Air Temperature Influenced the Vocal Activity of Birds in a Subtropical Forest in Southern Taiwan
Chun-Chieh Liao,
1)Bao-Sen Shieh,
1)Chao-Chieh Chen
1,2)【 Summary】
Singing activities of birds in the early spring are mainly controlled by the photoperiod and other environmental factors such as temperature, food, and rainfall. In this study, we monitored the vocal activity of a forest bird community in the Shan-Ping Forest Ecological Garden, southern Taiwan, to explore the influence of ambient temperature on the vocal behavior of birds. An acous- tic monitoring system was set up to record bird sounds throughout the morning hours in March of 2011 and 2012, and the air temperature was recorded on the hour by a data logger. Recorded soundscape files were sampled, transcribed, and analyzed. Results showed that both the number of bird species and vocal activity detected in 10-min intervals sampled from 6 morning hours were positively correlated with the air temperature in both years. At the species level, the relationship between vocal activity and air temperature was found to be significant for 6 of 16 common resident species. Furthermore, air temperature at 10:00 or 11:00 appeared to be a better predictor of vocal activities of birds in the study plot than those measured from 06:00 to 09:00. This study illustrated that the vocal activity of a forest bird community was strongly influenced by air temperature and closely fluctuated with the dynamics of cold fronts. We propose that this relationship could be developed into a bioassay to examine the influence of global warming on spring singing of birds through autonomous recording.
Key words: autonomous recording, bird community, cold front, singing behavior, soundscape.
Liao CC, Shieh BS, Chen CC. 2018. Air temperature influenced the vocal activity of birds in a subtropical forest in southern Taiwan. Taiwan J For Sci 33(4):291-304.
1)Department of Biomedical Science and Environmental Biology, Kaohsiung Medical University, 100 Shih-Chuan 1st Rd., San Ming District, Kaohsiung 80708, Taiwan. 高雄醫學大學生物醫學暨環境生
物學系,80708高雄市三民區十全一路100號。
2)Corresponding author, e-mail:chen5123@kmu.edu.tw 通訊作者。
Received April 2018, Accepted August 2018. 2018年4月送審 2018年8月通過。
研究報告
南臺灣亞熱帶森林氣溫變化對鳥類鳴叫行為之影響
廖俊傑
1)謝寶森
1)陳炤杰
1,2)摘 要
鳥類在繁殖季初期的鳴唱活動除了會受到光週期的調控之外,溫度、食物量及降雨量也都是重要 的環境影響因子。本研究在扇平森林生態科學園區藉由自動錄音監測系統探討早春氣溫變化對鳥類鳴 叫行為的影響。我們在2011年與2012年的三月份,錄下每天清晨到中午的鳥類鳴叫聲,並放置溫度監 測器進行溫度資料取樣。我們選取寒流來襲前後期間約兩週的錄音資料,並以人工方式進行轉錄。野 外錄音結果顯示,無論是鳥種數或鳴叫頻度皆與氣溫呈顯著正相關。16種當地常見的鳥類中,有6種的 鳴叫頻度與氣溫變化有顯著正相關。此外,本研究也發現每日上午10:00或11:00的溫度較上午06:00到 09:00的溫度更適合用來預測鳥類鳴叫頻度受氣溫變化的影響。綜合上述,鳥類群聚的鳴叫行為確實會 受到溫度變化的影響,且寒流所帶來的氣溫波動與鳥類鳴叫行為有共振的效應。我們認為自動錄音系 統非常適合用來探索氣候變遷對早春鳥類繁殖鳴叫行為的影響。
關鍵詞:自動錄音、鳥類群聚、寒流、鳴叫行為、聲景。
廖俊傑、謝寶森、陳炤杰。2018。南臺灣亞熱帶森林氣溫變化對鳥類鳴叫行為之影響。台灣林業科學 33(4):291-304。
INTRODUCTION
In spring, most adult males begin to sing for territorial defense and to attract mates for the coming breeding season (Morse 1970, Catchpole and Slater 1995). Although spring singing of birds is mainly controlled by intrin- sic mechanisms triggered by photoperiodic changes, fine-scale variations are mediated by other environmental factors such as tempera- ture, food resources, and rainfall (Slagsvold 1977, Ball 1999, Thomas 1999, Wingfield et al. 2003, Dawson 2008). The environmental factor of temperature has been more often studied than other environmental factors.
Garson and Hunter (1979) found a strong relationship between the overnight minimum temperature and singing activity the following morning in the Eurasian Wren (Troglodytes troglodytes) and Great Tit (Parus major). In addition, Higgins (1979) illustrated that the
vocal activity of the Song Thrush (Turdus philomelos) was significantly proportional to air temperature in the early morning. These findings indicate that birds might reduce their singing activity in order to save energy during times of low temperature, because a consider- able amount of energy and time is required for singing (Thomas 1999, Gil and Gahr 2002).
Previous studies exploring the influence of temperature on the vocal behavior of birds mainly focused on single bird species (Garson and Hunter 1979, Higgins 1979). However, it would be more representative if this rela- tionship could be applied to the entire bird community of a given ecosystem. Hence, we monitored changes in the vocal activity of a bird community in a subtropical forest to ex- amine the effect of temperature on the vocal behaviors of birds. This study was carried out
in early spring when cold fronts still prevailed and the fluctuation in temperature remained high. This ensured that a greater range of temperatures was recorded, and its effect on the vocal activity of birds could be clearly de- marcated.
This study was feasible through the use of an acoustic monitoring system, which has been used as a tool for estimating bird rich- ness and abundance in other studies (Hobson et al. 2002, Acevedo and Villanueva-Rivera 2006, Brandes 2008, Dawson and Effort 2009). Acoustic monitoring systems have the potential to operate continually around the clock (Acevedo and Villanueva-Rivera 2006, Chen et al. 2011). Therefore, we could moni- tor changes in the vocal activities of birds across days, and even during the entire period of a cold front.
In addition, 80~94% of birds detected during point counts are through sound dur- ing the breeding seasons (Best 1981, Scott et al. 1981, DeJong and Emlen 1985), and it is even higher in forest habitats (Brewster and Simons 2009, Celis-Murillo et al. 2012). Sim- ilarity indices of point count surveys between personnel and autonomous recording usually reach 80~95% (Hobson et al. 2002, Celis- Murillo et al. 2009). Therefore, the vocal activity of a bird community can substantially be constructed using an acoustic monitoring system with continuous recording.
The particular objectives of this study were to: 1) document the vocal activity of a forest bird community; 2) relate vocal activity with air temperature; and 3) establish an automated acoustic system as a standard bioassay of envi- ronmental effects on bird reproductive cycles.
MATERIALS AND METHODS Study area
The Shan-Ping Forest Ecological Garden,
a remote research station of the Taiwan For- estry Research Institute, is located in southern Taiwan (22°58’12’’N, 120°41’24’’E), with an annual average temperature of 20.6℃ and an annual rainfall of 3500 mm (Lu et al. 2009).
Broadleaf forests and montane streams char- acterize the site, and 134 bird species have been documented (King et al. 1990); howev- er, the species richness apparently decreased in a recent survey (Tsai et al. 2015). The main road to Shan-Ping Forest was heavily damaged by Typhoon Morakot in 2009 and tourism activities were not permitted during the study period. It turned out that Shan-Ping Forest is a very profitable site for acoustic re- cording research since tourist interference is nearly negligible.
Field procedures
We set up an acoustic monitoring sys- tem in the Shan-Ping Forest. The unit con- sisted of a mini-computer (ASUS, AS-D692, Taipei City, Taiwan), a 3-channel stereo microphone mixer (Rolls, MX54S Promix Plus, Salt Lack City, UT, USA), a micro- phone (Shure, WL183, Niles, IL, USA), and a temperature and light data logger (HOBO Pendant temp/light, part no. UA-002-64, Mi- crodaq.com (Ltd), Contoocook, NH, USA).
The acoustic monitoring system was located 10 m from a trail that runs beside a stream through a riparian forest. A cable line was pulled 15 m further into the forest, and the microphone was fixed under an aluminum roof that was tied to a tree trunk at a height of 3 m above the ground. The acoustic moni- toring system automatically recorded files (WAV) at 10-min intervals, from 05:25 to 11:25 every day, through recording software (High Criteria, Total Recorder, Richmond Hill, Ontario, Canada). In a preliminary test, small passerine birds such as Morrison’s Ful- vetta (Alcippe morrisonia) could be clearly
recorded at a distance of approximately 20 m. Larger species such as the Gray Treepie (Dendrocitta formosae) could be recorded as far as 100 m away from the recorder (Chen at al. 2011). The air temperature was record- ed on the hour by the HOBO pendant data logger, and data were periodically down- loaded.
Data analyses
In the lab, we selected soundscape files recorded during 14~28 March 2011 and 10~27 March 2012, each period of which included 2 cold fronts. We retrieved a section of 10-min recording from each morning hour (from 06:05~06:15 to 11:05~11:15), adding up to a total of 60 min of recording segments per day. We took a 1-min interval as a work- ing unit, and species detected by songs or calls each minute were transcribed onto a data sheet. Then, the number of different bird species recorded in the 10-min interval was enumerated. The occurrence of vocalizations of different bird species was summed for each minute, and the grand sum for the 10-min pe- riod was taken as a measure of vocal activity.
It represents the relative frequency of vocal events of all species occurring in the 10-min period. The air temperature (℃) was retrieved from the HOBO data logger, and the tempera- ture taken at 06:00 was used to represent the temperature for that hour, and the same for the other 5 h.
We used the GLMM procedure of SAS (SAS, Cary, NC, USA) with HOUR (sam- pling unit) as a repeated-measure to explore the relationship between air temperature and the number of bird species recorded in a 10-min interval each morning hour throughout the study periods of 2011 and 2012. The same analysis was repeated for the relationship between air temperature and vocal activity of birds detected within a 10-
min interval for 2011 and 2012, separately.
A simple linear regression was applied to formulate the relationship between the aver- age morning air temperature with either the average number of bird species detected or vocal activity on a daily basis. This analysis was separately conducted for the 2011 and 2012 datasets. Following this method, we formulated the relationship between aver- age air temperature and vocal activity for those common bird species that appeared in at least 25 d of recording, considering both 2011 and 2012. Finally, to determine either the maximum or minimum temperature that most influenced the vocal behavior of birds, we used temperatures from different morn- ing hours as the day’s temperature and redid the regression analyses.
RESULTS
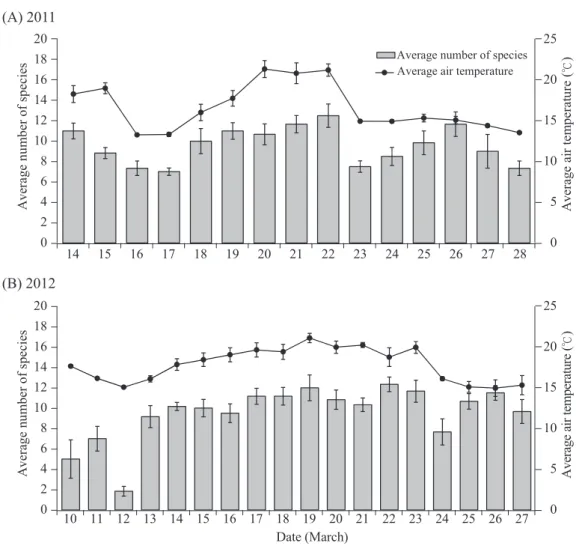
The number of bird species recorded in 10-min intervals of the 6 morning hours was positively correlated with air temperature in both 2011 (F1,83 = 21.71, p < 0.0001; Fig. 1A) and 2012 (F1,101 = 15.04, p < 0.001; Fig. 1B).
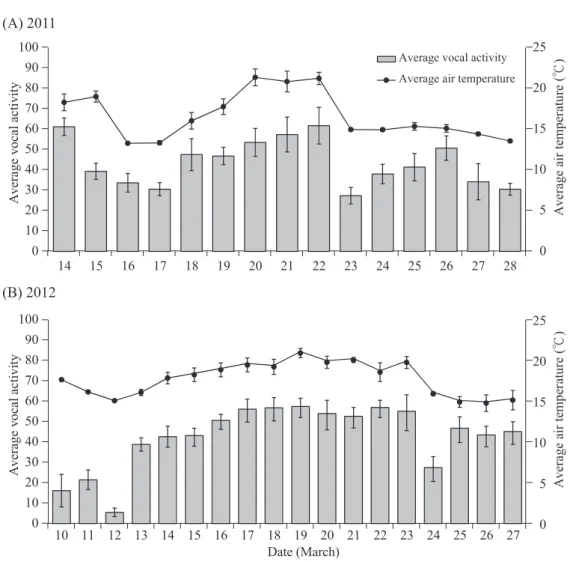
In addition, the vocal activity of birds as a whole in the community also showed a signif- icant positive correlation with air temperature in both 2011 (F1,83 = 29.22, p < 0.0001; Fig.
2A) and 2012 (F1,101 = 31.27, p < 0.0001; Fig.
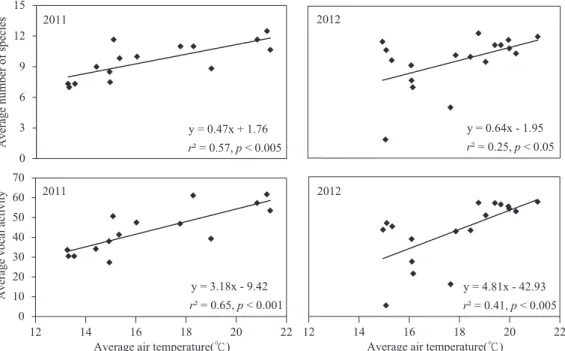
2B). Figure 3 depicts such a linear relation- ship on a daily basis. The average morning air temperature was highly correlated with the average number of species and vocal activity of birds recorded in a 10-min interval of the 6 morning hours in both 2011 and 2012 (Fig. 3).
We found 30 bird species (excluding nocturnal owls) from 33 days of acoustic recording in both 2011 and 2012. Vocal activity showed a significant positive cor- relation with air temperature in 6 of the 16 common bird species that appeared in at least
Fig. 1. The number of bird species recorded in a 10-min interval was significantly correlated with the air temperature (℃) of the 6 morning hours in both A) 2011 (F1,83 = 21.71, p < 0.0001) and B) 2012 (F1,101 = 15.04, p < 0.001) at Shan-Ping Forest Ecological Garden, southern Taiwan.
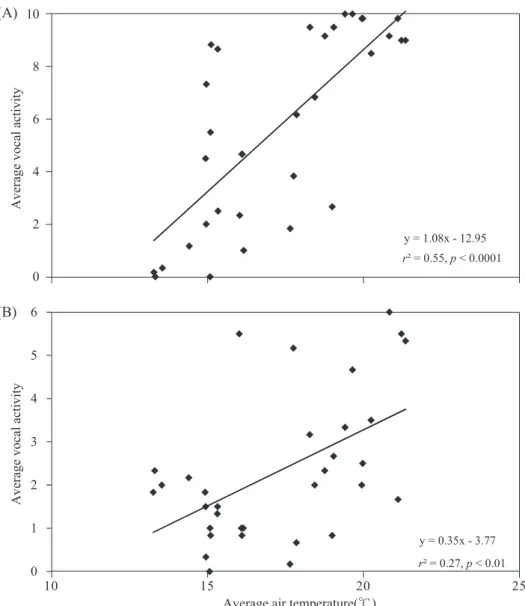
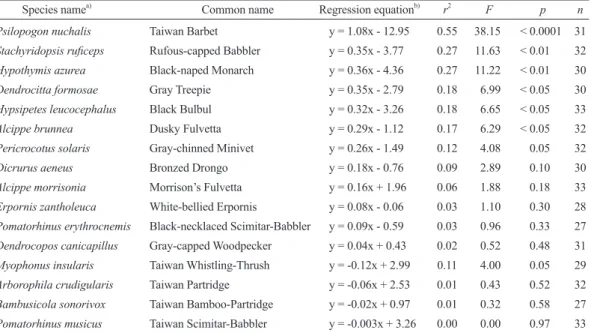
25 d of recording (Table 1). These included Psilopogon nuchalis (r2 = 0.55, p < 0.0001;
Fig. 4A), Stachyridopsis ruficeps (r2 =0.27, p < 0.01; Fig. 4B), Hypothymis azurea (r2 = 0.27, p < 0.01), Dendrocitta formosae (r2 = 0.18, p < 0.05), Hypsipetes leucocephalus (r2
= 0.18, p < 0.05), and Alcippe brunnea (r2 = 0.17, p < 0.05).
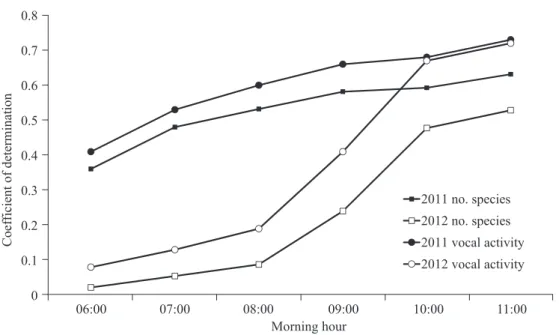
When the air temperature on the hour was replaced by the temperature from dif- ferent morning hours, the coefficient of de-
termination (r2) of the regression analyses increased from 06:00 to 11:00 (Fig. 5). The value of r2 reached a plateau at 10:00. Al- though both years contained 2 cold fronts in the sampled period, the fluctuation of tem- perature in 2011 (coefficient of variation (CV)
= 17.44) was greater than that in 2012 (CV
= 11.59). The influence of air temperature on the vocal behavior of birds in 2011 appeared to be stronger and thus resulted in a higher r2 than that in 2012 (Fig. 5).
0 5 10 15 20 25
0 2 4 6 8 10 12 14 16 18 20
Average air temperature (℃)
Average number of species
14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
Date (March) (A) 2011
0 5 10 15 20 25
0 2 4 6 8 10 12 14 16 18 20
Average air temperature (℃)
Average number of species
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 (B) 2012
Average number of species Average air temperature
Fig. 2. The vocal activity of birds recorded in a 10-min interval was significantly correlated with the air temperature (℃) of 6 morning hours in both A) 2011 (F1,83 = 29.22, p < 0.0001) and B) 2012 (F1,101 = 31.27, p < 0.0001) at Shan-Ping Forest Ecological Garden, southern Taiwan.
DISCUSSION
In this study, the vocal activity of a for- est bird community was found to be highly influenced by the air temperature during waves of cold fronts in early spring of both 2011 and 2012. Such a relationship was found to be stronger in 2011 when temperature fluctuations with the dynamics of cold fronts were greater. These results indicated that air temperature has a clear impact on bird vocal
behavior at the community level. Our results confirmed that air temperature can affect the vocal activity of birds as found in other stud- ies (Garson and Hunter 1979, Reid 1987, Thomas 1999, Hasan and Badri 2016). Gar- son and Hunter (1979) reasoned that having spent much energy in maintaining the body temperature on cold nights, birds would show a greater demand to feed the following morn- ing which would therefore lead to a decline in singing activity. Indeed, singing and foraging
0 5 10 15 20 25
0 10 20 30 40 50 60 70 80 90 100
Average vocal activity
Average vocal activity Average air temperature
Average air temperature (℃)
14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
0 5 10 15 20 25
0 10 20 30 40 50 60 70 80 90 100
Average air temperature (℃)
Average vocal activity
Date (March)
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 (A) 2011
(B) 2012
y = 4.81x - 42.93 r² = 0.41, p < 0.005
12 14 16 18 20 22
Average air temperature(℃) 2012
y = 0.47x + 1.76 r² = 0.57, p < 0.005 0
3 6 9 12 15
Average number of species 2011
y = 0.64x - 1.95 r² = 0.25, p < 0.05 2012
y = 3.18x - 9.42 r² = 0.65, p < 0.001 0
10 20 30 40 50 60 70
12 14 16 18 20 22
Average vocal activity
Average air temperature(℃) 2011
Fig. 3. The simple linear regression depicts the relationship between the daily average number of species and vocal activity of birds recorded in 10-min intervals of 6 morning hours with average morning air temperatures (℃) in both 2011 (n = 15 d) and 2012 (n = 18 d) at Shan-Ping Forest Ecological Garden, southern Taiwan.
are mutually exclusive or close to being so, and thus a tradeoff might occur between the amount of time that a bird can spend on either behavior (Thomas 1999).
The annual average temperature at our study site was 20.6℃, and the minimum, mean, and maximum temperatures during the study period (March 2011) were 12.6, 16.6, and 25.9℃. These values are much higher compared to those in temperate zones. As a re- sult, the energy spent on maintaining the body temperature at our study site may have been far less than that spent by temperate birds.
Therefore, we considered that the strong posi- tive relationship between the vocal activity of birds and air temperature at our study site might have resulted from spring singing in- stead of energy maintenance. At our study site, birds usually resumed singing immediately after a cold spell had ended. With consecutive
days of sunny weather and high temperatures in late winter and early spring, unleashed singing abruptly increased. It seems that birds regard such change as the arrival of spring.
The singing continued until the next cold front arrived, and the sudden drop in temperature once again made the forest as quiet as during a normal winter day. The dramatic fluctuation of temperature during waves of cold fronts in early spring clearly illustrates the effect of temperature on bird vocal behavior. A likely explanation for this pattern is spring singing.
Breeding males have to sing as early as pos- sible in order to hold a territory (Garson and Hunter 1979), because the security of a terri- tory will contribute greatly to their upcoming reproductive success.
Although the initiation of breeding in birds is mainly controlled by photoperiod changes in early spring (Wingfield et al. 1980,
y = 1.08x - 12.95 r² = 0.55, p < 0.0001 0
2 4 6 8 10
Average vocal activity
y = 0.35x - 3.77 r² = 0.27, p < 0.01 0
1 2 3 4 5 6
5 2 0
2 5
1 0
1
Average vocal activity
Average air temperature(℃) (B)
(A)
Fig. 4. The average vocal activity in a 10-min interval was significantly correlated with the average air temperature (℃) of the 6 morning hours for (A) Psilopogon nuchalis (r2 = 0.55, n = 31, p < 0.0001) and (B) Stachyridopsis ruficeps (r2 = 0.27, n = 32, p < 0.01). Data from both 2011 and 2012 were pooled for the analyses.
Wingfield and Farner 1993, Hau et al. 1998), environmental factors, such as temperature, food, and rainfall, were proven to play impor- tant roles in triggering reproduction (Strain and Mumme 1988, Thomas 1999, Hau 2001, Leit- neret al. 2003, Lehmann et al. 2012). Wingfield et al. (2003) found that the breeding cycles of the mountain White-crowned Sparrow (Zono-
trichia leucophrys oriantha) are consistent among years and controlled by the photoperiod length, but a slight difference in the timing of breeding is mainly attributed to climate varia- tions. Our results showed that the vocal activ- ity of birds at Shan-Ping Forest accordingly fluctuated with air temperatures in early spring.
This implies that temperature might play a cru-
Table 1. The relationship between air temperature (℃) and vocal activity for 16 common bird species at Shan-Ping Forest Ecological Garden
Species namea) Common name Regression equationb) r2 F p n
Psilopogon nuchalis Taiwan Barbet y = 1.08x - 12.95 0.55 38.15 < 0.0001 31 Stachyridopsis ruficeps Rufous-capped Babbler y = 0.35x - 3.77 0.27 11.63 < 0.01 32 Hypothymis azurea Black-naped Monarch y = 0.36x - 4.36 0.27 11.22 < 0.01 30 Dendrocitta formosae Gray Treepie y = 0.35x - 2.79 0.18 6.99 < 0.05 30 Hypsipetes leucocephalus Black Bulbul y = 0.32x - 3.26 0.18 6.65 < 0.05 33
Alcippe brunnea Dusky Fulvetta y = 0.29x - 1.12 0.17 6.29 < 0.05 32
Pericrocotus solaris Gray-chinned Minivet y = 0.26x - 1.49 0.12 4.08 0.05 32
Dicrurus aeneus Bronzed Drongo y = 0.18x - 0.76 0.09 2.89 0.10 30
Alcippe morrisonia Morrison’s Fulvetta y = 0.16x + 1.96 0.06 1.88 0.18 33 Erpornis zantholeuca White-bellied Erpornis y = 0.08x - 0.06 0.03 1.10 0.30 28 Pomatorhinus erythrocnemis Black-necklaced Scimitar-Babbler y = 0.09x - 0.59 0.03 0.96 0.33 27 Dendrocopos canicapillus Gray-capped Woodpecker y = 0.04x + 0.43 0.02 0.52 0.48 31 Myophonus insularis Taiwan Whistling-Thrush y = -0.12x + 2.99 0.11 4.00 0.05 29 Arborophila crudigularis Taiwan Partridge y = -0.06x + 2.53 0.01 0.43 0.52 32 Bambusicola sonorivox Taiwan Bamboo-Partridge y = -0.02x + 0.97 0.01 0.32 0.58 27 Pomatorhinus musicus Taiwan Scimitar-Babbler y = -0.003x + 3.26 0.00 0.00 0.97 33
a) Species names followed the eBird/Clements Checklist of Birds of the World Version 2017 (Clements et al. 2017).
b) x, temperature; y, vocal activity.
cial role in spring singing of birds and act as an important environmental factor in initiating the breeding process of birds.
Increased temperatures due to global climate change might directly or indirectly affect the breeding cycle of birds. The most conspicuous consequence found so far is the forwarded laying date for many northern spe- cies (Crick et al. 1997, Dunn and Winkler 1999, Both et al. 2004, Møller et al. 2006, Dolenec et al. 2011). As Carey (2009) pointed out, future studies should incorporate not only temperature data and the timing of the laying date but also the amount of population vari- ability in other life history traits. Hasan (2011) found that the onset of the dawn chorus is highly temperature dependent. The dawn cho- rus of some passerine birds in North America was shown to begin earlier with increasing temperature at nautical twilight (Bruni et al.
2014). Our study illustrated that the maxi-
mum temperature of the morning hours is a better predicator of the vocal behavior of birds than temperatures measured from earlier hours. This implies that the daily maximum temperature has a greater impact on the vo- cal behavior of birds. Since variations in temperatures in later hours of the morning are greater than those in the early hours, the maximum temperature tends to be a better indicator of the vocal activity of birds in the morning. In other words, birds are more sen- sitive to the maximum temperature of a day and respond with behavioral changes (Pattin- son and Smit 2017). As a consequence, global warming is likely to initiate spring singing at an earlier date and thus advance the breed- ing cycle of birds (Dunn and Winkler 2010, Møller et al. 2010). We consider that such a relationship can be further revealed through acoustic monitoring studies. Autonomous acoustic systems have been used as monitor-
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
Coefficient of determination
Morning hour
2011 no. species 2012 no. species 2011 vocal activity 2012 vocal activity
06:00 07:00 08:00 09:00 10:00 11:00
Fig. 5. The trend of the coefficient of determination (r2) in regression analyses when
temperatures (℃) at different morning hours were designated as the day’s temperature. Air temperatures at 10:00 and 11:00 had higher r2 values than those from earlier hours.
ing devices for avian biodiversity surveys (Hobson et al. 2002, Acevedo and Villanueva- Rivera 2006, Celis-Murillo et al. 2012, Snad- don et al. 2013). Advanced recording devices, such as SM4, can be scheduled and record for up to 250 h. Such recorded data could allow us to analyze the effects of climate change on the spring singing of birds.
Vocal activities of over 1/3 of the ana- lyzed species exhibited positive correlations with air temperature in this study. Of these, the Taiwan Barbet had the highest r2 value.
The Taiwan Barbet was recorded almost every hour in the recording, and it can be consid- ered the most vociferous species at the study site. This implies that species with persistent vocal activity are most suitable for autono- mous recording studies. Although the use of autonomous devices is sometimes criticized for failing to record as many birds compared to human observers (Huttoand Stutzman 2009), a selected set of vocal species would provide a reliable trend on spring singing to
detect possible influences of climate change.
The vocal activity of 4 bird species had a negative relationship with temperature, but it was only significant (p = 0.05) in the Taiwan Whistling-Thrush (Myophonus insularis).
We found this trend might have been due to behavioral features of these species. For example, Fang (2004) indicated that Taiwan Whistling-Thrushes often sing prior to sun- rise, and they are almost silent after 09:00.
This vocal activity pattern would result in a negative relationship with temperature which normally increases towards noon time.
CONCLUSIONS
This study showed that air temperature in early spring did affect the vocal activity of a forest bird community in Shan-Ping Forest, southern Taiwan. Furthermore, air tempera- tures close to noon (the maximum air tem- perature of a day) were better predicators of the vocal output of birds in that day. A bioas-
say by monitoring the co-occurrence of daily maximum air temperatures and initiation dates of spring singing of birds could be ap- plied to detect the influence of global warm- ing on the advance of spring singing in birds.
ACKNOWLEDGEMENTS
We thank Chia-Hua Wu, Chih-Wei Tsai, and Po-Jen Chiang for their advice on the operation of the acoustic monitoring system.
We thank the administration of the Shan-Ping Forest Ecological Garden for its great support during this study. We are especially grateful to Chao-Chin Lin, Yung-Hsio Chen, and Wen- Chih Lin for their assistance in this study and Hans-Uwe Dahms for critically reading an early draft of the manuscript.
LITERATURE CITED
Acevedo MA, Villanueva-Rivera LJ. 2006.
Using automated digital recording systems as effective tools for the monitoring of bird and amphibians. Wildl Soc Bull 34(1):211-4.
Ball GF. 1999. The neuroendocrine basis of seasonal changes in vocal behavior among songbirds. In: Hauser MD, Konishi M, editors.
The design of animal communication. Cam- bridge, MA: MIT Press. p 213-53.
Best LB. 1981. Seasonal changes in detection of individual bird species. Stud Avian Biol 6:252-61.
Both C, Artemyev AV, Blaauw B, Cowie RJ, Dekhuijzen AJ, Eeva T, et al. 2004. Large- scale geographical variation confirms that climate change causes birds to lay earlier. Proc Roy Soc Lond B Biol Sci 271(1549):1657-62.
Brandes TS. 2008. Automated sound record- ing and analysis techniques for birds survey and conservation. Bird Conserv Int 18:163-73.
Brewster JP, Simons TR. 2009.Testing the importance of auditory detections in avian
point counts. J Field Ornithol 80(2):178-82.
Bruni A, Mennill DJ, Foote JR. 2014. Dawn chorus start time variation in a temperate bird community: relationships with seasonal- ity, weather, and ambient light. J Ornithol 155(4):877-90.
Carey C. 2009. The impacts of climate change on the annual cycles of birds. Proc Roy Soc Lond B Biol Sci 364(1534):3321-30.
Catchpole CK, Slater PJB. 1995. Bird song:
biological themes and variations. Cambridge, UK: Cambridge Univ. Press. 323 p.
Celis-Murillo A, Deppe JL, Allen MF. 2009.
Using soundscape recordings to estimate bird species abundance, richness, and composition.
J Field Ornithol 80(1):64-78.
Celis-Murillo A, Deppe JL, Ward MP. 2012.
Effectiveness and utility of acoustic recordings for surveying tropic birds. J Field Ornithol 83(2):166-79.
Chen CC, Chiang PJ, Shieh BS, Lin CC.
2011. Diurnal timing of bird surveys using an acoustic monitoring system in the Shan-Ping Forest Ecological Garden. Taiwan J For Sci 26(4):313-21.
Clements JF, Schulenberg TS, Iliff MJ, Rob- erson D, Fredericks TA, Sullivan BL, Wood CL. 2017. The eBird/Clements checklist of birds of the world: v2017.
Crick HQP, Dudley C, Glue DE, Thomson DL. 1997. UK birds are laying eggs earlier.
Nature 388:526.
Dawson A. 2008. Control of the annual cycle in birds: endocrine constraints and plasticity in response to ecological variability. Phil Trans R Soc B 363(1497):1621-33.
Dawson DK, Efford MG. 2009. Bird popula- tion density estimated from acoustic signals. J Appl Ecol 46(6):1201-9.
DeJong MJ, Emlen JT. 1985. The shape of the auditory detection function and its implica- tions for songbird censusing. J Field Ornithol 56(3):213-23.
Dolenec Z, Dolenec P, Møller AP. 2011.
Warmer springs, laying date and clutch size of tree sparrows Passer montanus in Croatia.
Curr Zool 57(3):414-8.
Dunn PO, Winkler DW. 1999. Climate change has affected the breeding date of tree swallows throughout North America. Proc Roy Soc Lond B Biol Sci 266(1437):2487-90.
Dunn PO, Winkler DW. 2010. Effects of cli- mate change on timing of breeding and repro- ductive success in birds. In: Møller AP, Fielder W, Berthold P, editors. Effects of climate change on birds. Oxford, UK: Oxford Univ.
Press. p 113-28.
Fang JR. 2004. Territorial and vocal behavior of Formosan Whistling Thrush (Myiophoneus insu- laris) [Master’s thesis]. Taipei, Taiwan: School of Life Science, National Taiwan Normal Univ. 64 p.
[in Chinese with English abstract].
Garson PJ, Hunter ML Jr. 1979. Effects of temperature and time of year on the singing behaviour of Wrens Troglodytes troglodytes and Great Tit Parus major. Ibis 121(4):481-7.
Gil D, Gahr M. 2002. The honesty of bird song: multiple constraints for multiple traits.
Trends Ecol Evol 17(3):133-41.
Hasan NM. 2011. Comparison of the onset of dawn chorus of bulbuls and house sparrows in two different geographical locations: effect of climate, noise and light pollution. Res Opin Anim Vet Sci 1(4):220-5.
Hasan NM, Badri M. 2016. Effect of ambient temperature on dawn chorus of house spar- rows. Environ Ecol Res 4(3):161-8.
Hau M. 2001. Timing of breeding in variable environments: tropical birds as model system.
Horm Behav 40(2):281-90.
Hau M, Wikelski M, Wingfield JC. 1998.
A Neotropical forest bird can measure slight changes in tropical photoperiod. Proc Roy Soc Lond B Biol Sci 265(1391):89-95.
Higgins RM. 1979. Temperature-related variation in the duration of morning song
of the Song Thrush Turdus ericetorum. Ibis 121(3):333-5.
Hobson KA, Rempel RS, Hamilton G, Turn- bull B, Wilgenburg SLV. 2002. Acoustic sur- veys of birds using electronic recordings: new potential from an omnidirectional microphone system. Wildl Soc Bull 30(3):709-20.
Hutto RL, Stutzman RJ. 2009. Humans versus autonomous recording units: a com- parison of point-count results. J Field Ornithol 80(4):387-98.
King HB, Ho HJ, Chang NH. 1990. Bird sur- vey report at Shan-Ping and Nan-Fong-Shan areas. Taipei, Taiwan: Council of Agriculture, Ex- ecutive Yuan. [in Chinese with English abstract].
Lehmann M, Spoelstra K, Visser ME, Helm B. 2012. Effects of temperature on circadian clock and chronotype: an experimental study on a passerine bird. Chronobiol Int 29(8):1062-71.
Leitner S, Van’t Hof TJ, Gahr M. 2003.
Flexible reproduction in wild canaries is inde- pendent of photoperiod. Gen Comp Endocrinol 130(2):102-8.
Lu SS, Lin WC, Chen YH, Lin CC. 2009.
Application of wireless sensor network to study the defensive behavior of Apis cerana (Hymenoptera: Apidae). J Natl Park 19:1-8. [in Chinese with English abstract].
Møller AP, Flensted-Jensen E, Mardal W.
2006. Rapidly advancing laying date in a seabird and the changing advantage of early reproduction. J Anim Ecol 75(3):657-65.
Møller AP, Flensted-Jensen E, Klarborg K, Mardal W, Nielsen JT. 2010. Climate change affects the duration of the reproductive season in birds. J Anim Ecol 79(4):777-84.
Morse DH. 1970. Territorial and courtship songs of birds. Nature 226:659-61.
Pattinson, NB, Smit B. 2017. Seasonal behav- ioral responses of an arid-zone passerine in a hot environment. Physiol Behav 179:268-75.
Reid ML. 1987. Costliness and reliability in the singing vigour of Ipswich Sparrow. Anim
Behav 35(6):1735-43.
Scott JM, Ramsey FL, Kepler CB. 1981.
Distance estimation as a variable in estimating bird numbers from vocalizations. Stud Avian Biol 6:252-61.
Slagsvold T. 1977. Bird song activity in relation to breeding cycle, spring weather, and environ- mental phenology. Ornis Scand 8(2):197-222.
Snaddon J, Petrokofsky G, Jepson P, Willis KJ. 2013. Biodiversity technologies: tools as change agents. Biol Lett 9(1):20121029.
Strain JG, Mumme RL. 1988. Effects of food supplementation, song playback, and tempera- ture on vocal territorial behavior of Carolina Wrens. Auk 105(1):11-6.
Thomas RJ. 1999. Two tests of a stochastic dynamic programming model of daily singing routines in birds. Anim Behav 57(2):277-84.
Tsai CF, Lee YF, Chen YH, Chen WM. 2015.
Species turnover in tropical montane forest avifauna links to climate correlates. Glob Ecol Conserv 3:541-52.
Wingfield JC, Farner DS. 1993. Endocrinol- ogy of reproduction in wild species. Avian Biol 9:163-327.
Wingfield JC, Follett BK, Matt KS, Farner DS. 1980. Effect of day length on plasma FSH and LH in castrated and intact White-crowned Sparrows. Gen Comp Endocrinol 42(4):464-70.
Wingfield JC, Hahn TP, Maney DL, Schoech SJ, Wada M, Morton ML. 2003.
Effects of temperature on photoperiodically induced reproductive development, circulat- ing plasma luteinizing hormone and thyroid hormones, body mass, fat deposition and molt in mountain White-crowned Sparrows, Zono- trichia leucophrysoriantha. Gen Comp Endo- crinol 131(2):143-58.
Appendix 1. The mean, minimum, and maximum air temperatures (℃) recorded from 06:00 to 11:00 of each day in March of 2011 and 2012
March 2011 2012
Mean±SD Minimum Maximum Mean±SD Minimum Maximum
1 17.19±2.30 15.19 20.90 18.17±1.74 16.90 21.09
2 18.35±1.44 16.62 20.14 18.05±1.72 16.62 20.90
3 NULL NULL NULL 19.58±1.85 17.86 22.43
4 NULL NULL NULL 18.84±1.79 17.48 21.76
5 NULL NULL NULL 17.11±2.43 15.28 21.19
6 17.98±2.07 15.57 20.14 18.03±2.03 16.43 21.47
7 18.11±0.92 16.71 18.90 19.32±1.99 17.38 22.62
8 13.19±0.84 12.59 14.80 19.98±1.84 18.33 22.81
9 14.39±1.72 12.79 17.19 20.41±1.28 18.90 21.95
10 15.68±1.35 14.13 17.57 17.65±0.44 17.67 18.52
11 16.12±1.00 14.90 17.48 16.16±0.25 15.76 16.43
12 17.44±2.56 14.90 21.00 15.08±0.18 14.90 15.38
13 17.57±2.63 15.00 21.66 16.09±0.95 14.90 17.19
14 18.28±2.52 15.47 22.24 17.85±1.73 16.24 20.42
15 19.00±1.65 17.67 21.86 18.44±2.06 16.52 21.86
16 13.27±0.17 12.98 13.46 19.05±2.09 16.90 22.24
17 13.32±0.57 12.59 13.94 19.65±2.11 17.76 23.10
18 16.03±2.48 13.85 20.04 19.41±2.27 17.67 23.29
19 17.76±2.34 15.47 21.47 21.11±1.40 19.19 22.72
20 21.35±2.55 19.28 25.42 19.98±1.84 17.95 22.43
21 20.83±3.13 18.14 25.90 20.25±0.75 19.38 21.09
22 21.22±1.85 19.09 23.97 18.76±2.82 16.05 23.29
23 14.95±0.21 14.71 15.28 19.95±1.78 18.24 22.24
24 14.93±0.43 14.33 15.47 16.11±0.58 15.28 16.81
25 15.33±1.16 13.94 17.28 15.10±1.61 13.75 17.76
26 15.09±1.12 13.75 17.00 14.96±2.49 12.30 18.81
27 14.41±0.47 13.85 15.09 15.32±2.88 12.40 19.66
28 13.54±0.35 13.08 13.94 16.65±2.35 14.04 20.23
29 15.98±2.19 13.46 19.19 16.37±2.72 13.46 20.42
30 15.82±1.48 13.85 17.57 17.62±2.55 15.09 21.57
31 14.48±0.65 13.65 15.09 20.22±2.28 17.57 23.48