國立臺灣大學生命科學院生態學與演化生物學研究所 博士論文
Institute of Ecology and Evolutionary Biology College of Life Science
National Taiwan University Doctoral Dissertation
台灣入侵植物南美豬屎豆及其根瘤菌之共生關係 Symbiotic relationship between an invasive legume, Crotalaria zanzibarica, and its root-nodulating rhizobia
in Taiwan
黃承泰 Cheng-Tai Huang
指導教授:高文媛 博士 劉啟德 博士 Advisors: Wen-Yuan Kao, Ph.D.
Chi-Te Liu, Ph.D.
中華民國 107 年 5 月
May, 2018
致謝
本論文承蒙許多師長、同學、友人及家人的協助得以完成。首先感謝在成功 大學就學時期影響我最深的兩位老師,侯平君老師(已故)啟發我對生態學研究的 熱情,曾怡禎老師教導我微生物技術,為本論文之研究打下基礎。
感謝指導教授高文媛老師,願意收留我這個在離開大學四年後、走投無路的 學生,讓我的研究之路得以延續;雖然碩士班的表現頻頻讓老師失望,但老師還 是很有耐心地指導,也同意讓我挑戰不同的研究領域,更提供我念博士班的機會。
感謝劉啟德老師的共同指導,讓我有發揮微生物專長的舞台,除了專業建議之外,
劉老師還時常給予精神層面上的指引,是我在實驗挫折中的一盞明燈。
感謝王俊能老師、湯森林老師、江殷儒老師及黃政華老師詳細審閱這本論文,
各位老師不吝提供許多寶貴的建議,使得整本論文能更完善。感謝陳香君老師在 電子顯微鏡技術上的協助,讓我得到精美的實驗照片。
感謝好友陳仲麒、曾景漢、王郁盛在烈日下協助我採樣。感謝高老師及劉老 師實驗室的各位學長姐及同學們,這段日子有你們真好。感謝張菽娟小姐細心管 理溫室,讓我的植物頭好壯壯。感謝楊月鈴助教協助行政事務。
最後,感謝我的父母、小狗布魯與優比一直以來的支持與陪伴,讓我能順利 渡過這段艱苦的日子。謹以此論文獻給父母: 黃景潮先生與李繡淳女士。
摘要
南美豬屎豆 (Crotalaria zanzibarica Benth.) 是台灣歸化豆科植物中分布最廣 的一種,為多年生的木本植物,常見於道路旁、河岸及廢耕地。野外觀察發現南 美豬屎豆普遍具有根瘤,因此推測與固氮菌(根瘤菌)共生有助其在貧瘠棲地建立族 群,是讓此植物能夠在台灣廣泛分布的原因之一。然而,南美豬屎豆在台灣的共 生根瘤菌未曾被研究。本論文探討南美豬屎豆與根瘤菌之間的共生關係,以了解 此外來植物在台灣的共生根瘤菌之多樣性及可能來源,並檢驗不同根瘤菌株對植 物的共生表現。
從種植在台大溫室的南美豬屎豆根瘤中分離出具多形態的菌株 CzR2,分析此 菌株之 6 個管家基因 (atpD、dnaK、glnII、gyrB、recA 和 rpoB) 序列,結果顯示 CzR2 屬於 Bradyrhizobium arachidis。CzR2 在游離狀態時受到甘露醇或果糖誘導會 產生多形態細胞,此現象首次在根瘤菌中被發現。CzR2 在與南美豬屎豆共生時也 產生多形態的類菌體,然而有些類菌體其染色體具多倍體,有別於游離細胞的單 倍體及二倍體。此結果顯示 CzR2 與南美豬屎豆共生時其染色體有內複製的現象。
調查新店溪沿岸的南美豬屎豆及常見的六種共域豆科植物,這些植物具有不 同的根瘤(有限及無限)和類菌體(膨大及非膨大)形態。由分離菌株的 16S rRNA 序 列分析,得知南美豬屎豆、蠅翼草、異葉山螞蝗和鍊夾豆皆與 Bradyrhizobium 建 立共生,與田菁共生的根瘤菌為 Neorhizobium 和 Rhizobium,白花菽草則為 Rhizobium,含羞草的共生根瘤菌為 Cupriavidus 和 Paraburkholderia。雖然這些植
物具有不同的共生特徵,其葉片都具有類似的穩定性氮同位素比值(δ15N),其值約
為 -1 ‰;且這些植物根瘤的 δ15N 皆為正值(3.7-7.3 ‰),其中無限根瘤比有限根瘤
普遍具有較高的δ15N 數值。
以多基因序列 (dnaK-glnII-recA-rpoB) 分析從台灣北、中、南三河岸南美豬屎 豆族群所分離出之 59 株根瘤菌株,以及其他共域豆科植物族群所分離出之 54 株 根瘤菌株,結果顯示這些菌株皆屬於 Bradyrhizobium 且可區分成 21 個支序群。同
時比較分析其他共域豆科的根瘤菌群時,發現某些菌群似乎對南美豬屎豆有專一 性,然而某些菌群則廣泛出現在多種植物的根瘤中。多數南美豬屎豆根瘤菌具有 代表美洲起源的 nodA 共生基因,其次為亞洲起源及世界廣布型,顯示此物種在台 灣能夠與來自不同地理區的的根瘤菌建立共生。
藉由溫室實驗比較接種不同菌株對南美豬屎豆的影響,結果發現南美豬屎豆 與不同菌株共生時產生不同的根瘤且類菌體形態亦有所不同,顯示根瘤及類菌體 形態亦受到共生菌株的影響,不全受到宿主決定;南美豬屎豆植株生物量及植株 總氮量在不同菌株接種處裡下有顯著差異,然而,南美豬屎豆植株生物量分配、
氮含量、穩定性同位素比值和共生效率(總生物量變化/總根瘤生物量變化)受不同 接種菌株的影響並不顯著。
綜合野外調查和溫室實驗結果,證實南美豬屎豆能夠與多樣 Bradyrhizobium 菌種建立有效共生,顯示其為廣適性宿主,此特徵有助於該物種擴散到不同地點 時能找到相容根瘤菌。另一方面,來自南美洲的外來根瘤菌對於南美豬屎豆在台 灣與根瘤菌建立共生中扮演重要角色。
關鍵詞: 入侵植物、南美豬屎豆、豆科與根瘤菌共生、慢生型根瘤菌、共生特徵
Abstract
Crotalaria zanzibarica Benth., a perennial shrub native to Africa, is the most
widely-distributed naturalized legume in Taiwan. The plant, commonly distributed along roadsides and riverbanks, and in abandoned fields, established symbiosis with rhizobia forming root nodules. Root nodules are capable of fixing nitrogen. Accordingly, the symbiosis with nitrogen-fixing rhizobia might help C. zanzibarica colonizing nutrient-poor habitats. In this dissertation, I studied the symbiotic relationship between rhizobia and this legume, aiming to understand the diversity and possible origins of the symbiotic rhizobia and the beneficial effects of the symbiosis to C. zanzibarica.
A rhizobial strain, designed as CzR2, was isolated from the nodules of C.
zanzibarica grown in a greenhouse. This strain displayed pleomorphism, cell size
ranging from 2 to 10μm, in free-living state when cultivated in standard YEM medium which significantly differs from any known rhizobia. Based on the analysis of atpD-dnaK-glnII-gyrB-recA-rpoB gene set, CzR2 belongs to Bradyrhizobium arachidis.
Results of further experiments revealed that pleomorphism in this strain in its free-living state could be induced by mannitol, or fructose, but not by glucose.
Accordingly, the pleomorpism is substrate-dependent. CzR2 in its free-living state contained haploid and diploid cells, while that in symbiosis with C. zanzibarica was elongated with polyploidy, suggesting the occurrence of genomic endo-reduplication.
Legume-rhizobia symbioses of C. zanzibarica and six common legume species growing sympatrically along Xindian riverbank were investigated in Chapter 2. I found that these legumes form either determinate or indeterminate types of root nodules and harbored swollen or non-swollen bacteroids. Based on the 16S rRNA sequences, the symbionts of these legumes were classified as Bradyrhizobium, Neorhizobium, Rhizobium, Cupriavidus and Paraburkholderia. Irrespective of their possessing of
diverse symbiotic traits and nodule symbionts, the seven legume species had similar and consistently negative leaf δ15N values (mean of -1.2 ‰), and showed 15N enrichment (varying from 3.7 to 7.3 ‰) in their nodules. In addition, variations in the values of leaf δ13C (varying from -29 to -34‰) among the seven legumes were measured, indicating
their photosynthetic water use efficiencies were different. The results also suggested that C. zanzibarica could be nodulated by diverse rhizobia.
To compare the symbionts of C. zanzibarica and sympatric legumes growing along three distant riverbanks in Taiwan, I collected 59 isolates from this plant and 54 isolates from coexisting legumes. Based on the multilocus sequence analysis of concatenated dnaK-glnII-recA-rpoB gene sequences, the C. zanzibarica isolates were highly diverse,
belonging to 14 clades and varied among sampling sites, which can be either phylogenetically similar to or distinct from the isolates of coexisting legumes. The majority of C. zanzibarica isolates had nodA genes of American origin, following by
Asian origin, while others might be cosmopolitan.
To confirm the field isolates are able to nodulate C. zanzibarica and to compare the effects of symbionts on growth of this plant, I conducted single-strain inoculation experiment and investigated growth response, nodulation response, symbiotic efficiency and nitrogen relationship of C. zanzibarica inoculated with six rhizobial strains. The greenhouse inoculation experiment revealed that nodule and bacteroid morphologies in C. zanzibarica were rhizobial strain-dependent. Furthermore, C. zanzibarica plants
showed significant variation in total plant biomass and nitrogen accumulation among the strains inoculated, while there was very little variation in biomass allocation, nitrogen content, δ15N value and symbiotic efficiency among these tested plants.
Results of the greenhouse experiments and field investigations indicated that C.
zanzibarica was capable of forming effective symbiosis with diverse rhizobia, which
might confer the plant the ability of colonizing various habitats and contribute to its widely distribution in Taiwan.
Keywords: invasive plant, Crotalaria zanzibarica, legume-rhizobia symbiosis,
Bradyrhizobium, symbiotic traits
Contents
摘要...I
Abstract...III Contents…...VI LIST OF FIGURES…...VIII LIST OF TABLES…...X LIST OF ABBREVIATIONS...XI
General introduction………...1
Chapter 1 C. zanzibarica cultivated in the greenhouse was nodulated by a peculiar strain, CzR2...12
Abstract...……..13
Introduction...14
Materials and Methods...16
Results...20
Discussion...23
Figures and Tables...27
Chapter 2 Rhizobia symbiosis of C. zanzibarica and coexisting legumes growing along Xindian riverbank of Northern Taiwan...35
Abstract...36
Introduction...38
Materials and Methods...42
Results...45
Discussion...48
Figures and Tables...56
Chapter 3 Phylogenetic analyses of Bradyrhizobium symbionts associated with invasive C. zanzibarica and its coexisting legumes in Taiwan...64
Abstract...…….. 65
Introduction...66
Materials and Methods...68
Results...72
Discussion...76
Figures and Tables...83
Chapter 4 Evaluation the growth of C. zanzibarica inoculated by Bradyrhizobium strains...92
Abstract...……..93
Introduction...94
Materials and Methods...95
Results...97
Discussion...99
Figures and Tables...103
Conclusions...109
Future works...111
Literature cited...114
Supplementary materials...122
LIST OF FIGURES
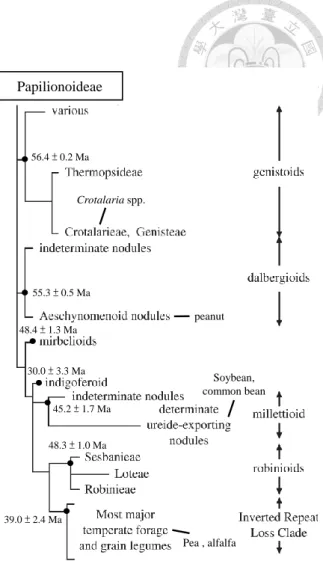
Figure A Phylogeny of the nodulating groups in Papilionoideae...4
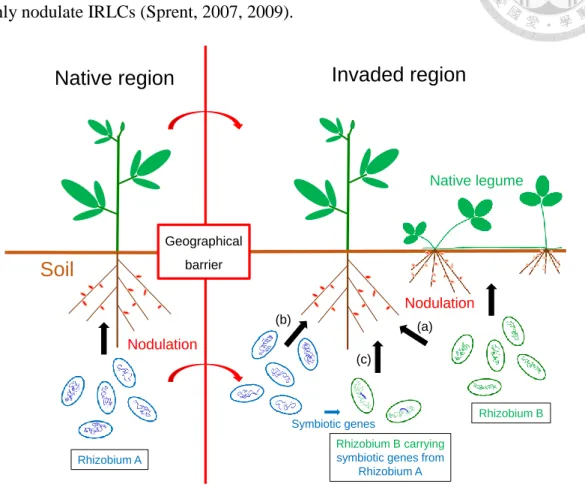
Figure B Three scenarios may contribute to the finding of exotic legume for the compatible rhizobia in the invaded areas...6
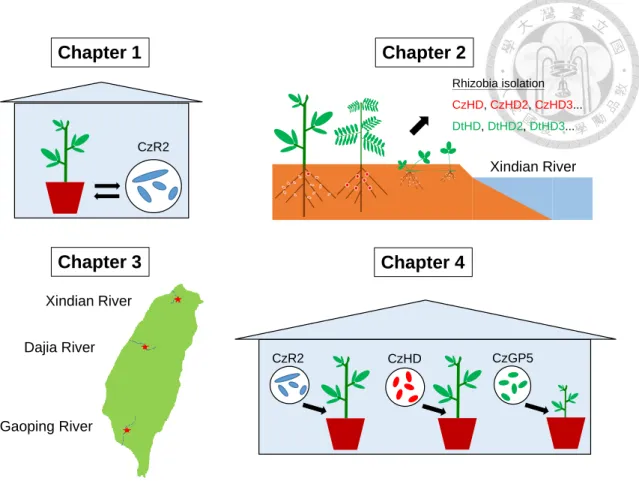
Figure C Illustration of the context of the four main chapters in this dissertation...11
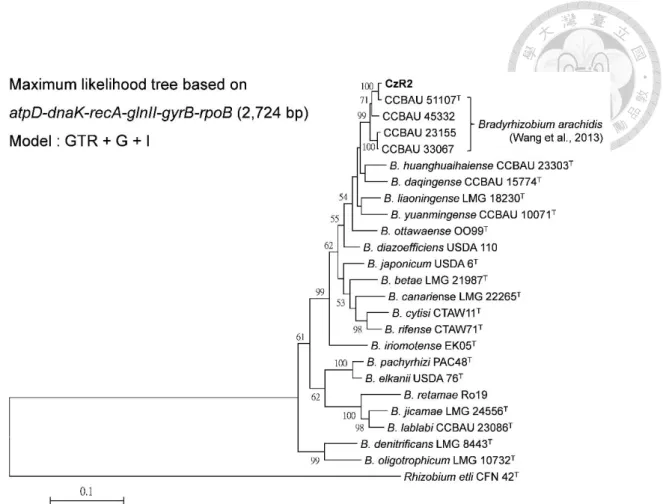
Figure 1-1 Phylogeny of strain CzR2 based on 16S rRNA genes...27
Figure 1-2 Phylogeny of strain CzR2 based on the six housekeeping genes...28
Figure 1-3 Cellular morphology of strain CzR2 and USDA 110 on YEM plates...29
Figure 1-4 Cell size variation during growth kinetics of strain CzR2 grown in YEM broth...30
Figure 1-5 Morphology of strain CzR2 grown with different substrates...31
Figure 1-6 Nodulation characteristics of C. zanzibarica infected by strain CzR2...32
Figure 1-7 Cellular morphology of strain CzR2 symbiotic with C. zanzibarica...33
Figure 1-8 Flow cytometry analyses of the DNA contents in free-living and symbiotic CzR2...34
Figure 2-1 The common legumes growing along Xindian riverbank...59
Figure 2-2 Nodule and bacteroid morphologies of common legumes growing along Xindian riverbank...60
Figure 2-3 Phylogenetic tree of 16S rRNA gene sequences of the nodule symbionts isolated from seven legumes growing along XD riverbank...61
Figure 2-4 Correlations between leaf and nodule δ13C values among the seven legume species coexisting along riverbank of Xindian River in Northern Taiwan...63
Figure 3-1 Phylogenetic identification of the Bradyrhizobium isolates from C. zanzibarica and its coexisting legumes by using MLSA technique...88
Figure 3-2 The nodA gene phylogenies of the Bradyrhizobium isolates from
C. zanzibarica and its coexisting legumes...90 Figure 4-1 Nodule morphology of C. zanzibarica inoculated with different rhizobial
strains at 41 DAS...105 Figure 4-2 Nodulation response for C. zanzibarica inoculated with six rhizobial
strains...106 Figure 4-3 Variation in growth of C. zanzibarica under different treatments...107 Figure 4-4 Relationship between total plant dry weight and total nodule dry weight in
C. zanzibarica inoculated with six rhizobial strains...108
LIST OF TABLES
Table 2-1 Characteristics of the seven leguminous species growing sympatrically along riverbank of Xindian River in Northern Taiwan...56 Table 2-2 Nitrogen contents and 15N signatures of seven legumes growing along XD
riverbank...57 Table 2-3 C ar bon con te nts a nd 1 3C signa tur es of sev en l eg u m es gr o w ing alo ng X D
riverbank...58 Table 3-1 Legume host and sampling sites of 113 Bradyrhizobium isolates analyzed in this study...83 Table 3-2 DNA polymorphism for Bradyrhizobium isolates collected from three riverbanks in Taiwan... ...84 Table 3-3 A summary of the number of isolates based on MLSA, from C. zanzibarica (Cz) and its coexisting native (CN) and exotic (CE) legumes in each of the three sampling sites (XD, DJ and GP) in Taiwan...85 Table 3-4 A summary of the number of isolates based on nodA, from C. zanzibarica
(Cz) and its coexisting native (CN) and exotic (CE) legumes in each of the three sampling sites (XD, DJ and GP) in Taiwan...86 Table 3-5 The possible origins inferred by nodA gene sequences of isolates collected
from C. zanzibarica and other native/exotic legumes in this study...87 Table 4-1 Profile of Bradyrhizobium strains used in this study...103 Table 4-2 Nitrogen concentration and stable isotope ratio of shoot, root and nodule of C. zanzibarica under different treatments...104
LIST OF ABBREVIATIONS
Culture medium
HM HEPES-MES
YEM Yeast extract mannitol
Phylogenetic analysis
NJ Neighbor-joining ML Maximum likelihood BI Bayesian inference
MLSA Multilocus sequence analysis
Plant physiology
δ13C Carbon stable isotope ratio δ15N Nitrogen stable isotope ratio WUE Water use efficiency
C
mass Carbon content on dry weight basis Nmass Nitrogen content on dry weight basisSampling site
XD River Xiandan River (新店溪) DJ River Dajia River (大甲溪) GP River Gaoping River (高屏溪)
General Introduction
As a consequence of globalization, many plant species were intentionally or accidentally transported into new ecosystems by human activities. Some of these exotic plants can colonize and invade into native communities in the introduced areas, disturbing the structure and function of ecosystems. Therefore, plant invasions have become one of the major threat to global biodiversity (Vitousek et al., 1997; Pimentel et al., 2005) and received much attention from the researchers.
Taiwan, with no exception, has 608 naturalized plant species which comprise 12%
of the total flora on this island (Wu et al., 2010). The family Fabaceae (Leguminosae) is one of the dominant families (79 species) among naturalized plants in Taiwan (Wu et al., 2010) and also the most contributor of the naturalized plants in the world (Pyšek, 1998).
In addition to their prevalence, naturalized legumes could have impacts on ecosystems.
Some naturalized legumes increased the soil nitrogen and altered the growth rate of other plant species, thus interfered the structure and function of plant communities (Vitousek, 1990; Yelenik et al., 2004). Wu et al. (2003) provided the background information (such as origin, habit, life form and minimum residence time) of naturalized legumes in Taiwan which was a starting point for evaluating the leguminous plant invasion in Taiwan. However, one of the most remarkable features of leguminous plants, the capability to form mutualistic symbiosis with rhizobia, is still poorly investigated in Taiwan. This thesis aims to understand the symbiotic relationship between the naturalized legume and its nodulating rhizobia in Taiwan.
In this chapter, I gave a brief introduction about the definitions of plant invasions, legume taxonomy, rhizobial symbionts of legumes, and studies of legume-rhizobia symbiosis in Taiwan. A summary of the objectives in this thesis was presented in the last section of this chapter.
1. Terminology of plant invasions
In 1958, Elton published the classic book ‘The Ecology of Invasions by Animals and Plants’ which first exploited the field of invasion ecology. However, the term
‘invasive species’ was not strictly defined in Elton’s book, and its meaning was not consistent among the published literatures, causing considerable confusion and misuse.
Some researchers suggested that exotic species which causing in economic and/or environmental damages or harm to human are considered invasive species (Cronk and Fuller, 1995; Mack, 1997). Some researchers defined the invasive species as exotic species which spread over large areas in the introduced sites (Richardson et al., 2000a;
Williamson, 1997).
The terminology used in this proposal followed the definitions of Richardson et al.
(2000a). The authors suggested that the spread of plants were limited by several barriers, including major geographical, environmental, reproductive and dispersal barriers., Plants introduced by human activities across major geographical barriers into new habitats are ‘alien plants’ (synonym: exotic plants). Alien plants which overcome the local environmental barriers in their introduced sites and sustain populations over many generations without human intervention, are referred as ‘naturalized plants’. If the naturalized plants could spread into new areas away from (> 100 m) their introduced sites, they are then defined as ‘invasive plants’. In addition, 50-80% of invasive plants which cause obviously negative impacts on native biodiversity are further categorized as ‘environmental weeds’. About 10% of invasive plants which are capable of changing the nature of ecosystem over considerable areas (such as nitrogen-fixer, sand stabilizers, fire promoters/suppressors...etc.) are referred as ‘transformers’.
Papilionoideae
56.4 ± 0.2 Ma
55.3 ± 0.5 Ma
48.4 ± 1.3 Ma
45.2 ± 1.7 Ma
48.3 ± 1.0 Ma
39.0 ± 2.4 Ma 30.0 ± 3.3 Ma
Modified from Sprent (2007) New Phytologist, 174:11-25 Crotalaria spp.
peanut
Soybean, common bean
Pea , alfalfa
2. Legumes of the world and in Taiwan
The family Leguminosae (Fabaceae), one of the largest plant families, contains more than 18,000 species throughout the world (Lewis et al., 2005; Sprent, 2009).Papilionoideae is the largest subfamily of legumes, and most of its members are able to form nodules with symbiotic rhizobia (Sprent, 2007, 2009). Lavin et al.
(2005) subdivided the major nodulating legumes within Papilionoideae into seven distinct groups, Genistoids, Dalbergioids, Mirbelioids, Indigoferoids, Millettioids, Robinioids and Inverted Repeat Loss clade (IRLCs) (Figure A), based on their fossil records and molecule data.
According to the book ‘Legume
Nodulation: A Global Perspective’ published by Sprent (2009), the global distribution of these papilionoid groups were distinctive. Crotalarieae and Genisteae, the two largest tribes in Genistoids, are mainly distributed in Africa and Mediterranean regions, respectively. Dalbergioids are largely in Central and South Americas. Millettioid is a pan-tropical group and world-wide spread. Mirbelioids are endemic to South Africa and Australia. Indigoferoid legumes are mostly distributed in Africa, considerable numbers are also found in Asia and Australia. Most of the genera in Robinioids and IRLCs are
Figure A. Phylogeny of the nodulating groups in Papilionoideae. Age estimates are shown at the nodes of each group based on the report of Lavin et al. (2005). Ma = million years. The important legumes belonged to these groups are indicated.
distributed in temperate regions.
Taiwan, a sub-tropical Pacific island, has 214 legume species, including 160 native species and 54 naturalized species (Huang, 1993; Wu et al., 2003). Thus, about 25% of the legume plants in Taiwan are introduced. Most of legumes in Taiwan belong to the subfamily Papilionoideae (including 140 native and 44 naturalized species). Among these papilionoid legumes, 127 species (106 native and 21 naturalized) belong to Milletioids, 26 species (20 native and 6 naturalized) to Genistoids, 16 species (4 native and 12 naturalized) to IRLCs, 11 species (7 native and 4 naturalized) to Dalbergioids and 4 species (3 native and 1 naturalized) to Robinioids.
3. Rhizobia play an important role in legume ecology
Rhizobia, comprising phylogenetically diverse soil bacteria, are able to induce nodules, a tumor-like structure hosting the nitrogen-fixing rhizobia, on stems or on roots of legume plants. Based on the 16S rRNA gene sequences, currently known rhizobia are divided into two classes, Alphaproteobacteria and Betaproteaobacteria, referred as alpha- and beta-rhizobia, respectively. The legume-rhizobia symbiosis is commonly found in Mimosoideae and Papilionoideae, but rarely in other subfamilies (Sprent, 2009). Mimosoideae usually establishes symbiosis with beta-rhizobia (such as Burkhoderia and Cupriavidus), whereas Papilionoideae is commonly nodulated by alpha-rhizobia (Sprent, 2007, 2009; Gyaneshwar et al., 2011). Alpha-rhizobia are highly diverse, including the classical rhizobia, such as Rhizobium, Sinorhizobium, Mesorhizobium, Bradyrhizobium and Azorhizobium (Young and Haukka, 1996), and the non-classical genus, Methylobacterium (Sy et al., 2001). The host specificity of alpha-rhizobia vary among genera, for examples Bradyrhizobium is a common
symbiont of Milletioid, Genistoid and Dalbergioid legumes, while Rhizobium spp.
mainly nodulate IRLCs (Sprent, 2007, 2009).
Rhizobia have been well known to improve legume yields in agriculture. Besides, rhizobia could also be critical factors affecting legume distributions in natural ecosystems. It has been reported that the growth of exotic legumes was limited by scarcity of compatible rhizobia when they were originally introduced to new habitats (Parker et al., 2006; Richardson et al., 2001b). Several scenarios allowing exotic legumes to find their nodule symbionts in the introduced areas have been reported (Figure B). For their successful establishment, the exotic legumes might recruit the
Native region Invaded region
Soil
Nodulation
Rhizobium A
Rhizobium B
Nodulation Native legume
Geographical barrier
Rhizobium B carrying symbiotic genes from
Rhizobium A Symbiotic genes (b) (a)
(c)
Figure B. Three scenarios may contribute to the finding of exotic legume for its compatible rhizobia in the invaded areas. (a) Exotic legume might recruit the local rhizobia from native legumes, (b) exotic rhizobia might be introduced accompanying the introduction of exotic legume, and (c) exotic legume might be associated with the rhizobia which are phylogenetically similar to local rhizobia but carrying the symbiotic genes from exotic rhizobia.
indigenous rhizobia in the introduced areas (Parker et al., 2006). Alternatively, the exotic legumes and their nodule symbionts might have co-introduced from native ranges into invaded regions (Rodríguez-Echeverría, 2010; Crisóstomo et al., 2013). For example, Horn et al. (2014) reported that an invasive legume, Cytisus scoparius in North America obtained its compatible Bradyrhizobium symbionts from heterogeneous ancestry. Some were associated with native rhizobia from native legumes of North America. Some utilized exotic rhizobia of European C. scoparius (the ancestral population). Besides, the authors also found that some symbionts of C. scoparius were phylogenetically similar to native rhizobia but displayed European originated symbiotic genes.
4. Studies of legume-rhizobia symbiosis in Taiwan
Some of Mimosa plants are notorious invasive plants in the world (Binggeli, 1996).
Among them, Mimosa pudica is listed as the world’s worst weed (Holm et al., 1991) and also as one of the 20 major environmental weeds in Taiwan (Chiang et al., 2003).
Chen et al. (2003) examined 190 nodule symbionts isolated from M. pudica and its congener, M. dipliotricha distributed in 14 sites across Taiwan Island. Based on 16S rRNA gene sequences, 93% of isolates belonged to Ralstonia taiwanensis (later renamed Cupriavidus taiwanensis). In contrast, M. pigra, a newly naturalized plant in Taiwan (Wu et al., 2010), was mainly nodulated by Burkholderia (Chen et al., 2005).
Barrett and Parker (2005) reported that M. pudica and M. pigra growing in their native ranges (Panama) were commonly associated with Burkholderia strains. These results suggested that naturalized legumes in Taiwan could utilized the compatible rhizobia distinct from or similar to those symbionts in their original regions.
Sesbania cannabina, belonging to Robinioid group, has been used as a common green manure in Taiwan and is also on the list of the 20 major environmental weeds in Taiwan (Chiang et al., 2003). Chen and Lee (2001) investigated 18 nodule symbionts isolated from S. cannabina grown in the sugarcane fields, locating in the southern part of Taiwan. According to amplified 16S rDNA restriction analysis (ARDRA) and 16S rDNA sequencing, 18 isolates were separated into four genotypes which belonged to Sinorhizobiun and Rhizobium. Furthermore, the symbiotic genes (nifH) of these isolates were also diverse. This result indicated that naturalized S. cannabina plants in Taiwan were nodulated by phylogenetically diverse rhizobial community even though they were grown in adjacent fields.
Hung et al. (2005) reported the nodule symbionts of seven native legumes, including six Millettioid legumes and one Genistoid, growing in the central part of Taiwan. Among the 83 isolates they examined, most of strains belonged to Rhizobium, Bradyrhizobium and Agrobacterium, whereas one Sinorhizobium strain and one Burkholderia strain were also isolated.
Taken together, the aforementioned studies revealed that rhizobia associated with legumes in Taiwan were highly diverse. However, most of these studies used the 16S rRNA gene analysis and phenotypic traits to characterize the isolated rhizobia. It has been suggested that the 16S rRNA genes and phenotypic traits of rhizobia are variable.
In addition, the origin of these rhizobia and the specific relationship between these rhizobia and their legume hosts were rarely studied.
5. Objectives and studies of this study
Crotalaria plants, containing around 600 species mainly native to Africa, were
introduced to many countries as green manures or fodders and became the common exotic legumes found throughout the world (Polhill, 1982). Nine naturalized Crotalaria species were recorded in Taiwan, including C. bialata Schrank, C. incana L., C.
lanceolata E. Mey, C. linifolia L. f., C. micans Link, C. pallida Ait., C. spectabilis Roth., C. triquetra Dalzell and C. zanzibarica Benth. (Wu et al., 2010). C. zanzibarica, first documented in 1934, has been evaluated as a naturalized species with the highest invasiveness in Taiwan (Wu et al., 2003, 2005). This plant mainly distributed in the nutrient-poor habitats, such as roadsides, riverbanks and abandoned fields, commonly formed root nodules (Wu et al., 2005). In a preliminary study, I cultivated C.
zanzibarica, neither inoculated nor fertilized, in a greenhouse of NTU and found some individuals formed root nodules and looked much healthier than the others without such structures. I therefor suspected that the nitrogen-fixing symbiosis of C. zanzibarica might contribute to its successful colonization and widely distribution in Taiwan.
Despite the fact that Crotalaria is the largest Genistoid legume and spreads worldwide (Sprent, 2009), the studies on its symbiotic characteristics are relatively few. In addition, the symbiotic relationship between legumes and nitrogen-fixing rhizobia in Taiwan were also rarely exploited. The observation and literature survey invoked my interests in studying the relationship between C. zanzibarica and its symbionts.
In this thesis, I focused on the symbiotic characteristics of naturalized C.
zanzibarica in Taiwan, and the specific objectives are: (1) to investigate the diversity, possible origin and host specificity of rhizobia associated with C. zanzibarica, (2) to document the symbiotic related traits of C. zanzibarica, including nodule type, bacteriod morphology, nitrogen content and nitrogen stable isotope, and (3) to understand how rhizobial strains collected from divergent sources varied in their
symbiotic performance on C. zanzibarica plant. To begin with, I isolated and identified the nodule symbiont of greenhouse-grown C. zanzibarica and characterized the nodule, bacteroid and symbiosome features. The results are presented in Chapter 1.
Following the results of Chapter 1, in order to understand whether field-growing and greenhouse-grown C. zanzibarica had similar symbiotic characteristics, I investigated C. zanzibarica growing along Xindian riverbank in the northern part of Taiwan. In addition, to further examine whether C. zanzibarica shared nodule symbionts with sympatric legumes along Xindian riverbank, several co-existing legumes were also analyzed. Results were presented in Chapter 2.
After investigating the Xindian population, I extended the investigation into two other C. zanzibarica populations growing along Dajia and Gaoping riverbanks, locating in the central and southern parts, respectively, of Taiwan. In Chapter 3, I reported the phylogeny and possible origins of rhizobia isolated from these three C. zanzibarica populations.
In previous studies (Chapter 1 and 2), I found that plants of C. zanzibarica cultivated in a greenhouse of NTU and growing along the three riverbank were nodulated by different Bradyrhizobium strains and displayed distinctive bacteroid sizes.
In Chapter 4, I conducted the single-strain inoculation experiment to confirm the symbiotic relationship.
A figure illustrating the main theme of each chapter in this study is shown in
Figure C.
Chapter 1 Chapter 2
CzR2
Xindian River
CzR2
Chapter 4
CzHD
Chapter 3
Xindian River
Dajia River
Gaoping River
Rhizobia isolation CzHD, CzHD2, CzHD3...
DtHD, DtHD2, DtHD3...
CzGP5
Figure C. Illustration of the context of the four main chapters in this dissertation.
Chapter 1
C. zanzibarica cultivated in the greenhouse was nodulated by a peculiar strain, CzR2
(This chapter has been published in Microbes and Environments 31: 410-417, 2016.
DOI information: 10.1264/jsme2.ME16063)
Abstract
Crotalaria zanzibarica is an exotic, widely-distributed leguminous plant in Taiwan.
To understand traits contributing to its successful invasion, I cultivated this plant in a greenhouse. Without fertilizer and rhizobia added, the plants still formed root nodules and had high leaf nitrogen contents (5-8%). A rhizobial strain, designed as CzR2, was isolated from these nodules. This strain displayed pleomorphism, cell size ranging from 2 to 10 µm, in free-living state when cultivated in standard YEM medium which significantly differs from any known rhizobia. In this study, I identified and studied the characteristics of the strain, and observed, the nodules formed by the strain. The objective of the study was to understand the relationship between C. zanzibarica and its rhizobial symbionts.
Results of multilocus sequence analysis of atpD, dnaK, glnII, gyrB, recA and rpoB genes confirmed that CzR2 belongs to Bradyrhizobium arachidis. CzR2 was uniformly rod-shaped in basal HM medium but it displayed pleomorphic cells when mannitol or fructose was added which indicates its unusual morphology in YEM medium is caused by the gradient mannitol. No infection thread was found but highly pleomorphic bacteriods and a symbiosome containing several bacteroids were often observed under FM and TEM observations of the indeterminate nodules of C. zanzibarica formed by the inoculation of CzR2. Results of the flow cytometric analysis revealed that CzR2 cells in YEM medium and in nodules of C. zanzibarica had two and three peaks of relative DNA contents, respectively. It suggests that elongated cells of CzR2 in free-living state might result from cell-cycle delay while those in symbiotic state from genomic endo-reduplication.
Introduction
Soil bacteria that establish symbiotic relationships with legumes and induce tumor-like, N2-fixing structure (nodule) on roots are referred to as rhizobia. Rhizobia have been shown to improve legume yields in agriculture (Thies et al., 1991). Most of the research of legumes-rhizobia symbioses focused on the legume crops and model legumes, such as Glycine, Pisum, Medicago and Lotus, but relatively rare in other legumes (Doyle and Luckow, 2003). Besides the agricultural benefits, rhizobia could also play a critical role in affecting legume distributions in natural ecosystems. It has been reported that the growth of exotic legumes were limited by scarcity of compatible rhizobia when they were first introduced to new habitats (Richardson et al., 2000b; Park et al., 2006). For their successful establishment, the naturalized legumes (“naturalized plants” in this report are exotic plants that can sustain populations over many generations without human intervention, defined by Richardson et al. (2000a) should acquire their symbionts from newly occupied habitats. Although legumes are the most contributors of the naturalized plants of the world (Pyšek, 1998), much less is known about their compatible rhizobia in their introduced regions and how specific these symbionts are to host plants.
The genus Crotalaria, containing around 600 species distributed in tropical and subtropical regions, is a member of the subfamily Papilionoideae and most of its members are native to Africa (Polhill, 1982). Some species of Crotalaria were introduced to countries as green manures or fodders (Polhill, 1982). Despite the fact that Crotalaria is a large genus and spread worldwide, studies on its symbiotic rhizobia are relatively few. According to the limited literatures, Bradyrhizobium strains are the most common symbionts isolated from Crotalaria species (Samba et al., 1999; Liu et al.,
2007; Aserse et al., 2012). However, some nonclassical rhizobial strains, such as Methylobacterium (Sy et al., 2001) and Burkhoderia (Liu et al., 2007), have also been reported to nodulate Crotalaria spp..
Crotalaria has been introduced into Taiwan for more than 80 years, and is the largest genus of the naturalized legumes of the island (Wu et al., 2003). Crotalaria zanzibarica is a perennial leguminous shrub, native to Africa, and becomes the most widely-distributed naturalized legume in Taiwan (Wu et al., 2005). This plant, mainly distributed along roadsides and riverbanks, and in abandoned fields, commonly established symbiosis with rhizobia forming root nodules. C. zanzibarica, despite being evaluated as a species of wide distribution and of highest invasiveness among the naturalized legume in Taiwan (Wu et al., 2003, 2005), the relationship between its distribution and the symbiotic rhizobia has not been addressed. In a preliminary study, I grew C. zanzibarica, neither inoculated nor fertilized, in a greenhouse and found some individuals formed root nodules and looked much healthier than the others without such structures. In addition, the nodules and leaves of the plant accumulated unusually high nitrogen contents, 6-9% and 5-8%, respectively (Huang and Kao, unpublished). The results prompted us to study the symbiotic relationship between the plant and its root-nodulating rhizobia.
In this study, a peculiar rhizobium, with pleomorphism both in YEM medium and in symbiotic state, from root nodules of greenhouse-grown C. zanzibarica was isolated.
Based on its slow-growing phenotype, I suspected that this isolate as a species of Bradyrhizobium. In this study, I analyzed 16S rRNA gene sequence and several housekeeping and symbiotic genes of this rhizobium to elucidate its species identity.
The phenotypic variations, in particular the causes of pleomorphism in YEM medium,
and symbiotic characteristics with C. zanzibarica of the rhizobium were also investigated.
Materials and methods
Isolation of rhizobia from root nodules of C. zanzibarica
Nodules of C. zanzibarica grown in a greenhouse of National Taiwan University (Taipei City, Taiwan) were collected. Fresh nodules were surface sterilized by immersion in 0.5% SDS for 1 min, then 70% ethanol for 5 min, and washed three times by sterile deionized-distilled water (DDW). Nodule suspension was obtained by crushing the nodule in DDW and spread onto YEM agar plate (grams per litter: NaCl, 0.2; MgSO4・7H2O, 0.2; K2HPO4, 0.5; FeSO4・7H2O, 0.005; mannitol, 10.0; yeast extract, 0.4; agar, 15.0; the pH was adjusted to 6) placing at 30℃. When putative rhizobial colonies were visible, single colony was picked and checked for unity by repeated streaking on YEM agar plate. A pure culture was obtained and designated as strain CzR2. For comparative purpose, a reference strain, Bradyrhizobium japonicum USDA 110 was purchased from Bioresource Collection and Research Center (BCRC, Hsinchu, Taiwan). Pure cultures of each strains were stored in 15% glycerol-YEM broth at -80℃.
Molecular identification of strain CzR2
Total genomic DNA was extracted from the pure culture of strain CzR2 grown in YEM broth until the late exponential phase of growth. Extraction of the DNA was performed by using Geneaid DNA Mini kit (Geneaid). The 16S rRNA genes were amplified by using bacterial universal primers (Marchesi et al., 1998).
The other six housekeeping genes (atpD, dnaK, glnII, gyrB, recA and rpoB genes) and three symbiotic genes (nodA, nodZ and nifH) were amplified by the primers taken from published literatures (Menna and Hungria, 2011; Stępkowski et al, 2005; Parker, 2012). Sequencing primers of all genes were the same primers used for PCR amplification.
The aforementioned sequences and reference sequences of housekeeping genes were aligned by using the CLUSTALW program (Thompson et al, 1994). Phylogenetic identification of strain CzR2 was conducted by analyzing 16S rRNA and combined atpD-dnaK-glnII-gyrB-recA-rpoB genes dataset, respectively. Maximum likelihood (ML) phylogenies and model tests were both performed by using software MEGA version 6 (Tamura et al., 2013). The best-fit models for ML analyses were T92+G+I for 16S rRNA gene and GTR+G+I for atpD-dnaK-glnII-gyrB-recA-rpoB genes dataset. The topology of the trees was evaluated by bootstrapping with 1,000 replications. All the sequences obtained in this study were deposited in GenBank database. The GenBank accession numbers of strain CzR2 generated in this study are KJ125399 (16S rRNA), KU315329 (atpD), KU000974 (dnaK), KU000975 (glnII), KU315331 (gyrB), KU001035 (recA), KU001095 (rpoB), KJ125402 (nodA), KJ135034 (nodZ) and KJ135031 (nifH).
Cellular morphology and growth dynamic in YEM medium
Both CzR2 and the reference strain were grown on YEM agar medium at 30°C for 7 days. Single colony of each strain was picked and stained with DAPI (4,6-diamidino-2-phenylindole; Sigma-Aldrich) at 50 μg/ml for 10 min at 25 ℃. The cell samples were examined by the microscopy (BX51, Olympus, Japan) under a bright
or fluorescent field, following the method reported by Liu et al. (2011). To measure the bacterial cell size, at least ten random fields of view were photographed and more than 1,000 cells of each sample were examined by using ImageJ 1.48 software.
For constructing the growth curves, strain CzR2 and USDA 110 were inoculated into 3 ml of fresh YEM broth (five replicates for each strain) and grown at 30 ℃, 200 rpm on a shaking table. The cell growth was evaluated as the number of CFU ml-1 and OD600 measurement. CFU ml-1 was counted every 24 hours and OD600 value was measured every eight to ten hours. The mean generation times were calculated from the exponential phase of growth based on the OD600 measurement. The cell size was also measured every 24 hours during cell growth.
Substrate tests in HM medium
Because YEM media containing yeast extract is a complex medium, the defined medium, HM (HEPES-MES) medium (Cole and Elkan, 1973) was used to conduct the substrate tests. Strain CzR2 were grown on the HM basal plates and these media comprised of different substrates (0.04% or 0.3% yeast extract, 0.5% mannitol, 0.5%
fructose and 0.5% glucose, respectively) at 30°C for 7 days.
To compare the growth response among different substrates, I also inoculated strain CzR2 into 3 ml HM basal broth and the broths added 0.04% yeast extract, 0.5%
mannitol, 0.5% fructose or 0.5% glucose. In all substrate tests, strain USDA 110 was also included for comparative purposes. Additionally, B. arachdis CCBAU 51107T was tested its morphology grown in YEM, HM basal broths and HM broths with different substrates.
Morphological features of CzR2 infected nodules
Seeds of C. zanzibarica were sterilized using 5% SDS and 10 mM NaCl for 5 min, then 70% ethanol for 30 min and finally drilled by sterilized needle to break seed coat.
The sterilized seeds were germinated in a Petric dish at room temperature for 2 days.
After germination, seedlings were transferred into 1L glass pots filled with a sterilized mixture of peat soil, vermiculite and perlite in a proportion of 5:2:2, respectively. Plants were irrigated with distilled water regularly. After 4-5 days of growing in the pots, plants in pots were inoculated either with 600 μl of strain CzR2 at exponential phase in YEM broth or with a YEM liquid media without rhizobial culture as a negative control.
All the seedlings were grown in a humidity controlled (70%) growth camber with a 12 hours of light period (photosynthetic active radiation of 100~150 μmol m-2 s-1, at 28℃) and a 12 hours dark period (at 25℃). When plants were 40 days old, they were transferred from the growth camber to a glasshouse and grown in the 2 L plastic pots in natural daylight. For histological observation, branched nodules from two months old plants were fixed by FPGA (formalin: propionic acid: glycerol 95%: EtOH:
H20=1:1:3:7:8) and sectioned to a thickness of 15μm by using a microslicer DTK-1000.
Nodule sections were stained with 1% toluidine blue O (TBO), and then examined by light microscopy.
Observations of symbiotic CzR2 (bacteroids) under fluorescent and transmission electron microscopies
Nodule samples were collected from mature C. zanzibarica plants (two months old) which were inoculated by strain CzR2 and grown in a NTU greenhouse. For FM observation, bacteroids were extracted from fresh nodules and stained with DAPI,
following the aforementioned protocol. For TEM observation, nodule samples were fixed with 2.5% glutaraldehyde, dehydrated in 15-100% acetone series, and then, embedded with Spurr’s resin. Ultrathin sections (80 nm) were conducted by using an ultramicrotome (PowerTome XL) and analyzed on a transmission electron microscope (Hitachi H-7650) at 100 kV.
DNA content of free-living and symbiotic CzR2
Free-living CzR2 was grown in YEM broth for 7 days and symbiotic bacteroids were extracted from mature root nodules of C. zanzibarica (67 days old). These cells were fixed in 90% ethanol for 16 h at 20°C, then washed twice with PBS followed by centrifugation for 2 min at 4,000 rpm. Pelleted cells were stained with propidium iodide (PI)-RNase staining buffer solution (BD Biosciences) for 30 min at room temperature.
For each flow cytometry experiment, the DNA content was measured in a population of 20,000 cells with a Cytomics FC500 analyzer (Beckman Coulter Ltd.). Data analysis was performed with CXP software (Beckman Coulter Ltd.).
Results
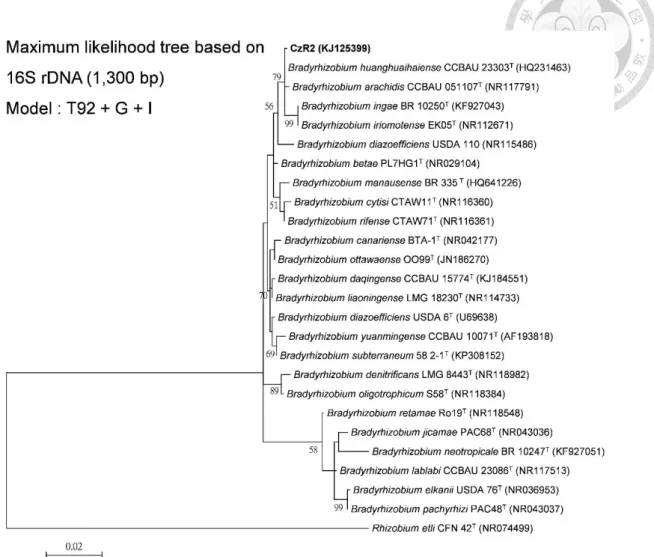
Phylogenetic identification of strain CzR2 by using 16S rRNA gene and MLSA of atpD, dnaK, glnII, gyrB, recA and rpoB genes
In the phylogenetic tree of 16S rRNA gene (Figure 1-1), strain CzR2 was grouped together with four Bradyrhizobium type strains, and all of them had nearly identical sequences. Strain CzR2 showed 99.92% similarity to B. huanghuaihaiense CCBAU 23303T, 99.77% similarity to B. arachidis CCBAU 51107T and 99.39% similarity to both B. iriomotense EK05T and B.ingae BR1025T.
Maximum likelihood analysis of the concatenated six housekeeping genes (atpD, dnaK, glnII, gyrB, recA and rpoB) revealed that strain CzR2 and four CCBAU strains formed a well-supported group (Figure 1-2). These four CCBAU strains were reported that they belonged to a novel species, Bradyrhizobium arachidis (Wang et al., 2013).
Comparison of these six genes among related strains, strain CzR2 was most similar (99.27% identity) to the type stain of B. arachidis, CCBAU 51107, which is peanut symbiont isolated from Hebei, China.
Similarities of symbiotic genes between strain CzR2 and CCBAU 51107
Beside the housekeeping genes, I also compared the symbiotic gene sequences between strain CzR2 and CCBAU 51107. These two strain had 98.71% similarity in nodA gene (460 bp) and 99.85% similarity in nifH gene (648 bp). The nodZ gene sequence of CCBAU 5117 was not available in GenBank.
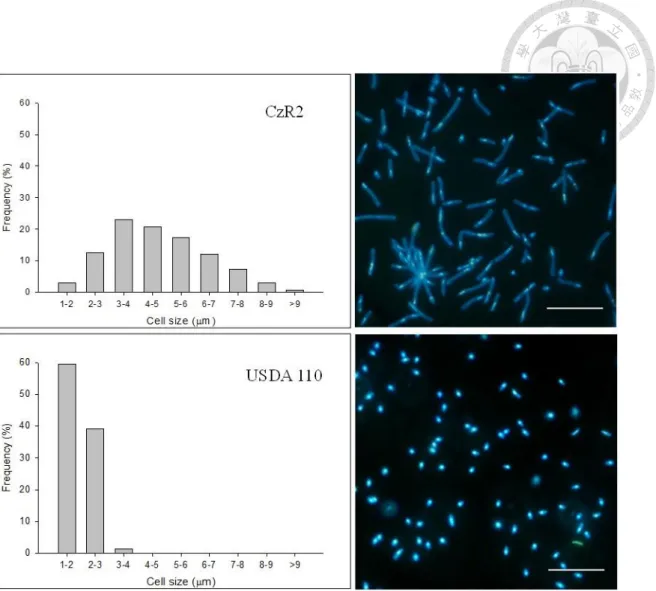
Comparison of cellular morphology and growth dynamic between strain CzR2 and USDA 110 in YEM medium
Strain CzR2 and USDA 110 both showed slow-growing phenotype which formed detectable colonies on YEM plate after 5-6 days of incubation. The cellular morphology of these two strains was observed by optical microscopy. As shown in Figure 1-3, strain CzR2 was highly various in cell size (1.4 to 10.9 μm in length) and branched cells were occasionally observed (1-2 % in examined cells). On the other hand, USDA 110 was uniformly rod-shaped and the cell length was from 1.3 to 3.4 μm under the same growth condition.
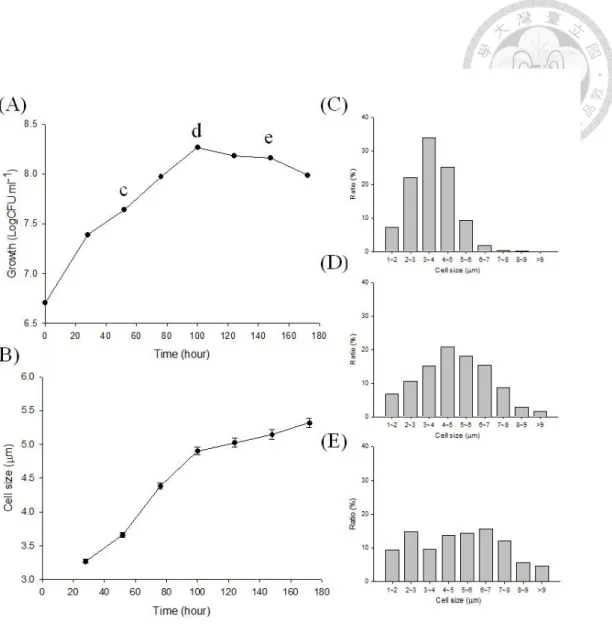
The mean length of strain CzR2 dramatically increased from 3.2 to 5.0 μm during
exponential growth phase (Figure 1-4), about 3% of these cells were branched. In contrast, the cell size of USDA 110 was consistent (about 2 μm) in any growth phases (data not shown).
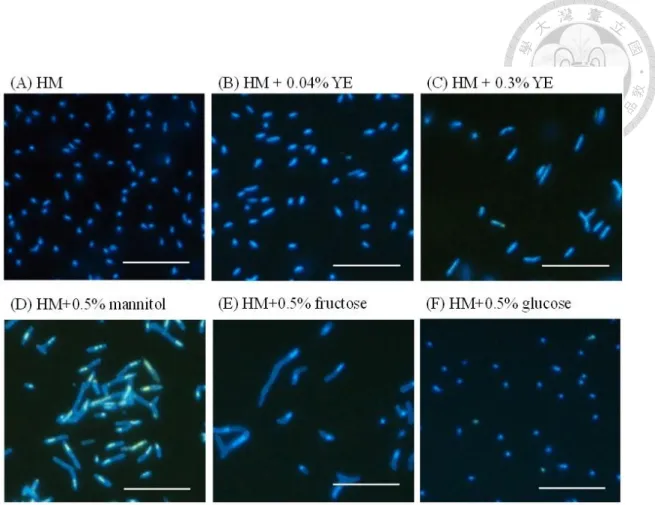
Effect of substrates on the morphology of free-living strain CzR2
Strain CzR2 was uniformly rod-shaped on the basal HM plate. When 0.04 or 0.3%
yeast extract were added, it remained uniformly rod-shaped. In contrast, when it was grown on the HM plate with 0.5% mannitol displaying pleomorphic cells. Furthermore, the pleomorphism of strain CzR2 can also be induced by adding fructose, but not by glucose (Figure 1-5). On the other hand, the cells of USDA 110 grown with any tested substrates were uniformly rod-shaped.
Symbiotic characteristics of strain CzR2 in C. zanzibarica
Strain CzR2 can induce C. zanzibarica to form visible nodules (1 mm) at 7 dpi (days post inoculation). The nodules of seedlings at 35 dpi became branched, and that of plants at 56 dpi displayed multi-lobed morphology (Figure 1-6 ABC). On the other hand, the control plants never formed any nodule in the growth chamber. Nodule sections showed that all plant cells in the infection zone were uniformly infected and no infection thread was formed (Figure 1-6 DE).
Bacteroids extracted from the multi-lobed nodules of C. zanzibarica were highly pleomorphic and some were longer than 10μm in length (Figure 1-7A). TEM images of nodule section also showed that strain CzR2 displayed pleomorphism inside the host cells. Besides, symbiosomes commonly comprised multiple bacteroids (Figure 1-7B,
C).
The results of flow cytometric analysis were shown in Figure 1-8. Symbiotic CzR2 isolated from nodules of C. zanzibarica showed three peaks (1C, 2C and 3C). On the other hand, free-living CzR2 was composed of two peaks (1C and 2C).
Discussions
In this study, I isolated a rhizobial strain, CzR2, from roots of greenhouse-grown C.
zanzibaria and found this strain displayed bacteroid-like morphology in free-living states. To my knowledge, this is the first report that free-living rhizobia showed pleomorphism in standard YEM medium. Based on the MLSA of six housekeeping genes, strain CzR2 can be classified as Bradyrhizobium arachidis. Strain CzR2 and B.
arachidis CCBAU 51107T were nearly identical (>99% similarity) in the both housekeeping and symbiotic gene sequences. Wang et al. (2013) reported that B.
arachidis is a peanut symbiont in China, with rod-shape of 1.30 to 1.97 μm in length, and its generation time was 8.8 hours in YEM broth. In consistent with their report, strain CzR2 is able to nodulate peanut (data not shown). However, cell length (Figure
1-3) and generation time (ca. 30.2 hours, data not shown) of CzR2 were significantly
different from those of B. arachidis CCBAU 51107T reported by Wang et al. (2013). To elucidate the discrepancy, I purchased and cultured B. arachidis CCBAU 51107T (LMG 26795), deposited in Belgian Coordinated Collections of Microorganisms, and confirmed this type strain also displayed pleomorphism in standard YEM medium (data not shown).Bacteria usually maintained their morphological uniformity due to equal cell-division (Young, 2006). A pure bacterial culture displaying diverse cell shape and size (called pleomorphism) was considered as the product of old culture (composing
ageing and dying cells) or contaminants (Wainwright, 1997). In this study, strain CzR2 in YEM broth gradually increased its cell size at the early exponential phase (Figure
1-4), when cells were young. Even when CzR2 was sub-cultured repeatedly or
re-isolated from legume hosts, pleomorphism was still present. In contrast, reference strain USDA 110 never showed such unusual morphology under the same culture conditions. Taken together, these results confirm that pleomorphic phenotype of CzR2 is a genuine feature rather than artifact.It has been reported that some bacteria produced filamentous cells in response to specific environmental cues. Steinberger et al. (2002) reported that cells of Pseudomonas aeruginosa grown in media with low nutrient was significantly elongated but their width remained unchanged. As consequence, the ratio of cell surface area to cell volume was increased which might enhance the absorbance of nutrients. In addition, several pathogenic bacteria produced elongated cells when phagocytes or antibiotics existed, hence reduced the chance of being killed (Justice et al., 2008). Elongated cells of free-living rhizobia have also been reported when they were cultured in the medium with dicarboxylate or high concentration of yeast extraction (Reding and Lepo, 1989;
Skinner et al., 1977). In this study, I showed that fructose and mannitol can induce strain CzR2 from uniformly rod-shaped cells to highly pleomorphic and elongated cells (Figure 1-5), which has not been found before. The biological roles of this unusual phenomenon are currently understudy.
When inoculated with CzR2, C. zanzibarica produced multi-lobe indeterminate nodules (Figure 1-6C), in consistence with the specific nodule type (crotalarioid) described by Corby (1988). Furthermore, the infection zone of nodules only contained infected cells and infection thread was never observed (Figure 1-6DC), suggesting that
nodulation process of C. zanzibarica was not though root-hair infection pathway. These results supported the idea that the nodulation of genistoid legumes were commonly by crack and epidermal infections, and few infected cells divided repeatedly to form uniform infected zone (Sprent, 2007). However, distinctive features were found inside the nodule cells of the C. zanzibarica inoculated by CzR2, for example, symbiosomes commonly contained multiple bacteroids with pleomorphism (Figure 1-7). In the nodules of pea, each symbiosome usually harbors a single but pleomorphic (such as swollen, elongated, or branched) bacteroid. In contrast, in the nodules of soybean, each symbiosome usually contains multiple bacteroids of uniformly rod-shape (Haag et al., 2013; Mergaert et al., 2006; Oono et al., 2009).
Although strain CzR2 formed filamentous cells in both free-living and symbiotic states, flow cytometric analysis revealed that those cells had different DNA contents (Figure 1-8). It suggests that elongated cells of strain CzR2 in free-living state might result from cell-cycle delay while those in symbiotic state from genomic endo-reduplication. The results that adding fructose to HM medium can induce cell elongation of CzR2 (Figure 1-5) suggest that cellular morphology of strain CzR2 in symbiotic state might be also regulated by fructose.
In conclusion, the rhizobium CzR2 isolated from nodules of C. zanzibarica belongs to a Bradyrhizobium arachidis strain. Pleomorphism of this strain in the free-living state can be in induced by adding mannitol, fructose, but not by glucose, to the HM medium, thus, it is substrate dependent. CzR2 in free-living contain haploid and diploid cells while when symbiotic with C. zanzibarica were elongated and with polyploidy, indicating the occurrence of genomic endo-reduplication. In this C.
zanzibarica-CzR2 symbiosis, symbiosomes commonly contained multiple, pleomorphic
bacteroids which is also an unusual feature.
Figure 1-1 Phylogeny of strain CzR2 based on 16S rRNA genes. Maximum
likelihood tree based on partial 16 rRNA genes (1,300 bp) showing the relationships among strain CzR2 and defined Bradyrhizobium species. Only bootstrap value > 50 are shown at the internodes. The scale bar represents 2 % nucleotide substitutions.Figure 1-2 Phylogeny of strain CzR2 based on the six housekeeping genes.
Maximum likelihood tree based on concatenated atpD-dnaK-glnII-gyrB-recA-rpoB gene sequences (2,724 bp) showing the relationships among strain CzR2 and defined Bradyrhizobium species. Only bootstrap value > 50 are shown at the internodes. The scale bar represents 10 % nucleotide substitutions.
Figure 1-3 Cellular morphology of strain CzR2 and USDA 110 on YEM plates.
Size variation and distribution of strain CzR2 (Top) and USDA 110 (down) grown on YEM plate for 7 days. In each strain, at least 1,000 cells were examined. The pictures showed these cells stained by DAPI. Bar, 10 μm.
Figure 1-4 Cell size variation during growth kinetics of strain CzR2 grown in YEM
broth. Growth was evaluated as the number of Log CFU ml
-1 (A) and mean cell size (B), error bars in (B) indicate standard errors (n >1000). Distribution and frequency of cells with different size were also calculated in the exponential phase (C), transition phase (D) and stationary phase (E).Figure 1-5 Morphology of strain CzR2 grown with different substrates. CzR2 was
cultivated on the plates of HM (HEPES-MES) basal medium (A), HM+0.04% yeast extract (YE) (B), HM+0.3% YE (C), HM+0.5% mannitol (D) HM+0.5% fructose (E) and HM+0.5% glucose (F) for 14 days. Cells were stained with DAPI, bar = 10 μm.Figure 1-6 Nodulation characteristics of C. zanzibarica infected by strain CzR2.
The nodules of C. zanzibarica inoculated by strain CzR2 were at 7 dpi (days post inoculation) (A), 35 dpi (B) and 56 dpi (C), bar = 1 cm. Nodules were longitudinally sectioned and stained with toluidine blue O (D and E). The darkly-stained infection zone (I) were surrounded by cortex (Ⅱ) and vascular bundles (black arrow). The apical nodule meristem (M) showed two distinct cell groups, uninfected cells (UC) and infected cells (IC).
Figure 1-7 Cellular morphology of strain CzR2 symbiotic with C. zanzibarica. (A)
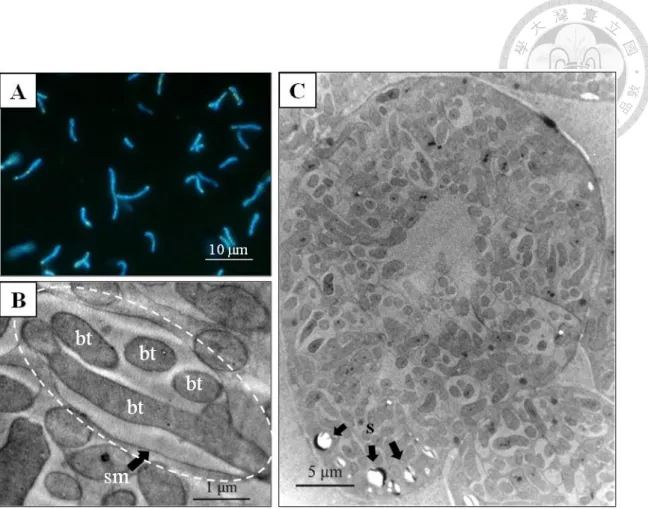
Bacteroids of strain CzR2 stained with DAPI displayed pleomophism. (B) TEM images of root nodule of C. zanzibarica showed that a single symbiosome (dashed circle) was encompassed by the symbiosome membrane (sm) and harbored multiple bacteroids (bt).(C) Infected cells were filled with bacteroids except for central region and starch grains (s) scattered on the periphery.
Figure 1-8 Flow cytometry analyses of the DNA contents in free-living and
symbiotic CzR2. Cells were stained with propidium iodide (PI). Free-living CzR2 (A)
was grown in YEM broth and sampled from late log phase, and bacteroids (B) were extracted from the nodules of 67 days old C. zanzibarica. The x axis shows fluorescence levels, indicating the DNA contents and the y axis shows cell counts. In each experiment, 20,000 cells were analyzed.Chapter 2
Rhizobia symbiosis of C. zanzibarica and coexisting legumes growing along Xindian riverbank of
Northern Taiwan
(This chapter has been published in Taiwania 68: 7-15, 2018. DOI information:
10.6165/tai.2018.63.7)
Abstract
Legume-rhizobia symbioses of seven leguminous species growing along Xindian riverbank of Northern Taiwan were investigated in this study. These legumes form either determinate or indeterminate types of root nodules. The determinate nodules of Alysicarpus vaginalis, Desmodium. triflorum, D. heterophyllum, Sesbania cannabina
and the indeterminate nodules of Mimosa pudica harbored bacteroids of morphological uniformity (length of 1-3 μm), while the indeterminate nodules of Crotalaria zanzibarica and Trifolium repens contained bacteroids of highly pleomorphism (size
varying from 1 to 5 μm). The enclosed bacteria were isolated from respective nodules,
and twenty slow-growing and nine fast-growing rhizobial isolates were recovered. The slow-growing isolates were classified to the genus Bradyrhizobium based on the 16S rRNA sequences, whereas the fast-growing rhizobia comprise four genera, Neorhizobium, Rhizobium, Cupriavidus and Paraburkholderia. Results of stable isotope
analyses revealed that the seven leguminous species had similar and consistently negative δ15N values in leaves (mean of -1.2 ‰), whereas the values were positive (varying from 3.7 to 7.3 ‰) in the nodules. These values were significantly higher in the
indeterminate nodules than those in the determinate ones. In addition, variations in the values of leaf δ13C (varying from -29 to -34‰) among the seven legumes were
measured, indicating their photosynthetic water use efficiencies were different. This is the first field survey to report the rhizobial diversity and the nutrient relationships of sympatric legume in Taiwan.
Introduction
Nitrogen, one of the most important nutrients for plant growth and reproduction, is often limited in the ecosystems (Vitousek and Howarth, 1991). Some plants evolved symbiosis with bacteria capable of nitrogen fixation and overcame the limitation.
Consequently, the nitrogen availability and the primary productivity of the ecosystems are improved by the symbiotic activity (Vitousek et al., 2002). Because the interaction plays an important role in affecting primary productivity and is of great application in agriculture, the symbiotic relationship has received much attention from the researchers worldwide.
Most leguminous plants are capable of fixing atmospheric N2 via symbiosis with rhizobia (Sprent, 2001, 2009). There were 217 legume species, including 148 native species and 59 exotic species, recorded in Taiwan (Huang, 1993; Wu et al., 2003).
Despite the fact that legume is one of the largest plant families in Taiwan (Huang, 1993) and the importance of the symbiotic relationship in contribution to the nitrogen availability and primary productivity of ecosystems, the bacteria symbionts have been investigated only in few species of leguminous plants in Taiwan (Chen et al., 2000, 2003, 2005; Chen and Lee, 2001; Hung et al., 2005; Huang et al., 2016). Field survey of root-nodulating rhizobia and concomitantly measurements of nutrient of sympatric
leguminous species are lacking in Taiwan.
Xindian River is located in Northern Taiwan. The soil along riverbank is mainly sandy and frequently subjected to disturbances, such as flooding and human activity.
Despite this, leguminous plants are prevalent in this habitat. The nitrogen-fixing symbiosis between the leguminous plants and the soil rhizobia might provide these plants competitive advantages in the nitrogen-poor and arid habitats. Seven leguminous species were commonly observed in this area, including three native species (Alysicarpus vaginalis, Desmodium triflorum and D. heterophyllum) and four exotic species (Crotalaria zanzibarica, Mimosa pudica, Sesbania cannabina and Trifolium repens) (Table 1). These species belong to distantly related legume groups, including
Genistoids (C. zanzibarica), Milletioids (A. vaginalis, D. triflorum, and D.
heterophyllum), Robinioids (S. cannabina), Inverted Repeat-lacking clade (T. repens)
and Mimosoids (M. pudica) (Lewis et al., 2005; Sprent, 2007). In addition, the geographic origins of these legumes are also disparate. A. vaginalis, D. triflorum, and D.
heterophyllum are native species in Taiwan, while C. zanzibarica, S. cannabina, T.
repens and M. pudica, are exotic and originated from Africa, Asia, Europe and America,
respectively (Wu et al., 2003). Among the seven species, the bacterial symbionts of A.
vaginalis, C. zanzibarica, M. pudica and S. cannabina in Taiwan have been reported