Estimating the Effects of Sediment from a
Bypass Tunnel on the Management of
Attached Algal Biomass
Yukio Miyagawa, Yukio Onoda, Masanao Sueyoshi, Yuichi Kayaba, Keigo Nakamura, Tetsuya Sumi, and Yasuhiro Takemon
Abstract
Supplying sediments to sediment deficient areas in downstream reaches of a dam can promote the detachment of overgrown algae. Therefore, permanent sediment supply should be secured to maintain the algal growth under control, e.g., using sediment bypass tunnel, because the attached algae can overgrow again within several months. However, long term effects of sediment supply via bypass tunnels on algal biomass have not been previously assessed. In this study, yearly algal biomass in downstream reaches of a dam equipped with a sediment bypass tunnel was measured to estimate the yearly reduction in the amount of algal biomass. In order to achieve this goal, a model for predicting algal biomass was developed. The model used involved multiple variables because attached algae are easily affected by several factors. The accuracy of the model in predicting algal biomass was proven; then, the changes in algal biomass in the presence and absence of sediments from the bypass tunnel were simulated. In the simulation, each scenario was evaluated using three different values of D60, the index of the average diameter on
riverbeds. The six simulations indicated that the model can quantify two types of effects of the sediment: the direct attack on algae during flood, which is effective in reducing overgrown algae, and the reduction in grain size of the riverbed material, which prevents algae from overgrowing.
Keywords: algae, riverbed, dam, friction force, quantitative assessment model
1 Introduction
Sediment supply to sediment deficient areas can promote the detachment of overgrown algae. This sediment effect on algae was previously proven in a study (Tsujimoto and Tashiro, 2004). However, algae must be monitored after the detachment because they can recover and overgrow again within several months (Biggs et al., 1999). Therefore, sediments should be supplied several times a year to prevent algae from overgrowing. Thus, a sediment supply system, such as a sediment bypass tunnel, may offer a constant control over algal biomass (Kondolf, et al., 2014). However, long term effects of sediment supply via bypass tunnels on algal biomass have not yet been assessed.
In this study, the long term effects of sediment supplied from a bypass tunnel on controlling algae were estimated by comparing algal biomass activity in the presence and absence of sediments.
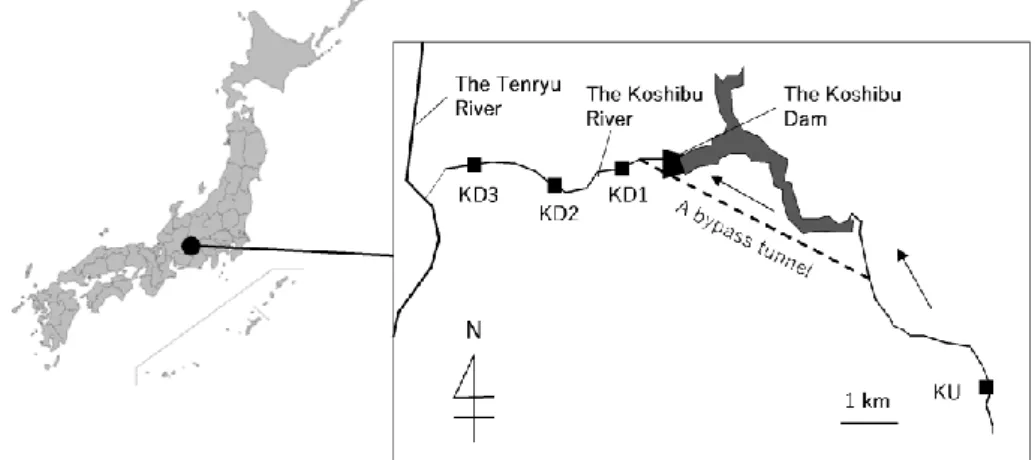
The study was conducted on the bypass tunnel constructed at the Koshibu Dam located at the Koshibu River in Nagano Prefecture, Japan in 2016 (Figure 1). The tunnel was originally constructed to remove part of the sediments from the upstream section to the downstream section of the river to prevent sediment deposition in the reservoir during floods.
Fig. 1: Location of the Koshibu Dam, a bypass tunnel of the dam, and monitoring locations in this study (Ku, KD1, 2, and 3)
2 Study area and methods
2.1 Collection of field monitoring data
Algal biomass was measured at several sites located at the upstream and downstream sections of the Koshibu Dam three or four times per year from April 2016 to December 2017 using the proxy of the primary photosynthetic pigment (chl-a) in mg/m2. During the period, sediments were supplied three times (September 2016, July 2017, and October 2017) to the study area via the bypass tunnel. Herein, they are referred to as the first, the second, and the third sediment. Three survey sites were distributed over 5-km downstream from the Dam (1.0, 2.8, and 4.0 km away from the dam, called KD1, 2, and 3, respectively) at the connection point of the main trunk of the Tenryu River (Figure 1). The upstream site (KU) was chosen to be near the minister office of Oshika village (Figure 1). The chl-a was measured by collecting three stones of moderate sizes (150– 200 mm) from riffles in the river and then scraping the algae from the surface of each stone in a 5 × 5 cm quadrat using a nylon brush. The chl-a content of each algal sample was measured using the SCOR/UNESCO method (SCOR/UNESCO, 1966). Moreover, photon flux densities on the riverbed and distributions of the riverbed material size were measured.
Similar information on algal biomass measured at the same locations from 2014 to 2017 was provided by the Tenryu River Dam Management Office, Chubu Regional Development Bureau, Ministry of Land, Infrastructure, Transport, and Tourism.
Flow and water quality information was obtained from the Hydrology Water Quality Database (http://www1.river.go.jp/). In these data, the volumetric flow at the downstream sites was estimated based on the outflow discharge of the dam, whereas that at the upstream site was estimated based on the inflow. In addition, the following water quality parameters were obtained for all sites: water temperature, and concentrations of suspended solid, nitrogen and phosphorus in the stream.
2.2 Construction of models for estimating algal biomass
The rates of primary production and detachment of algal biomass at KU and KD1 from April 2016 to December 2017 were estimated as follows (Uehlinger, et al., 1996).
De Re Pr dt
dM [1]
where M is the algal biomass content on the stone (mg-chl.a/m2), Pr is the rate of net primary production, Re is the metabolic rate, and De is the rate of detachment. Pr changes from day to night. M is calculated by integrating the previous expression using the daily values of Pr, Re, and De. These daily values are calculated by adding up the 24 hourly values of Pr, Re, and De respectively. The dissolved oxygen (DO) was almost at saturation in all the sites; thus, the equations used to calculate Pr and Re can be constructed following the information reported in Stevenson et al. (1996).
[2] [3] where Pmax and Rmax are the constant values of maximum rate of Pr and Re respectively,
T is the water temperature, Ib is the photon flux density on the riverbed, N and P are the
concentrations of nitrogen and phosphorus, and Ik, Nk, and Pk are the half-saturation
constants (assumed to be 0.05, 0.01, and 50, respectively; Stevenson et al., 1996). Typically, Pr and Re increase in the summer and decrease in the winter because these factors are affected by the water temperature and the photon flux density (eq. 2 and 3). The values of T, Ib, N, and P were measured or assumed based on the availability of the
data to calculate Pr and Re (section 2.1). Pmax and Rmax were also assumed based on the
rate of increase in algal biomass in the sites. This rate was calculated by dividing algal biomass value by the number of days between the day of a heavy flood and that of the observation, because the heavy flood can cause the detachment of algae, thereby the algae may start to growth again. Using this method, the average value of the rate was assumingly 1.33 mg-chl.a/m2/day, and Pmax and Rmax were approximately 0.6 and 0.05,
respectively. P P P N N N I I I P M Pr k k b k b T ( )20 max・1.047 ・ ) 20 ( max1.047 T R M Re ・ ・
Generally, two types of forces can cause the detachment of algae: the internal force caused by the death of old algae and the external force caused by materials in stream water attacking the algae (Biggs, 1996). Herein, the daily detaching rate caused by the internal force was assumed to be around 2.0% of algal biomass in case of mature algae with biomass value of over 50 mg-chl.a/m2. The external force Deex can be estimated based on
the volume of bed load (eq. 4; Tsujimoto and Tashiro, 2004).
De ex = β・γ・qs・d1/3・u*2/3・M [4]
where β is the resistant coefficient of algal detachment, γ is the coefficient related to material property of cobble (=4.94 × 105), q
s is the bed load volume, d is the average
diameter of sediment material particles (assumed 2 mm in this study), and u* is the effective shear velocity on sandy substratum (Tsujimoto and Tashiro, 2004). qs was
calculated using Meyer–Peter and Muller equation (eq. 5; Meyer-Peter and Muller, 1948), and the value u* was computed using Hey’s expression (Hey, 1979).
𝑞∗ = 8.0 ∗ (𝜏∗)32(1 −𝜏𝑐∗
𝜏∗) 3 2
(If τ* < τc*, q* is 0) [5]
β may alter depending on the diameter of the sediment particle and was assumed based
on the rate of algal detachment calculated by comparing algal biomass data in the downstream reaches of the dam before flood and that after the flood.
In eq.5, τ* is the dimensionless tractive force acting on the riverbed calculated via eq.6,
τc* is the dimensionless critical tractive force (assumed 0.05). In eq.6, R is the specific
gravity of bed material in water (=1.65), g is the gravitational acceleration (=9.8), and D60,
the index of average diameter on the riverbed, is the 60 % passing size of the bed material.
𝜏∗= 𝑢∗2
𝑅𝑔𝐷60 [6]
Moreover, the result estimated using eq. 1 was compared to that measured in the sites to confirm the accuracy of the model and to analyze the effects of sediment.
2.3 Estimating the effects of sediment on algal biomass
The algal biomass was estimated at KD1 from April 2016 to December 2017 using the constructed model and the assumed β values (section 2.2). The state of riverbed was considered in this simulation by extracting the changes in D60 from data on riverbed
material distribution. Two simulations were performed: one in the presence of sediments from the bypass tunnel and another in the absence of sediments. Each simulation process was conducted using three values of D60: 200, 100, and 50 mm (i.e., the calculations were
performed six times in total). The values of D60 were fixed during each simulation.
In the constructed model, whether riverbed materials are moved and bed load is caused (τ* > τc* in eq.5) is partly depended on the values of D60, because τ* in eq.5 is smaller as
3 Results
3.1 Assuming β in the model using the algal biomass data
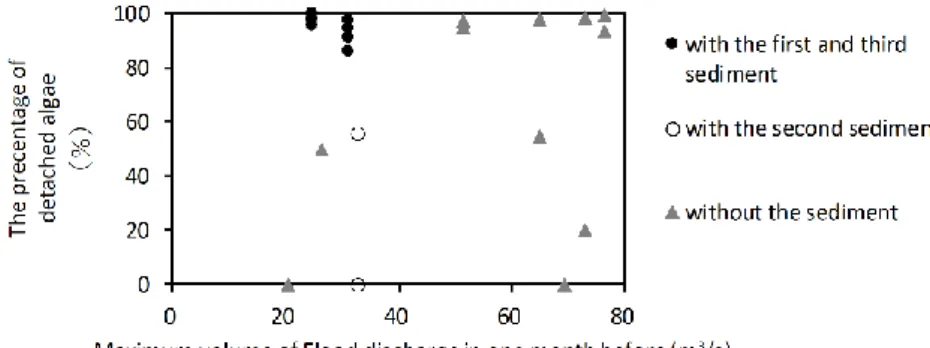
The algal detachment rates were over 80% when the first and the third sediment were supplied from the bypass tunnel (Figure 2). In contrast, the rate was lower (<60%) under a flood with a similar rate (~30 m3/s) in cases of the second sediment and no sedimentation from the bypass tunnel (Figure 2). Floods with no sediments sometimes do not cause a considerable detachment of algae even with flood discharge of over 60 m3/s (Figure 2). Therefore, the value of β in eq. 4 was set to 2.0 × 10-4 in the situations with the first and third sediment and to 2.0 × 10-5 in the other situations (including those without sediment
and with the second sediment).
Fig. 2: Comparison between the algal detachment rates after flood in the presence of sediments and those after flood in the absence of sediments in the downstream areas of the Koshibu Dam
3.2 Determining the model accuracy in estimating algal biomass
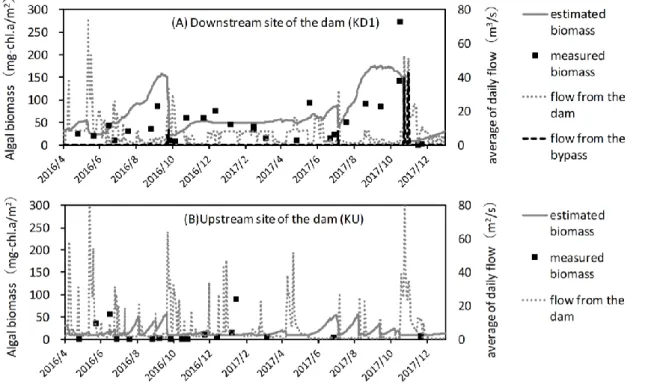
The results of estimating algal biomass in the constructed model showed that the algal biomass at KU and KD1 increased during the normal flow but decreased by the flood (Figure 3). Owing to the monthly flood at KU, algal biomass hardly increased at KU, and its value was always under 50 mg-chl.a/m2 in the estimation. The measured values at KU also showed the same trend. Conversely, some algal biomass values measured at KD1 in the summer and autumn were over 100 mg-chl.a/m2. The value of algal biomass at KD1 estimated using the model also showed an average increase rate of 1.05 mg-chl.a/m2/day, which is almost equal to the measured value of 1.33 mg-chl.a/m2/day (section 2.2).
The average detachment rate of the algae during the first and third sediment at KU1 was almost 90%, whereas it was 30% during the second sediment, similar to the measured values (Figure 2, and 3).
3.3 Estimating effects of sediment on algal biomass
The results of estimating algal biomass under six situations (Section 2.3) showed that the algal biomass decreased with the decrease in D60, particularly in summer and autumn
In addition, in September 2016, there was sudden decrease in algal biomass during the first sediment, not in the case without the first sediment at D60 of 200 or 100 mm (Figure
4). In contrast, at D60 of 50 mm, algal biomass remained under 100 mg-chl.a/m2 even in
the case with no sediment from the bypass tunnel (Figure 4).
Fig. 3: Results of estimating algal biomass at the downstream site (KD1) and the upstream site (KU) of the Koshibu Dam from April 2016 to December 2017 (algal biomass is illustrated by solid lines and dots, whereas the volumetric flow rate is illustrated by broken lines)
Fig. 4: The results of estimating algal biomass at the downstream reaches of the Koshibu Dam (solid lines show the scenario with the sediment from the bypass tunnel, and broken lines show that without the sediment)
4 Discussions
Using the constructed model can roughly estimate the yearly average algal biomass, even though the accurate daily biomass was still difficult to estimate. Furthermore, the model can predict the detachment timing of large algal volumes, such as in the cases of the first
and the third sediment in the flood season. The model can also predict the degree in which the sediment affects algae. This was confirmed by comparing the higher rates of algal detachment in the first and the third sediment and the lower ones in the second sediment (Figure 3). Practically, the calculated volume of the first and third sediment was over seven times higher than that of the second sediment. Thus, the model can predict how degree of flood and sediment may detach overgrown algae. These estimations could be used to establish a plan to manage the sediment supply from a bypass tunnel and algal biomass in the downstream part of a dam.
The sediment from a bypass tunnel may promote the detachment of algae via two mechanisms: directly attacking algae during flood and decreasing the size of riverbed material, which facilitates the movement of materials. The effect of the former mechanism may be predicted by the model because the results of simulations proved that algal biomass in September 2016 after the flood was less in the case with sediment compared to that without sediment (Figure 4). Therefore, for average floods, e.g., September 2016 flood, the effect of sediment is necessary for efficient detachment of algae. On the other hand, heavy floods, e.g., October 2017 flood, could cause complete detachment of algae even if there is no sediment. The model can be also used to estimate the effect of the latter mechanism quantitatively, because the results also proved that yearly algal biomass was restricted with the decrease in the D60 of riverbed materials
(Figure 4). Thus, the former mechanism is effective when algae already overgrows and detaching it is required, whereas the latter mechanism is effective in preventing the overgrowth of algae for a long period. Then, both mechanisms are important to control algal biomass.
In conclusion, the developed model can be used to partly estimate the effect of sediments on algal biomass. However, it isn’t still considered in this model that the sediment buries a part of the riverbed materials and decreases the area where algae are attached. Thus, the effect of sediment is still underestimated in this model. Therefore, another model for estimating algal biomass per square meter of riverbed is under development to target this issue.
5 Conclusions
Herein, a new method was developed for estimating the effect of sediment from a bypass tunnel on algal detachment and control. The values of yearly algal biomass in the presence and absence of sediments were estimated and compared using this method. Results showed that the developed model can estimate the yearly average value of algal biomass and the timing of algal detachment caused by flood. Results also indicated that the model can quantify two types of sediment effects: direct attack on algae during flood, which affects the overgrown algae, and decreasing the size of riverbed material, which prevents the overgrowth of algae.
Acknowledgement
The data used in this study were supplied by the Tenryu River Dam Management Office, Chubu Regional Development Bureau, Ministry of Land, Infrastructure, Transport, and Tourism.
References
Biggs B.J.F. (1996). Hydraulic habitat of plants in streams, River Research and Applications, 12, 131-144.
Biggs, B.J.F., Smith, R.A., Duncan, M.J. (1999). Velocity and sediment disturbance of periphyton in headwater streams: Biomass and metabolism, Journal of the North American Benthological
Society, 18, 222-241.
Hey, R.D. (1979). Flow resistance in gravel-bed rivers. Journal of the hydraulics division, 365-379. Kondolf, G.M., Gao, Y., Annandale, G.W., Morris, G.L., Jiang, E., Zhang, J., Cao, Y., Carling, P., Fu,
K., Guo, Q., Hotchkiss, R., Peteuil, C., Sumi, T., Wang, H.W., Wang, Z., Wei, Z., Wu, B., Wu, C., Yang, C.T. (2014). Sustainable sediment management in reservoirs and regulated rivers: Experiences from five continents, Earth’s Future, 2(5), 256-280.
Meyer-Peter, E., Muller, R., (1948). Formulas for Bed-Load transport. I.A.H.S.R. 2nd meeting,
Stockholm, 39-64.
Ministry of Land, Infrastructure, Transport, and Tourism. (2019). Water Information System. http://www1.river.go.jp. Last access January 29, 2019.
SCOR/UNESCO (1966). Determination of photosynthetic pigments in seawater, Report of
SCOR/UNESCO Working, Group 17, Monograph on Oceanographic methodology, UNESCO,
Paris, 69.
Stevenson, R.J., Bothwell, M.L., Lowe, R.L. (1996). Algal ecology, Academic Press, San Diego. Tsujimoto, T., Tashiro, T. (2004). Application of Population Dynamics Modeling to Habitat Evaluation–
Growth of Some Species of Attached Algae and Its Detachment by Transported Sediment–.
Hydroécologie Appliquée, 14, 161-174.
Uehlinger, U., Buhrer, H., Reichert, P. (1996). Periphyton dynamics in a floodprone prealpine river: Evaluation of significant processes by modelling, Freshwater Biology, 36, 249-263.
Authors
Yukio Miyagawa (corresponding Author) Yukio Onoda Masanao Sueyoshi Yuichi Kayaba Keigo Nakamura Tetsuya Sumi Yasuhiro Takemon
Aqua Restoration Research Center, Public Works Research Institute (PWRI), Japan Email: y-miyagawa@pwri.go.jp