charge heterogeneity. We have failed to purify caiman 8-crystallin by repeated gel filtra- tion and various ion-exchange chromatographies. The coeluted and contaminated e- and/]-crystal-
t 5 1o o_=_ G-K- IHT-F-Y-E-D tKtlt-F-0-G.
o-=- =i,-,-,-=%=-oi,-=-o
G-K- I15
R-S4 R-S~ ,L-N4 ,L-S~ .R-C4 .R-S] .R-Cq .H-CA IR-Sq 20 D - " b - M-S- M-H- M-S- .S-G- " S ~ :S-S- IIS-S-
-T-T-(C.ar~0 7I )
(Carp yI
I)
(Carp y I I I )(Carp yIv)
( F r o g yI I I ) ( C ~ a n yI I ) (Raty2-1
)
(Ca].~ 711 )
< HL.r, an ~2- I )Fig. 5. Comparison of N-termina~ sequences of ~,-crystallins from various species. The sequences listed for caiman, carp and frog crystallins were taken from this study and those for other zpecies from Ref. 15 and the references therein. The homology ~egion is
bo×ed. Amino-acid residues are denoted by one-letter symbols.
lins were always found in varied amounts with the • major &crystallin in the SDS gels. therefore, the amino-acid analysis was carried out on a purer 0-crystallin from the snake lens despite a much lower content of this crystallin in the snake species (Fig. 1 and Table I).
Circular dichroic study
CD spectra in the far-ultraviolet region for the caiman crystallin fractions are shown in Fig. 4. All crystallin fractions except Cm2 corresponding to /~-crystallin showed the predominance of fl-sheet structure. The estimations of the fractions of four basic structural elements (Table II) indicated about 35-45% of fl-sheet contribution t~r a-, fl- and ~,-crystallins, whereas about 52 +6% a-helical structure was found in Cm2. This is consistent with the previous conformation study of chicken
8- and duck e-crystallins [31,32], further corrobo- rating the existence of similar conformation of the peptide backbone in the orthologous crystallins of the vertebrate lens. The CD spectra of the frac- tions of the turtle and snake crystallins were also compared with the corresponding fractions of cai- man crystallins (data n o~ shown); essentially no qualitative differences could be found in the peptide region, except that there were small quantitative differences in the magnitude of el- liptieities. It is noteworthy that a-, 13- and ~,-crys- tallins demonstrated essentially similar secondary
structures even from the evolutionarily distant species despite their differences in primary se- quences and immunological properties (un- published results). This would indicate some con- formational relatedness among the major crystal- lin classes characterized here.
N-terminal sequence analysis
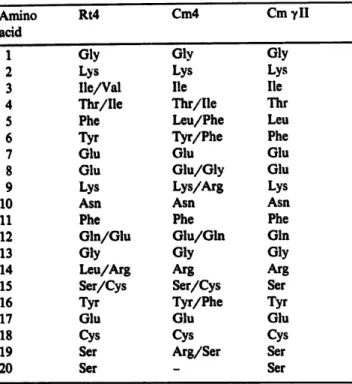
A more defined sequence comparison of these crystallins is needed for establishing the phylo- genetic relationships amongst different classes of crystallins. It is surprising to find that all crystal- lins except ~,-crystallins contain blocked N-termi- nal residues (Table III). We have also found a blocked N-terminus for the peak 4 fraction of the snake, strengthening our conclusion that the snake lens lacks ~,-crystallin fraction which is present in turtle and caiman species of reptiles. Hence, the preliminary sequence analyses were carried out on caiman and turtle 7-crystallin fractions. N-termi- nal sequence analyses of the crude (turtle) and purified ~,II-crystallin subfractions (caiman) from cation-exchange column indicated that they were closely related to each other with almost identical N-terminal sequences (Ref. 15 a:ad Table lII). Some differences and heterogenei'~y were detected along the sequences by microsequencing. There are several positions at which two amino acids have been identified, which is indicative of more than four ~,-crystallin polypeptides present in the
TABLE ill
THE AMINO-TERMINAL SEQUENCES OF REPTILIAN CRYSTALLINS
The residues with more than one amino acid denote those positions where more than one phenylthiohydantoin derivative was detected by automatic sequencing. Dashes indicate the unidentified amino acids. Cml-3, Skl-4 and Rtl-3 of Table II were found to be N-terminally blocked.
Amino Rt4 Cm4 Cm ~II
acid
1 Gly Gly Gly
2 Lys Lys Lys
3 lle/Val lie lie
4 Thr/lle Thr/lle Thr
5 Phe Leu/Phe Leu
6 Tyr Tyr/Phe Phe
7 Glu Glu Glu
8 Glu Glu/Gly Glu
9 Lys Lys/Arg Lys
10 Asn Asn Asn
11 Phe Phe Phe
12 Gln/Glu Glu/Gln Gin
13 Gly Gly Gly
14 Leu/Arg Arg Arg
15 Ser/Cys Ser/Cys Ser
16 Tyr Tyr/Phe Tyr
17 Glu Glu Glu
18 Cys Cys Cys
19 Ser Arg/Ser Ser
20 Ser - Ser
reptilian ~,-crystallin family. Comparison of the amino-terminal sequences of ~,-crystallin poly- peptides from the six different species encompass- ing four major classes of vertebrates, and wide evolutionary history showed a high degree of se- quence similarity (Fig. 5). 7-Crystallin was found in either decreased amounts or was missing in the lenses of the bird class [5,26] and therefore not included for comparison. The percentage of se- quence identity for the first 20 residues of all species is about 70~. The protein sequence study associated with the present report has indicated a great deal of difficulty encountered in the purifi- cation of the trypfic and cyanogen bromide frag- ments of ¥-crystallin polypeptides due to their inherent microheterogeneity, which makes the se- quence determination of crystallin polypeptides more feasible at the gene than at the protein level. The partial sequences reported here could also provide the crucial information in making several
oligonucleotide probes for the future molecular cloning of rept~an crystallin genes.
Conclusions
A systematic and general approach has been carried out to provide some basic information on the protein chemistry of reptilian crystallins. Some salient differences between different species be- longing to the same class have been found, which may be important in the comparison and tracing of the evolutionary origin amongst these related species.
Acknowledgement
We thank the generous support of the National Science Council, Taipei, Taiwan, R.O.C.
References
1 De Jong, W.W. (1981) in Molecular and Cellular Biology of the Eye Lens (Bloemendai, H., ed.), pp. 221-278, Jolm Wiley & Sons, New York.
2 Bloemendal, H. (1985) Exp. Eye Res. 41, 429-448. 3 Chiou, S.-H. (1986) FEBS Lett. 201, 69-73. 4 Chiou, S.-H. (1984) J. Biochem. (Tokyo) 95, 75-82. 5 Chiou, S.-H., Chang, W.-C., Kuo, J., Pan, F.-M. and Lo,
T.-B. (1986) FEBS Lett. 196, 219-222.
6 Chiou, S.-H., Chang, T., Chang, W.-C., Kuo, J. and Lo, T.-B. (1986) Biochim. Biophys. Acta 871, 324-328. 7 Chiou, S.-H., Chert, S.-W. and Lo, T.-B. (1986) FEBS Lett.
209, 107-110.
8 Chiou, S.-H., Chang, W.-C., Pan, F.-M., Chang, T. and Lo, T.-B. (1987) J. Biochem. (Tokyo) 101, 751-759.
9 Chiou, S.-H. (1987) Intern. J. Peptide Protein Res. 30, 108-116.
10 Chiou, S.-H. (1987) Lens Res. 4, 163-171.
11 Wistow, G.J., Mulders, J.W.M. and De Jong, W.W. (1987) Nature 326, 622-624.
12 Wistow, G.J. and Piatigorsky, J. (1987) Science 236, 1554-1556.
13 Carper, D., Nishimura, C., Shinohara, T., Dietzchold, B., Wistow, G., Craft, C., Kador, P. and Kinoshita, J.H. (1987) FEBS Lett. 220, 209-213.
14 Chiou, S.-H., Azari, P., Himmel, M.E. and Squire, P.G. (1979) Intern. J. Pepfide Protein Res. 13, 409-417. 15 Chiou, S.-H., Chang, W.-P., Lo, C.-H. and Chen, S.-W.
(1987) FEBS Lett. 221, 134-138.
16 Laemmli, U.K. (1970) Nature 227, 680-685.
17 Chen, S.-T., Chiou, S.-H., Chu, Y.-H. and Wang, K.-T. (1987) Intern. J. Peptide Protein Res. 30, 572-576. 18 Marchalonis, J.J. and Weltman, J.K. (1971) Comp. Bio-
19 Chang, C.T., Wu, C.-S.C. and Yang, J.T. (1978) Anal. Biochem. 91, 13-31.
20 De Jong, W.W., Terwindt, E.C. and Groenewoud, G. (1976) Comp. Biochem. Physiol. 55B, 49-56.
21 Zigler, J.S., Jr. and Sidbut$,, T B, Jr. (1976) Comp. Bio- chem. Physiol. 55B, 19-24.
22 Zigler, J.S., Jr. and Sidbury, J.B., Jr. (1977) Ophthalmic Res. 8, 92-98.
23 Williams, L.A. and Piatigorsky, J. (1979) Eur. J. Biochem. 100, 349-357.
24 De Jong, W.W., Stapel, S.O. and Zweers, A. (1981) Comp. Biochem. Physiol. 69B, 5,93-598.
25 Williams, L.A., Piatigors~ky, J. and Horwitz, J. (1982) Bio- chim. Biophys. Acta 708, 49-56.
26 Bindels, J.G., Bessems, G.J.J., De Man, B.M. and Hoenders, H.J. (1983) Comp. Biochem. PhysioL 76B, 47-55.
27 Bloemendal, H. (1977) Science 197, 127-138.
28 Harding, J.J. and Crabbe, M.J.C. (1984) in The Eye (Day- son, H., ed.), Vol. 1B, pp. 207-492, Academic Press, London
29 BjSrk, I. (1964) Exp. Eye Res. ~, 254-2C,1.
30 Williams, L.A. and Piatigorsky, J. (1979) Biochemistry 18, 1438-1442.
31 Honvitz, J. and Piatigorsky, J. (1980) Biochim. Biophys. Acta 624, 21-29.
32 Stapel, S.O., Zweers, A., Dodemont, H.J., Kan, J.H. and De Jong, W.W. (1985) Eur. J. Biochem. 147, 129-136.