國立交通大學
生物科技學系
博士論文
幽門螺旋桿菌熱緊迫蛋白 60 活化β1 轉化生長因子
訊息傳遞之機制
Helicobacter pylori
Heat Shock Protein 60 Trigger SMAD Signal
Pathway by Interacting with Transforming Growth Factor-β
receptor II
學 生: 林 靜 宜
幽門螺旋桿菌熱緊迫蛋白 60 活化β1 轉化生長因子訊息傳遞之機制
Helicobacter pylori Heat Shock Protein 60 Trigger SMAD Signal Pathway by Interacting
with Transforming Growth Factor-β receptor II
研 究 生:林靜宜 Student:Ching-Yi Lin
指導教授:廖光文 Advisor:Kuang-Wen Liao
國 立 交 通 大 學 生 物 科 技 學 系 博 士 論 文 A thesisSubmitted to Department of Biological Science and Technology College of Biological Science and Technology
National Chiao Tung University in partial Fulfillment of the Requirements
for the Degree of PhD in
Biological Science and Technology
December 2008
幽門螺旋桿菌熱緊迫蛋白 60 活化β1 轉化生長因子訊息傳遞之機制
學生:林靜宜 指導教授:廖光文 博士
國立交通大學 生物科技學系 博士班
摘 要
幽門螺旋桿菌為普遍存在人體的致病菌。根據統計,全球約有一半左右的人口感染此 菌。該菌會在胃部造成持續性的慢性感染,進而增加十二指腸潰瘍、胃癌或淋巴腫瘤之 罹患率。在病原慢性感染的機制中,抑制宿主的免疫反應扮演了極為重要的角色;而在 幽門螺旋桿菌的免疫抑制效應方面,目前雖然有若干的研究進行探討,然而其機制仍尚 未完全被釐清。幽門螺旋桿菌熱緊迫蛋白 60 為該菌的黏附因子之一;而由於其能引發 許多炎症細胞激素的產生,故該蛋白也常作為幽門螺旋桿菌疫苗研發的主要抗原。然 而,近來相關的研究顯示,熱緊迫蛋白 60 具有調控免疫反應的功能。本研究的目的在 探討幽門螺旋桿菌熱緊迫蛋白 60 的結構及其在免疫反應中所扮演的角色。首先,由氨 基酸序列比對、圓二色光譜、以及蛋白質膠體電泳的結果顯示:該蛋白質可形成雙體或 是四聚體,而這與目前已知的大腸桿菌熱緊迫蛋白 60 的結構極為不同。而在免疫功能 的探討方面,我們發現幽門螺旋桿菌熱緊迫蛋白 60 和人類β1 轉化生長因子的蛋白質結 構有部分的相似性,其中有一段相似區已知為β1 轉化生長因子與其第二型受器之結合 位。後續的結果顯示:在 ELISA 與細胞的實驗中均可證實熱緊迫蛋白 60 可與β1 轉化生 長因子受器結合。此種結合會啟動β1 轉化生長因子所調控的 SMAD 訊息傳遞路徑,開啟 下游基因表現,並抑制周邊血球細胞與單核球細胞株 THP-1 的免疫活性。綜合以上的實 驗數據,我們認為幽門螺旋桿菌熱緊迫蛋白 60 會藉由模仿β1 轉化生長因子的生物活性 來抑制宿主的免疫系統,以利於該菌的持續性感染。Helicobacter pylori Heat Shock Protein 60 Trigger SMAD Signal Pathway by
Interacting with Transforming Growth Factor-β receptor II
Student: Ching-Yi Lin Advisor: Dr. Kuang-Wen Liao
Department of Biological Science and Technology National Chiao Tung University
Abstract
Helicobacter pylori have explored multiple mechanisms to evade host immune surveillance
for chronic infection. But either of them is restricted by certain bacterial strains containing potential virulent factors, or those immune-restrained functions only limit to some specific immunocytes. However, the long-term persistence of H. pylori suggests a more comprehensive and powerful factor(s) hinds behind to regulate host immune system.
Helicobacter pylori heat shock protein 60 (HpHsp60) was previous identified as an adhesion
molecule or a potent immunogen. This study aims to study the structure of HpHsp60s and evaluates their functions on host immune responses. Analyzing the structure of HpHsp60 via amino acid blast, circular dichroism and electrophoresis indicated most recombinant HpHsp60s form dimers or tetramers that are quite different than E. coli Hsp60 protein structure. Moreover, a novel property of HpHsp60 was found, which is, by mimicking TGF-β1, HpHsp60 could exert immune regulatory effects. With structural homology to the receptor binding site of TGF-β1, HpHsp60 could interact with TGF-β receptor II, trigger SMAD pathway, and inhibiting the immune functions of THP-1 monocytic cells and peripheral mononuclear cells (PBMCs). Our study provides a new hint that H. pylori may employ Hsp60 to surrender host immunity.
致 謝
大家都說,好不容易畢業了,終於海闊天空了呀!但我卻覺得背上的重擔開 始真實了起來。不能再拿「我是學生」這個藉口胡作非為了呀!心裡默默的想著。 一切,很真實,也很不真實。「一起為了光明的未來而努力」這樣的話果然我是 說不出口的,想說的是,因為未來會遇到的一切都是未知,所以,認真的戰鬥吧, 「不要輸給灰暗時代而來呼吸吧」。 感謝一路陪著我走到這裡的人們,好同學亦涵以及國欽的鼓勵;實驗室的學 弟妹于鈴、彥谷、上知、弘育、詩涵、竣元、懷堯、韻茹、侑松、源庭、其翰、 依穎、昱丞、珮君、竺婷、莉沂、立筠、家弘、維瞳、筱薇、冠倫、堅甫、子慧 於生活及實驗上的協助與支持;萬能助理鈺珊對我在研究與心情上的多所照顧; 與你們相處的情誼會成為我此生美好記憶的一部份。 感謝 廖光文 老師對學生博士論文的指導,拜師於門下,不僅學習到治學之 道,對於待人接物的道理更是獲益良多。而老師對於學生的愛護與殷殷期盼常令 學生感到汗顏,希望在未來能繼續努力不讓老師失望。 最後,我要感謝我的父母與男朋友劉正吉,你們的支持與陪伴是我不停成長 的動力,未來的路將因為有你們的存在而寬廣。目 錄
中 文 摘 要 ………...…………..I
Abstract ………...…II
目錄 ………...….…III
Overview ………1
The immunodominant activity of microbial Hsp60 ………3
Immune responses to mammalian HSPs ……….5
Immunoregulatory roles of Hsp60 ………..6
Potential mechanisms for microbial or endogenous Hsp60s of immune-regulation ..9
Immune responses toward other HSPs ………..12
TGF-β plays an important role in immune-regulatory responses………..13
The interaction between TGF-β and Hsp60-mediated immunregulation …………..15
Characterizing the polymeric status of Helicobacter pylori heat shock protein 60 Abstract ……….28
Introduction ………...29
Materials and Methods ………..31
Results ………...36
Discussion ……….39
Figures and Legends ……….43
Acknowledgements ………...52
References ...……….…….53
with Transforming growth factor-β receptor II
Abstract ……….58
Introduction ………...59
Materials and Methods ………..63
Results ………...74
Discussion ……….80
Figures and Legends ……….85
References ………...108
Overview
Heat shock proteins (HSP) were initially found through a joyful accident involving the overheating of a Drosophia salivary gland preparation on a microscope stage. This was first reported as: “A new puffing pattern induced by temperature shock and DNP in Drosophila” in 1962 (F., 1962). HSPs carry out pivotal housekeeping functions and are molecular chaperones that are important for the survival pf prokaryotic and eukaryotic cells. On the basis of their molecular size they have been divided into several families: Hsp100, Hsp90, Hsp70, Hsp60, Hsp40, and small Hsps (Jolly and Morimoto, 2000; Lindquist and Craig, 1988). Heat shock proteins are strongly conserved and show high homology among all organisms, and some mammalian family members have highly conserved microbial homologous, which results in immunological cross-recognition between mammalian and microbial homologous. HSPs play roles in various aspects. Although they were investigated as molecular chaperones at first, recent studies have revealed their involving in regulation of cell metabolism or decision of cell fate. It has been found that glucocorticoid receptor fails to mature to a transcriptionally active form in the absence of Hsp90 (Picard, et al., 1990). Hsp27 and Hsp70 are also found they can both inhibit caspase dependent apoptosis pathways. Hsp70 inhibits a caspase-independent form of death by interacting with lysosomal membrane and preventing the activity of hydrolytic lysosomal enzymes (Daugaard, et al., 2007).
The first evidence of a role for HSPs as antigens in inflammatory responses was obtained in the 1980s in the rat model of heat-killed mycobacterial-induced adjuvant arthritis (van Eden, et al., 1988). It was found that T cells isolated from rats with disease were responding to mycobacterial Hsp60. Since then, immune reactivity to HSPs has been recognized in human or other experimental models of inflammatory diseases, Recently, several studies have shown that HSP-reactive T cells have an immunoregulatory phenotypes, indicating that HSPs --- and in particular, Hsp60 and Hsp70 --- constitute a group of autoantigens with the potential to trigger immunoregulatory pathways, which can suppress immune responses that occur in various inflammatory diseases. These results display one thing for certainty that HSPs have double-edged-sword effects on host immune systems, and both microbial and mammalian HSPs may possess these immune-regulation abilities. The effects of Hsp60 and other HSPs on host immunity will be reviewed in the followings.
1. The immunodominant activity of microbial Hsp60
Host immune responses to Hsp60 from bacteria, protozoa, fungi and helminthes have frequently been detected after infection with these organisms (Zugel and Kaufmann, 1999). During bacterial infections, the bacterial members of the Hsp60 and Hsp70 families are also common targets of humoral and cell-mediated immune responses in mammals (Kaufmann, 1990; Lydyard and van Eden, 1990). In fact, Hsp60 has been known as the common antigen of gram-negative bacteria before it was identified as a member of the Hsp60 family (Shinnick, et al., 1988; Thole, et al., 1988). Analysis of T-cell responses in mice immunized with Mycobacterium tuberculosis showed that ~10-20% of the M. tuberculosis-specific T cells responded to M. tuberculosis Hsp60 (Kaufmann, et al., 1987). Antibodies specific for Hsp60 of Chlamydia trachomatis have also been detected at high titers in the sera of patients infected with C. trachomatis. With prompting a high-titer antibody response, C. trachomatis Hsp60 has been implicated in devoting to the pathogenesis of C. trachomatis-mediated inflammation and conjunctival scarring (Dean, et al., 2008). Bulut and colleagues has identified the C. trachomatis Hsp60 can activate macrophages and endothelial cells through toll-like receptor 4 and MD2 to turn on NF-κB activation and promote inflammation (Bulut, et al., 2002). Antibodies and T cells specific for mycobacterial Hsp60 have also been detected in healthy humans not known to have been exposed to mycobacterial infections (Munk, et al., 1989). It revels
that Hsp60 can be an immunodominant antigen to evoke host immunity.
The occurrence of immunodominant immune responses to Hsp60 is likely to result from several factors that are related to their function. First, because they are essential in cell survival, HSP sequences are highly conserved between different bacterial species. So, each time the immune system encounters bacteria, its initial response might be targeted to the strongly conserved regions of the bacterial HSP that are recognized by memory cells arising from previous infections with unrelated bacteria. Second, during infection, the invading microorganism experiences stress (for example, a rise in temperature and exposure to reactive-oxygen species produced in host defense), and it upregulates expression of its own HSPs, making them easily available targets for the host immune cells. Third, HSP-specific responses might be immunodominant because, as molecular chaperones, HSPs associate with other, often unfolded, newly synthesized antigenic microbial proteins or with microbial proteins that are denatured or partially unfolded as a result of stress. As a consequence, B cells with cell-surface B-cell receptors specific for a (partially unfolded) microbial antigen can internalize the associated microbial HSP together with the microbial antigen, process them both, and present peptides derived from the two proteins in the context of MHC class II molecules for recognition by T helper (Th)
2. Immune responses to mammalian HSPs
In spite of the existence of immune tolerance to self antigens, host-HSP-specific T and B cells have been found in association with chronic inflammation, as well as in healthy individuals (Cohen, 1991). Because HSPs are intracellular proteins, in the presence of cellular stress, self-HSPs are efficiently presented by major histocompatibility complex (MHC) class I molecules to cytotoxic T lymphocytes (CTLs). An early study showed that CTLs raised against mycobacterial Hsp60 also recognized macrophages that were subjected to various forms of cellular stress, in the absence of exogenously added antigen, indicating that endogenous self-Hsp60 is presented in the context of MHC class I molecules to the CTLs (Koga, et al., 1989). The CTL-mediated lysis of macrophages that were prompted by treatment with IFN-γ was inhibited when Hsp60 expression was suppressed Hsp60-mediated antisense oligonucleotides (Zugel, et al., 1995), thereby confirming the specificity of the CTLs for Hsp60.
HSP-derived peptides can also be presented by MHC class II molecule. In vitro evidence has revealed that stressed antigen-presenting cells (APCs) were recognized by CD4+ T cells that were generated against a highly conserved mycobacterial Hsp60 sequence (Anderton, et al., 1995). In addition, after transferring these T cells to model animals, it was shown to protect against adjuvant-induced arthritis and to produce IFN-γ, interleukin-4 (IL-4) and IL-10. Therefore, recognition of HSPs expressed by stressed cells
might induce a disease-suppressive regulatory phenotype in responding T cells. These findings indicate the existence of CD4+ T cells with the capacity to recognize self-HSP epitopes and may manipulate immune responses associated with disease progression under certain conditions.
3. Immunoregulatory roles of Hsp60
The studies on previous section have implicated the potential role for Hsp60 as an immunoregulator. Because the T cells which can specifically recognize mycobacterial Hsp60 release IL-4 and IL-10 in animal model, which are so-called immune-suppressive cytokines, to protect hosts against inflammatory damages. In fact, the first findings that immune responses to HSPs have a regulatory role in inflammatory diseases were shown for mycobacteria-induced adjuvant arthritis (van Eden, et al., 1988). The animals pre-immunized with mycobacterial Hsp60 administered in incomplete Freund’s adjuvant (IFA) inhibited disease. Afterward many succeeding findings also showed the protective effects of microbial HSPs in various experimental disease models. So it is possible that immune responses to HSPs could be involved in the control of human chronic inflammatory diseases that have distinct, although as-yet-unknown, initiating
(RA), type 1 diabetes, systemic lupus erythematosus, atherosclerosis, dermatomyositis, inflammatory muscle disorders, organ transplantation and cardiac surgery. Although there is no clear and complete data on human diseases, a picture is emerging in which expression of HSPs or immune reactivity to HSPs seems to be related to downregulation of inflammation, rather than with induction or propagation of inflammation. Recent findings have revealed that arthritic synovial-tissue samples from animals with experimentally induced arthritis and patients with RA or Juvenile idiopathic arthritis (JIA) expressed increased levels of HSP60 (Boog, et al., 1992; de Graeff-Meeder, et al., 1990).
In an initial study, T cells from the synovial fluid and blood of patients with JIA or RA proliferated in response to mycobacterial HSP60 (De Graeff-Meeder, et al., 1991). However, in contrast to patients with RA or healthy control individuals, cells from patients with JIA had substantial proliferative responses to self-HSP60. Further studies showed that CD4+ T-cell responses to human HSP60 at the onset of JIA correlated with responses to mycobacterial HSP60 and with a favourable prognosis (Prakken, et al., 1996). So, these findings indicated that microbial-HSP-crossreactive responses of T cells to endogenous human Hsp60 might have a regulatory role in the course of self-remitting JIA.
Similar results are found in type 1 diabetes and atherosclerosis. Immunization with
Hsp60 has been shown to protect against type 1 diabetes in non-obese (NOD) mouse model (Elias, et al., 1990). Cohen and colleagues showed that the Hsp60-derived peptide
p277 could bind human MHC class II molecules and induce proliferative T-cell responses in patients with type 1 diabetes (Abulafia-Lapid, et al., 1999; Cohen, 2002). In addition, while the rabbits and mice receiving immunization of mycobacterial Hsp60 increased disease in the presence of atherosclerosis-promoting genetic and dietary conditions (Wick, et al., 2004), more recent studies have shown that mycobacterial Hsp60, when administered orally or intranasally, has the potential to suppress induced atherosclerosis (Harats, et al., 2002; Maron, et al., 2002).
4. Potential mechanisms for microbial or endogenous Hsp60s of immune-regulation
The analysis of immune responses to Hsp60 in experimental models and in patients has indicated the capacity of HSPs to induce regulatory T-cell responses. It is possible for HSPs with unique characteristics that endow them with this regulation-inducing capacity. Such characteristics might be related to the interactions of HSPs with receptors of the innate immune system, such as TLRs, or to their stress inducibility or their molecular conservation, which results in immune crossreactivity between bacterial and mammalian homologoues.
CD40, TLRs and scavenger receptors CD36, CD91 and lectin-type oxidized LDL receptor 1 (LOX1) (Binder, et al., 2004; Delneste, 2004). Such immune activation might contribute to the breaking of tolerance to autoantigens, such as Hsp70, leading to the induction of autoimmune disease (Millar, et al., 2003). In the previous studies, both human and microbial Hsp60 have been found to activate pro-inflammatory responses through TLR-2/4 (Takenaka, et al., 2004; Zhao, et al., 2007). However, recent researches have shown that mammalian Hsp60 has a direct innate effect on T cells through binding to TLR2 (Zanin-Zhorov, et al., 2003) and results in activation of T-cell adhesion and inhibition of chemokine-receptor expression. In addition, Hsp60 has recently been shown to modulate the expression of transcription factors involved in TH1-versus TH2-cell differentiation: Hsp60 downregulates expression of the TH1-cell promoting transcription factor T-bet, as well as NF-κB and the intracellular-signaling molecule NFATp, whereas it upregulates the expression of TH2 cell-promoting transcription factor GATA3. this results in decreased secretion of the pro-inflammatory cytokines TNF and IFN-γ and increased secretion of the regulatory cytokine IL-10 by Hsp60-activated T cells (Zanin-Zhorov, et al., 2005).
4.2 Central or peripheral selection of self-HSP-reactive T cells and lacking of co-stimulatory molecules generate HSP-specific regulatory T cells
studies, even in human umbilical cord lymphocytes (Fischer, et al., 1992). In the peripheral immune system, when the cells have left the central lymphoid organs, microbial HSP can be the full agonists for these T cells compatible with immuno-dominant character of these HSP. At the same time, when self-HSP is expressed in the periphery, under conditions of cell stress, the self HSP can act as a partial agonist producing a regulated or actively regulatory response in these T cells.
In addition, at the site of inflammation HSP will be up-regulated in all (stressed) cells of which the majority will be tissue cells lacking co-stimulatory molecules. In the absence of co-stimulatory molecules T cells will adopt a state of anergy or regulation (Taams, et al., 1998). Through these mechanisms HSP-specific T cells can adopt a regulatory phenotype upon antigen recognition in the periphery of the immune system. It is also possible that this tendency of these cells to stay in a tolerant or regulatory state is further promoted by mucosal tolerance in the gut associated lymphoid system (GALT) for abundantly present microbial HSP from the gut microbiota will be dominated by tolerance fro the conversed sequences, as these are shared among the variety of bacterial species present.
immune responses. Hsp70 share almost the same characteristics as Hsp60 on the host immunity. Both mammalian and microbial Hsp70 were found to activate inflammatory responses (Cohen, 1991) while they also suppress NF-κB activation to establish an anti-inflammatory microenvironment at the same time (Bender, et al., 2007; Zheng, et al., 2008). Human Hsp70 levels were in positive correlation with monocyte TLR4, IL-6, and TNF-α expression, which are index for inflammation (Satoh, et al., 2006). Moreover, Campisi and colleagues have revealed that E. coli Hsp72 can stimulate rat splenocytes and macrophages to elevate nitric oxide (NO), TNF-α, IL-1β, and IL-6 to facilitate host recovery from bacterial inflammation (Campisi, et al., 2003). In contrast to Hsp70-mediated immune activation, they also display immune-regulatory effects to protect rats from aggravation of stroke or patients from obesity-induced insulin resistance (Chung, et al., 2008). As for Hsp90, there is a report indicating the inhibition of Hsp90 can attenuate inflammation in endotoxin-induced uveitis (Poulaki, et al., 2007).
6. TGF-β plays an important role in immune-regulatory responses
TGF-β is a regulatory cytokine with pleiotropic effects on cell proliferation,
differentiation, migration, and survival that affect multiple biological processes, including development, carcinogenesis, fibrosis, wound healing, and immune responses (Blobe, et al., 2000). As for immune modulation, TGF-β has also performed multifaceted functions
on a variety of immune cells. TGF-β exerts the greatest impact on T lymphocytes. T cell proliferation, differentiation, and survival are all affected by TGF-β (Cerwenka and Swain, 1999; Gorelik and Flavell, 2002), and it also induces the FoxP3-expressing CD4+CD25+ Tregs and can therefore indirectly influence T cell activation.
TGF-β inhibition of T cell proliferation is influenced by the differentiation status of T cells and the integrated signals from cytokines and costimulatory molecules on T cells, which correlates with reduced TGF-β receptor II expression (Cottrez and Groux, 2001). In addition to influence T cell proliferation, TGF-β also regulates helper T (Th) cell differentiation, and it generally inhibits the acquisition of Th cell functions (Gorelik and Flavell, 2002). TGF-β-mediated inhibition of Th cell differentiation occurs even in the presence of IL-2, in which case T cell proliferation is largely unaffected (Sad and Mosmann, 1994). As for CD8+ cytotoxic T cells, TGF-β has been found to potently inhibit cytotoxic T cell (CTL) differentiation. Early studies showed the CD8+ T cells activated in the presence of TGF-β do not acquire CTL functions (Ranges, et al., 1987), which is likely due to TGF-β inhibition of perforin expression in activated CD8+ T cells (Smyth, et al., 1991). TGF-β also inhibits Fas-ligand (FasL) expression in T cell lines (Genestier, et al., 1999), thereby affecting the death receptor cytotoxic pathway of CD8+
inhibits the expression of multiple effector molecules of CTLs. Investigators have shown that TGF-β can convert CD4+CD25- T cells to Treg in vitro (Chen, et al., 2003; Fantini, et al., 2004; Horwitz, et al., 2003; Wan and Flavell, 2005; Zheng, et al., 2004). Overexpression of TGF-β in the islets of the pancreas also expands the Treg population that protects nonobese diabetic (NOD) mice from type 1 diabetes (Peng, et al., 2004). However, the function of TGF-β in Treg generation/maintenance under physiological conditions remains controversial and poorly understood.
7. The interaction between TGF-β and Hsp60-mediated immunregulation
Since Hsp60 are highly expressed in immune-dysregulated diseases such as RA or JIA (Boog, et al., 1992; de Graeff-Meeder, et al., 1990) and some pathogen infections, the pathogenic role they play in disease progression is becoming attractive. In the animal models of rheumatoid arthritis (RA), T cell responses to self Hsp60 protect against the induction of arthritis. The same results were shown in RA patients since human Hsp60-reactive synovial fluid (SF) T cells produced significantly lower amounts of IFN-γ and higher amount of IL-4 (van Roon, et al., 1997). In the other case, severe injury may lead to immunosuppression with various factors involvement such as hsp60 and hsp70. Locally released Hsps at the site of soft tissue trauma would impaired the capacity of mononuclear cells (MNCs) to release TNF-α and cause monocytopenia restricted to
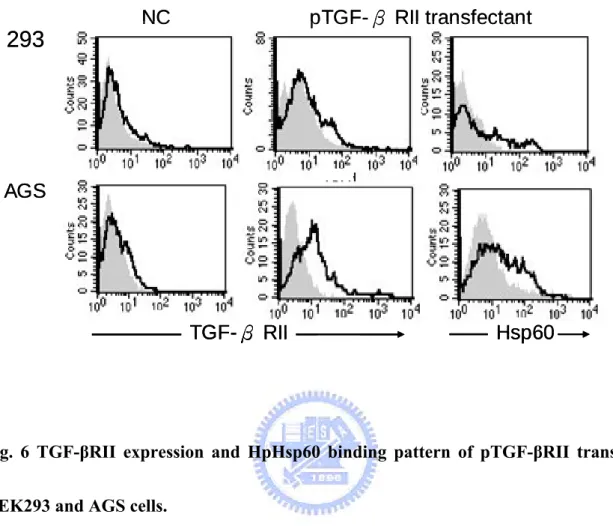
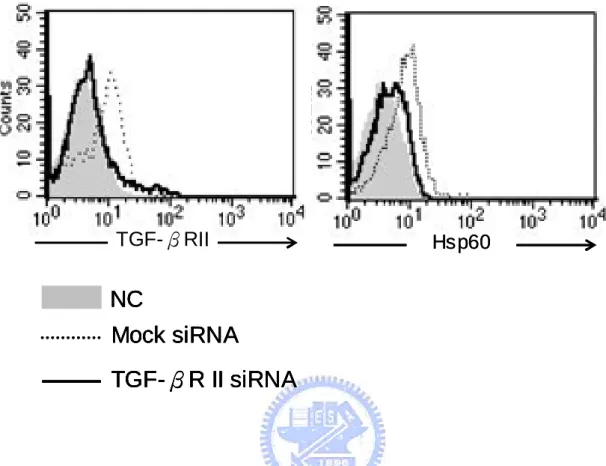
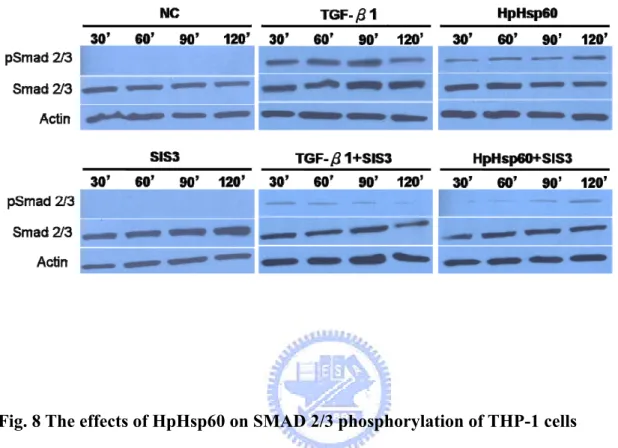
wound fluids (Flohe, et al., 2007). Chlamydia Hsp60 was also associated with activating Th2 immunity and persistent chronic infection (Kinnunen, et al., 2001). Although the exact mechanisms of Hsp60-mediated immune inhibition are still unmasked, previous studies have revealed some hints. Klimartin and the colleagues proved that anti-IL-10 neutralizing antibodies could restore the suppression of TNF-α production of monocytes treated with human Hsp60 (Kilmartin and Reen, 2004). Furthermore, Adjuvant arthritis (AA) patients vaccinated with human Hsp60 or hsp65 genes were found to enhance both IL-10 and TGF-β1 releases (Quintana, et al., 2002). Together, these results shed us twilight about the immunoregulatory effects of Hsp60; however, there is must something new and comprehensive needing us to explore. H. pylori are important factors of gastric cancer, and host immunosuppression caused by them is viewed as one of the major effectors to maintain the persistent infection and carcinogenesis. Since the recent research has indicated that H. pylori Hsp60 (HpHsp60) is vital to pathogen survival and could not be knockout, this protein must play important character in H. pylori infection. In this study, we attempt to investigate the immunoregulatory effects of HpHsp60 from a brand-new aspect. Quite different from previous reports, we found HpHsp60 not only surrender host immunity by secreting immunosuppressive cytokines, they also directly
target genes and suppress immune activities of peripheral mononuclear cells (PBMCs) and monocytic cells. These evidences broaden the view of Hsp60-mediated immunoregulation and strengthen its role in pathogen survival and persistent infection.
REFERENCES
Abulafia-Lapid, R., Elias, D., Raz, I., Keren-Zur, Y., Atlan, H. and Cohen, I. R. (1999). T cell proliferative responses of type 1 diabetes patients and healthy individuals to human hsp60 and its peptides. J Autoimmun 12, 121-9.
Ahmadzadeh, M. and Rosenberg, S. A. (2005). TGF-beta 1 attenuates the acquisition and expression of effector function by tumor antigen-specific human memory CD8 T cells.
J Immunol 174, 5215-23.
Anderton, S. M., van der Zee, R., Prakken, B., Noordzij, A. and van Eden, W. (1995). Activation of T cells recognizing self 60-kD heat shock protein can protect against experimental arthritis. J Exp Med 181, 943-52.
Bender, T. O., Riesenhuber, A., Endemann, M., Herkner, K., Witowski, J., Jorres, A. and Aufricht, C. (2007). Correlation between HSP-72 expression and IL-8 secretion in human mesothelial cells. Int J Artif Organs 30, 199-203.
Binder, R. J., Vatner, R. and Srivastava, P. (2004). The heat-shock protein receptors: some answers and more questions. Tissue Antigens 64, 442-51.
Blobe, G. C., Schiemann, W. P. and Lodish, H. F. (2000). Role of transforming growth factor beta in human disease. N Engl J Med 342, 1350-8.
interleukin-15-mediated interferon-gamma production in human T lymphocytes.
Scand J Immunol 50, 612-8.
Boog, C. J., de Graeff-Meeder, E. R., Lucassen, M. A., van der Zee, R., Voorhorst-Ogink, M. M., van Kooten, P. J., Geuze, H. J. and van Eden, W. (1992). Two monoclonal antibodies generated against human hsp60 show reactivity with synovial membranes of patients with juvenile chronic arthritis. J Exp Med 175, 1805-10.
Bulut, Y., Faure, E., Thomas, L., Karahashi, H., Michelsen, K. S., Equils, O., Morrison, S. G., Morrison, R. P. and Arditi, M. (2002). Chlamydial heat shock protein 60 activates macrophages and endothelial cells through Toll-like receptor 4 and MD2 in a MyD88-dependent pathway. J Immunol 168, 1435-40.
Campisi, J., Leem, T. H. and Fleshner, M. (2003). Stress-induced extracellular Hsp72 is a functionally significant danger signal to the immune system. Cell Stress Chaperones 8, 272-86.
Cerwenka, A. and Swain, S. L. (1999). TGF-beta1: immunosuppressant and viability factor for T lymphocytes. Microbes Infect 1, 1291-6.
Chen, W., Jin, W., Hardegen, N., Lei, K. J., Li, L., Marinos, N., McGrady, G. and Wahl, S. M. (2003). Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J Exp Med 198, 1875-86.
Chung, J., Nguyen, A. K., Henstridge, D. C., Holmes, A. G., Chan, M. H., Mesa, J. L., Lancaster, G. I., Southgate, R. J., Bruce, C. R., Duffy, S. J., Horvath, I., Mestril, R., Watt, M. J., Hooper, P. L., Kingwell, B. A., Vigh, L., Hevener, A. and Febbraio, M. A. (2008). HSP72 protects against obesity-induced insulin resistance. Proc Natl Acad Sci
U S A 105, 1739-44.
Cohen, I. R. (1991). Autoimmunity to chaperonins in the pathogenesis of arthritis and diabetes. Annu Rev Immunol 9, 567-89.
Cohen, I. R. (2002). Peptide therapy for Type I diabetes: the immunological homunculus and the rationale for vaccination. Diabetologia 45, 1468-74.
Cottrez, F. and Groux, H. (2001). Regulation of TGF-beta response during T cell activation is modulated by IL-10. J Immunol 167, 773-8.
Daugaard, M., Rohde, M. and Jaattela, M. (2007). The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett 581, 3702-10.
De Graeff-Meeder, E. R., van der Zee, R., Rijkers, G. T., Schuurman, H. J., Kuis, W., Bijlsma, J. W., Zegers, B. J. and van Eden, W. (1991). Recognition of human 60 kD heat shock protein by mononuclear cells from patients with juvenile chronic arthritis. Lancet 337,
D., Maini, R. N., Kuis, W., Rijkers, G. T. and Zegers, B. J. (1990). Antibodies to the mycobacterial 65-kd heat-shock protein are reactive with synovial tissue of adjuvant arthritic rats and patients with rheumatoid arthritis and osteoarthritis. Am J Pathol 137, 1013-7.
Dean, D., Kandel, R. P., Adhikari, H. K. and Hessel, T. (2008). Multiple Chlamydiaceae species in trachoma: implications for disease pathogenesis and control. PLoS Med 5, e14.
Delneste, Y. (2004). Scavenger receptors and heat-shock protein-mediated antigen cross-presentation. Biochem Soc Trans 32, 633-5.
Elias, D., Markovits, D., Reshef, T., van der Zee, R. and Cohen, I. R. (1990). Induction and therapy of autoimmune diabetes in the non-obese diabetic (NOD/Lt) mouse by a 65-kDa heat shock protein. Proc Natl Acad Sci U S A 87, 1576-80.
F., R. (1962). A new puffing pattern induced by temperature shock and DNP in Drsosophila.
Experientia 18, 571-573.
Fantini, M. C., Becker, C., Monteleone, G., Pallone, F., Galle, P. R. and Neurath, M. F. (2004). Cutting edge: TGF-beta induces a regulatory phenotype in CD4+CD25- T cells through Foxp3 induction and down-regulation of Smad7. J Immunol 172, 5149-53. Fischer, H. P., Sharrock, C. E. and Panayi, G. S. (1992). High frequency of cord blood
1667-9.
Flohe, S. B., Bangen, J. M., Flohe, S., Agrawal, H., Bergmann, K. and Schade, F. U. (2007). Origin of immunomodulation after soft tissue trauma: potential involvement of extracellular heat-shock proteins. Shock 27, 494-502.
Genestier, L., Kasibhatla, S., Brunner, T. and Green, D. R. (1999). Transforming growth factor beta1 inhibits Fas ligand expression and subsequent activation-induced cell death in T cells via downregulation of c-Myc. J Exp Med 189, 231-9.
Gorelik, L. and Flavell, R. A. (2002). Transforming growth factor-beta in T-cell biology. Nat
Rev Immunol 2, 46-53.
Harats, D., Yacov, N., Gilburd, B., Shoenfeld, Y. and George, J. (2002). Oral tolerance with heat shock protein 65 attenuates Mycobacterium tuberculosis-induced and high-fat-diet-driven atherosclerotic lesions. J Am Coll Cardiol 40, 1333-8.
Horwitz, D. A., Zheng, S. G. and Gray, J. D. (2003). The role of the combination of IL-2 and TGF-beta or IL-10 in the generation and function of CD4+ CD25+ and CD8+ regulatory T cell subsets. J Leukoc Biol 74, 471-8.
Jolly, C. and Morimoto, R. I. (2000). Role of the heat shock response and molecular chaperones in oncogenesis and cell death. J Natl Cancer Inst 92, 1564-72.
Kaufmann, S. H., Vath, U., Thole, J. E., Van Embden, J. D. and Emmrich, F. (1987). Enumeration of T cells reactive with Mycobacterium tuberculosis organisms and specific for the recombinant mycobacterial 64-kDa protein. Eur J Immunol 17, 351-7. Kilmartin, B. and Reen, D. J. (2004). HSP60 induces self-tolerance to repeated HSP60
stimulation and cross-tolerance to other pro-inflammatory stimuli. Eur J Immunol 34, 2041-51.
Kinnunen, A., Paavonen, J. and Surcel, H. M. (2001). Heat shock protein 60 specific T-cell response in chlamydial infections. Scand J Immunol 54, 76-81.
Koga, T., Wand-Wurttenberger, A., DeBruyn, J., Munk, M. E., Schoel, B. and Kaufmann, S. H. (1989). T cells against a bacterial heat shock protein recognize stressed macrophages.
Science 245, 1112-5.
Lindquist, S. and Craig, E. A. (1988). The heat-shock proteins. Annu Rev Genet 22, 631-77. Lydyard, P. M. and van Eden, W. (1990). Heat shock proteins: immunity and
immunopathology. Immunol Today 11, 228-9.
Maron, R., Sukhova, G., Faria, A. M., Hoffmann, E., Mach, F., Libby, P. and Weiner, H. L. (2002). Mucosal administration of heat shock protein-65 decreases atherosclerosis and inflammation in aortic arch of low-density lipoprotein receptor-deficient mice.
Circulation 106, 1708-15.
(2003). Hsp70 promotes antigen-presenting cell function and converts T-cell tolerance to autoimmunity in vivo. Nat Med 9, 1469-76.
Munk, M. E., Schoel, B., Modrow, S., Karr, R. W., Young, R. A. and Kaufmann, S. H. (1989). T lymphocytes from healthy individuals with specificity to self-epitopes shared by the mycobacterial and human 65-kilodalton heat shock protein. J Immunol 143, 2844-9. Peng, Y., Laouar, Y., Li, M. O., Green, E. A. and Flavell, R. A. (2004). TGF-beta regulates in
vivo expansion of Foxp3-expressing CD4+CD25+ regulatory T cells responsible for protection against diabetes. Proc Natl Acad Sci U S A 101, 4572-7.
Picard, D., Khursheed, B., Garabedian, M. J., Fortin, M. G., Lindquist, S. and Yamamoto, K. R. (1990). Reduced levels of hsp90 compromise steroid receptor action in vivo.
Nature 348, 166-8.
Poulaki, V., Iliaki, E., Mitsiades, N., Mitsiades, C. S., Paulus, Y. N., Bula, D. V., Gragoudas, E. S. and Miller, J. W. (2007). Inhibition of Hsp90 attenuates inflammation in endotoxin-induced uveitis. Faseb J 21, 2113-23.
Prakken, A. B., van Eden, W., Rijkers, G. T., Kuis, W., Toebes, E. A., de Graeff-Meeder, E. R., van der Zee, R. and Zegers, B. J. (1996). Autoreactivity to human heat-shock protein 60 predicts disease remission in oligoarticular juvenile rheumatoid arthritis. Arthritis
DNA vaccine encoding human heat shock protein 60. J Immunol 169, 3422-8.
Ranges, G. E., Figari, I. S., Espevik, T. and Palladino, M. A., Jr. (1987). Inhibition of cytotoxic T cell development by transforming growth factor beta and reversal by recombinant tumor necrosis factor alpha. J Exp Med 166, 991-8.
Sad, S. and Mosmann, T. R. (1994). Single IL-2-secreting precursor CD4 T cell can develop into either Th1 or Th2 cytokine secretion phenotype. J Immunol 153, 3514-22.
Satoh, M., Shimoda, Y., Akatsu, T., Ishikawa, Y., Minami, Y. and Nakamura, M. (2006). Elevated circulating levels of heat shock protein 70 are related to systemic inflammatory reaction through monocyte Toll signal in patients with heart failure after acute myocardial infarction. Eur J Heart Fail 8, 810-5.
Shinnick, T. M., Vodkin, M. H. and Williams, J. C. (1988). The Mycobacterium tuberculosis 65-kilodalton antigen is a heat shock protein which corresponds to common antigen and to the Escherichia coli GroEL protein. Infect Immun 56, 446-51.
Smyth, M. J., Strobl, S. L., Young, H. A., Ortaldo, J. R. and Ochoa, A. C. (1991). Regulation of lymphokine-activated killer activity and pore-forming protein gene expression in human peripheral blood CD8+ T lymphocytes. Inhibition by transforming growth factor-beta. J Immunol 146, 3289-97.
Taams, L. S., van Rensen, A. J., Poelen, M. C., van Els, C. A., Besseling, A. C., Wagenaar, J. P., van Eden, W. and Wauben, M. H. (1998). Anergic T cells actively suppress T cell
responses via the antigen-presenting cell. Eur J Immunol 28, 2902-12.
Takenaka, R., Yokota, K., Ayada, K., Mizuno, M., Zhao, Y., Fujinami, Y., Lin, S. N., Toyokawa, T., Okada, H., Shiratori, Y. and Oguma, K. (2004). Helicobacter pylori heat-shock protein 60 induces inflammatory responses through the Toll-like receptor-triggered pathway in cultured human gastric epithelial cells. Microbiology
150, 3913-22.
Thole, J. E., Hindersson, P., de Bruyn, J., Cremers, F., van der Zee, J., de Cock, H., Tommassen, J., van Eden, W. and van Embden, J. D. (1988). Antigenic relatedness of a strongly immunogenic 65 kDA mycobacterial protein antigen with a similarly sized ubiquitous bacterial common antigen. Microb Pathog 4, 71-83.
van Eden, W., Thole, J. E., van der Zee, R., Noordzij, A., van Embden, J. D., Hensen, E. J. and Cohen, I. R. (1988). Cloning of the mycobacterial epitope recognized by T lymphocytes in adjuvant arthritis. Nature 331, 171-3.
van Roon, J. A., van Eden, W., van Roy, J. L., Lafeber, F. J. and Bijlsma, J. W. (1997). Stimulation of suppressive T cell responses by human but not bacterial 60-kD heat-shock protein in synovial fluid of patients with rheumatoid arthritis. J Clin Invest
Wick, G., Knoflach, M. and Xu, Q. (2004). Autoimmune and inflammatory mechanisms in atherosclerosis. Annu Rev Immunol 22, 361-403.
Zanin-Zhorov, A., Bruck, R., Tal, G., Oren, S., Aeed, H., Hershkoviz, R., Cohen, I. R. and Lider, O. (2005). Heat shock protein 60 inhibits Th1-mediated hepatitis model via innate regulation of Th1/Th2 transcription factors and cytokines. J Immunol 174, 3227-36.
Zanin-Zhorov, A., Nussbaum, G., Franitza, S., Cohen, I. R. and Lider, O. (2003). T cells respond to heat shock protein 60 via TLR2: activation of adhesion and inhibition of chemokine receptors. Faseb J 17, 1567-9.
Zhao, Y., Yokota, K., Ayada, K., Yamamoto, Y., Okada, T., Shen, L. and Oguma, K. (2007). Helicobacter pylori heat-shock protein 60 induces interleukin-8 via a Toll-like receptor (TLR)2 and mitogen-activated protein (MAP) kinase pathway in human monocytes. J
Med Microbiol 56, 154-64.
Zheng, S. G., Wang, J. H., Gray, J. D., Soucier, H. and Horwitz, D. A. (2004). Natural and induced CD4+CD25+ cells educate CD4+CD25- cells to develop suppressive activity: the role of IL-2, TGF-beta, and IL-10. J Immunol 172, 5213-21.
Zheng, Z., Kim, J. Y., Ma, H., Lee, J. E. and Yenari, M. A. (2008). Anti-inflammatory effects of the 70 kDa heat shock protein in experimental stroke. J Cereb Blood Flow Metab
Zugel, U. and Kaufmann, S. H. (1999). Immune response against heat shock proteins in infectious diseases. Immunobiology 201, 22-35.
Zugel, U., Schoel, B., Yamamoto, S., Hengel, H., Morein, B. and Kaufmann, S. H. (1995). Crossrecognition by CD8 T cell receptor alpha beta cytotoxic T lymphocytes of peptides in the self and the mycobacterial hsp60 which share intermediate sequence homology. Eur J Immunol 25, 451-8.
Characterizing the Polymeric Status of Helicobacter pylori Heat
Shock Protein 60
Abstract
Helicobacter pylori heat shock protein 60 (HpHsp60) was first identified as an adhesion
molecule associating with H. pylori infection. With highly homology among species, however; HpHsp60 has also been investigated as a potent immunogen and elicits host proinflammatory immune responses. Here we have analyzed the structure of HpHsp60 via amino acid blast, circular dichroism and electrophoresis and the results indicated most recombinant HpHsp60s form dimers or tetramers that are quite different than E. coli Hsp60. Treatment of human monocytic cells THP-1 with HpHsp60 was found to up-regulate a panel of cytokines including IL-1α, IL-8, IL-10, IFN-γ, TNF-α, TGF-β1, GRO and RANTES, while carboxymethylated HpHsp60s which were switched their oligomeric status could further enhance NF-κB-mediated IL-8 and TNF-α secretion in THP-1 cells comparing to intact HpHsp60s. The oligomeric status of HpHsp60s may participate in regulating host inflammation to facilitate H. pylori persistent infection.
Introduction
Helicobacter pylori (H. pylori) has been attributed with the pathogenic role in gastric
disease. This organism is associated with pathogenesis of upper gastrointestinal disorders, including chronic gastritis, peptic ulcer, gastric cancer and gastric mucosa-associated lymphoid tissue (MALT) lymphoma (Algood and Cover, 2006). The virulent factors such as CagA (Peek and Blaser, 2002), Vac A (Molinari, et al., 1997), BabA (Boneca, et al., 2003), or
rocF-gene encoded arginase (Gobert, et al., 2001) have been demonstrated to play important
roles in H. pylori colonization and persistent infection.
The expression of heat shock protein (Hsp) 60 (also known as chaperonin (Cpn) 60) by
H. pylori was previously identified as an adhesion molecule to interact with host gastric
epithelial cells and mucin (Huesca, et al., 1996). It was a little contradictory that even though Cpn60 proteins have significant sequence conservation among species, they own extreme immunogenicity (Young DB, 1987). As a potent antigen of H. pylori, Hsp60 could activate myeloid cells to produce proinflammatory cytokines including IL-1β (Maguire, et al., 2005), IL-6 (Lin, et al., 2005; Maguire, et al., 2005), and IL-8 (Maguire, et al., 2005; Zhao, et al., 2007), and it also stimulates KATO III human gastric epithelial cells to secrete IL-8 (Takenaka, et al., 2004).
Although the structure of H. pylori Hsp60s has not been investigated, previous work was established on the resolution of Escherichia coli Hsp60 (Braig, et al., 1994). E. coli Hsp60 is
a porous cylinder of 14 subunits consisting of two approximately 7-mer rotationally symmetrical rings stacked back-to-back. Due to the profound stability of the bacterial chaperonin system and their evolutionary conservation among species, both mammalian and prokaryotic Hsp60s have been assumed to function in a similar way. However, an increasing number of studies are showing that there are variations in structures and functions of other species’ Hsps (Chen and Sigler, 1999; Shimamura, et al., 2004; Zahn, et al., 1996). Possibly, these differences reflect a special adaptation of specific homologues to their microenvironments and endogenous substrates.
We have cloned and expressed recombinant H. pylori Hsp60 (rHpHsp60) and analyzed the structure via sequence alignment, gel electrophoresis and circular dichrosim. Although it contains highly conserved amino-acid sequences and similar secondary structure compositions, the oligomerization of Hsp60s from H. pylori and E. coli are quite different. This difference has been reflected on the divergence of proinflammatory cytokine profiles produced by monocytic cell line THP-1 treated with Hsp60s. Blocking the thiol group within Cys residue of H. pylori Hsp60 altered its molecular conformation and the abilities of cytokine inductions and NF-κB activation. This would imply that HpHsp60 may regulate inflammatory responses via variant oligomer composition.
Materials & Methods Cell culture
Human monocytic cell line THP-1 was obtained from Bioresource Collection and
Research Center (HsinChu, Taiwan) and maintained in RPMI media supplemented with 0.05 mM 2-mercaptoethanol, 2 mM L-glutamine, 50 μg/ml of Penicillin/Streptomycin and 10% heat-inactivated FBS (Biological Industries, Haemek, Israel) at 37℃, 5% CO2.
Amino acid sequence analysis
The amino acid sequences of H. pylori and E. coli hsp60 were aligned using the Vector
NTI Suite 9 software (Invitrogen, USA). Variable amino acid positions and their relative frequencies were identified from the alignment.
Expression and purification of H. pylori Hsp60
Genomic DNA of H. pylori as the template, the DNA fragments containing the gene of Hsp60 was amplified by polymerase chain reaction and cloned into T7 promoter-driven pET 30a (+) expression vector (Novagen, Darmstadt, Germany). The sequences of HpHsp60 were sequenced, verified and submitted to Genbank (accession No.: DQ674277). The proteins were then expressed in Escherichia coli (BL21 strain) and purified using HisTrap affinity (Ni-NTA) chromatography (General Electric, NY, USA) followed by sephadex G-25 column (General
Electric, NY, USA) to remove salts associated with proteins. The purity of HpHsp60 was determined by SDS-PAGE, western blot assay (detected by HRP conjugated goat polyclonal antibody to polyhistidine; Novus, CO, USA) and confirmed with mass spectrometry analysis.
Carboxymethylation of H. pylori Hsp60
For carboxymethylation, 5 mg of H. pylori Hsp60 were dissolved in 5 ml of 0.1 M Tris-HCl buffer (pH 8.6) containing 5.4 M urea and 1% v/v β-mercaptoethanol. After being flushed with nitrogen and incubated at room temperature for 2 h, the reaction mixture was added with 20 mg of iodoacetic acid and maintained at pH 8.6 by the addition of 0.1 M NaOH for another 2 h. In the end, the carboxymethylated (CM)-HpHsp60 was dialyzed in 1X PBS buffer (pH 7.4) to desalt.
Gel electrophoresis
Reducing SDS-PAGE was carried out on 10% using 1 mm thick slab gels with the BioRad MiniProteanTM 3 system (BioRad, CA, USA). The final volume of the resolving gel was 4.0 ml with 1.3 ml of 30% acrylamide solution, 1 ml of 1.5 M Tris-HCl (pH 8.8), 1.6 ml of distilled water, 40 μl of 10% SDS, 20 μl of 10% APS, and 3.6 μl of TEMED. Proteins
incubated in 95℃ for 15 minutes and then loaded into each well and electrophoresis was run. All procedures were followed as described above for non-reducing SDS-PAGE except for removing β-mercaptoethanol from the 3X sample buffer. Native-PAGE was performed as sequential processes on 7.5% native gels as SDS-PAGE except SDS was definitely excluded from 1X sample tracking dye, stacking/separating gels, and electrophoresis running buffer. Proteins analyzed by native gel were directly incubated with 1X sample tracking dye without boiling.
Circular dichrosim (CD) spectroscopy
CD measurements for the H. pylori Hsp60 were conducted on AVIV Model410 Circular
Dichroism spectropolarimeter. Instrument optics and sample chamber were continuously flushed with 10 l/ml of dry N2 gas. The purified proteins were dissolved in 0.1 M phosphate
buffered saline (PBS) at pH 7.4 to obtain a protein concentration of 0.5 mg/ml. Spectra were recorded in the far-ultraviolet (UV) region (200-260 nm). Protein samples were measured at room temperature in a 1 mm path-length quartz cell (300 μl) by using a scan speed of 50 nm/min, a time response of 1 s, a bandwidth of 1 nm and average of 8 scans.
Human protein cytokine array
per well and incubated at 37 in a 5% CO℃ 2 atmosphere for 2 hours. 10 μg HpHsp60s were
then added into cells. After 16-h incubation, supernatants were harvested and assayed by human protein cytokine array (RayBiotech, GA USA) according to the manufacturer’s instructions. Membranes were briefly blocked with a blocking buffer, and then 1 ml of medium from either HpHsp60-treated or untreated THP-1 cell culture was added and incubated at room temperature for 2 hours. After incubation, membranes were treated for 1 hour with biotin-conjugated anti-cytokine antibodies, reacted with horseradish peroxidase-sreptoavidin and then developed using ECL.
IL-8 and TNF-α production inducted by H. pylori Hsp60 treatments on THP-1 cells THP-1 cells were seeded in the same condition described in previous description.
Following, 10 μg naïve or CM-HpHsp60 proteins were added into the well for a 16-h incubation period. Supernatants were then harvested and cytokines were assayed for TNF-α and IL-8 by DuoSet® ELISA development systems and operated according to the manufacturer’s specifications (R & D Systems, MN, USA).
pylori Hsp60-treated THP-1 cell. Transfections of THP-1 cells with pNF-κB-hrGFP were
carried out by electroporation method with Multiporator® (Eppendorf, Hamburg, Germany). THP-1 cells (3 × 106 cells/ml) were incubated with 75 μg of plasmids followed by electroporation (350 V, 80 μs, 1 pulse). The cells were harvested at 48 hours after electro-transfection and hrGFP expression was assayed by flow cytometer.
Statistical analysis
The results are presented as means ± SEM. Significant differences were evaluated with ANOVA . A P value of < 0.05 was considered statistically significant.
Results
Analysis of amino acid sequences of E. coli and H. pylori Hsp60
The sequences of E. coli and H. pylori hsp60 proteins were retrieved from the NCBI
Protein database (http://www.ncbi.nlm.nih.gov/sites/entrez? db=Protein). The accession numbers of E. coli and H. pylori Hsp60 proteins are AAS75782 and DQ674277 respectively. The protein sequence alignment showed that the positives and identity between HpHsp60 and
E. coli Hsp60 are 73.0% and 59.9% (Fig. 1a). There is only one cysteine in the protein
sequence of HpHsp60 (blue frame) while there is three cysteines in E. coli Hsp60 (red frame). The results of circular dichroism measurements were provided for an overview of the secondary structures of H. pylori Hsp60 proteins. Spectra were recorded in 0.1 M phosphate buffered saline (PBS) at pH 7.4 with a protein concentration of 0.5 mg/ml (Fig. 1b). The quantitative evaluations of the data in terms of secondary structure components were shown in figure 2b. The CD spectra of H. pylori Hsp60 displayed a major α-helix conformation composed of 42.2% regular and 24.7% distorted forms, 15.3% β-sheet (with 3.5% regular and 11.8 distorted forms), 8.3% Turn and 9.7% Unordered structure (Fig. 1c).
proteins were composed of monomer and only few could form the dimmer by intermolecular disulfide bonds. Results of native PAGEs indicated that there were three major forms for the rHpHsp60. According to the migration rate of molecular weight marker, recombinant H.
pylori Hsp60 could form multiple oligomerization consisting of dimer, tetramer, and decamer
(Fig. 2b). The results indicated that the cystein residue should not be involved in oligomerization of rHpHsp60 by intermolecular disulfide bonds.
Cytokine expression profiles of THP-1 cells induced by HpHsp60
Microarray membranes were scanned after reacted with the culture medium of THP-1
cells with or without HpHsp60 treatment (Fig. 3a) and the density of each spot was assessed by densitometry. The relative intensities of each cytokine profiles were respectively normalized to control spots on each membrane. The data showed conditioned medium from HpHsp60-treated THP-1 cells up-regulated several different target cytokines such as IL-1α (1.86 fold comparing to NC), IL-8 (11.67 fold), IL-10 (2.03 fold), TNF-α (1.86 fold), IFN-γ (1.79 fold), and TGF-β1 (1.5 fold). The chemokines such as GRO (2.24 fold) and RANTES (1.52 fold) were also elevated in response to HpHsp60 (Fig. 3b).
The effect of cysteine residue in H. pylori Hsp60 on the structure and the induction activities of proinflammatory cytokines
Although the cysteine residue contained within H. pylori Hsp60 did not involve in the formation of intermolecular disulfide bond, dissimilar distribution between E. coli and H.
pylori Hsp60 is concerned if cysteine residues were relevant to any function. Therefore, the
thiol group of cysteine residue within H. pylori Hsp60 was carboxymethylated and sequentially analyzed its status of oligomerization. The migration rate of CM-modified HpHsp60 became slower than the rHpHsp60 and the status of oligomeric structure changed when the only thiol group of HpHsp60 switched to the methyl group (Fig. 4a).
To determine whether the conformation change induced by carboxymethylation of Cys could cause the influence on the activities of HpHsp60, the proinflammatory cytokine releases of THP-1 cells induced by intact or CM-modified HpHsp60 were investigated. The results showed CM-modified HpHsp60 could elicit higher proinflammatory cytokine secretions, since IL-8 and TNF-α production were significantly elevated (Fig. 4 b,c, p < 0.05) comparing to those induced by rHpHsp60. It is known that activation of NF-κB pathway results in the transactivation of a multiple responsive genes that contribute to the inflammatory phenotype including IL-8 (Berger, 2002) and TNF-α (Simmonds and Foxwell, 2008) from monocytes, so the status of NF-κB activation is next investigated. CM-modified HpHsp60 was found to drive more NF-κB activation than rHpHsp60 did (Fig. 4d, p < 0.05).
Discussion
Hsp60s are among the most conserved proteins in all living organisms, either prokaryotic
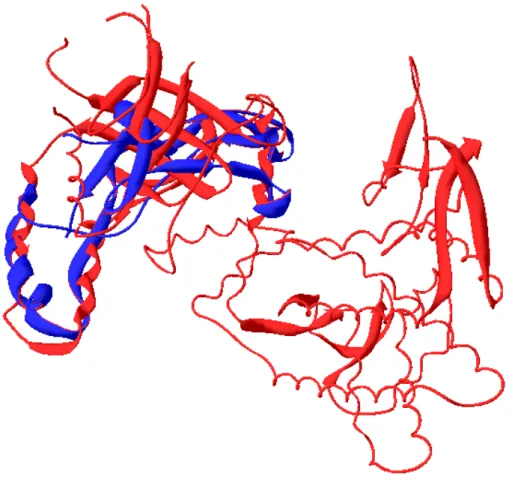
or eukaryotic. X-ray-determining structures from E. coli (Braig, et al., 1994) revealed that the Hsp60 oligomers are a porous cylinder of 14 subunits consisted of two approximately 7-fold rotationally symmetrical rings stacked back-to-back. Each Hsp60 subunit contains three structurally distinct domains: a large equatorial domain that constructs the foundation of the assembly at the waist and brings the ring together, an apical domain that forms the end of the cylinder, and a small intermediate domain that connects the two. However, although previous studies have been continuously disclosed that Hsp60s (or named as GroEL) from some prokaryotes or even yeast (Houston, et al., 1990) and pea chloroplast (Hendrix, 1979) have similar structures by electromicroscope analysis, no current evidence indicates that all Hsp60s shall possess the exact same pattern in response to truly complicated microenvironments of diverse species. The alignment in figure 1 between the Hsp60 amino acid sequences of H.
pylori and E. coli shows over 70% positive sequences among them. It shows that the two
Hsp60 genes have close relationship in evolution but still can not indicate they have the same activities or oligomerization. Furthermore, it has been shown that the ATP bound to GroEL is in direct contact with Cys-137 (Bochkareva and Girshovich, 1994), however, HpHsp60 did not own this cysteine residue. This revealed that even though there is high conservation between protein sequences, the Hsp60s coming from different species could have variant
activities or conformation. Thus, the oligomerization of H. pylori Hsp60 analyzed by native gel electrophoresis demonstrated that it is strictly different from E. coli Hsp60, since most HpHsp60 could form dimers and tetramers instead of heptameric E. coli Hsp60 (Fig. 2b). Hsp60 class is one of the two families of chaperonins which have been identified, with members in the bacterial cytoplasm (GroEL) and in endosymbiotically derived mitochondria (Hsp60) and chloroplasts (Krishna, et al., 2007). Being chaperonins, Hsp60s are essential in mediating ATP-dependent polypeptide chain folding in a variety of cellular compartments. In addition to protein-folding assistants, Hsp60s are also designated as a potent immune regulator. Both mammalian and microbial hsp60s functions not only as active immunogens, but also regulate host innate or adaptive immunity (Tsan and Gao, 2004; van Eden, et al., 2005). The conserved sequences and multifaceted properties of Hsp60s imply that it is important for all species to help the organisms adapt to certain improper circumstances. Therefore the different oligomerization happened in certain species is reasonable.
It is known that hsp60s play important roles in triggering host immune responses. Several microbial homologues have been reported they can elicit pro-inflammatory responses. The responses comprise the induction of inflammatory mediators such as IL-6, TNF-α and NO (Tsan and Gao, 2004) and the activation of the T helper cells type 1 by promoting
exposed by this study. Similar to Hsp60s derived from other species (Maguire, et al., 2005), H.
pylori Hsp60 could stimulate THP-1 monocytes to produce IL-1α, IL-8, and TNF-α which
manage the host inflammation. On the other hand, H. pylori Hsp60 also engages in IL-10 and TGF-β1 induction which potentially manipulates immune regulation and GRO and RANTES secretion associated with chemoattraction (Sager, et al., 1991), cellular communication, survival and differentiation (Wong and Fish, 2003). Although sharing ~70% homology with E.
coli Hsp60, their physiological functions seem not exactly the same. The findings are
supported by previous reports that although H. pylori, Chlamydia and human Hsp60s also share homologous amino acid sequences, their potency in cytokine inductions such as IL-1β, IL-6, and IL-8, and vascular endothelial cell adhesion protein expressions are quite different from each other (Maguire, et al., 2005).
Although the single cystein residue within HpHsp60 was not involved in the formation of intermolecular disulfide bond, but it did contribute to HpHsp60 structural integrity and be relevant to proinflammatory cytokines IL-8 and TNF-α production. H. pylori Hsp60 was found to induce IL-8 release in both gastric epithelial cells (Takenaka, et al., 2004) and monocytes/macrophages (Zhao, et al., 2007) through the Toll-like receptor (TLR)-2/4-mediated signal pathway. By engagement with TLR-2/TLR-4, HpHsp60 activates mitogen-activated protein kinase (MAPK), extracellular signal-regulated kinase (ERK) and p38 and then induces the nuclear translocation of NF-κB to trigger TNF-α and IL-8
production (Zhao, et al., 2007). Our data was consistent with the previous reports that HpHsp60 could significantly stimulate THP-1 cells to release IL-8 and TNF-α via NF-κB activation; however, cystein blockage of HpHsp60 was found to strengthen this proinflammatory response. CM-modified HpHsp60 inspired the exacerbation of both NF-κB transcription factor/DNA binding activity and following-up cytokine secretions in monocytic cells. This phenomenon may disclose the importance for the oligomerization of HpHsp60 in inflammation. With proper structural composition, HpHsp60 could moderate host immune responses and may facilitate their survival and persistent infection within gastric microenvironments.
In this study, we have shown results for the oligomerization of HpHsp60. Quite different from other homologues, HpHsp60 could form dimers and tetramers and the oligomeric status which could affect the functions to activate NF-κB signaling and induce the releases of proinflammatory cytokines.
Figures and Legends
b.
200 220 240 260 280 300 -40 -30 -20 -10 0 10 20 30 40 md eg Wavelength(nm)c.
0 10 20 30 40 S ec onda ry s truc ture ra ti o(% )Fig. 1 Primary and Secondary structure analysis of H. pylori Hsp60. (a) Sequence
alignment of H. pylori and E. coli Hsp60s. The amino acid sequences of E .coli and H. pylori Hsp60s were retrieved from Pubmed database and analyzed by Vector NTI Suite 9 software. The blue and red frames indicated the cysteins within H. pylori and E. coli Hsp60s. CD spectra (b) and secondary structure estimation (c) of H. pylori Hsp60. Measurements were performed in 0.1 M phosphate buffered saline (PBS) at pH 7.4.
a.
b.
a.
HpHsp60 treated NC
b.
Human inflammatory cytokine levels elevated from HpHsp60-treated THP-1 cellsHuman Cytokine Protein Fold Increase
IL-8 Interleukin-8 11.67 fold
GRO Growth-related oncogene 2.24 fold
IL-10 Interleukin-10 2.03 fold
TNF-α Tumor necrosis factor-α 1.86 fold
IL-1α Interleukin-1α 1.86 fold
IFN-γ Interferon-γ 1.79 fold
RANTES Regulated on activation normal T cell expressed and secreted
1.52 fold
Fig. 3 Human cytokine up-regulated from THP-1 monocytic cells treated with HpHsp60.
Human protein-array analysis was used to determine the relevance in the release of HpHsp60-stimulated THP-1 cells. (a) Data from the array analyzed with filter 1, treated with HpHsp60 (Left) and filter 2, medium control (Right). (b) The intensities of the relative expression level of cytokines were quantified by densitometer (GE, USA). The values from scans were normalized based on the intensity of control spots on the filter corners, and the level increase for specific cytokines were shown.
a.
b.
0 10 20 30 40 50 60 70 80 NC HpHsp60 CMHpHsp60 IL -8 ( n g /m l) p < 0.05 0 10 20 30 40 50 60 70 80 NC HpHsp60 CMHpHsp60 IL -8 ( n g /m l) p < 0.05 p < 0.05c.
0 200 400 600 800 1000 1200 1400 1600 NC HpHsp60 CMHpHsp60 TN F-α ( pg/ m l) p < 0.05 0 200 400 600 800 1000 1200 1400 1600 NC HpHsp60 CMHpHsp60 TN F-α ( pg/ m l) p < 0.05 p < 0.05d.
Fig, 4 The effects of rHpHsp60 versus CM-HpHsp60s on proinflammatory cytokine induction and NF-κB transcriptional activity. (a) Native-PAGE analysis of HpHsp60 and
carboxymethylated (CM) HpHsp60. (b) & (c) THP-1 cells were incubated with rHpHsp60 (10 μg/ml) for 16 hours at 37 in 5% CO℃ 2 and conditioned media were harvested to be
determined for the IL-8 (b) and TNF-α(c) levels with sandwich ELISA. Data are expressed as means ± SEM for three replicates from one of three representative experiments. (*, P < 0.05.) (d) THP-1 cell were transiently transfected with NF-κB-dependent reporter plasmid (pNF-κB-hrGFP) and cotreatment with the oligomeric or monomeric HpHsp60s. The fluorescence was analyzed with flow cytometer.
Acknowledgments
We thank to Core Facilities for Proteomics and Structural Biology Research (Institute of Biological Chemistry, Academia Sinica, Nakang, Taiwan) and National Synchrotron Radiation Research Center (Hsinchu, Taiwan) in the service of MALDI-TOF-MS and circular dichroism.
REFERENCES
Algood, H. M. and Cover, T. L. (2006). Helicobacter pylori persistence: an overview of interactions between H. pylori and host immune defenses. Clin Microbiol Rev 19, 597-613.
Berger, M. (2002). Inflammatory mediators in cystic fibrosis lung disease. Allergy Asthma
Proc 23, 19-25.
Bochkareva, E. S. and Girshovich, A. S. (1994). ATP induces non-identity of two rings in chaperonin GroEL. J Biol Chem 269, 23869-71.
Boneca, I. G., de Reuse, H., Epinat, J. C., Pupin, M., Labigne, A. and Moszer, I. (2003). A revised annotation and comparative analysis of Helicobacter pylori genomes. Nucleic
Acids Res 31, 1704-14.
Braig, K., Otwinowski, Z., Hegde, R., Boisvert, D. C., Joachimiak, A., Horwich, A. L. and Sigler, P. B. (1994). The crystal structure of the bacterial chaperonin GroEL at 2.8 A.
Nature 371, 578-86.
Chen, L. and Sigler, P. B. (1999). The crystal structure of a GroEL/peptide complex: plasticity as a basis for substrate diversity. Cell 99, 757-68.
Gobert, A. P., McGee, D. J., Akhtar, M., Mendz, G. L., Newton, J. C., Cheng, Y., Mobley, H. L. and Wilson, K. T. (2001). Helicobacter pylori arginase inhibits nitric oxide production by eukaryotic cells: a strategy for bacterial survival. Proc Natl Acad Sci U S A 98,
13844-9.
Hendrix, R. W. (1979). Purification and properties of groE, a host protein involved in bacteriophage assembly. J Mol Biol 129, 375-92.
Houston, L. S., Cook, R. G. and Norris, S. J. (1990). Isolation and characterization of a Treponema pallidum major 60-kilodalton protein resembling the groEL protein of Escherichia coli. J Bacteriol 172, 2862-70.
Huesca, M., Borgia, S., Hoffman, P. and Lingwood, C. A. (1996). Acidic pH changes receptor binding specificity of Helicobacter pylori: a binary adhesion model in which surface heat shock (stress) proteins mediate sulfatide recognition in gastric colonization. Infect
Immun 64, 2643-8.
Krishna, K. A., Rao, G. V. and Rao, K. R. (2007). Chaperonin GroEL: structure and reaction cycle. Curr Protein Pept Sci 8, 418-25.
Lin, S. N., Ayada, K., Zhao, Y., Yokota, K., Takenaka, R., Okada, H., Kan, R., Hayashi, S., Mizuno, M., Hirai, Y., Fujinami, Y. and Oguma, K. (2005). Helicobacter pylori heat-shock protein 60 induces production of the pro-inflammatory cytokine IL8 in monocytic cells. J Med Microbiol 54, 225-33.
Molinari, M., Galli, C., Norais, N., Telford, J. L., Rappuoli, R., Luzio, J. P. and Montecucco, C. (1997). Vacuoles induced by Helicobacter pylori toxin contain both late endosomal and lysosomal markers. J Biol Chem 272, 25339-44.
Peek, R. M., Jr. and Blaser, M. J. (2002). Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat Rev Cancer 2, 28-37.
Pockley, A. G., Muthana, M. and Calderwood, S. K. (2008). The dual immunoregulatory roles of stress proteins. Trends Biochem Sci 33, 71-9.
Sager, R., Haskill, S., Anisowicz, A., Trask, D. and Pike, M. C. (1991). GRO: a novel chemotactic cytokine. Adv Exp Med Biol 305, 73-7.
Shimamura, T., Koike-Takeshita, A., Yokoyama, K., Masui, R., Murai, N., Yoshida, M., Taguchi, H. and Iwata, S. (2004). Crystal structure of the native chaperonin complex from Thermus thermophilus revealed unexpected asymmetry at the cis-cavity.
Structure 12, 1471-80.
Simmonds, R. E. and Foxwell, B. M. (2008). Signalling, inflammation and arthritis: NF-kappaB and its relevance to arthritis and inflammation. Rheumatology (Oxford) 47, 584-90.
Takenaka, R., Yokota, K., Ayada, K., Mizuno, M., Zhao, Y., Fujinami, Y., Lin, S. N., Toyokawa, T., Okada, H., Shiratori, Y. and Oguma, K. (2004). Helicobacter pylori heat-shock protein 60 induces inflammatory responses through the Toll-like