Aristolochia yujungiana (Aristolochiaceae): A New Species from Taiwan
Chang-Tse Lu,1) Jenn-Che Wang1,2)
【Summary】
Aristolochia yujungiana C.T. Lu & J.C. Wang, a new species from Taiwan is described and illustrated. Compared to its congeners in Taiwan and neighboring areas, A. yujungiana is similar to A. shimadae, A. heterophylla, A. kaempferi and A. liukiuensis. However, it can be differentiated from A. shimadae by the perianth color and shape, from A. heterophylla by the bracteole shape and limb surface, and from A. kaempferi and A. liukiuensis by the limb-lobe morphology and perianth- tube shape. A morphological description, diagnosis, line drawings, photographs, and conservation status of A. yujungiana as well as a key and a comparison table to morphologically similar species are provided to aid in identification.
Key words: Aristolochia yujungiana, Aristolochiaceae, new species, Taiwan.
Lu CT, Wang JC. 2014. Aristolochia yujungiana (Aristolochiaceae): a new species from Taiwan.
Taiwan J For Sci 29(4):291-9.
1)Department of Life Science, National Taiwan Normal Univ., 88 Tingchow Rd., Sec. 4, Wenshan District, Taipei 11677, Taiwan. 國立台灣師範大學生命科學系,11677台北市文山區汀州路四段88號。
2)Corresponding author, e-mail:biofv017@ntnu.edu.tw 通訊作者。
Received September 2014, Accepted December 2014. 2014年9月送審 2014年12月通過。
研究簡報
台灣產馬兜鈴屬植物之一新種—裕榮馬兜鈴
呂長澤1) 王震哲1,2)
摘 要
本文描述台灣產的新種馬兜鈴屬植物—裕榮馬兜鈴。與台灣及鄰近地區的同屬植物做比較,此新 種與台灣馬兜鈴、異葉馬兜鈴、大葉馬兜鈴與琉球馬兜鈴相似。但是,能以花萼筒的顏色與形狀與台 灣馬兜鈴區分,以小苞片形狀與花萼筒檐部表面毛被差異與異葉馬兜鈴區別,以及用花萼筒檐部裂片 形態與花萼筒形狀與大葉馬兜鈴及琉球馬兜鈴做區分。文中除形態描述及分類特徵摘要外,並提供線 描圖、照片、保育等級以及與其近緣之各物種的檢索表與比較表,以助於物種鑒別之用。
關鍵詞:裕榮馬兜鈴、馬兜鈴科、新種、台灣。
呂長澤、王震哲。2014。台灣產馬兜鈴屬植物之一新種—裕榮馬兜鈴。台灣林業科學29(4):291-9。
INTRODUCTION
Aristolochia L. sensu lato, comprising ca. 450 species distributed in tropical and temperate regions worldwide, is the largest and most diverse genus in the family Aris- tolochiaceae (Wagner et al. 2014). Recent phylogenetic studies of the genus based on morphological and molecular data suggested a subdivision of Aristolochia into 3 subgen- era: Aristolochia, Siphisia (Raf.) Duchartre, and Pararistolochia Huteh & Dalz. (Wanke et al. 2006).
In Taiwan, 5~7 Aristolochia species (Liu and Lai 1976, Hwang 1981, Ma 1989, Hou 1996, Huang et al. 2003, Yang 2007) have been documented. These species can be sort- ed into the 2 subgenera of Aristolochia and Siphisia. Aristolochia foveolata and A. zollin- geriana belong to the subgenus Aristolochia based on their slightly curved or rectilinear tube with a sharply distinct utricle and tube, a 1~3-lobed perianth limb, and a gynostemium with more than 3 lobes, each of which carries a single anther on its outer surface. On the other hand, all the other species are placed in
the subgenus Siphisia due to their U-shaped perianth without a sharply distinct utricle or tube, a 3-lobed limb, a gynostemium with 3 segments, and paired anthers on the outer sur- face of each gynostemium segment.
In the species of the subgenus Siphisia of Taiwan, A. shimadae is a controversial species. This species was first described by Hayata (1916), and some taxonomists (Lai 1973, Liu and Lai 1976) followed Hayata’s concept and treated it as an endemic species, but others regarded it as A. kaempferi (Hwang 1981, Huang et al. 2003), or as a variety of A. kaempferi (Hwang 1988), A. mollis (Ma 1989), or A. heterophylla (Hou 1996). Re- cently, Murata (2006) revised the Japanese Aristolochia, and considered that A. kaempferi was an endemic species of Japan, and A. shi- madae should be treated as a distinct species, distributed in Taiwan and Japan (central Hon- shu, Kyushu, and the Ryukyu Islands) (Mu- rata 2006, Watanabe et al. 2006). Based on Murata’s concept, A. shimadae can be defined as a species in which the floral characters are
conserved with a creamy inside perianth, and a yellow mouth that is rarely purple dotted, but the leaf morphology is polymorphic from triangulate-ovate to lanceolate-ovate (Murata 2006). Recently, Yang (2007) revised the Taiwanese Aristolochia again, and accepted Murata’s (2006) opinions which treat it as A.
shimadae. Accordingly, A. heterophylla and A. kaempferi that are recorded in the Flora of Taiwan 2nd edition (Hou 1996) should be treated as A. shimadae (sensu Murata 2006).
Recently, we received an unknown Aris- tolochia species found in the central part of Taiwan by Mr. Yu-Jung Hung. After a field investigation and comparisons with congeners from Taiwan and neighboring areas, we are certain that this unknown plant should be as- signed to the subgenus Siphisia, and consider it to be a new species. In this paper, we de- scribe this new species. Besides the diagnosis and morphological description, line drawings, photographs, and a key and a comparison table are provided to aid in the identification of it and its close relatives.
Repositories of specimens examined in- clude the following: HAST, Herbarium, Bio- diversity Research Center, Academia Sinica, Taipei, Taiwan; IBSC, South China Botanical Garden, Guangzhou, People’s Republic of China; TAI, National Taiwan Univ., Taipei, Taiwan; TAIF, Taiwan Forestry Research Institute, Taipei, Taiwan; TI, Univ. of Tokyo, Tokyo, Japan; TNS, National Museum of Na- ture and Science, Tsukuba, Japan; TNU, Na- tional Taiwan Normal Univ., Taipei, Taiwan.
TAXONOMIC TREATMENT
Aristolochia yujungiana C.T. Lu & J.C.
Wang, sp. nov.—TYPE: Taiwan. Nantou County, Yuchi Township, Peishankan, elev.
ca. 400 m, 8.II.2008, Chang-Tse Lu 1635 (ho- lotype TAIF; isotype TNU) 裕榮馬兜鈴(Figs.
1, 3B, D, F, H, J, K)
Diagnosis: Species A. shimadae Hayata similis, sed tubo perianthii ore nigro fauce viridi-flavo et atro-purpureo-maculato, semi- nibus late ovatis plano-concavis differt.
Woody, perennial climber. Young branches terete, pubescent. Leaves lamina herbaceous to leathery; petiole 2~5 cm long;
lamina 10~20 cm long, 2.5~3 cm wide, linear to lanceolate, rarely 3~5 lobed; base cordate, with 2 small basal lobes directed downward;
apex acute to acuminate, adaxially green and glabrescent, abaxially grayish-green with pubescence along the veins, hairs appressed;
nerves elevated on abaxial surface. Flower solitary in axils of prophylls of lateral branch- es; pedicel 4~5 cm long, bracteole ovate, ca.
3 mm long, inserted at base of pedicel. Peri- anth tube U-shaped, 3~4 cm long, upper 1/3 more slender, 5~6 mm wide at utricle, outer surface creamy, pubescent, and inner surface smooth and yellowish-green with dark-purple speckles; mouth subspherical to elliptical, ca.
5×8 mm, dark-purple sometimes with yel- low dots, annulus distinct; limb obtriangular in front view, ca. 2 cm wide, deeply 3-lobed, lobes spread, recurved apically, dark-purple, sparsely pubescent. Stamens 6, adnate to style column, anthers oblong, ca. 2 mm long.
Gynostemium ca. 4 mm long; stigmatic lobes 3, lobe triangular; ovary inferior, cylindrical, ca. 8 mm long, pubescent. Capsules hairy, el- lipsoid to oblong, with 6 ridges, 4~5 cm long, 2.5~3 cm in diameter, and dehiscent from apex. Seeds flat, widely ovate, plano-concave, 5.2~6 mm long, and 5.2~5.5 mm wide.
Distribution: Aristolochia yujungiana is endemic to Taiwan and so far found only in the type locality, along the forest edge in low- elevation mountain areas of central Taiwan (Fig. 2).
Phenology: Aristolochia yujungiana blooms in December to March and fruits in March to August.
Etymology: The specific epithet is derived from the forename of Mr. Yu-Jung Hung, who discovered this new species.
Conservation state: Human activities like agriculture and reclamation have largely altered low-elevation mountain areas of Tai- wan, especially in western Taiwan. The cur- rent population of A. yujungiana of fewer than 50 individuals covers an area of about 100 m2. We consider the current meager distribution of this new species a likely remnant of a more-
widespread population in the past. According to the International Union for Conservation of Nature (IUCN) threat categories (IUCN 2012), this species should be categorized as critically endangered (CR): B1ab (iii,v).
Taxonomic notes: By comparison to the congeners from Taiwan and neighboring ar- eas, we found that A. yujungiana most closely resembles A. shimadae Hayata, especially the population that is distributed in eastern Taiwan. They both have linear to lanceolate Fig. 1. Aristolochia yujungiana (from the holotype). A, habit; B, abaxial surface of the leaf;
C, front view of the flower; D, bisection of the perianth tube; E, gynostemium: E1, pistillate stage; E2, staminate stage showing anther dehisced and stigma folded; F, fruit; G, seeds, adaxial view (left) and abaxial view.
laminae, and the base is cordate with 2 small lateral lobes directed downward and acute at the apex. But A. yujungiana can be dif- ferentiated from A. shimadae by the flower color and shape. For example, the color of the mouth of the perianth tube is dark-purple and sometimes with yellow dots in A. yujungiana, while it is yellow in A. shimadae; and the di- ameter of tube of A. yujungiana is more slen- der (3~4 mm) than that of A. shimadae (5~7 mm) (Fig. 3).
Aristolochia yujungiana also resembles A. heterophylla from China. Compared to the type specimen and protologue of A. hetero- phylla (see Hemsley 1891), we found that the former can be distinguished from the latter by the bracteole shape (ovate vs. rounded) and limb surface (sparsely pubescent vs. com- pactly papillose).
Additionally, A. yujungiana is also simi- lar to A. kaempferi and A. liukiuensis of Japan in flower color, but the perianth tube shape and limb morphology differ from those of the latter 2 species. Aristolochia yujungiana has a slender tube at 3~4 mm in diameter, while
A. kaempferi and A. liukiuensis has thicker tubes at 5~7 mm in diameter. The limb of A.
yujungiana is deeply 3-lobed, the lobes are spread, recurved apically, and obtriangular in front view, but those of A. kaempferi and A.
liukiuensis are shallowly 3-lobed, the lobes are not spread, are incurved apically, and are widely obovate in front view. Based on these differences, we treated it as a new species.
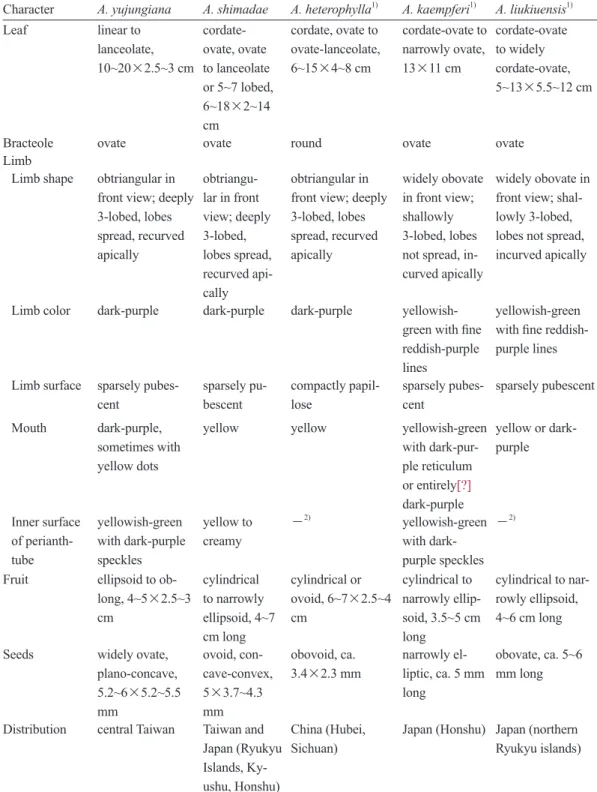
Detailed comparisons between A. yujungiana and its close relatives are given in Table 1.
Key to the Aristolochia yujungiana and its close relatives
1. Leaf lamina linear to lanceolate ...2 2.Perianth-tube 3~4 mm in diameter; mouth
dark-purple, sometimes with yellow dots ...A. yujungiana 2.Perianth-tube 5~7 mm in diameter; mouth
yellow ... A. shimadae 1.Leaf lamina widely cordate-ovate to nar- rowly ovate ...3 3.Perianth limbs deeply 3-lobed, lobes
spread, recurved apically, obtriangulate in front view ...4 Fig. 2. Geographical distribution of Aristolochia yujungiana (★).
4.Limb papillose; bracteole round ...
...A. heterophylla 4.Limb sparsely pubescent; bracteole
ovate ... A. shimadae 3.Perianth limbs shallowly 3-lobed, lobes
not spread, incurved apically, widely obovate in front view ...5
5.Perianth tube mouth > 7 mm in diameter ... A. liukiuensis 5.Perianth tube mouth < 7 mm in
diameter ...A. kaempferi
Additional specimens examined:
Aristolochia heterophylla Hemsley. CHI- NA. Hubei, Ichang, October 1887, A. Henry 3493, Fl. (syntype K photo!), Hupeh, March 1889, A. Henry 6417, Fr (syntype K photo!), Hupeh, Fang, March 1889, A. Henry 6417A, Fr. (syntype K photo!), Hupeh, March 1889, A. Henry 6490, Fr. (syntype K, photo!), Hu- peh, A. Henry 4665, Fl. (syntype NY photo!);
Ichang, Patung Dist., fl., March 1886, A.
Henry 642 (K photo!).
Fig. 3. Comparison of the floral morphology of Aristolochia shimadae (A, C, E, G, I, L) and A.
yujungiana (B, D, F, H, J, K). A and B, habitat; C and D, leaf variations; E and F, front view of flower (note the different colors on the mouth of the perianth tube; G and H, bisection of the perianth tube showing the gynostemium and inner surface of the perianth tube; I and J, fruit; K and L, seeds.
Table 1. Comparisons of Aristolochia yujungiana with its 4 morphologically close relatives Character A. yujungiana A. shimadae A. heterophylla1) A. kaempferi1) A. liukiuensis1) Leaf linear to
lanceolate, 10~20×2.5~3 cm
cordate- ovate, ovate to lanceolate or 5~7 lobed, 6~18×2~14 cm
cordate, ovate to ovate-lanceolate, 6~15×4~8 cm
cordate-ovate to narrowly ovate, 13×11 cm
cordate-ovate to widely cordate-ovate, 5~13×5.5~12 cm
Bracteole ovate ovate round ovate ovate
Limb
Limb shape obtriangular in front view; deeply 3-lobed, lobes spread, recurved apically
obtriangu- lar in front view; deeply 3-lobed, lobes spread, recurved api- cally
obtriangular in front view; deeply 3-lobed, lobes spread, recurved apically
widely obovate in front view;
shallowly 3-lobed, lobes not spread, in- curved apically
widely obovate in front view; shal- lowly 3-lobed, lobes not spread, incurved apically
Limb color dark-purple dark-purple dark-purple yellowish- green with fine reddish-purple lines
yellowish-green with fine reddish- purple lines Limb surface sparsely pubes-
cent
sparsely pu- bescent
compactly papil- lose
sparsely pubes- cent
sparsely pubescent
Mouth dark-purple, sometimes with yellow dots
yellow yellow yellowish-green with dark-pur- ple reticulum or entirely[?]
dark-purple
yellow or dark- purple
Inner surface of perianth- tube
yellowish-green with dark-purple speckles
yellow to creamy
-2) yellowish-green with dark- purple speckles
-2)
Fruit ellipsoid to ob- long, 4~5×2.5~3 cm
cylindrical to narrowly ellipsoid, 4~7 cm long
cylindrical or ovoid, 6~7×2.5~4 cm
cylindrical to narrowly ellip- soid, 3.5~5 cm long
cylindrical to nar- rowly ellipsoid, 4~6 cm long Seeds widely ovate,
plano-concave, 5.2~6×5.2~5.5 mm
ovoid, con- cave-convex, 5×3.7~4.3 mm
obovoid, ca.
3.4×2.3 mm
narrowly el- liptic, ca. 5 mm long
obovate, ca. 5~6 mm long
Distribution central Taiwan Taiwan and Japan (Ryukyu Islands, Ky- ushu, Honshu)
China (Hubei, Sichuan)
Japan (Honshu) Japan (northern Ryukyu islands)
1)Comparison characters of A. heterophylla were extracted from the prolotogue of A. heterophylla (Hemslay 1891), and those of A. kaempferi and A. liukiuensis were extracted from the Flora of Japan (Murata 2006).
2)−, Information unavailable.
Aristolochia kaempferi Willd. JAPAN.
HONSHU: Manatsuru Peninsula, Manatsuru- machi, Ashigara-shimo-gun, elev. ca. 70 m, fr., June 1958, T. Kawasaki 6499 (HAST);
Shizuoka Pref., Numazu-shi, elev. ca. 80 m, fl., 29 April 1991, F. Konta & K. Okada 69 (TNS); Numazu-shi, elev. ca. 150 m, fl., 5 June 1991, F. Konta et al. 225 (TNS); Shimizu City, elev. ca. 250 m, 17 June 1988, F. Konta 17046 (TNS); Shimoda City, elev. ca. 30 m, 28 May 2003, F. Konta et al. 23090 (TNS);
Chiba Pref., Kanto Distr., Amatsukominato- machi, Mt. Kiyosumi, fr., 5 June 1990, Y.
Tateishi et al. 15787 (IBSC); Miyazaki Pref., Koyu-gun, Kawanami-machi, elev. ca. 200 m, fr., 27 May 1983, Idzumi & Togashi s.n.
(TI); Prov. Ohsumi, Ohdomari, Satacho, fl., 29 April 1962, Hatusima & Sako 26891 (TI); Prov. Satsuma, Kawashiri, near Mt.
Kaimon, fr., 18 May 1946, Togashi s.n. (TI).
Aristolochia liukiuensis Hatusima. JA- PAN. Kagoshima Pref., Uken-Son (Uken Vil- lage), Ooshima-gun (Ooshima District), elev.
ca. 330 m, fl., 12 March 2002, Noshiro et al.
D488 (TI); Kagoshima Pref., Is. Tokunoshi- ma, Oshima-gun, Isen-machi, Kozima, fl., elev. ca. 100 m, 10 February 1982, Murata
& Endo 79 (TI); Isl. Amami-Oshima, near Kosyuku, fl. & fr., elev. ca. 100 m, 22 March 1958, Hatusima & Sako 21728 (TI); Isl.
Okinawa, fl., 5 March 1998, Takushi s.n.
(TI); Okinawa Pref., Mt. Yonaha, Kunigami- son, Kunigami-gun, fl., 13 December 1992, Kobayashi 1895 (TI); Kanna, Ginoza-son, Kunigami-gun, Okinawa-jima, fl., March 1983, Nishi & Ishizuka 158 (TI).
Aristolochia shimadae Hayata. JAPAN.
OKINAWA: Miyako-jima: Gusukube-cho W. of Aragusuku, elev. ca. 50 m, fl. & fr., 10 March 1978, Tateishi 4012 (TI); Isl. Ishigaki, upper stream of Miyara-gawa, E. foot of Mt.
Omoto-dake, elev. 50~100 m, fl. & fr., 28 March 1973, H. Koyama et al. 349, 350 (TNS);
Yaeyama-gun, Taketomi-cho, Isl. Itiomote, Funaura, elev. 10~20 m, fl., 16 April 1998, K. Yonekura et al. 98051 (HAST). TAIWAN.
TAIPEI: Yangmingshan, Chingshan Rd., elev.
ca. 240 m, 3 May 1996, S. C. Wu 977 (TAIF);
Kuanyinshan, elev. ca. 616 m, 4 February 1973, J. F. Wang s.n. (TNU). HSINCHU:
Shimpo, Shinchikucho, 15 December 1915, Y.
Shimada s.n. (holotype: TI, photo!; merotype:
TAIF!); Shintiku, Shinpo, fr., 20 September 1917, Shimada s.n. (TI, fruit voucher of A.
shimadae in Icon. Pl. Form. 8: 110, 1919);
Kuanhsi Town, No. 16 County Rd. 6.3 km, Minan Bridge-Nanho, elev. ca. 102 m, 28 June 2003, C. M. Wang 7005 (TNM); Paoshan Res- ervoir, elev. ca. 150 m, 26 March 2002, S. C.
Wu 2438 (HAST); Kuanwu, elev. 1950~2000 m, 16 May 2000, S. W. Chung 2371 (TAIF).
MIAOLI: Kungkuan, elev. ca. 120 m, 4 April 1973, I. S. Chen 2929 (TAI); Hsihu Hsiang, Chinshih Village, elev. ca. 138 m, 21 February 2007, C. L. Yang 460 (TNU). TAICHUNG: en route from Chunghsing Ling to Takeng, elev.
ca. 570 m, 2 November 1986, C.-I Peng 9981, 9966 (HAST); Ssuyuan, elev. ca. 1900 m, 19 April 1997, S. Y. Lu s.n. (TAIF); Wuling Farm, elev. ca. 2000 m, 4 July 2000, S. W. Chung 2645 (TAIF); Hohuanshan Tunnel to Lishan, elev. ca. 2482 m, 7 June 2002, C. K. Yang 92 (TNM). NANTOU: Meifeng to Tsuifeng, elev.
2000~2300 m, 21 May 2005, J. H. Lii 1181 (TAI); Tunyuan, elev. 1900~2100 m, 22 April 2000, C. H. Chen 3201 (TAIF); Hohuanshan, elev. ca. 3000 m, 19 May 1997, C. C. Hsu 254 (TAIF). HUALIEN: Chingshuishan, elev.
1500~2400 m, 25 July 1986, K. C. Yang 12861 (TAI). TAITUNG: Tulanshan, elev. ca. 1100 m, 8 May 2002, S. W. Chung 5245 (TAIF).
ACKNOWLEDGEMENTS
We heartily thank Dr. Qiner Yang of the South China Botanical Garden, China, for
improving the Latin diagnosis, and Dr. Aleck Tsung-Yu Yang of the National Museum of Natural Science, Taiwan, who helped us to get photographs of the type specimens of A.
heterophylla from the herbarium of Royal Botanic Gardens (K), KEW, U.K. England.
Authors also thank the English editor and two anonymous reviewers for kindly improving our manuscript.
LITERATURE CITED
Hemsley WB. 1891. All enumeration of all the plants known from China proper, Formosa, Hainan, Corea, the Luchu Archipelago, and the Island of the Hongkong, together with their distribution and synonymy, part X. J Linn Soc Bot 26:361-2.
Hou D. 1996. Aristolochiaceae. In: Boufford DE, et al., editors. Flora of Taiwan, 2nd ed., Vol.
2. Taipei, Taiwan: Editorial Committee of the Flora of Taiwan. p 636-42.
Huang SM, Kelly LM, Gilbert MG. 2003.
Aristolochia. In: Wu ZY, et al., editors. Flora of China, Vol. 5. Beijing, China: Science Press and St. Louis, MO: Missouri Botanical Garden Press. p 258-69.
Hwang SM. 1981. Materials for Chinese Aris- tolochia. Acta Phytotaxon Sin 19(2):222-31. [in Chinese with English summary].
Hwang SM. 1988. Aristolochia. In: Kiu HS, Ling YR, editors. Flora Reipublicae Popularis Sinicae, Tomus 24. Beijing, China: Science Press. p 199-245.
International Union for Conservation of Na- ture (IUCN). 2012. IUCN red list categories
and criteria: Version 3.1. 2nd Edition. Gland, Switzerland and Cambridge, UK: IUCN Spe- cies Survival Commission. 32 p.
Lai MJ. 1973. A taxonomic revision of Raffle- siaceae, Balanophoraceae and Aristolochiaceae of Taiwan. [Master’s thesis]. Taipei, Taiwan:
National Taiwan Univ. 58 p.
Liu TS, Lai MJ. 1976. Aristolochiaceae. In:
Li HL, et al., editors. Flora of Taiwan, Vol. 2.
Taipei, Taiwan: Epoch Publishing. p 571-6.
Ma JS. 1989. A revision of Aristolochia Linn.
from E. & S. Asia. Acta Phytotaxon Sin 27:
352-6.
Murata J. 2006. Aristolochia. In: Iwatsuki K, et al., editors. Flora of Japan, Vol. 2a. Tokyo, Japan: Kodansha. p 366-8.
Wagner ST, Hesse L, Sandrine I, Samain MS, Bolin J, Maass E, et al. 2014. Major trends in stem anatomy and growth forms in the perianth-bearing Piperales, with special focus on Aristolochia. Ann Bot-Lond 113:1139-54.
Wanke S, Gonzalez F, Neinhuis C. 2006.
Systematics of pipevines: combining morpho- logical and fast-evolving molecular characters to investigate the relationships within subfam- ily Aristolochioideae (Aristolochiaceae). Int J Plant Sci 167(6):1215-27.
Watanabe K, Kajita T, Murata J. 2006.
Chloroplast DNA variation and geographical structure of the Aristolochia kaempferi group (Aristolochiaceae). Am J Bot 93:442-53.
Yang CL. 2007. A Taxonomic study of Aristo- lochia L. (Aristolochiaceae) in Taiwan [mas- ter’s thesis]. Taipei, Taiwan: National Taiwan Normal Univ. 91 p. [in Chinese with English summary].