行政院國家科學委員會專題研究計畫 成果報告
子計畫一:屏東大鵬灣及附近海域浮游動物群聚多樣性之研
究與回顧(IV)
計畫類別: 整合型計畫 計畫編號: NSC93-2621-Z-110-002- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立中山大學海洋資源學系(所) 計畫主持人: 羅文增 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 11 月 7 日
行政院國家科學委員會補助專題研究計畫成果報告
高屏海域陸海交互作用及永續經營與管理—子計畫三: 屏東大鵬灣
及附近海域浮游動物群聚多樣性之研究與回顧 (IV)
- 屏東大鵬灣蚵棚拆除前後橈足類群聚組成及豐度分布之比較研究
Community structure and distribution of copepods in Tapong Bay SW of
Taiwan, comparison before and after removal of oyster-culture pens
計畫類別:□個別型計畫! ! ■整合型計畫
計畫編號:NSC 93-2621-Z-110-002
執行期間:
93 年 08 月 01 日至 94 年 07 月 31 日止
計畫總主持人:洪 佳 章
子計畫三主持人:羅
文 增
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立中山大學! 海洋生物科技暨資源學系
中華民國 93 年 10 月 31 日
高屏海域陸海交互作用(III)—
子計畫三:
屏東大鵬灣及附近海
域浮游動物群聚多樣性之研究與回顧
- 屏東大鵬灣蚵棚拆除前後橈足類群聚組成及豐度分布之比較研究
執行單位:國立中山大學海洋資源學系 計畫編號:NSC 93-2621-Z-110-002 計畫主持人:羅 文 增 共同主持人: 研究人員:徐培凱、謝泓諺、廖震亨 聯絡方式:高雄市蓮海路70 號! 國立中山大學海洋生物科技暨資源學系 E-Mail: [email protected]一、中文摘要
本研究是比較大鵬灣蚵架移除前(1999年8月至2001年6月)及移除後(2003 年2月至2004年12月)浮游橈足類群聚組成及豐度之差異外,並探討水文環境因 子對浮游橈足類群聚時空分布可能之影響。結果發現在蚵架移除後大鵬灣內之浮 游橈足類種類數目及豐度均分別較移除前高出數倍之多,但種歧異度指數則差異 不大,浮游橈足類豐度於季節間及測站間差異的幅度亦較在蚵架移除前小。浮游 橈足類種類數目及種歧異度指數經常是北站高於南站,冬季高於夏季之趨向。大 鵬灣內蚵架移除前後浮游橈足類優勢種類組成有一定程度的改變;蚵架移除前五 個 最 優 勢 種 或 分 類 群分 別 為 Acartia sp. 、橈 足 類 無 節 幼 蟲 、 Oithona sp.、 Paracalanus sp.及Paracalanus crassirostris ,這些優勢種彼此間的優勢量十分相當,介於10-15%之間;但蚵架移除後則是以 Oithona oculata 為最優勢種,佔全部
橈 足 類 數 量 的39.8 % , 其 他 優 勢 種 依 序 為 P. crassirostris (10.9 % ) 、 Acartia sinjiensis (9.4%)、Bestolina amoyensis (7.2%)及 Acartia sp. (6.9%)。以主成份分析 法分析結果顯示,蚵架移除前後的橈足類群聚的差異程度較測站間及季節間明
相關,但蚵架移除後則僅和葉綠素a呈顯著的負相關;橈足類種類數目在蚵架移 除前和溫度呈顯著的負相關,但在蚵架移除後和環境因子則無顯著相關。我們綜 合研究結果推測,大鵬灣內蚵架的移除會減低水域浮游植物被牡蠣率食的量,並 降低水中光衰減速率及增加基礎生產力外,也會造成在蚵架移除前經常大量出現 於灣內的海月水母之無性花蟲體因缺乏可著生的基底而明顯地減少,因而也減低 對橈足類的攝食衝擊,而使得橈足類數量較蚵架移除前高出數倍 。 關鍵詞:大鵬灣、 橈足類、種類組成、分布、蚵架
ABSTRACT
This study compares the species composition and abundance of planktonic copepods before (August 1999 to June 2001) with after (February 2003 to December 2004) the removal of oyster culture pens from Tapong Bay and their relationships with hydrographic conditions. The species number and abundance of copepod were apparently higher after removal, but showed no obvious difference in species diversity index between before and after removal. The variation in abundance of copepods between seasons and stations was smaller after removal than before. The species number and diversity index of copepods were generally higher at St. N and winter, and lower at St. C and summer. The species composition of copepod showed difference between before and after removal. Acartia sp., copepod nauplius, Oithona sp., Paracalanus sp. and Paracalanus crassirostris were the 5 most dominant species before the removal, each comprised 10-15% of the total copepods; while Oithona oculata was the most dominant species after removal and comprised up to 39.8% of the total copepods; other common species included P. crassirostris (10.9%)、Acartia sinjiensis (9.4%)、Bestolina amoyensis (7.2%)及 Acartia sp. (6.9%). Result of principle component analysis showed that the variation of copepod assemblage was greater between before and after removal than that between stations and seasons. The abundance of copepod exhibited significant and positive correlations with temperature and chlorophyll a before removal but was negatively correlated with chlorophyll a after
removal; while the species number of copepod showed only negative correlation with temperature before removal. Our results implied that the removal of oyster culture pens from Tapong Bay means exempting the filtering rate of oyster on phytoplankton and reducing the extinction coefficient of light in the water column and consequently increasing the primary productivity; furthermore, it may greatly decrease the abundance of moon-jelly that usually aggregated in the inner Bay and used to settle down their asexual Scyphistoma on the pens before removal. Thus, with relatively low or without feeding impact of moon-jelly, the abundance of copepod was naturally and apparently increasing in the Bay after removal.
Keyword: Copepods, Tapong Bay, Species composition, Distribution, Oyster culture pen.
二、前言
橈足類是海洋浮游動物群聚中最優勢的一個類群,其在海洋生態食物鏈中主 要是扮演次級生產者的角色,大多數以浮游植物為食。近一、二十年來浮游橈足 類在海洋生態系的重要性已廣受到海洋生物學者的瞭解與重視,尤其是其對浮游 植物族群之攝食衝擊及物質通量之影響;例如,Miller et al. (1991)在北太平洋亞極區海域、Landry et al. (1994)在南加州海灣所做之研究結果、及Dam, et al. (1994)
所參與之JGOFS (the Joint Global Ocean Flux Study)計畫在赤道太平洋海域之研究 均證明了此論點。同樣地,浮游橈足類亦為其他海洋較高營養階動物之食物,故 其在海域中之分布狀況亦會影響魚類資源動態。綜合以上所言,可知浮游橈足類 在海洋食物鏈中對於物質與能量之傳輸上有其重要之地位。
橈足類群聚組成之消長容易受到水文環境因子變動之影響,尤其是在河口 域、沿岸海域及較為封閉的潟湖及海灣。例如在Sacca del Canarin 河口的潟湖 區,由於有兩個出口與外洋相通,所以此區的橈足類以全浮游性種類為主(Ferrari et al., 1985);但是在Arcachon海灣,由於只有一個出海口,灣內外水流交換量較 小,因此該海灣以半浮游性橈足種類居多(Castel and Courties, 1982)。此外,
Ferrari et al. (1985)亦發現在漲潮時,以大洋浮游性種類為主,但在退潮時則以底 棲性或半浮游性種類較多。許多學者(Ferrari et al., 1985; Hopcroft et al., 1998 ; Sautour and Castel, 1999)對潟湖區的研究結果亦都發現,橈足類在浮游動物總豐度 中均佔有相當大比例;例如,Sautour and Castel (1999)在法國Marennes-Olron海灣 中發現,橈足類的豐度經常佔全部浮游動物的61~65%,有時更高達90%。
在國內早期關於橈足類之研究十分缺乏,僅有Tan (1967) 及Tseng (1972)進行 概略性的研究;近幾年來才有較多的學者參與,例如,Shih and Young (1995) 首 次收集並編輯台灣附近海域橈足種類之區系名錄,共記載了431種;其他有關橈 足類之研究,包括有攝食行為(Hwang and Turner, 1995)、日週性垂直遷移(Su, 1996)、與仔稚魚之相關性(Cheng, 1998)、種類多樣性與水團的關係 (Shih and Chiu, 1998)、攝食速率 (Hwang, et al., 1998)、河口域之分布(Hsieh and Chiu, 1998)及其 與季風之關係(Chen, et al., 1998)等,均已陸續被發表。近五年來於台灣西南附近 海域有關橈足類的研究亦有數篇文章陸續被發表出來,所發現的橈足種類數目約 有一百種以上 (Shih and Chiu, 1998)。
本研究區域—大鵬灣,為台灣西南部的大型潟湖之一。根據劉(2000)的研究 指出,大鵬灣屬於一淺水且半封閉型的潟湖,平均水深為2.2公尺,水體積為9.7 ×106立方公尺,面積為4.4×106平方公尺,灣內水位的變化主要是由全日潮的混 合潮所造成的。大鵬灣北邊有一出口與外海連接且有一小的魚塭排水溝注入,在 其東邊有牛埔溪支流注入,另外在其東南亦有一大排水溝,主要收集林邊鄉及大 鵬灣附近魚塭排放之廢水,較早期在灣內亦有頗多蚵架及箱網養殖漁業,是屬於 一過度開發利用之高優養化及污染區。民國86年11月政府成立大鵬灣國家風景區 管理處,除了規劃大鵬灣整體發展計畫外,並開始積極整頓灣內各項養殖沉苛, 經多年協調斡旋及各項救濟金的發放,終於在民國91年底全部將灣內的蚵架及箱 網養殖沉苛拆除或移除完畢,讓大鵬灣海域環境景觀大為改變。回顧以往文獻 中,較早期在灣內生態調查有由謝等(1990)及鄭等(1990)進行浮游植物及浮游動物 的基本調查;管理處成立時亦曾委託台加工程顧問股份有限公司(1999)針對灣內 水生生物(包括浮游生物、底棲生物及魚類)進行生態調查。不過,上述生態調查 報告有關浮游動物的研究大多僅止於大類之分類,對於灣內最優勢浮游動物類群
--橈足類的研究則相當缺乏。本研究團隊自民國88年選定大鵬灣海域開始進行研究 至93年止已有五年,期間含蓋了大鵬灣開發前後的採樣監測。本研究乃針對大鵬 灣海域浮游橈足類群聚多樣性之時空分布進行研究,並比較大鵬灣蚵架移除前後 之浮游橈足類群聚組成及豐度之差異外,並探討水文及其他環境因子對浮游橈足 類群聚分布可能之影響。
三、採樣及研究方法
本研究是於1999年8月至2001年6月及2003年2月至2004年12月期間,每兩個 月租用小船在大鵬灣內之北、中及南三個測站(Fig. 1)以北太平洋標準浮游動物網 (網口直徑45 cm,網長180 cm,網目為100 µm)進行近表層之水平拖網,每次 拖網時間為5分鐘,網口中央繫一流量計以估算過濾之水體積。採得之浮游動物 樣品均在船上直接以5~10%之中性福馬林(PH=7.4~7.6)固定保存。每次採樣前 亦以手提式溫鹽儀測定海水之溫度及鹽度,並記錄天候資料,另外亦採取一公升 表水樣品置入暗瓶冷藏,俾便進一步測定水體葉綠素a之濃度。 浮游動物大類之鑑定及計數是先以分樣器將中性福馬林保存之浮游動物樣品 切割子樣品至約含500個橈足類個體,再置於解剖顯微鏡或光學顯微鏡下進行鑑 定計數,並儘可能鑑定至種或屬。海水樣品葉綠素a濃度測定是依照Strickland and Parsons (1972)的方法,以濾膜(孔徑0.45μm,濾膜半徑20 mm)過濾500 ml的海水 後,將濾膜置入5 ml之90%的丙酮內,並放入-20 ℃之恆溫冰箱蔽光保存24小時, 使其充分萃取葉綠素後,以離心機速率3300 rpm離心20分鐘,取上層液以螢光光 度計於加酸前後(0.1N HCl) 測量吸光值,再計算得到葉綠素a濃度。 本研究除了依目的進行基礎之圖表分析外,亦以Shannon種歧異度指數來探 討每個樣品中橈足類群聚種類的多樣性及各種類數量分布之均勻程度,此外亦利 用SYSTATS統計軟體的變方分析(ANOVA)及線性迴歸分析法(Linear regression)來 探討水體溫度、鹽度、葉綠素a等環境因子及橈足類豐度在時空上的差異性及其 間之相關性。浮游橈足類的豐度數據經轉換成對數值後,亦以主成份分析法(Principal component analysis)來分析橈足類群聚在不同月別及測站間資料,再將 所得前兩個主成份軸之轉載值進行點狀分散圖,以探討測站間及季節間橈足類的 群聚組成及數量的變異程度 。
四、結果
大鵬灣內水文環境因子之時空變化
屏東大鵬灣水域溫度及鹽度有明顯的季節變化(Fig. 2)。夏季時水溫經常超過 30℃,尤其是在2000年8月之中站,水溫達31.8℃;最低溫則是出現在1999年12月 的南站(水溫為17℃),而該月的北站水溫則為23℃;顯見冬季時測站間的水溫差 距較夏季大,北站(St. N)的季節性溫度變化範圍明顯較中站(St. C)及南站(St. S) 小,蚵架移除後之水溫季節變化幅度亦略較移除前小。以變方分析結果顯示水溫 在月別間是有顯著性的差異(p < 0.001)(Table 1)。鹽度的季節變化則和水溫相反, 以冬季時相對較高,夏季較低,夏季時灣內南站之鹽度一般明顯較北站低許多, 顯見受到陸源淡水之影響;但蚵架移除後之鹽度季節變化幅度較移除前小了許 多,且年平均鹽度亦較移除前高出許多。以變方分析結果顯示大鵬灣水域鹽度在 測站間、月別間及蚵架移除前後均有顯著性的差異(p < 0.001)。蚵架移除前大鵬 灣的葉綠素a含量在測站間及季節間的變化並沒有明顯及一致性差異,但是在蚵 架移除後季節間的變化頗大,經常在夏季及南站出現較高的葉綠素a含量,蚵架 移除後之葉綠素a年平均含量亦明顯較移除前高出許多。以整體季節性變化來 看,冬季的葉綠素a濃度普遍較夏季時低,不過夏季時各測站之濃度差異較大(Fig. 2)。以變方分析結果顯示葉綠素a濃度在月別間及蚵架移除前後是有顯著性的差異 (p < 0.05,p < 0.001)。大鵬灣地區降雨量也有很明顯的季節變化,在夏季時經常 有甚高的降雨量,在2003年8月時全月累積降雨量高達743 mm,冬季時則甚少降 雨(Fig. 2)。浮游橈足類群聚組成及豐度之時空分布
大鵬灣內蚵架移除後之浮游橈足類平均種類數目及平均豐度均分別較移除前 高出2倍及5.6倍,但平均種歧異度指數則差異不大(Table 2)。在蚵架移除前浮游 橈足類之豐度經常於冬季時出現較低值,接近灣口的北站豐度亦經常高於灣內的 中站及南站,差異可達數十至百倍;不過在蚵架移除後,各季節間及測站間變化 的幅度變得比較小,冬季時也不在出現較低值之現象,反而在2004年12月的南站 出現最高豐度 (Fig. 3)。各測站浮游橈足類種類數目經常以北站較高,南站較低; 冬季時種類數目一般亦較夏季高些。冬季浮游橈足類種歧異度指數一般有較夏季 高的趨向,而南站及中站在冬春季時因少數種類站較高的優勢量而使得種歧異度 指數呈現較低的情形。以變方分析結果顯示大鵬灣浮游橈足類豐度在蚵架移除前 後呈現顯著性的差異(p < 0.001),但在測站間及月別間的差異並不顯著性;橈足 類種類數目在測站間、月別間及蚵架移除前後均有顯著性的差異,但種歧異度指 數則均無顯著差異(Table 1)。 大鵬灣內蚵架移除前後浮游橈足類優勢種類組成也有一定程度的改變(Table2)。蚵架移除前五個最優勢種或分類群(taxa)分別為 Acartia sp. (copepodites, 佔全
部橈足類數量的15.6%)、橈足類無節幼蟲 (copepoda nauplius, 15.3%)、Oithona sp.
(copepodites, 14.0 % ) 、 Paracalanus sp. (copepodites, 13.9 % ) 及 Paracalanus crassirostris (10.0%),這些優勢種多為幼生或小型種,且彼此的優勢量也十分相
當;蚵架移除後則以 Oithona oculata 為最優勢種,佔全部橈足類數量的39.8%,
其他依序為 P. crassirostris (10.9%)、Acartia sinjiensis (9.4%)、Bestolina amoyensis (7.2%)及 Acartia sp. (6.9%)。絕大多數的優勢種或分類群在蚵架移除後的豐度均
明顯較蚵架移除前增加了許多,其中 O. oculata、B. amoyensis、Temora turbinata
及 Acartia erythraea更增加了一百倍以上。各優勢種或分類群在季節及測站間的分 布情況各有不同。O. oculata通常在春夏季時有較高的豐度,並以灣內的南站居 多,不過在蚵架移除後的2004年秋冬季10月及12月南站卻出現甚高的數量,但同 年年初冬季(2月)中站則呈現最低值(Fig. 4)。P. crassirostris是台灣西南沿岸海 域常見的種類,其在接近灣口的北站豐度較高,灣內中及南站明顯較低,在秋冬 季時通常有較高的數量。A. sinjiensis 的豐度以灣內的南站居多,北站則相對少很
多,在降雨量較低的冬春季 通常有較高的數量。Acartia sp. 除了在1999年夏秋季 (8月及10月)外,通常在冬季時有較高的豐度,在蚵架移除前常以北站有較高 的豐度,但是在蚵架移除後則以南站居多。橈足類無節幼蟲豐度在蚵架移除前之 季節及測站間變化趨向不明顯,但是在蚵架移除後則於冬季灣內的南站及中站出 現較高豐度。Bestolina amoyensis通常是在夏季內灣測站豐度較高,並在2003年8 月時出現最高豐度。Oithona sp.在春夏季時出現較高豐度,冬季一般較低,在測 站間的變化趨向則不一致。Oithona dissimilis在蚵架移除前於夏季北站及南站有較
高 的 豐 度 , 但 是 在 蚵 架 移 除 後 則 以 冬 季 時 居 多 。Oithona simplex 及 Oithona brevicornis兩種均以接近灣口的北站數量較高,季節變化不明顯,但分別在2004 年10月及12月於南站出現最高豐度。
橈足類之樣品群分析
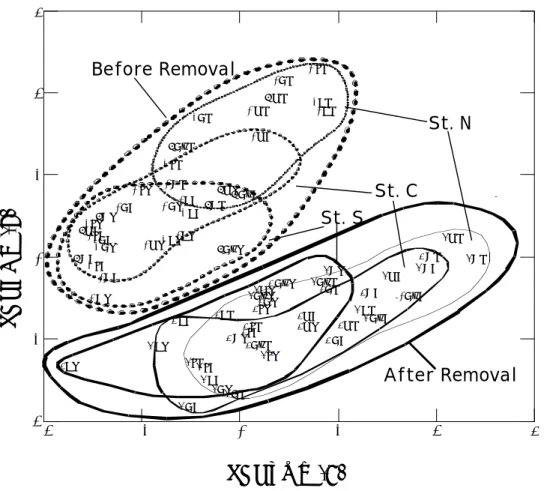
由主成份分析法分析橈足類群聚組成及數量在測站間、季節間及蚵架移除前 後的變異程度結果可知(Fig. 5),蚵架移除前後的橈足類群聚組成及數量是有很明 顯的差異。蚵架移除前及移除後各測站間似乎有由北向南一定程度偏移的趨向, 顯示灣口至灣內測站間之橈足類群聚有一定程度的差異性,但其間有季節性變化 所造成的影響因素;這種測站間的差異程度以蚵架移除前較為明顯,蚵架移除後 測站間的變異範圍重疊程度較高,顯示灣口至灣內測站間橈足類群聚的差異程度 較小。我們嘗試區劃蚵架移除前及移除後各月間及年間的變異範圍,但發現彼此 重疊程度高。浮游動物與環境因子之相關性
以直線迴歸分析結果(Table 3)可知,大鵬灣內浮游橈足類豐度在蚵架移除前 和水域溫度及葉綠素a濃度均呈顯著的正相關(p < 0.01及p < 0.05),但蚵架移除後 則僅和葉綠素a呈顯著的負相關(p < 0.05)。橈足類之種類數目在蚵架移除前和溫 度呈顯著的負相關(p < 0.05),但在蚵架移除後和三種環境因子(溫度、鹽度及葉 綠素濃度)之相關性全部都不顯著。種歧異度指數在蚵架移除前後均和三種環境因子盛不顯著相關。在十個主要優勢種或分類群中,在蚵架移除前只有Oithona sp.
和溫度 及葉 綠素a濃 度呈顯 著的 正相 關(分 別為p < 0.001及p < 0.05);而P.
crassirostris 僅和 葉綠素a濃度呈顯著的正相關(p < 0.05) 顯著的相關。在蚵架移除
後僅有 Oithona dissimilis和鹽度呈顯著的正相關(p < 0.05)和葉綠素a濃度呈顯著的
負相關(p < 0.01);而Acartia sp.及橈足類無節幼蟲則和溫度呈顯著的負相關(分別 為p < 0.01及 p < 0.05) 。
五、討論
屏東大鵬灣是屬於優養化且半封閉的亞熱帶型潟湖,由於水產養殖過度地開 發利用,加上灣內外水體的交換僅靠單一潮口進出,以致於底質沈積物裡的有機 物含量明顯由潮口向灣內遞增 (謝,2000)。根據洪(2000)的研究報告顯示,大鵬 灣高度優養化及高基礎生產力的重要原因可能是水域系統磷過剩所造成,但由於 灣內水體面積不大,其水文及水質狀態易受到海水潮汐、降雨量及廢水排放等影 響,而有明顯的季節性及測站間的變化。高密集蚵棚及箱網養殖區幾乎遍及整個 大鵬灣水域(佔水域面積的82%),更大大地影響潮汐水流的交換速率,因此也 造成潮口與灣內生地化的差異性。本研究結果亦印證了上述的論調,大鵬灣除了 冬季外,一般水溫都能達25℃以上,這可能是位於亞熱帶地區及受到黑潮支流及 南海水所彙集的沿岸水團影響所致(Shaw, 1989;吳,1996);冬季時灣內南站及中 站水溫則明顯受到氣候的影響而較其他季節低,北站雖較易與沿岸海水交換,但 亦較沿岸海域之水溫低了約2℃。大鵬灣地區有明顯的乾濕季節,在夏季(6∼9月) 時之月平均降雨量較其他月份高(Fig. 2)。本研究結果亦顯示大鵬灣在夏季時鹽度 都較其他月別為低,尤其是在南站及中站,鹽度更明顯有淡化的現象。大鵬灣這 種明顯的季節性差異,可能是造成灣內水域橈足類群聚組成及數量在測站間及季 節性消長的重要原因之一。不過在蚵架移除後大鵬灣內各測站間及季節間鹽度變 化的幅度似乎有明顯趨緩的現象,我們推測此可能是由於大鵬灣潮汐水流的交換 速率較蚵架移除前增加所導致,但仍須水流資料來印証。此外,在蚵架移除後大鵬灣內葉綠素a濃度明顯較蚵架移除前增加許多,各測站間及季節間變化的幅度 亦較大,尤其是最內灣的南站。蚵架移除後大鵬灣內葉綠素a濃度明顯地增加似 乎和潮汐水流的交換速率增加的假說有矛盾之處,但是以生物群聚消長的情狀推 測仍有其可道之處。大鵬灣內蚵架的移除必大大地減低水域浮游植物被率食的 量,並可降低水中的光衰減速率及增加基礎生產力。 在以往有關海洋浮游動物的文獻報告,大多著重在中大型橈足類的研究,而 忽略或低估了小型橈足類在海洋生態系的重要性。直到近幾年來,小型橈足類的 研究才逐漸受到重視,尤其是在溫熱帶及較貧營養鹽之海域,其數量經常佔有絕 對的優勢(Morales, et al., 1991);在微生物圈食物網物質傳輸的過程中,小型橈足 類及其幼生亦扮演著重要的角色(Roff, et al., 1995; Wickham, 1995)。例如,Calbet et al. (2001)在地中海西北海域所進行的浮游動物研究亦十分強調小型種的重要 性,在他們的研究結果顯示以53 µm網目所採得的數量是200 µm網目所採得的8 倍。本計畫在1999-2001年的研究亦得到類似的結果,大鵬灣內小型橈足類是浮游 動物最優勢的類群,其中幼生個體即占了絕大部分;小型橈足類在北站的數量經 年地佔浮游動物總豐度的90%以上,但是在南站及中站,可能因水母幼生及枝角 類分別在秋或冬季及春季時偶爾大量出現,而使得小型橈足類不具優勢外,一般 亦能保持在80%以上的優勢量。在蚵架移除後,灣內海月水母無性花蟲體頓時缺 乏可著生的基底,以致於明顯地減少,因而也減低對橈足類的攝食衝擊,而使得 橈足類更具優勢量,並較蚵架移除前高出數倍。 Rios-Jaha (1998)在波多黎哥Phosphorescent海灣的研究指出,在乾濕季節間橈 足類豐度有明顯的差異,橈足類在低溫乾季時的豐度較高溫濕季時多出4倍。大 鵬灣內浮游橈足類豐度也有明顯的乾濕季節變化,但測站間之季節變化趨勢卻不 甚相同,北站橈足類在秋季經常出現較高豐度,南站則在每年夏季出現高峰量, 但是在其他月份均明顯較低。但是在蚵架移除後,隨著灣內水域環境的變動,橈 足類群聚組成及數量也有很大的改變,豐度及種類數目均較蚵架移除前增加許 多。優勢種類組成也有互為消長的現象,在蚵架移除前主要的5個優勢種十分均 勢,每種之相對豐度介於10∼15%之間並有明顯測站間之分布區隔;但是在蚵架 移除後則是由Oithona oculata佔最優勢量,達總豐度的40%(Table 2)。橈足類群聚
組成及數量在蚵架移除前後的變動情狀亦可從主成份分析結果(Fig. 5)得到証 明,蚵架移除前後橈足類群聚的變動較測站間明顯,而測站間的變異趨向似乎又 較月別間明顯。
大鵬灣內浮游橈足類種類組成及分布除受到物理因子(溫度、鹽度及潮汐等) 的影響外,生物本身習性及其他生物因子的影響亦頗為重要。 根據陳及章 (1965)、陳等(1974)、Nishida (1985)、Yamaji (1991) 及Chihara and Murano (1997)
對於橈足種類地理分佈特徵的描述, Oithona spp. 及Acartia spp.是沿岸內灣及海 水或汽水養殖池中常見的類群,可以適應較低鹽度的環境;南站因附近有一大的 排水溝,在夏天雨季來臨時,常會有大量淡水或養殖廢水流入,使得水域鹽度降 低,因此經常成為主要的優勢類群。北站由於較接近灣口,種類數目及豐度經常 較 其 他 測 站 高 , 所 出 現 的 種 類 亦 以 沿 岸 常 見 的 廣 鹽 性 種 類 居 多 , 例 如 P. crassirostris等就可能是隨著潮汐的作用進入灣內並出現在北站,這種類在乾季時 通常有較高的豐度。此外,南站及中站在蚵架移除前於冬季時明顯受到海月水母 攝食的影響,而夏季時則受到養殖池排放水及鹽度淡化的影響較大;北站則相對 較易受到沿岸水團及潮汐的影響。無論是環境因子、橈足類種類組成及季節分佈 特徵,北站皆與其他兩測站有明顯的不同,北站主要以暖水高鹽之沿岸性種類居 多,由於其接近灣口而較易出現大洋性種類,而棲息在南站及中站之種類也可能 會隨著退潮水流至北站,因此外洋種及在南站、中站棲息之種類皆有可能出現於 北站,而使北站的種類數較其他測站高;南站的橈足類種類組成除暖海性灣內種 外,附近養殖池所排入的養殖池種亦為主要優勢種,加上其平均鹽度較北站及中 站低,因此一些河口低鹽性種類(如Genus Acartia)亦經常出現,因此南站的環境 可說是類似河口地區之環境;中站橈足類豐度則可能因水母攝食的影響使得橈足 類豐度較兩測站為低,但是在蚵架移除後南中北測站間橈足類群聚組成數量上差 異的程度似乎較蚵架移除前小一些。海月水母對於浮游動物之攝食衝擊近年來亦 受到許多學者的重視,例如Omori et al. (1995) 在東京灣所進行之研究結果發現, 海月水母的攝食率為2000~3500 prey/day,而大型水母(>14cm)之攝食率更達到 28320~54000 prey/day;Stoecker et al. (1987) 在實驗室針對水母攝食微小型浮游動 物之研究結果指出,水母對於微小型浮游動物之攝食率雖然不高,但是仍可能會
影響生態系中微小型浮游動物之族群量。我們在蚵架移除前於大鵬灣內中站及南
站所進行海月水母的研究結果發現,在冬(乾)季(12~2月)期間,海月水母的數
目明顯較夏(濕)季(6-10月)高出許多,尤其是在2000年1月,海月水母的密度達11
ind./m2,由該季所採得44隻水母樣品進行胃內容物之分析結果,估算共發現有729
個殘體分屬於15個浮游動物類群,其中以橈足類(copepoda)所佔的比例最高,佔 總數量的75%,其次為介形類(ostracoda, 11%),橈足類幼生(copepod nauplius, 5%) 則居第三高量,顯見大鵬灣內海月水母大多以小型的浮游橈足類為食。此外,我 們亦在同一期間研究並估算大鵬灣內橈足類對浮游植物之攝食衝擊程度,結果並 不大(僅3.5%),但數量眾多的橈足類本身卻可能是灣內其他魚蝦蟹類幼生主要的 食物來源,對於灣內水域生態系統物質及能量傳遞的貢獻上仍有其不可輕忽的地 位。
六、參考文獻
Calbet, A., Garrido, S., Saiz, E., Alcaraz, M. and Duarte, C. M. (2001) Annual zooplankton succession in coastal NW Mediterranean waters: the importance of the smaller size fractions. J. Plankton Res. 23: 319-331.
Castel, J. and Courties, C. (1982) Composition and differential distribution of zooplankton in Arcachon Bay. J. Plankton Res. 4: 417-433.
Cheng, C. C. (1998) The distributional correlation between planktonic copepods and ichthyoplankton in the western North Pacific. M.Sc. Thesis, National Taiwan University.
Chihara M. and Murano, M. (1997) An Illustrated Guide to Marine Plankton in Japan. Tokai Uni. 649-1004.
Dam, H. G., Zhang, X., Butler, M., and Roman, M. R. (1994) Mesozooplankton grazing and Metabolism on the Equator at 140° W during the JGOFS EQUPAC study. EOS, Trans Am Geophy Union 75(3): 50.
24-hour cycle of zooplankton sampling in a lagoon of the Po River Delta. J Plankton Res. 7: 849-865.
Hopcroft, R. R., Roff, J. C. and Lombard, D. (1998) Production of tropical copepods in Kinston harbor, Jamaica: the importance of small species. Mar. Biol. 130: 293-604. Hsieh, C. H. and Chiu, T. S. (1998) Abundance composition of the copepods in Tanshui
River estuary and adjacent waters. Acta Zool. Taiwanica 9: 1-9.
Hwang, J. S., Chen, Q. C. and Wong, C. K. (1998) Taxonomic composition and grazing impact of calanoid copepods in coastal waters of northern Taiwan. Crustaceana 71: 378-389.
Hwang, J. S. and Turner, J. T. (1995) Behavior of cyclopoid, harpacticoid and calanoid copepods from coastal waters of Taiwan. Mar. Ecol. 16: 207-216.
Landry, M. R., Lorenzen, C. J., and Peterson, W. K. (1994) Mesozooplankton grazing in the Southern California Bight. II. Grazing impact and particulate flux. Mar. Ecol. Prog. Ser. 115: 73-85.
Miller, C. B., Frost, B. W., Booth, B., Wheeler, P. A., Landry, M. R., and Welschmeyer, N. (1991) Ecological processes in the Subarctic Pacific: Iron limitation cannot be the whole story. Oceanogr. 4: 71-78.
Morales, C. E., Bedo, A., Harris, R. P., and Tranter, P. R. G. (1991) Grazing of copepod assemblages in the north-east Atlantic: the importance of small size fraction. J. Plankton Res. 13: 455-472.
Nishida, S. (1985) Taxonomy and distribution of the family Oithonidae (Copepda, Cyclopoida) in the Pacific and Indian Oceans. Bulletin of the Ocean Research Institute University of Tokyo 1-167.
Omori, M., Ishii, H. and Fujinaga, A. (1995) Life history strategy of Aurelia aurita (Cnidaria, Scyphomedusae) and its impact on the zooplankton community of Tokyo Bay. ICES J. Mar. Sci. 52: 597-603.
Rios-Jaha, E. (1998) Spatial and temporal variations in the zooplankton community of Phosphorescent Bay, Puerto Rico. Est. Coastal Shelf Sci. 46: 797-809.
Roff, J. C., Turner, J. T., Webber, M. K. and Hopcroft, R. R. (1995) Bacterivory by tropical copepod nauplius, extent and possible significance. Aquat. Microb. Ecol. 9:
165-175.
Sautour, B. and Castel, J. (1999) Grazing activity of mesoplanktonic copepods in a shallow bay during an algal spring bloom (Marennes-Oleron Bay, France). J. Mar. Biol. Asso. U.K. 79: 73-84.
Strickland, J. D. H. and Parsons, T. R. (1972) A practical Handbook of seawater analysis. Bull. Fish. Res. Board Can. 167: 1-310.
Shaw, P. T. (1989) The intrusion of water masses into the sea southern of Taiwan. J. Geophys. Res. 94(C12): 18213-18226.
Shih, C. T. and Chiu, T. S. (1998) Copepod diversity in the water masses of the southern East China Sea north of Taiwan. J. Mar. Systems 15: 533-542.
Shih, C. T. and Young, S. S. (1995) A checklist of free-living copepods, including those associated with invertebrates, reported from the adjacent seas of Taiwan. Acta Zool. Taiwanica 6: 65-81.
Stoecker, D. K., Michaels, A. E. and Davis, L. H. (1987) Grazing by the jellyfish, Aurelia aurita, on microzooplankton. J. Plankton Res. 9: 901-915.
Su, C. H. (1996) Vertical migration of five dominant copepods in the northern South China Sea. M. Sc. Thesis, Nat’l Taiwan Ocean Univ., Keelung, pp. 1-60.
Tan, T. H. (1967) Distribution of biomass and abundance of zooplankton in water surrounding Taiwan. Acta Oceanogr. Taiwanica 1: 127-136.
Tseng, W. Y. (1972) On Copepoda of the family Candaciidae in the northeast sea-waters of Taiwan. In: Sugawara, K. (ed.). The Kuroshio II. Proceedings of the Second Symposium on the Results of the Cooperative Study of the Kuroshio and Adjacent Regions. Tokyo. September 28 - October 1, 1970. Saikon Publishing Comp. Ltd. Tokyo.
Wickham, S. A. (1995) Trophic relations between cyclopoid copepods and ciliated protists. Complex interactions link the microbial and classic food webs. Limnol. Oceanogr. 40: 1173-1181.
Yamaji, I. (1991) Illustrations of the Marine Plankton of Japan. Hoikusha Publishing Co., Ltd., Osaka 294-387.
台加工程顧問股份有限公司(1999)交通部觀光局大鵬灣風景特定區水質改善研 究及規劃。交通部觀光局大鵬灣風景特定區管理局研究計畫報告。 吳德泰(1996)高屏峽谷水文特性之調查及研究。國立中山大學海洋資源研究所 碩士論文。 洪佳章(2000)高屏海域生地化作用與通量研究:碳、營養鹽及微量元素(I)。 2000年國科會永續會永續發展研討會論文集摘要。 劉祖乾(2000)高屏海底峽谷大鵬灣交互影響之近岸作用及懸浮沈積物傳輸研 究。2000年國科會永續會永續發展研討會論文集摘要。 鄭新鴻、謝介士、蘇茂森(1990)大鵬灣之生態環境調查-II 浮游生物之分布。省 水產試驗所東港分所。 謝介士、鄭新鴻、蘇茂森、蔡雪貞、葉瑾瑜(1990)大鵬灣之生態環境調查-I 水 質現況。省水產試驗所東港分所。 謝蕙蓮(2000)高屏海域啐屑食物網之研究:啐屑來源及分布(I)。2000年國科會 永續會永續發展研討會論文集摘要。 陳清朝、章淑珍(1965)黃海和東海的浮游橈足類I--哲水蚤目。海洋科學彙刊7: 20-131頁。 陳清朝、章淑珍、朱長壽(1974)黃海和東海的浮游橈足類II--劍水蚤目和猛水蚤 目。海洋科學彙刊9: 27-76頁。
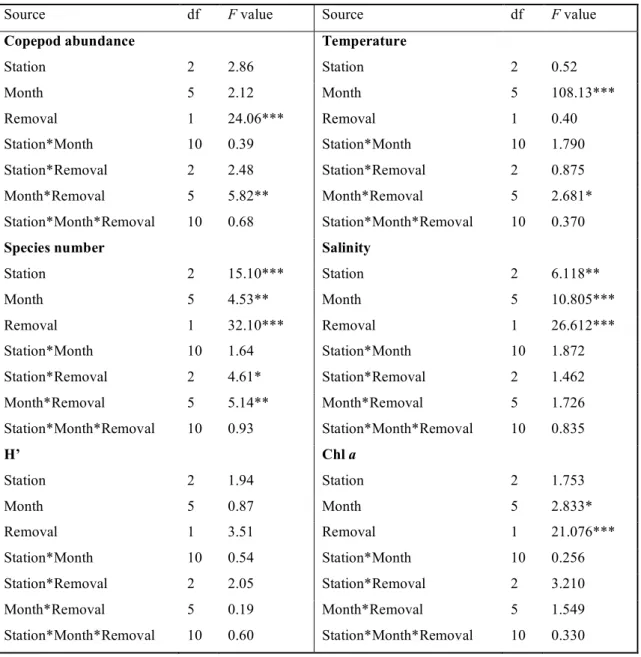
Table 1. Three-way ANOVA of copepod parameters and environmental variables measured at 3 stations in Tapong Bay. *: p < 0.05, **: p < 0.01, and ***: p < 0.001.
Source df F value Source df F value
Copepod abundance Temperature
Station 2 2.86 Station 2 0.52 Month 5 2.12 Month 5 108.13*** Removal 1 24.06*** Removal 1 0.40 Station*Month 10 0.39 Station*Month 10 1.790 Station*Removal 2 2.48 Station*Removal 2 0.875 Month*Removal 5 5.82** Month*Removal 5 2.681* Station*Month*Removal 10 0.68 Station*Month*Removal 10 0.370
Species number Salinity
Station 2 15.10*** Station 2 6.118** Month 5 4.53** Month 5 10.805*** Removal 1 32.10*** Removal 1 26.612*** Station*Month 10 1.64 Station*Month 10 1.872 Station*Removal 2 4.61* Station*Removal 2 1.462 Month*Removal 5 5.14** Month*Removal 5 1.726 Station*Month*Removal 10 0.93 Station*Month*Removal 10 0.835 H’ Chl a Station 2 1.94 Station 2 1.753 Month 5 0.87 Month 5 2.833* Removal 1 3.51 Removal 1 21.076*** Station*Month 10 0.54 Station*Month 10 0.256 Station*Removal 2 2.05 Station*Removal 2 3.210 Month*Removal 5 0.19 Month*Removal 5 1.549 Station*Month*Removal 10 0.60 Station*Month*Removal 10 0.330
Table 2. Mean abundance (individuals/m3) and relative abundance (RA, %) of copepods before (1999-2001) and after (2003-2004) the removal of oyster culture pens in Tapong Bay.
Before removal (B) After removal (A) Copepod species! Mean ± SE RA Mean ± SE RA
Ratio of A/B Oithona oculata 69 ± 42 1.39 11089 ± 5642 39.81 160.7 Paracalanus crassirostris 499 ± 218 10.04 3029 ± 849 10.87 6.1 Acartia sinjiensis 74 ± 67 1.50 2629 ± 1117 9.44 35.5 Acartia sp. 773 ± 450 15.55 1920 ± 1223 6.89 2.5 Copepoda nauplius 759 ± 271 15.28 1575 ± 1238 5.65 2.1 Bestolina amoyensis 16 ± 7 0.31 2002 ± 1038 7.19 125.1 Oithona sp. 696 ± 165 14.00 429 ± 244 1.54 0.6 Oithona dissimilis 412 ± 177 8.29 673 ± 408 2.41 1.6 Oithona simplex 111 ± 79 2.23 848 ± 578 3.04 7.6 Oithona brevicornis 27 ± 9 0.55 831 ± 696 2.98 30.8 Paracalanus sp. 691 ± 211 13.91 32 ± 10 0.11 0.1 Pseudodiaptomus sp. 29 ± 17 0.59 464 ± 316 1.67 16.0 Temora turbinata 1 ± 1 0.03 315 ± 118 1.13 315.0 Paracalanus parvus 3 ± 2 0.06 292 ± 106 1.05 97.3 Acartia tsuensis 312 ± 251 6.28 8 ± 8 0.03 0.03 Acartia erythraea 2 ± 1 0.04 292 ± 148 1.05 146.0 Oithona rigida 0 0 220 ± 174 0.79 - Euterpina acutifrons 0 0 211 ± 54 0.76 - Oithona atteneuta 26 ± 14 0.52 127 ± 55 0.46 4.9 Zausodes sp. 149 ± 46 3.01 0 0 - Other copepods 319 ± 192 6.42 870 ± 630 3.12 2.7 Total copepods 4969 ± 1334 100.0 27858 ± 11023 100.0 5.6
Total species number 36 (15) 69(17) 1.9
Table 3. Linear regression coefficients of the 10 most dominant copepod species in the Tapong Bay. Species as dependent variables are listed in row, and environmental factors as independent variables are listed in columns.
*: p < 0.05, **: p < 0.05, ***: p < 0.001.
Before Removal After Removal Temp. Sal. Chl a Temp. Sal. Chl a
Oithona oculata 0.146 -0.768 -0.403 1.280 -0.665 -0.601 Paracalanus crassirostris 0.599 0.183 2.487* -0.064 -0.222 0.053 Acartia sinjiensis -0.098 -0.578 -0.261 0.235 -0760 -0.454 Acartia sp. 1.740 -0.575 0.832 -2.823** 1.273 -1.592 Copepoda nauplius 1.337 0.552 1.577 -2.680* -0.129 -0.478 Bestolina amoyensis 1.095 -0.976 0.880 1.794 -0.864 0.700 Oithona sp. 4.069*** -0.391 2.597* -1.290 -0.665 -1.496 Oithona dissimilis 0.791 -1.138 -0.314 -1.487 2.159* -3.732** Oithona simplex 0.975 1.481 -0.319 -0.055 0.500 -0.434 Oithona brevicornis 0.348 0.750 0.494 -0.298 -0.618 0.279 Total copepods 2.859** -0.387 2.452* -1.530 0.513 -2.520* Species number -2.431* 1.989 -1.475 -1.881 0.840 -1.949 Species diversity index -0.612 0.187 -1.029 -1.747 0.813 -1.907
Taiwan 23 22 24 25 120 121 122 119 E N Taiwan Strait Pacific Ocean
St. N
St. C
St. S
Tapong Bay
Taiwan Strait
Tung Kang
Datan
Linpan Dike Tidal Inlet 1 kmFig. 1. Location of sampling stations in Tapong Bay, southwestern Taiwan. Fishponds are indicated by the lighter shading.
15 20 25 30 35 Temperature (°C) A O D F A J A O D F A J F A J A O D F A J A O D 20 25 30 35 Salinity (psu) A O D F A J A O D F A J F A J A O D F A J A O D 0 10 20 30 40 Chorophyll a (µg/L) A O D F A J A O D F A J F A J A O D F A J A O D 0 200 400 600 800 Precipataion (mm) A O D F A J A O D F A J F A J A O D F A J A O D Month/Year
Before removal After removal
Mean St. S
St. C St. N
1999 2000 2001 2003 2004
Fig.2. Seasonal changes of Temperature, Salinity, chlorophyll a and precipitation in Tapong Bay before and after the removal of oyster culture pens. Dashed lines indicate annual means.
1 2 3 4 5 6 Abundance (log(n+1)/m 3 ) A O D F A J A O D F A J F A J A O D F A J A O D 0 10 20 30 40 Species number A O D F A J A O D F A J F A J A O D F A J A O D 1.0 1.5 2.0 2.5 3.0 3.5 4.0
Species diversity index (H')
A O D F A J A O D F A J F A J A O D F A J A O D Month/Year 1999 2000 2001 2003 2004 Mean St. S St. C St. N 4.4 3.7 16 20 2.6 2.4
Before removal After removal
Fig.3. Seasonal changes in numerical abundance, species number and diversity of copepods in Tapong Bay before and after the removal of oyster culture pens. Dashed lines indicate annual means.
0 1 2 3 4 5 6 A OD F A J A OD F A J F A J AO D F A J A OD Oithona oculata 0 1 2 3 4 5 6 A OD F A J A OD F A J F A J A OD F A J A OD 0 1 2 3 4 5 6 A O D F A J A O D F A J F A J A O D F A J A O D Acartia sinjiensis 0 1 2 3 4 5 6 A O D F A J A O D F A J F A J A O D F A J A O D Acartia sp. 0 1 2 3 4 5 6 Abundance (log(n+1)/m 3 ) AO D F A J AO D F A J F A J A O D F A J AO D Copepoda nauplius 0 1 2 3 4 5 6 AO D F A J AO D F A J F A J A O D F A J AO D Bestolina amoyensis 0 1 2 3 4 5 6 A OD F A J A O D F A J F A J A OD F A J A OD Oithona sp. 0 1 2 3 4 5 6 AO D F A J AO D F A J F A J A O D F A J AO D Oithona dissimilis 0 1 2 3 4 5 6 A OD F A J A O D F A J F A J A OD F A J A OD Oithona simplex 0 1 2 3 4 5 6 A OD F A J A O D F A J F A J A OD F A J A OD Month/Year Oithona brevicornis 1999 2000 2001 2003 2004 1999 2000 2001 2003 2004 Paracalanus crassirostris
Before removal After removal Before removal After removal
Mean St. S
St. C St.N
Fig.4. Seasonal changes in numerical abundance of the 10 most dominant copepods in Tapong Bay before and after the removal of oyster culture pens.
-2 -1 0 1 2 3 -2 -1 0 1 2 3 3FS 1JC 0DS 1AC 1AS 9DC 9OC1JS0JC 0DC 9DS 0FS 0OS 1FC 0AC 1FS 0AS 9DN 0JS 4FS 1JN 0FC 0DN 9AuN 9OS 4AC 3FC 9AuC 4JN4JC 1AN 4FC 4AS4AN 4DC 4DN 4ON 3DN 4DS 0AuC 4OC 3DC 4AuC 4FN 3ON 3AN 4AuN 0FN 1FN 0JN 3OC 3AC 3AuS 3JS 0AN 3AS 3FN 9ON 3JN 4OS 4AuS 4JS 3JC 0OC 3AuN 0ON 9AuS 3DS 3OS
Fig. 5. PCA plot based on a Pearson correlation matrix of logarithmic abundances of the 41 most common copepods collected in Tapong Bay, southwestern Taiwan, during 1999~2004. Compound symbols represent years (the 1st number: 9, 1999; 0, 2000; 1, 2001; 3, 2003; and 4, 2004), months (F, Feb; A, Apr; J, Jun; Au, Aug; O, Oct; D, Dec) and stations (N, S, and C)
Before Removal After Removal