Volume 15,Number 10,1996
Mary
AnnLiebert,Inc.Pp.
827-844Expression,
Characterization,
and
Genomic Structure of
Carp
JAK1 Kinase Gene
MAU-SUN
CHANG,1
GEEN-DONG
CHANG,2
JIANN-HORNG
LEU,3
FORE-LIEN
HUANG,1-3
CHEN-KUNG
CHOU,4
CHANG-JEN
HUANG,3
andTUNG-BIN
LO2-3
ABSTRACT
A
3.7-kb cDNA encodes the carp
JAK1
kinase of
1,156
amino
acid residues. The overall amino acid
sequenceidentity
between carp
JAK1
andmurine
JAK1, JAK2, JAK3,
andhuman
TYK2is
57%, 35.5%, 31.3%,
and
42.4%,
respectively.
Inaddition,
carp
JAK1
shows
higher
sequence
homology
tomammalian
JAK1
inboth
thekinase-like
(JH2)
and kinase
(JH1)
domains
(approximately
70%
identity).
Therefore,
carp
JAK1
is
aho-molog
of mammalian
JAK1.
To
investigate
the
possible
function of
JH2
domain,
full-length,
and various
trun-cated
forms of
carpJAK1
wereproduced
in the baculovirus
system.
Our
results demonstratethat
c-JHl
and
C-JH2
associate with each other and
C-JH2
can betyrosine-phosphorylated
by
c-JAKl
andby
c-JH(l
+2).
The JAK
I gene wasalso isolated from
acarp
genomic library
and characterized. This
geneis divided into 24
exonsspanning
atleast31
kbof
genomic
DNA. Exon 1 contains
the5'-untranslated
region
and
exon 2con-tains the
putative
translation initiation site. The
2.5-kb DNA
region
upstream
of the
transcription
initiation
site contains
numerouspotential binding
sites for
transcription
factors
including
NF-IL6, HNF-5, API,
GHF-5,
andE2A. When this
DNAfragment
wasplaced
upstream
of the
chloramphenicol acetyltransferase
(CAT)
reporter
gene
andtransfected into
a carpCF
cellline,
it could drive the
synthesis
of CAT enzyme 16 times
more
efficiently
than
thepromoterless
pCAT-Basic.
Deletion
analysis
defined
apositive
regulatory
region
be-tween
—1,023
and—528.
A smallerregion
(—181
to+59)
without
anytypical
TATA-box sequences, G
+C-rich sequences,
or otherbinding
sequencesfor
knowntranscription
factors still
hadpromoter
activity.
Constructs without this
region
did
not have detectablepromoter
activity.
This
suggests
thatthis
region
of
DNA may
play
animportant
rolein
theexpression
of
carpJAK
I gene.INTRODUCTION
of the externalsignals
tothe cell.Upon binding
of theligand,
thekinaseactivity
of thereceptor
PTK isinduced,
resulting
inTYROSiNE
PHOSPHORYLATIONcatalyzed by protein tyrosine
ki-phosphorylation
of thereceptor
and cellular substratesontyro-nases
(PTKs)
isakey
step
intransducing signals
fromex- sine residues(Ullrich
andSchlessinger,
1990).
By
contrast, the ternal stimulitothe nucleus.Therefore,
PTKsplay
veryimpor-
nonreceptor PTKs,
duetothe lack of extracellulardomains,
can-tantroles in the
regulation
of cellproliferation
and differentiation notdirectly
interact with extracellularligands.
Therefore,
the(Hunter
andCooper,
1985;
Hanks etal,
1988).
On the basis ofphysiological
functions of thesenonreceptor
PTKs remained ob-their structuralsimilarities,
PTKscanbe divided intotwomajor
scureforquite
some time until the observation thatp56lck
was groups:receptorandnonreceptorPTKs.Receptor
PTKs contain associated with the CD4 and CD8co-receptors
(Veillette
etal,
extracellular, transmembrane,
andcytoplasmic
domains. Theex-1988).
Thissuggests
thatby
binding
nonreceptor PTKs,
some tracellular domainsareabletoassociate withligands,
where the transmembranereceptorslacking
TKdomainscanactinaman-cytoplasmic catalytic
domains areresponsible
for transmission neranalogous
tothereceptorPTKs.'Department
ofZoology,
and2GraduateInstitute of BiochemicalSciences,National TaiwanUniversity,
Taipei,
Taiwan,instituteof
Biological Chemistry,
AcademiaSinica,Taipei,
Taiwan.4Department
of MedicalResearch,VeteransGeneralHospital, Taipei,
Taiwan.The
polymerase
chainreaction(PCR) (Mullís
andFaloona,
1987)
has beenemployed
toclonepotential
PTKs,
using
de-generate
oligonucleotide primers
corresponding
to conservedpeptide fragments
inkinase domains. The JAK(Janus kinase)
family
wasfirst identifiedby
thisstrategy(Wilks,
1989;
Wilks etal, 1991).
Thisfamily belongs
tothenonreceptor
PTKs andcurrently
consists of JAK1(Wilks
etal, 1991;
Yang
etal,
1993),
JAK2(Harpur
etal,
1992),
JAK3(Witfhuhn
etal,
1994),
and TYK2(Firmbach-Kraft
etal,
1990)
in mammals. These kinases lack the Srchomology
domains SH2 and SH3(Pawson
andSchlessinger,
1993)
butbear an unusual feature ofhaving
akinase-like domain. The functional role of TYK2 was first demonstratedby
itsparticipation
ininterferon(IFN)
signal
transductionpathway
and its association with the IFN-areceptor
(Velazquez
etal,
1992).
Recently,
manycytokine
re-ceptors
have been foundtoassociate with one ortwo or more members of the JAKfamily, including
thereceptors
forery-thropoietin
(EPO) (Witthuhn
etal,
1993),
growth
hormone(Argetsinger
etal,
1993),
prolactin (Campbell
etal, 1994;
Da Silva etal, 1994;
Rui etal,
1994),
interleukin 2(IL-2)
(Miyazaki
etal,
1994;
Russell etal, 1994;
Witthuhnetal,
1994),
IL-3(Silvennoinen
etal,
1993),
IL-4(Witthuhn
etal,
1994),
IL-6(Stahl
etal,
1994),
IL-12(Bacon
etal,
1995),
on-costatinM,
leukemiainhibitory
factor(LIF)
(Stahl
etal,
1994),
granulocyte-macrophage colony-stimulating
factor(GM-CSF)
(Quelle
etal,
1994),
granulocyte
colony-stimulating
factor(G-CSF) (Nicholson
etal,
1994),
andIFN-y (Muller
etal, 1993;
Watling
etal,
1993).
Although lacking
protein
kinasedomains,
the above members of the
cytokine
receptor
superfamily
cancouple ligand binding
with thetyrosine phosphorylation
of downstreameffectors,
which is mediatedby
association with the JAKs. The activated JAKs thenphosphorylate cytoplasmic
transcription
factors STATsor thesignal
transducer andacti-vators of
transcription.
Aftertyrosine
phosphorylation,
thesetranscription
factorsarehomo-orheterodimerized,
translocated into thenucleus,
and inducetranscriptional
responses(Shuai
etal, 1993;
Darnelletal, 1994;
Ihleetal, 1994;
Schindler andDarnell,
1995).
In
fish,
growth
hormone andprolactin
from manyspecies
have been isolated and cloned(Chang
etal, 1992a,b;
Bemardi etal,
1993),
whereasonly
oneIFNfrom flatfish has beenpu-rified and cloned
(Tamai
etal,
1993).
Basedon avariety
of studies frommammals,
the JAKfamily
membersareshowntobe involved in the
signal
transduction ofgrowth
hormone(Argetsinger
etal,
1993),
prolactin (Campbell
etal, 1994;
DaSilvaetal,
1994),
and interferons(Velazquez
etal,
1992).
Therefore,
as aninitialstep
tounderstand thesignal
transduc-tion infish,
westartedtoisolate the carp JAK kinasefamily
by
PCR. As
reported
in this paper,amember of the JAKfamily
wascloned anddesignated
carp JAK1 kinase. The carp JAK1 kinase has ahigher
sequencehomology
in both JH1 and JH2 domains(70%
identity)
tohuman and murine JAK1(Wilks
etal, 1991;
Yang
etal,
1993).
Because the function of the JH2 domain isunknown,
weexplored
itsactivity
inabaculovirusexpression
system,which has been usedtoexpress mammalianJAK2 kinase
(Quelle
etal,
1994;
Duhe andFarrar,
1995).
We made constructs that encodefull-length
JAK1(c-JAKl),
the JH1 and JH2 domains alone(c-JHl
andc-JH2),
and the JH1 and JH2 domains in combination[c-JH(l
+2)]
andexpressed
them in the baculovirus system. Coinfection andimmunopre-cipitation experiments
showed thatc-JHl and C-JH2 could as-sociate with eachother,
and that C-JH2 could betyrosine-phos-phorylated by
c-JAKlorc-JH(l
+2).
Our resultsmayprovide
cluesconcerning
thefunction of the JH2 domain.Studies
by
others showed thatexpression
ofmostmembers of the mammalian JAK kinasefamily
suchasJAK1, JAK2,
andTYK2,
isubiquitous
whereasonly
JAK3 ismainly expressed
inhematopoietic
cells(Ihle
andKerr,
1995).
As afirststep
toexplore
the molecular basisunderlying
theregulation
of carp JAKl geneexpression,
we also cloned and characterized the carp JAKl gene.We havemapped
the carp JAKltranscription
start site and characterized itspromoter.
With nested deletionmutantsof the
5'-flanking
region
fusedtothechloramphenicol
acetyltransferase
(CAT)
reporter gene, the promoteractivity
was characterized
by
transfection into afish cell line. To ourknowledge,
this is the firstreport
tocharacterize thegenomic
structure and the
promoter
region
of JAK kinases.MATERIALS AND METHODS
Materials
Insect
Spodoptera
frugiperda
(Sf9)
cells were grown in Grace's mediumcontaining
10% fetal calf serum(FCS;
LifeTechnologies,
Inc.,
Gaithersburg,
MD).
Monoclonalanti-phos-photyrosine IgG
clone 4G10 waspurchased
fromUpstate
Biotechnology
Inc.(Lake
Placid,
NY).
Construction
of
arandom-primed
cDNAlibrary
Total RNAwasisolated from carp brainby
theguanidinium
isothiocyanate
method(Chomczynski
andSacchi,
1987),
fol-lowedby
the use of aQuickTrack
mRNA isolation kit(Stratagene,
LaJolla,
CA).
The isolated mRNA was used for thesynthesis
ofrandom-primed
cDNA.Amplification of
DNAfragments
by
PCRDegenerate
primers
(Walks, 1989)
weredesigned
tofit the amino acid sequences thatare conserved among PTKs(Hanks
etal,
1988; Hanks,
1991).
The amino acid sequences of thetwo
opposing primers
are HRDLAAR(subdomain
VI inFig.
2,
below)
and DVWSFG(subdomain IX).
The first-strand carp brain cDNA was usedastemplate.
The conditions for the re-action were94°C for 1min,
45°C for 1min,
and 72°C for 2 min in 30cycles.
cDNA
library screening
An
oligo-(dT)-primed
carp liver cDNAlibrary
in the lamda ZAP II(Short
etal,
1988)
obtained fromStratagene
was screenedby using
aPCR-amplified
DNAfragment
as aprobe
(see Results).
Theprobe
was labeledusing
a DIG DNALabeling
Kit(Boehringer
Mannheim, Mannheim,
Germany).
Hybridization
andwashing
werecarried outaccording
tothe standardprocedures
(Sambrook
etal,
1989).
Signals
were de-tectedusing
the DIG Luminescent Detection Kit for Nucleic Acids(Boehringer Mannheim).
Putativepositive plaques
werepurified
through
several rounds ofrescreening
atlower densi-ties and then excisedaspBluescript plasmids according
tothe manufacturer's instructions. To isolate thefull-length
cDNA,
FISH TYROSINE KINASE
JAKl
the
300-bp
DNAfragment
from the5' end ofpJ21 (Fig.
1)
wasamplified
by
PCR, DIG-labeled,
and then usedas aprobe
toscreenanother
random-primed
carp brain cDNAlibrary
as de-scribed above.Sequence
analysis
cDNA clonesweresubclonedinto
plasmid pUC18
(Yanisch-Perron etal,
1985)
andsequenced by
thedideoxy
chain-terminationmethod
(Sanger
etal,
1977)
using
aSequenase
kit(United
States BiochemicalCorp.
Cleveland,
OH).
Several pro-grams from IntelliGenetics(Mountain
View,
CA)
wereusedtoanalyze
the nucleotide sequences.DNA
constructionsDNA
fragments encoding
the entire carp JAKl were ampli-fied withoverlap
extension PCR(Ho
etal,
1989)
fromJ9 and J21 cDNA(Fig.
1)
by using
thefollowing primers: primer
F1.5'-ATATCAGGATCCATGCCAGAACTAGCAGTCATG-3' andprimer
Rl,
5'-GTGGAAACTCAGCTGGCTGAC-3';
primer
F2,
5'-GTCAGCCAGCTGAGTTTCCAC-3'andprimer
R2,
5'-CAGTAC
AAGÇTTCTC
ATCTTATTTCCATAGTTA-3'. The nucleotide sequences of
primers
Rl and F2 arecom-plementary
andthey
are locatedatthe end of clone J9. OtherDNA
fragments encoding
the kinase-likedomain,
kinasedo-main,
and both domainswereamplified
with the PCR from J21 cDNAby
using
thefollowing primers:
c-JH2(residues
572-875),
5'
-ATATCAGGATCCCAGCTGAGTTTCC
ACC-GCATC3'(F3)
and5'-CAGTACAAGCTTGAGGAACCTC-TTCTCAAACAC-3'(R3);
c-JHl(residues 869-1,156),
5'-ATATC
AGGA1ÇCGTGTTTG
AGAAGAGGTTCCTC-3'(F4)
and 5'-CAGTAC
AAGÇTTCTC
ATCTTATTTCCATAGTTA-3'(R2); c-JH(l
+2) (residues 572-1,156),
5'-ATATCA-GGATCCCAGCTGAGTTTCCACCGCATC-3'
(F3) and
5'-CAGTACAAGCTTCTCATCTTATTTCCATAGTTA-3'(R2).
Each
primer
encodesaunique
restrictionsite(underline) (Bam
HI, GGATCC;
and HindIII,
AAGCTT).
The PCRproducts
bases o carp JAKl
s'-f
—n 3500 3730K
J9 J21 HIFIG.1. Restriction maps of carp JAKl cDNA clones. The ki-nase-like
(JH2)
and kinase(JH1)
domains arerepresented
by
thehorizontally striped
and blackboxes,
respectively.
Restriction enzyme sites are shown above carp JAKl cDNA. Thepoly
(A)
tail isrepresented by
AAA.The cDNAclones,
Jl 1 andJ21,
were isolated from acarp liverlibrary
whereas the clone J9 wasisolated fromacarp brainlibrary.
werethen restricted with Bam HI and HindIIIand
ligated
intopQE30
(QIAGEN
Inc., Chatsworth,
CA).
DNAsequenceanaly-sis was
performed
to confirm the accuracy of the PCR prod-ucts.Expression
inEscherichia coli
All the
resulting plasmids (pQEJAKl, pQEJH(l
+2),
pQEJHl,
andpQEJH2)
were transformed into E. coli strain JM109. Thetransformantsweregrownovernight
at37°C in LB broth and diluted 1:10 into fresh medium. After incubation at37°C for1
hr,
isopropyl-/3-D-thiogalactopyranoside
(IPTG)
was added to a final concentration of 1mM,
and incubation was continued for 3 hr. Cellswerecollectedby centrifugation.
Duetoall the recombinant
proteins
containastretchof sixhistidines,
therefore theseproteins
canbepurified by using
Ni2+-nitrilo-triacetate-agarose (Ni-agarose) (QIAGEN
Inc.)
according
totheprocedures
asdescribed(Huang
etal,
1995).
Antibodies
Purifiedrecombinant
protein
c-JHl orc-JH2wasdissolved inphosphate
buffersaline(PBS)
and mixedthoroughly
withanequal
volume of Freund'scomplete adjuvant
for the firstin-jection
of Freund'sincomplete
adjuvant
for the second and thirdinjection. Approximately
100 pg of recombinantprotein,
for eachinjection,
wassubcutaneously injected
into the back ofguinea pig.
Expression
inthe
baculovirussystem
All of the
pQE30
constructs described in the E. coliex-pression
systemweredigested
with EcoRIand XbaIand thenligated
into the transfer vector PVL1393(PharMingen,
SanDiego,
CA).
Plasmids(0.5
pg ofeach)
were co-transfected into Sf9 cells with 0.1 pg of"Baculogold"
virus DNA(PharMingen),
whichcontainedalethaldeletion.Theparental
baculovirus is unabletoinfect insect cells andonly
recombi-nant baculoviruscontaining
theplasmid
constructcanrepli-catein Sf9 cells.
Sf9 cells were infected with various baculovirusat a
multi-plicity
of infection(moi)
of 10plaque-forming
units/cell.
After 60hr,
cells wereharvested, washed,
andlysed
inabuffercon-taining
150 mMNaCl,
1% TritonX-100,
and50 mM TrispH
7.5. The Triton-insoluble cellpellets
weresolubilized inabuffercontaining
1.5%/V-lauroylsarcosine,
25 mMtriethanolamine,
and 1 mM EDTApH
8.0(Frankel
etal,
1991).
Both Triton-soluble and -insoluble fractions were incubated with theNi-agarose.
After extensive
washing
with 0.5 MNaCl,
proteins
wereelutedby
0.1 M EDTApH
8.0. Anequivalent
amountofprotein
wassubjected
toNaDodS04-polyacrylamide
gel electrophoresis
(PAGE),
blottedtonitrocellulose paper, andprobed
with differ-entantibodies.Western
blot
The methodofTricine
NaDodSGj-PAGE (Schagger
andvonJagow, 1987)
wasused. Thegel
concentrationwas7.5%. AfterNaDodS04-PAGE,
theproteins
wereanalyzed
by
immunoblotusing
anti-PYmAborpolyclonal
antibodiesspecific
for JH1 or JH2 domainaccording
totheprocedures
asdescribed(Huang
etImmunoprecipitation
Sf9 cellswerecoinfected with vAcJH2 and vAcJHl. After 60
hr,
cellswerecollectedand solubilizedby
resuspension
and incubation in theimmunoprecipitation
buffer(50
mM TrispH
7.4,
150 mMNaCl,
1 mMNa3V04
and 1%NP-40).
Immunoprecipitation
wasperformed
by
the addition ofanti-JHl oranti-JH2 antiserumatadilution of 1:100orwithpreimmune
serum as a control reaction. The mixture(200
pi)
was incu-batedat4°C for 2 hrfollowedby
anincubation with 50pi
of 50mg/ml slurry
of ProteinA-Sepharose
(Pharmacia,
Uppsala,
Sweden)
for 1 hr. TheSepharose
was collected and washed three times with theimmunoprecipitation
buffer andtwotimes with 20 mM TrispH
7.4,
1 mMEDTA/1
mMEDTA.Samples
werethen boiled ina0.5%NaDodS04
sample
buffertodisrupt
thecomplexes.
Productsweresubjected
toNaDodS04-PAGE
and
analyzed by
immunoblotusing
anti-PY mAborpolyclonal
antibodiesspecific
forthe JH1 orJH2 domain.Reverse
transcription
andPCR
mRNA
(0.5 pg)
isolated from carp brain and liver tissues wasincubatedat65°C for 5 min inabuffercontaining
50 mMTris-HCl,
75 mMKC1,
3 mMMgCl2,
10 mMdithiothreitol,
2 units of RNasin and 1.25 mMdeoxynucleotide
triphosphatates
(dGTP,
dATP, dTTP,
anddCTP),
and thenkept
onice. Two hundred units ofSuperscript Moloney
murine leukemia virus reversetranscriptase
(RT;
GIBCOBRL,
Gaithersburg,
MD)
andoligo(dT)i2_i8 primer
(2 pg)
and random hexamerprimers
(2
pg)
wereadded and incubatedat 37°C for 1 hr. Thereaction was thenstopped by
incubationat95°C for 5 min.Fivesets of
specific primers,
Fl/Rl, F1/R2, F3/R3, F4/R2,
and
F3/R2
wereused and their sequencesweredescribed in the DNA Construction section. PCRwasthen carriedoutby
addi-tion of the heat-treated RTmixture,
PCRbuffer,
andTaq
poly-merase(Promega,
Madison, WI).
Only
theamplified
DNAfrag-ments
by
F3/R3
andF4/R2
from brain tissues and PCRproduct
by
Fl/Rl
from liver tissueswereisolated fromanagarosegel
and subcloned into thepGEM-T
vector(Promega).
Otherana-lytical
methods were the same as described in the section ofSequence Analysis.
Isolation
of
carpJAKl
genomic
clones
A
commercially
available carp liver lambdaFIX IIgenomic
library (Stratagene)
was usedto isolate 15-to 18-kbgenomic
DNAclones
containing
the gene that encodes carp JAKl.Full-length
carp JAKl cDNA was labeledusing
a DIG DNALabeling
Kit(Boehringer
Mannheim).
Approximately
1 X106
independent
cloneswereplated
atadensity
of 5 X104
plaque
forming
units/150-mm
Petri dish.Hybridization
andwashing
werecarriedoutasdescribed above.Signals
weredetectedus-ing
the DIG Luminescent Detection Kit for Nucleic Acids(Boehringer
Mannheim).
Putativepositive plaques
werefurtherpurified through
several rounds ofrescreening
atlower densi-ties. Four distinct carpgenomic
DNAclones(designated
Jl, J2,
J3,
andJ4)
wereisolated,
and DNAwasprepared
from each clone. These carpgenomic
clones weredigested
with various restriction endonucleases. The DNAfragments
were fraction-atedby electrophoresis
in 1% agarosegels
and transferredtonylon
filters(Sartorius
AG,
Goettingen, Germany).
Theresult-ing
DNAblotswereprobed
under theabove conditions ofhigh
stringency
with the DIG-labeled whole carp JAKl cDNA.Sequence analysis
of
genomic
clones that encode
carp JAKlPhage
DNA wasdigested
with NotI,
SacI, 5a/1,
and XbaI and subcloned into either the
pUC18
orpBluescript
vector.Each subclone was
sequenced by
thedideoxy
chain-termina-tion method(Sanger
etal,
1977)
using
theSequenase
proto-cols from United States Biochemical. Thenucleotide sequences of each DNA
fragment
were determinedusing
T3 and T7primers
from both ends. Additional nucleotide sequenceswere determinedusing
different sets of 20-nucleotideprimers
syn-thesized basedonthe known sequences from both ends of each DNAfragment.
The sizes of the intronsweredetermined eitherby
restriction enzymemapping
ofgenomic
clones orby
PCRusing
sense and antisensespecific
primers
that are from twodifferentexons.The
genomic
DNAsequencewasanalyzed
with the GeneticsComputer Group
software program. Thetran-scription
factorrecognition
site data bases(releases
7.3 and6.5)
wereusedto
identify potential transcription
factor motifs within the5'-flanking region
of the carp JAKl gene.Primer extension
analysis
An antisense
oligonucleotide
(5'-GlTT
AATTTCGATCAT-TACAGACGTTTG-3')
complementary
to the 5' end of carp JAKl cDNA clone J9(Fig.
1)
was labeledatthe 5'end with["y-32P]ATP
by
T4polynucleotide
kinase andpurified
on aStratagene
NucTrap probe purification
column. The labeledprimer
was annealedto5pg ofpoly(A)+RNA
prepared
from adult carp liver totalRNAand extendes withMoloney
murine leukemia virus(M-MLV)
reversetranscriptase
(GIBCO BRL)
asdescribedpreviously
(Sambrook
etal,
1989).
The extendedproducts
wereanalyzed
on a5%polyacrylamide,
7Murease-quencing
gel.
The size of the extendedproducts
was inferred fromasequencing
ladder of the JAKl gene obtained from the sameprimer
used forprimer
extension.Plasmid construction
for
CATfunctional
analysis
A 3-kbSal I
fragment encompassing
for5'-flanking
region
was subcloned intopBluescript
and various deletion mutantswere
generated
from this cloneby
PCRusing specific primer
FIG. 2. Amino acid sequence
alignment
of the carp,mouse(Yang
etal,
1993)
and human(Wilks
etal,
1991)
JAKlproteins.
Alignments
wereinitially
madeby
computeranalysis
andweresubsequently aligned by inspection. Gaps
areintroducedtoop-timize
alignment
and shownas dots. The identical residuesarerepresented by
dashes. Theputative
kinase domains are delin-eated witharrows. Subdomains Ia-XIa of the kinase-line domain and subdomains I-XI of the kinase domain aredenotedac-cording
to theprevious
reports
(Hanks
etal, 1988; Hanks,
1991).
The sequence of carp JAKl has beendeposited
in theJAKl
cJAKl MPELAVMELGRQLCGKMKKQRKAEMTVLTVMKGLEIHFYLPDTHQLEYFKDCHTAEDL 58 mJAKl MQYLNIKEDCNAMAF-A--RSFK-T-VKQWPEP-V-VT-L-REP-RLGSGEY-E- 60 hJAKl MQYLNIKEDCNAMAF-A--RSSK-T-VNLEAPEP-V-VI-S-REP-RLGSGEY-E- 6 0 cJAKl CVEGCQEDATSHLCADNLFALSEESQDLWYAPNHAFKITEETSIKLHYRMRFYFTNWHAP 118 mJAKl -IRAA-ECSI-P--H.-YD--TK-RIITVDDK--LR-GT 119 hJAKl -IRAA-ACRI -P- -H.-YD-NTK-RTITVDDKM-LR-GT 119 cJAKl VRTESPVWRHSLFKHKGVSVSPKGPEGTPLLDAASLEYLFAQGQYDFLRGLAPVRAPQNE 178 mJAKl NDN-QS-PK-Q-NGYEKKRV-EA-S-LIKC-I-D-KT- 17 9 hJAKl NDN-QS-PK-Q-NGYEKKKI-DA-S-LVKC-I-D-KT- 179 cJAKl AEKHEIENECLGMAVLAITHHAKSNDLPLSGVGAETSYKRFIPDSLNRTIKQRNFSHVYV 23 8 mJAKl QDG-D-S-MMKKMQ-PELPKDI-Y--ET--KS-R-LLTRMR 23 9hJAKl QDG-D-S-MMKKMQ-PELPKDI-Y--ET- -KS-R-LLTRMR 23 9
cJAKl YNNVFKNFLNEFNSKTIQDSNITLYDLKVKYLSTLETLTQGLGRETIEPKILKVSGESDG 2 98
mJAKl I-D--K-N-C--SVSTH-A-KHY-A-IF-TSM-LI-S-NEL 2 99
hJAKl I-D--K-N-C--SVSTH-A-KHY-A-IF-TSM-LI -S-NEM 299
cJAKl SPALTLPSGDDG.LGYEVQVSGTTGISWRRKPVPNILIVKDKTKSKKNKADKQSKKEMTK 357
mJAKl -RCHSND--NVL.YEVM-TGNLGIQWRQKPNV-PVEKEKNKLKRK-LEYNKHKKDD-RN- 358
hJAKl NWFHSNDG-NVLYYEVM-TGNLGIQWRHKPNV-SVEKEKNKLKRK-LEYNKDKKDE-KN- 3 5 9
cJAKl RKTVMTIFSDFFEITHIVIKESCATIYSQDNKTMELDLFYRDAALSFAALVDGYFRLTVD 417
mJAKl LREEWNN--Y-P-WS-NK--N-N-K-SSHEE-VS-A- 418
hJAKl IREEWNN--Y-P-WS-NK--K-N-K-SSHEE-VS-A- 419 cJAKl AHHYLCTEVAPSSWQNLENGCHGPICTEYAIHKLRQEGNEEGTYVLRWSCTDYNYIIMT 4 77 mJAKl -D-PLI-H-IQ-N-S-M-FDN-L-- 4 78 hJAKl -D-PLI-H-IQ-N-S-M-FDN-L-- 4 79 CJAKl WCIEMDLCESRPVPQYKNFQIETSPQGYRLYGTDTFRPTLKELLEHLQGQNLRTENLRF 53 7 mJAKl -T-F-KSE.VLGGQK-F-VQKGR-S-H-SMDHF-S-RD-MN--KK-I-D-IS- 53 7 hJAKl -T-F-KSEQVQGAQK-F-VQKGR-S-H-S-RSF-S-GD-MS--KK-I-D-IS- 53 9 »*** ja **** CJAKl QPVLVGLGQPRKISNLLVMTRDREPDSQRQPQVSQLSFHRILKEEIVQGEHLGRGTRTNI 5 97
mJAKl VLKRCCQPK--E-A-KKAQEW.-PVYSM-D-KDLI-H- 596
hJAKl MLKRCCQPK- -E-A-KKAQEW
.
-PVYPM-D-KDLI-H- 598
**IIa** *** Ilia *** ***j_va***
cJAKl YSGVLKLKSEDDDDMGGYSQEVKVILKVLGSGHRDISLAFFETASMMRQISHKHIALLYG 6 57 mJAKl -T-LDYKDEEGIAEEKK. . I-DPS-A-V-VY- 6 54 hJAKl -Y-MDYKD-EGTSEEKK. . I-DPS-A-V-VY- 6 56 ******Va****** *****yj;a*****+ cJAKl VCVRHQENIMVEEFVQYGPLDLFMRRQSIPLSTAWKFQVAKQLAGALSYLEDKKLVHGNV 717 mJAKl -DV-EG-H-K-DA-T-P-K-S-D- 714 hJAKl -DV-EG-H-K-DV-T-P-K-S-D- 716 ++**Viia**** * *VIIIa** *** cJAKl CSKNILVARDGLDGEGGPFIKLSDPGIPITVLSREECVDRLPWIAPECVQDTANLSIAAD 777
mJAKl -T--L-L--E-I-SDI-VS--T-Q--IE-I-E-SK-V- 774
hJAKl -T--L-L--E-I-S-C-IT-Q--IE-I-E-SK---V--- 776 *j_Xa**** ***** xa *** ***xia*** cJAKl KWGFGTTLWEICYNGEIPLKDKKLTEKERFYAAQCQLASPDCEELAKLMTHCMTYDPRQR 837 mJAKl --S-T-I-ESR-RPVT-S-K-D-R--N-N-- 834 hJAKl --S-T-I-ESR-RPVT-S-K-D-R--N-N-- 836 < i i to ********** j **** CJAKl LFFRAIVRDIDMVEKQNPSIQP...VPMLEVDPTVFEKRFLKKIRDLGEGHFGKVELCRY 8 94 mJAKl P-M-NKL-E-D-VS.EKQPTT-H-R- 8 93 hJAKl P-M-NKL-E-D-VSRKKNQPT-H-R- 896
** ***ij**** **III* * **IV**** ***
cJAKl DPRGDRTGELVAVKSLKPENREEQSSNLWREIHILRELYHENIVKYKGIWHEEGGRSIKL 954 mJAKl --E--N-Q-SGGNHIAD-KK--E-N-CM-D--NG- 953 hJAKl --E--N-Q-SGGNHIAD-KK--E-N-CM-D--NG- 956 ***** v ******* *** * ** VI ********* CJAKl IMEFLPAGSLKEYLPRNKAHIDLKTLLNYAVQICQGMDLLASRNYIHRDLAARNVLVENE 1014 mJAKl -S-K--NK-N--QQ-K--I-K-Y-G--Q-V-S- 1013 hJAKl -S-K--NK-N--QQ-K--V-K-Y-G--Q-V-S- 1016 ***YII**** ***YIII***** ***jx***** CJAKl NTVKIGDFGLTKSIKDNEGYYTVKDDLDSPVFWYAPECLIHCKFYRASDVWSFGVTMYEL 1074 mJAKl HQ-A-ETDKE-R-Q-I-LH-- 1073 hJAKl HQAETDKERMQSILH -1076 * *x**** **xi** CJAKl LTYCDISCSPMSVFLTMIGPTHGQMTVTRLVKVLEEGKRLPKPDGCSDRLYCLMRRCWEA 1134 mJAKl -SDF-AL--K-T-K-C-PN-P-EV-Q-K-F 113 3 hJAKl -SDS-AL--K--.NT-K-C-PN-P-EV-Q-K-F 113 6 cJAKl TPEKRIDFKGLIANFQQMIDNQ 1156 mJAKl Q-SN-TT-QN--EG-EALLK 1153 hJAKl Q-SN-TS-QN--EG-EALLK 1156
582 JH2 847 875 JH1 1156 c-JAKl
C-JH2
c-JHl
c-JH(l +2)
FIG. 3. Schematic
diagrams
offull-length
and the various truncated forms of carp JAKl constructs. Thekinase-like and kinase domainsarerepresented by horizontally striped
and darkbox,
respectively.
Truncated moleculeswereconstructedas de-scribed in thetext.normalize the
samples
for differencesintransfectionefficiency.
CAT and/3-Gal
activities in the extracts were measuredac-cording
totheprevious procedures
(Herbomel
etal,
1984).
Theacetylated
products
of the CAT assaywereseparated by
thin-layer chromatography developed
with chloroform-methanol(95:5, vol/vol),
visualizedby autoradiography
andquantitated
by using
aPhospholmager (Bio-Imaging
Analyzer
BAS2000,
Fuji, Japan).
RESULTS
Isolation
of
carpJAKl cDNA
We
amplified
carp brain cDNAby
PCRusing
degenerate
primers,
anapproach
showntobeeffective inisolating
human and murine JAKl gene(Wilks
etal, 1991;
Yang
etal,
1993).
sets. PCR
products
were then cloned intopolylinker regions
of thereporter
vectorpCAT-Basic (Promega).
ClonepJPl
contains theflanking region
fragment
JP1,
nucleotides—2,541
to +59. JP1 wassynthesized using
PCRprimers
5'-TTG
AAGÇTTTCCTCCTAGG
ATC AGG-C AG A-3'(nucleotides
-2,541
to-2,522)
and 5'-GGGTCTAGAT-GCTTCAGTCGT-CATGATCAA-3'(nucleotides
+39 to+59)
onthe 3-kb Sal Ifragment.
Theoligonucleotide primers
have additional sequences of Hind III and Xba I sites ateachend,
respectively.
Fragment
JP1 waspurified
from thegel,
di-gested
with Hind III and XbaI,
and then cloned intothesame sites ofpCAT-Basic.
ClonespJP2, pJP3, pJP4, pJP5,
andpJP6
weresimilarly
constructed, except
that the JP1fragment
wasreplaced
with JP2(nucleotides
-2,541
to-181),
JP3(nu-cleotides
-2,541
to-901),
JP4(nucleotides -1,023
to+59),
JP5
(nucleotides
-528to+59),
and JP6(nucleotides
-181 to+59),
respectively.
ThepRSV-CAT
(Gorman
etal,
1983)
was usedas apositive
control.Transfection,
CAT,
and
ß-galactosidase
assayCarp
finepitheloid
cells,
CF(Chen
andKou,
1986),
were maintained in Leibovitz's L-15 mediumsupplemented
with 10% fetal calfserum(FCS)
at27°C. Subconfluent cultures(ap-proximately
60%confluent,
24 hr afterplating)
in60-mmcul-turedishes werewashed twice with Leibovitz's L-15
medium,
and incubated with
DNA-Lipofectamine complexes containing
20 pg of the different CATconstructstogether
with 5 pg ofpSV-/3-galactosidase (ß-GAP)
vector(Promega)
induplicate.
Transfection was carried out for 5hr,
and cellswere washed with fresh Leibovitz's L-15 medium and fed with the same mediumsupplemented
with 10% FCS. After 40hr,
cellswereharvested,
washed inphosphate-buffered
saline(PBS),
andlysed
with 25 mMTrisphosphate pH
7.8,
containing
2 mMdithiothreitol,
2 mMEDTA,
10%glycerol,
and 1% Triton X-100 at roomtemperature
for30 min. The totallysates
werescraped
fromthe dish and transferredtomicrocentrifuge
tubes. Cell debriswasremovedby centrifugation
and theextractswere frozenat—
70°C untiluse.Protein concentrationwasmeasured
by
the Bio-Radprotein
concentrationquick-assay
method(Bio-Rad, Richmond,
CA).
ThepSV-ß-Gal
vector(Promega)
carry-ing
the SV40promoter
linked tothe/3-Gal
gene wasused to¿w»
kDa M 1 94 75 45 28 22 17ML
— C-JH1/JH2#vvs
M c-JHlB
kDa 94 75 45 28 22 17 anti-PYFIG. 4. Demonstration of
tyrosine
kinaseactivity
of c-JHl in E. coli. Strain JM109 cellsweretransformed withpQE30
(lane
1),
pQEJH2
(lane 2),
andpQEJHl
(lane 3),
respectively.
After induction with 1 mM IPTG for 3hr,
proteins
wereprepared
andseparated by
NaDodS04-PAGE
and stained with Coomassie blue(A)
orimmunoblotted with anti-PY mAb(B).
Positions of c-JHl and c-JH2 are shown on theright.
LaneM,
Prestained molecularmassmarkers.FISH TYROSINE KINASE
JAKl
Amplified fragments
ofabout220bp
werepurified
andligated
intopGEM-T (Promega). Among
the 20clonessequenced,
15 clones contained asingle
JAK-related sequence.Using
this DNAfragment
as aprobe,
we screened anoligo(dT)-primed
carp liver cDNAlibrary.
Schematics of thetworepresentative
carp liver JAKl clonesobtainedfrom thisscreen areshown in
Fig.
1. On the basis ofDNAsequenceanalysis,
thesetwoclones,
pJl
1 andpJ21,
areidenticalexceptthatpJl
1 is0.7 kb whereaspJ21
is 2.1 kb inlength.
It was found thatpJ21
encodes the carphomolog
of mammalianJAKl,
containing
residues from465 to
1,156.
To obtain thefull-length
cDNAclone,
a300-bp
DNAfragment
from the 5' end ofpJ21
wasusedas aprobe
toscreen acarp brain cDNA
library.
Thisscreening yielded pJ9
(1.9
kb),
asshown inFig.
1.Therefore,
thefull-length
carpJAKl cDNA sequencesarederived from the two
overlapping
clones,
pJ9
andpJ21,
and containa5'-untranslatedregion
of 186nucleotides,
acoding
region
of3,468 nucleotides,
anda3'-untranslated
region
of65nucleotides. The totallength
of carp JAKl cDNA is3,719
nucleotides.(The
sequence has beende-posited
in GenBank withanaccession numberL24895.)
Structure and the
activity
of
carpJAKl cDNA
The
complete
sequenceof carp JAKl cDNA encodesanopenreading
frame of1,156
amino acid residues withapredicted
mol-ecularmassof 129 kD and theprotein
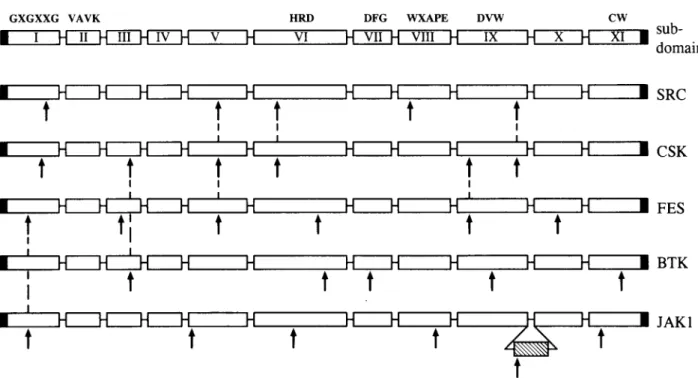
wastermed carp JAKl.Allmembers of the JAK
family
havesevenhomologous
domains in the molecule that have been namedasJHsorJAKhomology
domains. JH1 is acarboxy-terminal kinase-catalytic
domain whereas JH2 isakinase-likedomain;
the other five JHsarepre-sentin the farammo-terminal
part.
Aminoacidsequencecom-parison
of carp JAKl with human and murine JAKl(Wilks
etal, 1991;
Yang
etal,
1993)
is shown inFig.
2. There isahigher
sequencehomology
in both JH1 and JH2(70%
identity).
The overall sequenceidentity
between carp andhuman/murine
JAKl is about 57%. When the deduced amino acid sequences of carp JAKl werecompared
with those of murine JAK2(Harpur
etal,
1992),
murine JAK3(Witthuhn
etal,
1994),
and human TYK2(Firmbach-Kraft
etal,
1990),
itwasfound that the sequenceho-mology
is lower in JH1(50,
46,
and 56%identity, respectively)
andJH2
(45,
43,
and51%identity, respectively).
The overall se-quenceidentity
between carp JAKl and murineJAK2,
murineJAK3,
and humanTYK2is35%, 31%,
and42%,
respectively.
Totestwhether the JH1orJH2 domains possess
kinase-cat-alytic activity,
wegenerated His-tag
fusionproteins
of the JH2 domain(c-JH2)
and theJH1 domain(c-JHl)
ofcarp JAKl(Fig.
3),
andexpressed
thesefusionproteins
in E.coli(Fig.
4A).
Thetyrosine
kinaseactivity resulting
from each fusionprotein
can be detectedby anti-phosphotyrosine
monoclonal antibodies(anti-PY mAb).
Duetothe lack ofendogenous tyrosine
kinases in E.coli,
there is littleornocross-reactivebackground
for the anti-PY Western blot(Fig.
4B,
lane1).
Among
theexpressed
fusionproteins (Fig.
4A),
only
c-JHldisplayed tyrosine
kinaseactivity
and severaltyrosine-phosphorylated
bandsincluding
aband
corresponding
toc-JHl weredetected(Fig.
4B,
lane3).
By
contrast, C-JH2(lane
2)
didnotexpress any observablety-rosinekinase
activity.
These resultsareconsistentwithanear-lier
report
(Wilks
etal, 1991)
andsuggestthatonly
JH1 isafunctionally
active kinase domain. c-JHl and C-JH2were fur-therpurified by Ni-agarose affinity chromatography
and usedtogenerate
polyclonal
antibodies(see
below).
Full-length
and varioustruncated
forms of
carpJAKl
areproduced
in insectcells
using
the
baculovirus
system
Figure
5,
A andB,
show thatc-JAKl,
c-JH(l
+2),
c-JHl,
and C-JH2were
present
inboth the soluble andTriton-A kDa 94 75 45 . 28 22 . 17 B kDa
VN^?V
„(*»»
sS» vN°'JB^WW*1
10„KcVO^
( anti-JH2 S&„ce«»
3ft0+ï>
..mJ**1
-« c-JAKl ., c-JHO+2) -« pp60 -. c-JHl c kDa anti-JHl vf* 10 — c-JAKl — c-JH(l+2) -. pp60 — c-JHl anti-PYFIG.5.
Expression
offull-length
and various truncated formsofcarp JAKl in thebaculovirus system.Sf9 cellswereinfected
with
wild-type
baculovirus,
AcMNPV(lanes
1 and2)
and avariety
of recombinantbaculovirus(lanes 3-10),
respectively.
After 60hr,
cells wereharvested andseparated
into Triton-soluble(lanes
1, 3, 5, 7,
and9)
andTriton-insoluble fractions(lanes
2, 4, 6, 8,
and10).
Both fractionswerefurther boundtoNi-agarose gels
asdescribedin thetextandequivalent
amountsofbound
patients
wereseparated
by NaDodS04-PAGE,
trans-ferredto
nitrocellulose,
and immunoblotted withpolyclonal
an-tibodies
specific
for JH2 domain(anti-JH2;
A),
for JH1 domain(anti-JHl; B),
and a mAb forphosphotyrosine
(anti-PY; C).
Positions of
prestained
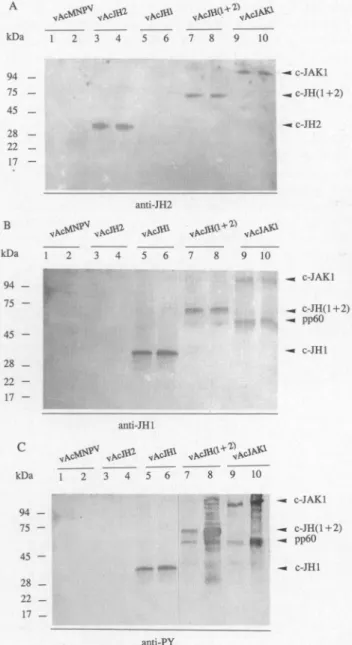
molecularmassstandardsand theirinsoluble fractionsas revealed
by
antibodiesspecific
for JH2(Fig.
5A)
and for JH1(Fig.
5B).
Again,
the anti-PYmAbswere usedtodetect thetyrosine
kinaseactivity
of eachfusionpro-tein. As a
control,
Sf9 cells were infectedby
wild-type
bac-ulovirus and there was little orno cross-reactivebackground
for the anti-PY Western blot(Fig.
5C,
lanes1 and2).
As shown inFig.
5C(lanes
3 and4),
c-JH2 didnotdisplay tyrosine
ki-naseactivity.
Other fusionproteins,
c-JAKl,
c-JH(l
+2),
and c-JHl allexpressed tyrosine
kinaseactivity (Fig.
5C,
lanes5-10)
andautophosphorylation by
themselves seemedtooccur as wecompared
theirelectrophoretic
mobilities inFig.
5,
A andB,
and thecorresponding
mobilities in the anti-PY Western blot(Fig.
5C).
Interestingly,
in cellsexpressing
c-JAKl andc-JH(1
+2),
anadditionalprotein
of60 kDwasdetectedby
anti-JHl antibodies and anti-PY mAb(Fig.
5B,
lanes7-10,
andFig.
5C,
lanes7-10).
Moreover,
there were moretyrosine-phos-phorylated proteins
bound toNi-agarose
in the Triton-insolu-ble fraction of insect cellsexpressing
c-JAKl andc-JH(l
+2)
(Fig.
5C,
lanes 8 and10)
than cellsexpressing
c-JHl. The pro-teinsanalyzed
werefractions boundtoNi-agarose, presumably
viatheHis-tag
of recombinantproteins.
Therefore,
they
were eithercleavage products
of JAKl relatedproteins
orassociatedproteins.
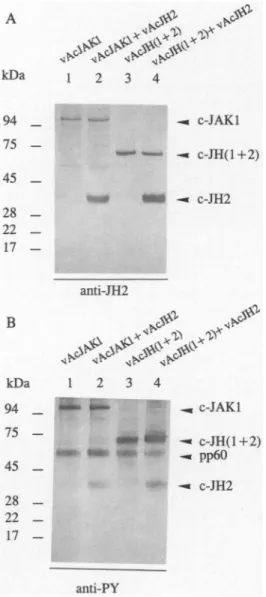
c-JHl and C-JH2
interactwith each other
To demonstrate the association of c-JHl and
c-JH2,
anti-JHlantibody
was used toprecipitate
c-JHl from the extract of vAcJHl- and vAcJH2-co-infectedcells,
and the associated c-JH2wasdetectedby blotting
withanti-JH2antibody.
As shown inFig.
6A,
C-JH2 isco-precipitated by
c-JHl.Similarly,
c-JHlis also
co-precipitated
by
c-JH2(Fig.
6B).
Because bothpro-teins are
highly expressed
under theseconditions,
there is thepossibility
that this interaction is somehownonspecific.
Therefore,
another recombinant baculovirusvAcCAT,
which carries thechloramphenicol acetyltransferase
(CAT)
gene un-der the control of the samepolyhderin
promoter,
was usedto coinfect with vAcJHl or vAcJH2. Theimmunoprecipitation
data showed that neither anti-JHl noranti-JH2antibody
was abletoprecipitate
CATprotein
from thecoinfected cellextract(data
notshown).
Moreover,
neither anti-JH2noranti-JHlan-tibody
wasabletoprecipitate
c-JHlorC-JH2 from theextractsfrom cellsinfectedwith vAcJHl orvAcJH2 aloneasshown in
Fig.
6(lane 1).
Therefore,
C-JH2seemstointeractspecifically
with c-JHl andpossibly by
tyrosine-phosphorylated by
c-JHl(see
textsbelow).
JH2 domain
istyrosine-phosphorylated
by
c-JAKl and
c-JH(l +2)
Because c-JHl and C-JH2were associated with each other
(Fig.
6),
wesought
toinvestigate
whether c-JH2 isasubstrate forc-JH1by
co-infectionexperiments
in which insect cellswere co-infected with vAcJH2 and vAcJAKl or vAcJH2 andvAcJH(l
+2),
instead of co-infection with vAcJH2 and vAcJHl. This avoidedambiguity
indatainterpretation
due tothe similar
mobility
of c-JHl andC-JH2onNaDodS04-PAGE.
Theexperimental procedures
werethesame asthose described in the vAcJHl and vAcJH2 co-infectionexperiment.
Proteins boundtoNi-agarose gels
wereanalyzed by blotting
with either anti-JH2antibody
oranti-PY mAb. The controlexperiment
was carried outby infecting
insect cells with eithervAcJAKl or##
B
kDa
94
-75 -45-28
-22 -17-IP
F
& &
1 2 3#^
ir jo w *<§
*Blot
:anti-JH2
kDa
94
-75 -45 -28 -22 -17-IP:
9
Cv ¡y » « SS¡ S
Blot
:anti-JHl
FIG. 6. Association ofc-JHl with C-JH2. Sf9cells wereinfectedwith vAcJH2 alone
(a,
lane1),
vAcJHl alone(B,
lane1),
and co-infected with vAcJHl and vAcJH2
(lanes
2and3),
respectively.
TheTriton-soluble fractionswereimmunoprecipitated
with normalguinea pig
antiserum(pre-imm;
Iane2),
anti-JHl antibodies(A,
lanes 1 and3)
and anti-JH2 antibodies(B,
lanes 1 and3).
The associatedproteins
wereanalyzed by
NaDodS04-PAGE,
transferredtonitrocellulose,
and immunoblotted with anti-JFK(A)
oranti-JHl antibodies(B).
Arrowheadindicatespositions
of c-JHl andC-JH2,
respectively.
JAKl
vAcJH(l
+2)
alone. As shown inFig.
7B,
C-JH2indeedwastyrosine-phosphorylated
by
c-JAKl(lane 2)
andby
c-JH(l
+2)
(lane 4).
This suggests thattransphosphorylation
of C-JH2by
c-JHl mayoccurwhenthey
areexpressed
inhigh
levelsto-gether.
RT-PCR
of
brain and liver mRNAAsdescribed
above,
twooverlapping
clones,
pJ9
andpJ21,
wereisolated from different tissues. Toinvestigate
whethertis-sue-specific
alternativesplicing
occursandtoanalyze
the un-clonedregions
inthetwotissues,
fivesets ofspecific primers
wereusedtoperform
RT-PCRonmRNAderived fromcom-4.4 kb—
2.3 kb
_
2.0kb—
0.6 kb—
*
if
<o
&
& .# %
Q %
&
BLBLBL BL BL kDa 94 _ 75 -45 _ 28 _ 22 _ 17-B
kDa 94 . 75 -45 . 28 -22 -17c#
<^.
VÄ^>*>
MF
»i* tr<fif?
- c-JAKl <—— ^c-JH(l+2)
anti-JH2Hi
&
^c^>:>
4*"
4P
MF
-, c-JAKl • -«c-JH(l+2)
-*pp60
» -« C-JH2 anti-PYFIG. 7.
Transphosphorylation
ofC-JH2by
c-JAKl andc-JH(1
+2).
Sf9 cellswereinfectedwith vAcJAKl alone(lane
1),
both vAcJAKl and vAcJH2(lane
2), vAcJH(l
+2)
alone(lane 3),
and bothvAcJH(l
+2)
and vAcJH2(lane 4),
respec-tively.
The Triton-insoluble fractions wereprepared
and then incubated withNi-agarose
gels
andequivalent
amounts of boundproteins
wereseparated by
NaDodS04-PAGE,
trans-ferredto
nitrocellulose,
andimmunoblottedwith JH2 anti-bodies(A)
or anti-PY mAb(B).
Prestained molecular mass markers(in kilodaltons)
areshownonthe left.FIG. 8. RT-PCR
analysis
ofthe carp JAKl kinasetranscripts.
mRNAs derived from brain(B)
and liver(L)
tissues wereprimed
witholigo(dT)
and randomprimers
andsubjected
to re-versetranscription.
Theresulting
cDNAwasamplified
with fiveprimer
sets,i.e., F3/R3,
R4/R2,
F3/R2,
Fl/Rl,
andF1/R2,
and thePCRproducts
wereanalyzed
by electrophoresis
on a 1% agarosegel.
The sequence of eachprimer
wasdescribed in theDNAConstructionsection.
moncarp brain and liver tissues. PCR
products
from both tis-sues were identical inlength (Fig.
8).
Only
the unclonedre-gions
inthetwotissues,
i.e.,
PCRproducts
fromprimers
F3/R3
and
F4/R2 (from brain),
and the DNAfragment
fromFl/Rl
(from
liver)
weresequenced.
All of the nucleotide sequences werethesame as thosewedeposited
in GenBank withan ac-cession number L24895. These sequences,aswellasthose from twooverlapping
clones,
pJ9
andpJ21,
suggestthatatleasttwoidentical
transcripts
from carp brain and liver tissues encode the samefull-length
carp JAKl kinase.Genomic
organization
of
the
carpJAKl
geneAsaninitial
step
toinvestigate
theregulation
of carp JAKl geneexpression,
we also cloned and characterized the carp JAKlgene. Fourpositive phage
clones,
termed Jl toJ4,
wereisolated from a
Stratagene
carp livergenomic library
with aDIG-labeled
full-length
carp JAKl cDNAas aprobe.
We are unable tofill the gap betweenphage
cloneJl and J2by
PCRamplification
ofgenomic
DNA with a setofoligonucleotides
thatcorrespond
to one end of clone Jl andthe other end of cloneJ2.To locate allexons,thesephage
cloneswereanalyzed
by
Southernblotting, subcloning,
andsequencing.
As shown inFig.
9,
the restriction map of eachgenomic
clone wascon-structed
by digesting
thephage
DNA with apanel
of restric-tion enzymesseparately
orin variouscombinations:5a/1,
Bgl
n,
HindIII,
XhoI,
and Eco RI. On thebasis of the nucleotide sequences of subclonedfragments,
the carp JAKl gene iscom-posed
of24exonsthat spans atleast 31 kb ofDNA(Fig.
9).
Thesequencesaroundthe exon-intron boundarieswere deter-mined andareshown in Table 1. Allexon-intron boundaries
identifiedconformedtothe
GT/AG
splice donor/acceptor
rule(Breathnach
etal,
1978).
Some exons wererelatively
small(88-108
bp),
whereas the first and the sixth exons werelarge
(237
bp
and 349bp).
The size ofintronsvariedconsiderably,
ranging
from >3 kb(intron
1)
to 100bp
(intron 19).
The first exoncontainedthe5'untranslatedregion,
andthe secondexonexon untranslatedexon
I_L
J_L
i i2S
o— CJ00 Wffl_u_
> Di o -Ü CO — -, Sx m a_LL
J_L
> oíExon
no l 2 3 4 5 6 7 8 9 10 11 1213 14JAK,
^JIJUL/IIII
IJUJU1JLJUI
15 1617181920 21 22 23 24 2.5kb >3kb 1.8kb lkb 1.2kb 1.5kb 2.3kb 2.2kb 1.5kb 1.8kb 1.5kb 1.4kb
Jl
/A
J2
J3
J4
"//
FIG. 9. Genomic
organization
of the carp JAKl gene. Exons are indicatedby
boxes numbered from 1 to24.Solid boxesin-dicatethe carp JAKl
coding region
whereas open boxesrepresent
the 5' and3' untranslatedregions.
Intronsand the5'-and3'-flanking regions
are indicatedby
the solid lines. The entire gene spans atleast 31 kb inlength
andcontains 24exons. A re-striction mapwasshown above thegenomic
structure.Threeoverlapping phage
clones,
J2-J4 andonenonoverlapping
clone, Jl,
were isolated fromaLambda FIXII carpgenomic library.
Agap betweenphage
clones Jl and J2was notobtainableby
PCRamplification
ofgenomic
DNA.containedthe
putative
translation initiation site. Thelargest
in-tron
(>3 kb)
separates
exons1 and 2. The JH2 domainwas lo-catedon exons 11-17 and thecatalytic
JH1 domainwaslocated on exons 18-24. Exon 24containedthe last 33 amino acidsaswellas the 3' untranslated
region.
Thepromoter
andexon/in-tronsequences of carp JAKl kinase gene have been
deposited
in GenBank with 10 serial accession
numbers,
from U53685to U53694.Table 1. Exon-Intron Organizationof theCarp JakI Gene
exon exon size
number
(bp)
3'end of
5'end of
approximate
3'end of
5'end of
amino
acid
the
exonthe intron
size
(bp)
the intron
the
exoninterrupted
1 242 TGC CTG ACG AG gtaaggacga >3000 tatcctgcag T GTC TGG ATG2 208 CAA GAA GAT GCC gtaagcgaga 1800 ttttccccag ACA TCT CAC Ala67 (3)
3 125 TAC CGT ATG AG gtgagtgggc 1000 gtttacacag G TTT TAT TTT Argl09 (2)
4 154 CTG TTT GCA CAG gtgaggtaca 400 tatgttttag GGC CAG TAT Glnl60 (3)
5 164 GCTGAG ACC AG gtgaggtaca 148 atctttgtag C TAC AAG CGA Ser215 (2)
6 349 AAG CCT GTA CCG gtgagtttag 250 cttttgatag AAT ATT CTG Pro331 (3)
7 180 AAT AAA ACT ATG gtaatttcca 600 ttgcccacag GAG TTG GAC Met391 (3)
8 158 GGA CCT ATC TG gtatgaccat 1200 ttctctacag C ACA GAG TAT Cys444 (2)
9 115 GTC TGC AH GAG gtacacacac 1500 tctctctcag ATG GAC CTA Glu482 (3)
10 199 CAA CCC AGA A gtaccagcac 2300 tttccttcag AA ATT TCC AAC Lys549 (1)
11 110 GAG ATT GTA CAG gtgatatttt 900 ttttattcag GGT GAG CAT Gln585 (3)
12 150 GAT ATC TCT CTG gtaagatgca 187 ctcctctcag GCT TTC TTT Leu635 (3)
13 89 CAT CAG GAG AA gtaagtacct 2200 ctgtctgtag T ATC ATG GTG Asn665 (2)
14 127 CTC AGC TAT CTG gtaaagaaac 1500 aatcctgtag GAG GAC AAG Leu707 (3)
15 136 AGCAGA GAA G gttgagagat 820 gtgtctacag AG TGT GTG GAC Glu753 (1)
16 152 AAGCTC ACA GAG gtaacagcat 111 ctctgagcag AAG GAG AGG Glu803 (3)
17 152 GAG AAA CAG AA gtatgaccta 457 ttctctccag T CCT TCC ATT Asn854 (2)
18 88 GAT CTTGGA GAG gtattttctg 110 ctcttttcag GGT CAC TTT Glu883 (3)
19 193 CAC GAA GAA G gtaaagccac 100 ttgtttcaag GT GGG AGA TCC Gly948 (1)
20 125 CAG ATC TGC CAG gtaacatcat 1800 ttaactccag GGC ATG GAC Gln989 (3)
21 173 CCA GTG TTCTG gtaagaatca 170 attttcccag G TAT GCC CCA Trpl047(2)
22 118 AGC CCT ATG TCG gtaagtggcc 1500 aaccattcag GTG TTC CTT Serl086(3)
23 111 TGC TCA GAC AGG gtaatatata 1400 gttttgacag TTG TAC TGT Argll23(3)
24 164 ATG AH GAC AAT CAG TAACTATGGAAATAAGATGAGATGCAGACTCCACCTCTTTTGTAAGAGGAAGTTCCAGAGAGACCAAAAAAAAAAA
_
FISH TYROSINE KINASE
JAKl
.NF-IL6 . -2541 TCCTCCTACGATCAGGCAGAAAATGTTTTnGACATTTATTTTGACATTATGTTGTTGTTTTGGGAAATAAAAAATCATTAAAATATTCACnnCACTC -2441 ATGTTTTTCCTGTAGACATCACGACTATAAAGAAAATTAAGACATCATTTCAGTTTGCAnTCATTTAATGTTnCTACAATTTCnTTCTTTCTGTGTC -2241 TTTTGGCACAGAAATATCCTTTTCAAATAGACACnGTAATCAAACTTTGCACAGACTTTAGTnCTGTGTTTnCAAAGACTACAAAATGTATATAAGT HNF-5. ... -2141 TAAAACAACAnGTACATCCTTATTTAnATGCATATnGTGTAAATATTAGCAGAAATGGGAAnCCATCCTTnACATATAnAACATTTGnTTAGG -2041 GCGACnACTACAAAAAGATGnTACnATATGAAACAATTTGACAACCAAACMATAGAAGTTTTAAAAAAAAAATAATACCTGCTTTTCTGTTCCCAT -1941 AAAACTGCTGAnTGTGATGTCAGACATnAAAAGCCATGAAGAGAAAGATATCATGGCCAAACCCCAAAACATTTGGAATCTCTGAAAAGCCCAATATG NF-IL6 .... -1741 CAGnCTAGGAGGnAATATAnGAATATCATTTTGTACAATATCAAAGCATATATGATGTATAAATTAGTTACTAAACAnnAnCAATTCATAGCAT -1641 nGGACACACACCATGAAGATnACAGTGTAGTATGATAAGTCnnCTGCCCATnCGTnATATAAnATAGACCATnTCCCATGATGTTAGTAAAT .API -1541 TAGACCCAGGTATTATCCTCTGTCACTTGCCCCnGnACATCAGCCCTnAACTCTTGAGTCATCTGAACATTTTAGAATCATATAGTAAGGTGAGTAA -1441 GTGGTCTGTAATGnGTGATATATGGAAATGCAGnATACAATnATTTTTTCnTTITnCAGAAAATAAAAATGTTTTCTCATCTGAAAAGCTGGGTC NF-IL6 . HNF-5 .... -1341 ATTnCAAnTGATGTTTACATGTATTTCGGAAATCCTCATCTAAATATTTGCTGAATnCTGTCTTAGCTTTTAGATGAGTGTTTATGGCAGTAnGCA . API -1241 CGTAGACTCATCACGGACCAAnATAGCCCATGACTCATCCAACGATAGGAGTTTCCAACTGGACTTTTTCCCCCCAGTGTnGTTTAAATAnAnGTC -1141 AACATATCTCAACACCATATTTGACATAAATATGTAnGTTGCAAATCATTnGACATGTCTGACAACATTTATATGGAAATGTTGATAAGGATAGACAA -1041 AAATGTATTTAATAGTAGAATnGAACTGAACGTTnACTCTAAAAACCTATTAAACTCCAATACACACAGTCTAATAnGTTTATATATCTACCCGAAC -941 nCTGnGAACTTAnGATCAGTCACAnaCCCAATGCTCTGCAGCCCACAGTTTCTGGCAACTCATATGATGCTGAGGACGGAAGTCCCTCCCTTTCA -841 CGGCAGGCnAAACCTCATGTCnAAAGACCATCTGCGTAATCAGAGAGGGGTTTGTGTTnGCTGATACCTATTTTAAAAACAGACACGTACCTGTAAA E2A ... -741 GCGGAAACAGCTGATTGATTMCTTAATGTnAGGAnAGGACTTrATGGACAAAAATAAACAACAGTAATAAAATGTAACAGnCAnAATAATATTTC NF-IL6 . . GHF-5 ... -641 TATTTACATTTTAnTCTGAAATCTAAAnATnATGAnATATCCAnATnATGAATCTGCAAMCATCTTTTACAGTGnGTGCTGTTACTGTTTAT -541 ATAGACTAGATAACTGCAGTATCTAAGCGnACnTTAAATGAGAAGCATAATAATGCnAGnAGCAGTAAnATTTAATATATGAATCAAnATTAAT -441 GGTTGTTATATnAAAGTGGTCATGTGGCATACCAnCAATGGACAAAATTnATCCGAGATATCTTTnAATACTACATATTTAACGAAGTGAAATCAG . TATA-box. -341 CGCnAAGGATCnCATGTAnATGAnCTAATCAnTTCAAACCATTTGAATCAnGGAGAGAGAGAAAAAAAAATGATATGTATAAATATGATGTTiT NF-IL6 ... -241 GGGAAATTTATGCATTTATGTTTACATTACAGCAATGCACTCTAAGCGAGAGTTTTGATCGCATGCATAAATAGTAAnCGTCTTTTAACTATATACAGT -141 ACAGAGGCAATGCGTCCAGTAGCCACAACAAAAAATCGCTGGCTTACTCTCTGCnAAGGGnAAAGCCAnGGTACAGAATTTGAnGACAGCCGCATG . CCAAT ... .+1 .... . -41 AGCCAATAAGCGGCATGnACAGACCTCGTCGnACCCTATTnCGGGAATGTGTTCGACGCTTGACAGGAAAGGTTAGTTGATCATGACGACTGAAGCA.*
( transcription
start site of carp JAKl gene)
.+60 CGGnTGTAGCACTCTGAAGTGAAGTnGTnTGAAAATAAACCTCGTGGAATAAACCAnATACTATCGGACAnAnnGGAGTGAAATAACGAGCAC
+250 +160 TTTTACATCAACAAAAAACCAAGAGGAAGTGTTCAAACGTCTGTAATGATCGAAAnAAACGGAAGTAGCTTTGCCTGACGAGTGTCTGGATGCCAGAAC
(
translation start site)
FIG. 10. Nucleotidesequence of the
5'-flanking region
of the carp JAKl gene. A 28-mer antisenseoligonucleotide
usedforprimer
extensionanalysis
is underlined. The candidatetranscription
startsiteby primer
extension(see
Fig.
11)
is indicated with anucleotidenumber(+1)
andanasterisk,
which is locatedat249bp
upstream
tothetranslationstartsite.Potentialbinding
sites foravariety
oftranscription
factors arealso marked and underlined. Thepromoter
andexon 1 sequences of carp JAKl kinase gene has beendeposited
inGenBank,
accessionnumber U53685.Determination
of
thetranscription
initiation siteThe
transcription
startsitewasdeterminedby
primer
exten-sion
analysis using
poly(A)+RNA
from carp liver. We useda 28-meroligonucleotide
labeled with32P
atthe5' end. Theex-act