Differential effect of sorbitol and polyethylene glycol on antioxidant
enzymes in rice leaves
Shih-Yung Hsu and Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China; *Author for correspondence (e-mail: [email protected])
Received 5 March 2002; accepted in revised form 6 June 2002
Key words: Lipid peroxidation, Oryza sativa, Polyethylene glycol, Sorbitol
Abstract
Polyethylene glycol (PEG) and sorbitol (ST) have each been used in osmotically induced water stress studies in plants, however, these osmotica may not have equivalent effects in plants. The present study was designed to examine whether antioxidant enzyme responses in rice leaves are different for PEG and ST of osmotic potential −1.5 MPa. As judged by relative water content, PEG treatment resulted in a higher degree of water stress in rice leaves than ST treatment. PEG treatment markedly increased lipid peroxidation, judged by malondialdehyde con-tent, in rice leaves. However, ST treatment had no effect on lipid peroxidation. An increase in peroxidase (POX), ascorbate peroxidase (APX) and glutathione reductase (GR) activities was observed in rice leaves treated with ST. PEG treatment had no effect on POX and APX activities and decreased GR activity in rice leaves. The de-crease in superoxide dismutase activity induced by PEG was more pronounced than by ST. Cycloheximide blocked the enhanced activities of POX, APX and GR by ST, indicating de novo synthesis of the enzymes. Re-sults suggest that ST but not PEG treatment can up-regulate antioxidant system in rice leaves.
Abbreviations: APX – ascorbate peroxidase, CAT – catalase, GR – glutathionine reductase, MDA –
malondial-dehyde, PEG – polyethylene glycol, POX – peroxidase, RWC – relative water content, SOD – superoxide dis-mutase, ST – sorbitol
Introduction
Drought is a major stress that dramatically limits plant growth and productivity (Boyer 1982). Leaves are known to close their stomata under water stress (Yordanov et al. 2000). During water stress leaf sto-matal closure limits water loss and the influx of CO2.
Lowered CO2influx leads to a decrease in carbon
re-duction by the Calvin cycle and to a decrease in oxi-dized NADP+to serve as an electron acceptor in
pho-tosynthesis. As a result, electrons flow to the alternative electron acceptor, O2, producing
superox-ide radical and consequently other reactive oxygen species, the most damaging of which is the hydroxyl radical (Scandalios 1993). It has been shown that th-ylakoid membrane electron leakage to O2 increased
in sunflower (Sgherri et al. 1996) and in wheat
(Bie-hler and Fock 1996) after drought. Reactive oxygen species can directly damage proteins, amino acids and nucleic acids and cause peroxidation of membrane lipids (Dat et al. 2000).
Plants have evolved specific protective mecha-nisms, involving enzymatic and non-enzymatic anti-oxidants in order to defend themselves against reac-tive oxygen species. Non-enzymatic antioxidants include glutathione and ascorbate (Alscher 1989; Dat et al. 2000), while enzymatic antioxidants include su-peroxide dismutase (SOD, EC 1. 15. 1. 1), catalase (CAT, EC 1. 11. 1. 6), peroxidase (POX, EC 1. 11.1. 7), ascorbate peroxidase (APX, EC 1. 11. 1. 11) and glutathione reductase (GR, EC 1. 6. 4. 2) (Bowler et al. 1992; Dat et al. 2000). It has been demonstrated that GR (Gamble and Burke 1984; Smirnoff and Co-lombé 1988; Baisak et al. 1994; Boo and Jung 1999),
APX (Smirnoff and Colombé 1988; Mittler and Zi-linskas 1992; Baisak et al. 1994; Mittler and Zilins-kas 1994), and POX (Zhang and Kirkham 1994) were all stimulated upon exposure to water stress. On the other hand, SOD (Quartacci and Navari-Izzo 1992; Baisak et al. 1994) and CAT (Mukherjee and Choudhuri (1983, 1985); Quartacci and Navari-Izzo 1992; Zhang and Kirkham 1994) activities which can catalytically scavenge superoxide radical and H2O2,
respectively, declined in water stressed plants. Activi-ties of cytosolic and chloroplastic Cu/Zn SOD in-creased during drought of pea plants (Mittler and Zi-linskas (1992, 1994)) and osmotic stress enhanced Mn SOD transcript abundance in maize (Zhu and Scandalios 1994). It appears that different compo-nents of the reactive oxygen scavenging system are modulated differently under water stress conditions.
Sorbitol (ST) and polyethylene glycol (PEG) have each been used to induce water stress in plant tissue. In experiments where osmotic stress is imposed on plant tissues by ST, it is difficult to separate purely osmotic effects from effects due to water stress gen-erated during such treatments. It is known that PEG does not enter the cell wall space (Rubinstein 1982) and ST does (Flores and Galston 1984). PEG mole-cules with a molecular weight greater than 3000 are apparently not absorbed at all (Tarkow et al. 1966). Thus, PEG and ST may not have equivalent effects. This study was therefore designed to investigate whether antioxidant enzyme responses in rice leaves are different for ST and PEG.
Materials and methods Plant material and treatments
Rice (Oryza sativa c.v. Taichung Native 1) was cul-tured in a stainless net floating on half-strength Johnson’s modified nutrient solution (pH 4.2) in a 500-ml beaker (Lin et al. 1999). The nutrient solution was replaced every three days. Rice plants were grown for 12 days in a greenhouse, under natural light and the day/night temperature of 30/25 °C. The api-cal 3 cm of the third leaf of 12-day-old seedling was used for the experiment. A group of 10 segments floated in a Petri dish containing 10 ml of distilled water served as controls. For induction of water stress, leaf segments were exposed to PEG-6000 or ST solution of osmotic potential −1.5 MPa. All
sam-ples were kept at temperature at 27 °C and irradiance of 40 mmol m−2
s−1
for 4, 8 and 12 h.
RWC, chlorophyll and protein measurements
RWC, defined as water content of leaf tissue as a per-centage of that of the fully turgid tissue, was deter-mined by the method of Weatherley (1950). Chloro-phyll was determined according to Wintermans and De Mots (1965) after extraction in 96% (v/v) ethanol. For protein extraction, leaf segments were homoge-nised in 50 mM sodium phosphate buffer (pH 6.8). The extracts were centrifuged at 17,600 g for 20 min, and the supernatants were used for determination of protein by the method of Bradford (1976) and for en-zyme assays.
Determinations of lipid peroxidation and H2O2
Malondialdehyde (MDA), routinely used as an indi-cator of lipid peroxidation, was extracted with 5% (w/v) trichloroacetic acid and determined according to Heath and Packer (1968). The H2O2 content was
colorimetrically measured, as described by Jana and Choudhuri (1981). H2O2 was extracted by
homoge-nising 50 mg leaf tissue with 3 ml of phosphate buffer (50 mM, pH 6.5). The homogenate was centrifuged at 6 000 g for 25 min. To determine H2O2contents, 3
ml of extracted solution was mixed with 1 ml of 0.1% titanium chloride in 20% (v/v) H2SO4. The mixture
was then centrifuged at 6,000 g for 15 min. The in-tensity of the yellow colour of the supernatant was measured at 410 nm. H2O2 level was calculated
us-ing the extinction coefficient 0.28 mmol−1cm−1.
Determinations of antioxidant enzymes
POX activity was measured using modification of the procedure of MacAdam et al. (1992). Activity was calculated using the extinction coefficient (26.6 mM−1cm−1at 470 nm) for tetraguaiacol. CAT
activ-ity was assayed by measuring the initial rate of dis-appearance of H2O2 (Kato and Shimizu 1987). The
decrease in H2O2 was followed as the decline in
op-tical density at 240 nm, and activity was calculated using the extinction coefficient (40 mM−1cm−1at 240
nm) for H2O2 (Somashekaraiah et al. 1992). SOD
was determined according to Paoletti et al. (1986). APX was determined according to Nakano and Asada (1981). The decrease in ascorbate concentration was followed as the decline in optical density at 290 nm
and activity was calculated using the extinction coef-ficient (2.8 mM−1
cm−1
at 290 nm) for ascorbate. GR was determined by the method of Foster and Hess (1980). One unit of activity for CAT, POX, SOD, APX, and GR were defined as the amount of enzyme which broke down 1 mmol of H2O2per min, caused
the formation of 1 mmol tetraguaiacol per min, inhib-ited 50% the rate of NADH oxidation observed in control, broke down 1 mmol of ascorbate per min, and decreased 1 A340 per min, respectively.
Experimental design
Chlorophyll, protein, H2O2 and MDA contents were
expressed per initial g fresh weight. Enzyme activi-ties were expressed as unit per mg protein. In the present investigation, rice seedlings were grown for 12 days in a greenhouse, where natural light was pro-vided. The growth of rice seedlings is sensitive to light and varies with different light intensities. Exper-iments were carried out at the different time of the year. Thus, absolute levels of each measurement var-ied among experiments because of seasonal effects. However, the patterns of response to PEG or ST were reproducible. For biochemical analysis, four indepen-dent extractions were performed for each treatment. All experiments described here were repeated at least three times. Similar results and identical trends were obtained each time. The data reported here are from a single experiment.
Results
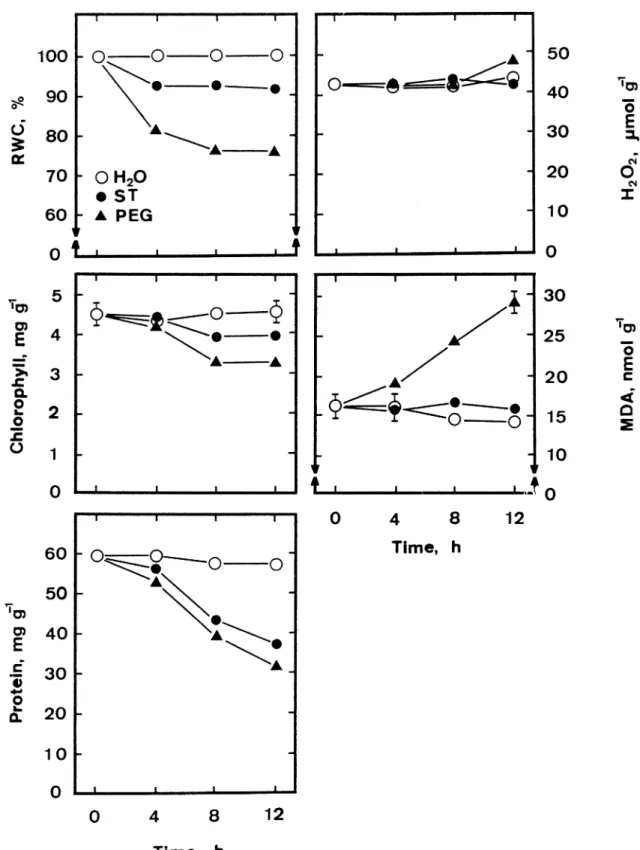
RWC of detached rice leaves exposed to ST or PEG solution of −1.5 MPa, with light exposure, decreased during 12-h of incubation, suggesting that both ST and PEG treatments in our study did indeed cause water stress in the leaves. The decrease in RWC in PEG-treated leaves was greater than that in ST-treated leaves, indicating that water stress induced by PEG is more severe than ST (Figure 1).
The obvious character of leaf senescence is yel-lowing. Chlorophyll loss has been the principal crite-rion of senescence for the largest number of workers. The protein break down during leaf senescence has been realized from earliest studies. In the present pa-per, ST- or PEG-induced senescence of the leaves was assessed by the decrease in chlorophyll and protein contents. It is clear from Figure 1, both ST and PEG decreased chlorophyll and protein contents.
The time courses of lipid peroxidation and H2O2
content in detached rice leaves treated with ST and PEG are given in Figure 1. MDA was used as an in-dicator of lipid peroxidation. MDA contents remained almost unchanged in control leaves during 12-h incu-bation (Figure 1). However, there was a significant increase in MDA content in PEG-treated leaves (Fig-ure 1). It is clear that PEG treatment resulted in an increase in lipid peroxidation in detached rice leaves. In contrast, ST treatment had no effect on lipid per-oxidation (Figure 1). Figure 1 also demonstrated that there was no significant difference in the content of H2O2between control and ST- or PEG-treated leaves.
There was no difference in the activity of CAT, the enzyme responsible for eleminating H2O2, between
control and ST-treated leaves (Figure 2). CAT activ-ity in PEG-treated leaves was lower than water-treated leaves at 4 h after treatment (Figure 2). ST treatment resulted in a higher activity of POX as com-pared with the control leaves (Figure 2). In contrast, no difference in POX activity was observed between PEG- and H2O-treated leaves (Figure 2). Figure 2
also demonstrated that ST treatment resulted in higher activities in SOD, APX and GR than PEG treatment. Since ST treatment caused an increase in the ac-tivities of POX, APX and GR, the effect of cyclohex-imide, an inhibitor of protein synthesis, was studied in order to know if these enhancements were due to new protein synthesis. Cycloheximide treatment pre-vented any ST-induced increase in POX, APX and GR activities (Table 1) indicating that the increases in POX, APX and GR activities in leaves incubated in ST solution are probably due to de novo protein synthesis.
Table 2 shows that rice leaves pretreated with ST resulted in a reduction of toxicity of paraquat, judged by the changes in protein contents.
Discussion
In the present investigation, ST and PEG were used to induce water stress. As judged by RWC, PEG treat-ment resulted in a higher degree of water stress in rice leaves than ST treatment (Figure 1). ST molecules can be absorbed by plant cells (Flores and Galston 1984). Thus, a lower degree of water stress induced by ST seems to result from a certain amount of os-motic adjustment, due to the accumulation of ST. Baisak et al. (1994) reported the enhancement of lipid peroxidation in the leaves subjected to higher degree
Figure 1. Changes in RWC, chlorophyll, protein, H2O2and MDA contents of detached rice leaves floated on water, ST (−1.5 MPa) or PEG
Figure 2. Changes in the activities of antioxidative enzymes in detached rice leaves floated on water, ST (−1.5 MPa) or PEG (−1.5 MPa)
of water stress but not subjected to mild stress. Par-allel to these observations, we also noticed an in-crease in lipid peroxidation in detached rice leaves treated with PEG, on the other hand, detached rice leaves did not exhibit an increase in lipid peroxida-tion with exposure to ST (Figure 1). The occurrence of lipid peroxidation is an indicator of the prevalence of free radical reactions and a change in the balance between O2
·-/H2O2 in leaves. However, we were not
able to provide any evidence for the increased forma-tion of O2
·-. Increase in the contents of H2O2in Vigna
catjang seedlings subjected to water stress had been
reported (Mukherjee and Choudhuri (1983, 1985)). In contrast, Boo and Jung (1999) demonstrated that H2O2content decreased in rice seedlings in response
to water stress. However, neither PEG nor ST had any effect on H2O2 content in detached rice leaves
(Fig-ure 1).
In the present investigation, we were able to show that ST can up-regulate its antioxidant system by in-creasing the activities of POX, APX and GR (Fig-ure 2). We also demonstrated that SOD activity in ST-treated leaves were higher than that in PEG-treated leaves (Figure 2). All these results would ex-plain why ST treatment resulted in no increase in lipid peroxidation.
It has been demonstrated that transgenic plants overexpressing POX, APX and GR had increased re-sistance to paraquat-mediated oxidative stress (Aono et al. (1991, 1993); Sen Gupta et al. 1993; Yun et al. 2000). Thus, it would be interesting to know whether ST-treated detached rice leaves are resistant to paraquat. To test this, detached rice leaves were pre-treated with either ST or water for 12 h and then transferred to either water or paraquat (10M) for 12 h in the light. It is indeed that ST pretreatment re-duced paraquat toxicity, judged by the decrease in protein content (Table 2). It seems that ST induction of POX, APX and GR activities could be a possible cause of reduced paraquat-mediated oxidative stress in rice leaves.
Water stress usually enhances the senescence of leaves (Thomas and Stoddart 1980). We also demon-strated that ST and PEG enhanced senescence of de-tached rice leaves (Figure 1). Since ST had no effect on lipid peroxidation (Figure 2), it is unlikely that ST-enhanced senescence of detached rice leaves is linked to reactive oxygen-mediated lipid peroxida-tion.
Treatment of detached rice leaves with cyclohex-imide, an inhibitor of protein synthesis, prevented ST induced increase in the activities of POX, APX and GR (Table 1) indicating that ST stimulates the de novo synthesis of POX, APX and GR proteins prob-ably as an adaptive mechanism to protect the leaves against oxygen radical damage. It is not known as to how ST lead to the stimulation of POX, APX and GR activities at a time when soluble protein content was declining. It has been shown that O2
·-and H
2O2
gen-erated in the leaves during water stress might be re-sponsible for the induction of GR (Pastori and Trippi 1992; Baisak et al. 1994). Since paraquat treatment, which generates superoxide radical internal, resulted in a decrease in POX activity and H2O2treatment had
no effect on POX activity in detached rice leave (Fang and Kao 2000), it seems that ST-induced POX may not be mediated through reactive oxygen species.
In conclusion, the results presented here suggest (a) that PEG treatment results in higher degree of water stress in detached rice leaves than ST treatment, (b) that PEG but not ST treatment results in a signif-icant increase in lipid peroxidation, and (c) that ST but not PEG treatment can up-regulate antioxidant system. It should be noted that the conclusion ob-tained from the present work is simply based on the studies using detached rice leaves. Since cutting in-creased POX activity and prevented the normal
de-Table 1. Effect of cycloheximide on the induction of POX, APX
and GR activities by ST (−1.5 MPa).
Treatment Enzyme activity, units mg−1protein
POX APX GR
H2O 0.62 ± 0.02 1.29 ± 0.03 0.31 ± 0.03
CHI 0.29 ± 0.03 1.38 ± 0.15 0.36 ± 0.03
ST 2.01 ± 0.09 1.80 ± 0.14 0.60 ± 0.06
ST + CHI 0.32 ± 0.02 1.55 ± 0.06 0.38 ± 0.04 The concentration of cycloheximide was 10M. Enzyme activi-ties were determined 12 h after treatment in the light. Means ± SE, n = 4.
Table 2. Effect of ST pretreatment on contents of protein in
de-tached rice leaves treated with paraquat.
Pretreatment Treatment Protein content, mg m−1FW
H2O H2O 54.23 ± 1.60
Paraquat 11.65 ± 0.33
ST H2O 40.36 ± 1.02
Paraquat 19.59 ± 1.69
Detached rice leaves were pretreated with water or ST (−1.5 MPa) for 12 h and then transferred to either water or paraquat (10M) for 12 h in the light. Means ± SE, n = 4.
crease in CAT activity in tobacco leaves (Parish (1968a, 1968b)), one may argue that the changes in antioxidant enzymes are probably a wound response. When detached leaves were used as an experimental system, wounding is always a problem. However, in the present work, each long and narrow rice leaf was cut once transversely, the aera of wounding was very small. Since no changes in RWC and proline content were observed in control rice leaves during 12-h in-cubation in the light (Figure 1, Cheng et al. (2002) (in press)), the results from the present investigation are unlikely to be associated with the changes in wa-ter-channel function or stomatal movement. Never-theless, detached leaf system lacks the natural fea-tures of transport in the whole plant. Thus, the results from experiments using detached rice leaves are not necessarily similar to those using whole plant system.
Acknowledgements
This work was supported by grant NSC 90-2313-B-002-267 from the National Science Council of the Republic of China.
References
Alscher R.G. 1989. Biosynthesis and antioxidant function of glu-tathione in plants. Physiol. Plant. 77: 457–464.
Aono M., Kubo A., Saji H., Natori T., Tanaka K. and Kondo N. 1991. Resistance to active oxygen toxicity of transgenic
Nic-otiana tabacum that expresses the gene for glutathione
reduc-tase from Escherichia coli. Plant Cell Physiol. 32: 691–697. Aono M., Kubo A., Saji H., Natori T., Tanaka K. and Kondo N.
1993. Enhanced tolerance to photooxidative stress of transgenic
Nicotiana tabacum with high chloroplastic glutathione
reduc-tase activity. Plant Cell Physiol. 34: 129–135.
Baisak R., Rane D., Acharya P.B.B. and Kar M. 1994. Alterations in the activities of active oxygen scavenging enzymes of wheat leaves subjected to water stress. Plant Cell Physiol. 35: 489– 495.
Biehler K. and Fock H. 1996. Evidence for the contribution of the Mehler-peroxidase reaction in dissipating excess electrons in drought-stressed wheat. Plant Physiol. 112: 265–272. Boo Y.C. and Jung J. 1999. Water deficit-induced oxidative stress
and antioxidative defense in rice plants. J. Plant Physiol. 155: 255–261.
Bowler C., Van Montagu M. and Inzé D. 1992. Superoxide dismu-tase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43: 83–116.
Boyer J.S. 1982. Plant productivity and environment. Science 218: 443–448.
Bradford M.M. 1976. A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the princi-ple of protein-dye binding. Anal. Biochem. 72: 248–254. Cheng F.-Y., Hsu S.-Y. and Kao C.H. 2002. Nitric oxide
counter-acts the senescence of detached rice leaves induced by dehy-dration and polyethylene glycol but not by sorbitol. Plant Growth Regul. (in press).
Dat J., Vandenabeele S., Vranová E., Van Montagu M., Inzé D. and Van Breusegem F. 2000. Dual action of the active oxygen spe-cies during plant stress responses. Cell. Mol. Life Sci. 57: 779– 795.
Fang W.-C. and Kao C.H. 2000. Enhanced peroxidase activity in rice leaves in response to excess iron, copper and zinc. Plant Sci. 158: 71–76.
Flores H.E. and Galston A.W. 1984. Osmotic stress-induced polyamine accumulation in cereal leaves. I. Physiological pa-rameters of the response. Plant Physiol. 75: 102–109. Foster J.G. and Hess J.L. 1980. Responses of superoxide
dismu-tase and glutathione reducdismu-tase activities in cotton leaf tissue exposed to atmosphere enriched in oxygen. Plant Physiol. 66: 482–487.
Gamble P.E. and Burke J.J. 1984. Effect of water stress on the chloroplast antioxidant system. I. Alterations in glutathione re-ductase activity. Plant Physiol. 76: 615–621.
Heath R.L. and Packer L. 1968. Photoperoxidation in isolated chlo-roplasts. Arch. Biochem. Biophys. 125: 189–198.
Jana S. and Choudhuri M.A. 1981. Glycolate metabolism of three submerged aquatic angiosperm during aging. Aquat. Bot. 12: 345–354.
Kato M. and Shimizu S. 1987. Chlorophyll metabolism in higher plants. VII. Chlorophyll degradation in senescing tobacco leaves: phenolic-dependent peroxidative degradation. Can. J. Bot. 65: 729–735.
Lin J.N., Wang J.W. and Kao C.H. 1999. Effect of abscisic acid and water stress on the senescence of detached rice leaves. Biol. Plant. 42: 313–316.
MacAdam J.W., Nelson C.J. and Sharp R.E. 1992. Peroxidase ac-tivity in the leaf elongation zone of tall fescue. Plant Physiol. 99: 872–878.
Mittler R. and Zilinskas B.A. 1992. Molecular cloning and charac-terization of a gene encoding pea cytosolic ascorbate peroxi-dase. J. Biol. Chem. 267: 21802–21807.
Mittler R. and Zilinskas B.A. 1994. Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. Plant J. 5: 397–405.
Mukherjee S.P. and Choudhuri M.A. 1983. Implications of water stress induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 58: 166–170.
Mukherjee S.P. and Choudhuri M.A. 1985. Implications on hydro-gen peroxideascorbate system on membrane permeability of water stressed Vigna seedlings. New Phytol. 99: 355–360. Nakano Y. and Asada K. 1981. Hydrogen peroxide is scavenged
by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 22: 867–880.
Paoletti F., Aldinacci D., Mocali A. and Capparini A. 1986. A sen-sitive spectrophotometric method for determination superoxide dismutase activity in tissue extracts. Anal. Biochem. 154: 536– 541.
Parish R.W. 1968a. Stduies on senescing tobacco leaf disks with special reference to peroxidase. I. The effect of cutting and in-hibition of nucleci acid and protein synthesis. Planta 82: 1–13. Parish R.W. 1968b. Sduies on senescing tobacco leaf disks with special reference to peroxidase. II. The effect of interactions of proline, hydroxyproline and kinetin. Planta 82: 14–21. Pastori G.M. and Trippi V.S. 1992. Oxidative stress induces high
rate of glutathione reductase synthesis in a drought-resistant maize strain. Plant Cell Physiol. 33: 957–961.
Quartacci M.F. and Navari-Izzo F. 1992. Water stress and free rad-ical mediated changes in sunflower seedlings. J. Plant Physiol. 139: 621–625.
Rubinstein B. 1982. Regulation of H+excretion. I. Effects of
os-motic shock. Plant Physiol. 69: 939–944.
Scandalios J.G. 1993. Oxygen stress and superoxide dismutase. Plant Physiol. 101: 7–12.
Sen Gupta A., Webb R.P., Holaday A.S. and Allen R.D. 1993. Overexpression of superoxide dismutase protects plants from oxidative stress. Plant Physiol. 103: 1067–1073.
Sgherri C.L.M., Pinzino C. and Navari-Izzo F. 1996. Sunflower seedlings subjected to increasing stress by water deficit: Changes in O2·-production related to the composition of
thy-lakoid membranes. Physiol. Plant. 96: 446–452.
Smirnoff N. and Colombé S.V. 1988. Drought influences the activ-ity of enzymes of the chloroplast hydrogen peroxide scaveng-ing system. J. Exp. Bot. 39: 1097–1108.
Somashekaraiah B.V., Padmaja K. and Prasad A.R.K. 1992. Pho-totoxicity of cadmium ions on germinating seedlings of mung bean (Phaseolus vulgaris): involvement of lipid peroxides in chlorophyll degradation. Physiol. Plant. 85: 85–89.
Tarkow H., Feist W.C. and Southerland C.F. 1966. Interaction of wood with polymeric materials. Penetration versus molecular size. Forest. Prod. J. 16: 61–65.
Thomas H. and Stoddart J.L. 1980. Leaf senescence. Annu. Rev. Plant Physiol. 31: 83–111.
Weatherley P.E. 1950. Studies in the water relation of cotton plant. I. The field measurement of water deficits in leaves. New Phy-tol. 49: 81–97.
Wintermans J.F.G.M. and De Mots A. 1965. Spectrophotometric characteristics of chlorophyll a and b and their pheophytins in ethanol. Biochem. Biophys. Acta 109: 448–453.
Yordanov I., Velikova V. and Tsonev T. 2000. Plant responses to drought, acclimation, and stress tolerance. Photosynthetica 38: 171–186.
Yun B.-W., Huh G.-H., Lee H.-S., Kwon S.-Y., Jo J.-K., Kim J.-S. et al. 2000. Differential resistance to methyl viologen in trans-genic tobacco plants that express sweet potato peroxidases. J. Plant Physiol. 156: 504–509.
Zhang J. and Kirkham M.B. 1994. Drought-stress-induced changes in activities of superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell Physiol. 35: 785–791.
Zhu D. and Scandalios J.G. 1994. Differential accumulation of mangase-superoxide dismutase transcripts in maize in response to abscisic acid and high osmoticum. Plant Physiol. 106: 173– 178.