交通大學生物科技學系
博士論文
骨型態發生蛋白的訊息途徑對細胞功能之探討:BMP-4 和流體剪力對
骨母細胞分化作用及腫瘤細胞生長作用之影響
Controlling Cell Fate by Bone Morphogenetic Protein Signaling:
Roles of BMP-4 and Fluid Shear Stress in Osteoblast Differentiation
and Tumor Cell Growth
Student: 張順福(Chang, Shun-Fu)
Advisor: 張 正 教授(Prof. Chang, C. Allen) 裘正健 教授(Prof. Chiu, Jeng-Jiann)
Institute of Biological Science and Technology National Chiao Tung University
Hsin-Chu, Taiwan, Republic of China 中華民國九十七年十一月
致 謝 博士班研究生活艱辛、漫長,除了需要自我不斷努力、隨時精進外,老師及 同學間的指導和討論也是使本論文能順利完成的因素之一,故完成博士論文研究 的最後階段,我要衷心感謝這些一路陪伴我的人。 首先要感謝的是我的指導教授交通大學張正博士及國家衛生研究院裘正健 博士,感謝您們在我進行論文研究時,包含學業上和研究方向的選擇、實驗設計 和最後論文撰寫上給予悉心的指導。也感謝加州大學聖地牙哥分校的錢煦院士在 論文研究上的指導以及將我的研究成果推薦到國際知名期刊(PNAS)。最後感謝 陽明大學鄭誠功博士、台大醫院孫瑞昇醫生和交通大學吳東昆博士在論文口試時 提供許多寶貴的意見,使本論文能更加的完整。 接著要感謝的是我的研究所學長姐及同學:何狗、丸子、俊傑、小羊、弘毅、 鴻萱、隆勇,在學業上、生活上及精神上給予的鼓勵與幫忙,由於我是在學校外 進行論文研究,所以特別感謝在學校的何狗、丸子及弘毅能隨時提醒及幫助我各 方面的事情。另外,也感謝實驗室的學長姐及學弟妹:政男、欣欣、全人、佩玲、 芷懿、定宇、連、依婷、育銘對部分實驗的幫忙及研究過程中的互相討論及勉勵; 其餘的研究室成員,也感謝你們在研究生活上所給予的歡樂和指教。 最後要感謝的是一路無怨無悔陪伴我、支持我拿到博士學位的家人,因為你 們無私的付出,讓我可無後顧之憂的全力完成學業,我只能再說一聲,我愛你們、 謝謝你們。 拿到博士學位代表著一個時期的結束,也代表著一個新時期的開始,身上所 擔負的責任也將更加沈重。生物科技的世界瞬間萬變,但也因此充滿著無限樂 趣,我將以最愉悅的心情,竭盡所能的在自己最有興趣的生物研究領域上貢獻一 己之力。
TABLE OF CONTENTS 致謝……….……... i 目錄………ii 表目錄………...v 圖目錄………...vi 中文摘要……….viii 英文摘要…..………...xi Background………..xiv Chapter I. Introduction...1
1.1 Bone morphogenetic proteins (BMPs)...2
1.2 BMPs signaling in osteogenesis...5
1.3 BMPs signaling in tumor biology...7
1.4 Mechanical flow forces biology...8
1.5 Mechanical flow forces and tumor………..10
Chapter II. Hypothesis and Specific Aims………...13
Chapter III. Materials and Methods……….16
3.1 Cell lines and cell culture………...17
3.2 Materials………..17
3.3 Flow apparatus………...18
3.4 Flow cytometric analysis………18
3.5 ALP specific activity assay……….19
3.6 RNA isolation and quantitative real-time PCR………...19
3.9 Treatments with RGD peptides and mAbs………..21
3.10 Reporter gene construct, DNA plasmids, siRNA, transfection, and luciferase assay………..21
3.11 Electrophoretic mobility shift assay (EMSA)………...21
3.12 Statistical analysis……….22
Chapter IV. Results……….24
4.1 Bone morphogenetic protein-4 induces odteoblast G0/G1 arrest and differentiation via increased expressions of p21CIP1 and p27KIP1: roles of integrins, extracellular signal-regulated kinase, and smad………...25
4.1.1 BMP-4 induces G0/G1 arrest and increased expressions of p21CIP1 and p27KIP1 in osteoblast-like cells………...25
4.1.2 BMP-4-induced MG63 cell differentiation is mediated by p21CIP1 and p27KIP1……….25
4.1.3 BMP-4-induced p21CIP1 and p27KIP1 expressions and G0/G1 arrest in MG63 cells are mediated by the BMPRIA/Smad5 pathway………...26
4.1.4 ERK mediates BMP-4-induced Smad phosphorylation in MG63 cells………...27
4.1.5 The BMP-4-induced ERK and Smad phosphorylations in MG63 cells are mediated by β3, but not αv and β1, integrins………..28
4.2 Tumor cell cycle arrest induced by shear stress: roles of integrins and Smad…...29
4.2.1 Shear stress induces G2/M arrest and corresponding changes in cell cycle regulatory protein expression in tumor cells………..29
4.2.2 Shear stress induces sustained phosphorylations of Smad1/5 in MG63 cells through BMPRIA………...29
4.2.3 Shear-induced G2/M arrest in tumor cells is mediated by Smad1/5………30
4.2.4 αvβ3 and β1 integrins mediate shear-induced Smad1/5 phosphorylation and G2/M arrest in tumor cells……….31
4.2.5 Shear stress inhibits MG63 cell differentiation and Runx2 binding activity in the
nucleus………..………32
4.2.6 αvβ3 and β1 integrins and Smad5 mediate shear-induced inhibition of MG63 cell differentiation………...32
Chapter V. Discussion……….…....55
5.1 Bone morphogenetic protein-4 induces odteoblast G0/G1 arrest and differentiation via increased expressions of p21CIP1 and p27KIP1: roles of integrins, extracellular signal-regulated kinase, and smad………...56
5.2 Tumor cell cycle arrest induced by shear stress: roles of integrins and Smad…...60
Chapter VI. Conclusion……….….67
Chapter VII. Future potential programs...69
Chapter VIII. References...72
Chapter IX. Notes added in proof...90
9.1 BMP-4-induced ERK1/2 and Smad1/5 activations through the association of β3 integrin, FAK, and Shc in MG63……….91
9.2 BMP-4-induced Smad activations and hence p21CIP1 and p27KIP1 expression are mediated by ERK2, but not ERK1, in MG63………...92
9.3 BMP-4-induced G0/G1 arrest in MG63 cells is mediated by Smad1 and ERK2 activation and p21CIP1 and p27KIP1 expression………...92
Chapter X. Publications...98
LIST OF TABLES
Table 1-1 Aberrations of BMP signaling in human tumor, the functional relevance and clinical correlations………...12 Table 4-1 BMP-4 induces G0/G1 arrest in human MG63 and mouse MC3T3-E1
osteoblast-like cells………..……….34 Table 4-2 BMP-4-induced G0/G1 arrest in MG63 cells is dose-independent…..……….…….35
Table 4-3 BMP4-induced G0/G1 arrest in MG63 cells is mediated by BMPRIA and
Smad5………..…….…….36 Table 4-4 Shear stress induces a G2/M cell cycle arrest in tumor cells…………..…………..37
Table 4-5 Shear-induced G2/M arrest in tumor cells is mediated by αvβ3 and β1 integrins and
Smad1/5………39 Table 9-1 BMP4-induced G0/G1 arrest in MG63 cells is mediated by Smad1, ERK2, p21, and
LIST OF FIGURES
Figure 3-1 Schematic diagram of the cell culture flow model……….….…23 Figure 4-1 BMP-4 regulates expression of cell cycle regulatory proteins in MG63 cells………...40 Figure 4-2 BMP-4-induced MG63 cell differentiation is mediated by p21CIP1 and
p27KIP1………..….42
Figure 4-3 BMP-4-induced p21CIP1 and p27KIP1 expressions are mediated by BMPRIA/Smad5 in MG63 cells………....43 Figure 4-4 BMP-4-induced Smad1/5 activation is mediated by ERK in MG63 cells………..45 Figure 4-5 BMP-4-induced ERK and Smad1/5 activations are mediated by β3 integrin in
MG63 cells………...46 Figure 4-6 Shear stress regulates expressions of cell cycle regulatory proteins in MG63 cells………...48 Figure 4-7 Shear stress induces sustained phosphorylation of Smad1/5 in MG63 cells through BMPRIA………...49 Figure 4-8 Shear-induced G2/M arrest in MG63 cells is mediated by Smad1/5………..……51
Figure 4-9 Shear-induced Smad1/5 activations are mediated by αvβ3 and β1 integrins in
MG63 cells………...52 Figure 4-10 MG63 cell differentiation and Runx2 binding activity in the nucleus were inhibited by shear stress acting through αvβ3 and β1 integrins and Smad5………..…53
Figure 5-1 Schematic representation of the signaling pathways regulating BMP-4-induced cell cycle arrest and differentiation in osteoblast-like cells………...65 Figure 5-2 Schematic representation of the signaling pathways regulating cell cycle and differentiation in tumor cells in response to shear stress……….……...66 Figure 9-1 BMP-4-induced ERK1/2 and Smad1/5 activations through the association of β
integrin, FAK, and Shc in MG63 cells………..96 Figure 9-2 BMP-4-induced Smad1/5 activations and hence p21CIP1 and p27KIP1 expression are mediated by ERK2 in MG63 cells………97
中文摘要
骨 型 態 發 生 蛋 白 ( bone morphogenetic proteins, BMPs ) 是 屬 於 轉 型 蛋 白 (transforming growth factor-beta, TGF-β)中的一種生長因子,因其具有在體內誘 導異位(ectopic site)骨頭或軟骨生成作用的能力而被發現與命名,但近幾年研究發 現,骨型態發生蛋白在腫瘤細胞上也具有重要的影響性。骨型態發生蛋白-4 為骨格生成 作用中最重要的生長因子之一,但經由其他研究發現,骨型態發生蛋白-4 也對其他種類 細胞的細胞功能具有調控作用,包含正常細胞和腫瘤細胞的生長或細胞凋亡。機械微環 境(mechanical microenvironment)在組織的發展、維持、功能或致病上相當重要, 近期研究發現,機械流體力學(mechanical flow forces)對腫瘤細胞的生長具有影響 性,也會調控 Smad 訊息途徑來影響細胞功能。此外,科學家在哺乳動物的細胞功能研 究上發現,影響細胞週期的運行結果可調控或決定細胞功能的作用。總和以上的論述, 我們假設骨型態發生蛋白的訊息途徑可調控細胞週期的運行進而影響骨型態發生蛋白 -4 所調節骨母細胞(osteoblasts)增生(proliferation)及分化作用(differentiation) 和機械流體力學調節腫瘤細胞生長作用。本論文的研究目的為(1)觀察骨型態發生蛋 白-4 是否會調控細胞週期的運行進而調節骨母細胞的分化作用;(2)研究骨型態發生蛋
白-4 是否藉由調節細胞週期調控因子(cell cycle regulators)的表現來影響此發生 過 程 ;( 3 ) 證 實 細 胞 膜 上 的 integrins 是 否 與 骨 型 態 發 生 蛋 白 的 細 胞 膜 上 受 體 (receptors)交互作用,因而促進及加強骨型態發生蛋白-4 的影響;(4)觀察機械流 體力學是否會調控細胞週期的運行進而調節腫瘤細胞的生長作用;(5)研究機械流體力 學是否藉由骨型態發生蛋白的訊息途徑來調節細胞週期調控因子的表現進而影響此發 生過程;(6)觀察機械流體力學所刺激骨型態發生蛋白的訊息途徑是否是經由刺激細胞 自體分泌(autocrine)骨型態發生蛋白所引起;(7)證實細胞膜上科學家認定的機械
接受器(mechanosensors)-integrins 是否為腫瘤細胞接受機械力的受體,因而影響細 胞內部骨型態發生蛋白的訊息途徑。
研究發現,骨型態發生蛋白-4 的刺激會遏止兩種不同的骨母細胞的細胞週期,並令 其停止在 G0/G1時期,此作用主要是藉由增加細胞週期調控因子 p21CIP1 和 p27KIP1的蛋白
表 現 所 引 起 , 並 因 此 造 成 細 胞 進 行 分 化 作 用 。 研 究 中 利 用 小 干 擾 RNA ( small interfering RNA, siRNA)技術發現此作用主要是受到骨型態發生蛋白的訊息途徑:骨 型態發生蛋白的膜上第一型受體 A 和 Smad5 所調控。此外,骨型態發生蛋白-4 也會刺激 ERK 基酶的短暫磷酸化,研究發現當 ERK 基酶的活性受到抑制時會抑制骨型態發生蛋白 的訊息蛋白 Smad1/5 的活化,且抑制細胞膜上 integrin β3的表現時,會同時造成骨型 態發生蛋白-4 刺激 ERK 和 Smad1/5 的活化被抑制。此結果提出骨型態發生蛋白-4 所造 成的反應及訊息途徑會受到 integrin β3-ERK 所調控。 在機械流體力學對腫瘤細胞生長調控的研究發現,當給予機械流體力學所產生的剪 力刺激四種腫瘤細胞 24 和 48 小時後,會遏止細胞週期的運行,並使其停止在 G2/M 時期, 此作用主要是藉由增加細胞週期調控因子 cyclin B1 和 p21CIP1 的蛋白表現及降低 cyclins A, D1, and E, Cdk-1, -2, -4, and -6, and p27KIP
的蛋白表現以及 Cdk1 的 活性所引起。研究中利用抗體(antibodies)和小干擾 RNA 技術發現此作用主要是透過
膜上 integrins αvβ3 和β1經由骨型態發生蛋白的訊息途徑:骨型態發生蛋白的膜上第
一型受體 A 和 Smad1/5 所調控。此外,研究也發現剪力會透過骨型態發生蛋白的訊息途 徑抑制腫瘤細胞的分化作用,包含降低轉錄因子 Runx2 與 DNA 結合的活性和分化蛋白骨 鈣素(osteocalcin)和鹼性磷酸酶(alkaline phosphatase, ALP)的基因和蛋白表現。
本論文的研究結果證實骨型態發生蛋白的訊息途徑可藉由調控細胞週期的運行而 調節骨型態發生蛋白-4 所誘導的骨母細胞分化作用和機械流體力學所誘導的腫瘤細胞
生長抑制作用。也證實機械微環境具有調控腫瘤細胞的分子機制進而影響細胞功能的作 用。我們提出對骨型態發生蛋白的訊息途徑活性的調控作用可能對治療骨骼或腫瘤相關 疾病提供新的方法,另外,機械流體力學與骨型態發生蛋白的訊息途徑之間的相互關係 對於治療腫瘤病人可能也可提供新的研究方向。
ABSTRACT
Bone morphogenetic proteins (BMPs), members of the transforming growth factor-β superfamily, were originally identified by their unique ability to induce ectopic bone and cartilage formation in vivo. In recent studies, BMP signaling has been suggested to play an important role not only in bone cells but also in tumor cells. BMP-4 is one of the most potent inducers of bone formation and is also demonstrated to be a potent growth factor for modulating other cells’ functions, including normal and tumor cells’ growth or apoptosis. The importance of the mechanical microenvironment in tissue development, maintenance, function and pathogenesis has been well established for several decades. Recent studies demonstrated that mechanical flow forces may affect the growth of tumor cells. Moreover, it is also suggested that mechanical flow forces may mediate the Smad signaling pathway. In mammals, the regulation of cell cycle distribution can mediate cell functions. We hypothesize that BMP signaling may regulate the cell cycle distribution, thereby mediating osteoblast proliferation and differentiation under BMP-4 treatment and mediating tumor cell growth under mechanical flow forces. The aims of this study were: (1) to observe if BMP-4 regulates the cell cycle distribution in order to induce differentiation in osteoblasts; (2) to investigate if BMP-4 signaling mediates the expression of cell cycle regulators and hence modulates this process; (3) to demonstrate if membrane integrins interact with BMP-4 receptors for optimization of the BMP-4 effects; (4) to observe if mechanical flow forces regulate the cell cycle distribution in order to affect tumor cell growth; (5) to investigate if mechanical flow forces mediate the expression of cell cycle regulators through BMP signaling and hence modulate this process; (6) to observe if mechanical flow force-stimulated BMP signaling comes from BMP autocrine effects; and (7) to demonstrate if the mechanosensor (i.e., integrin) response to mechanical flow forces induces intracellular BMP signaling.
induced the increased expression of p21CIP1 and p27KIP1 and hence cell differentiation, but had no effect on the expression of cyclins A, B1, D1, and E, and Cdk-2, -4, and -6. Using specific small interfering RNA (siRNA), we found that the BMP-4-induced G0/G1 arrest and increased
expressions of p21CIP1 and p27KIP1 were mediated by BMP receptor type IA-specific Smad5. In addition, BMP-4 induced transient phosphorylation of extracellular signal-regulated kinase (ERK); transfection with ERK-specific siRNA inhibited BMP-4-induced Smad1/5 activation. Moreover, transfection with specific siRNA for β3, but not αv and β1, integrins inhibited the
BMP-4-induced ERK and Smad1/5 phosphorylation, suggesting that the BMP-4-induced responses are mediated by the β3 integrin through ERK.
In four tumor cell lines, incubation under static conditions for 24 or 48 h led to G0/G1
arrest; in contrast, shear stress (12 dynes/cm2) induced G2/M arrest. Shear stress induced
increased expression of cyclin B1 and p21CIP1 and decreased expression of cyclins A, D1, and E, and cyclin-dependent protein kinases (Cdk)-1, -2, -4, and -6, and p27KIP1, as well as a decrease in Cdk1 activity. Using specific antibodies and siRNA, we found that the shear-induced G2/M arrest and corresponding changes in G2/M regulatory protein expression
and activity were mediated by αvβ3 and β1 integrins through bone morphogenetic protein
receptor type IA-specific Smad1 and Smad5. Shear stress also down-regulated runt-related transcription factor 2 (Runx2) binding activity and osteocalcin and alkaline phosphatase expression in tumor cells; these responses were mediated by αvβ3 and β1 integrins through
Smad5.
Our findings indicate that BMP signaling may mediate the cell cycle distribution to regulate BMP-4-induced osteoblast differentiation and the mechanical flow forces-induced tumor cell growth arrest. Our findings also provide new insights into the mechanisms by which the mechanical microenvironment modulates molecular signaling, gene expression, the cell cycle, and functions in tumor cells. Modulation of the activities of BMP signaling may be
useful in establishing new approaches to the treatment of a variety of bone or tumor disorders. Moreover, the communications between mechanical flow forces and BMP signaling may contribute new research directions for treating tumor patients, and further detailed investigations are needed.
BACKGROUND
Cells are constantly influenced by their biochemical and mechanical microenvironments. Chemical ligand binding to their specific receptors under biochemical stimulation and mechanical force regulating membrane mechanosensors, e.g., integrins, can transduce information into the cell and hence modulate cell functions. Thus, modulation in cell signaling, gene expression, structure and function by both chemical and mechanical factors plays important role in health and disease. Bone is living, growing tissue, and is constantly renewed through a two-part process called remodeling. Bone is continuously formed and absorbed by the actions of osteoblasts and osteoclasts, respectively. During bone formation and remodeling, osteoblast and osteoclast function may be regulated by chemical factors, such as BMPs, and mechanical factors, including flow-induced shear stress. Both chemical and mechanical stimuli are essential for the maintenance of skeletal integrity and bone mass. Suppression of these stimuli under conditions such as menopause or spaceflight results in bone loss.
Osteoporosis, osteomalacia, etc, are metabolic bone diseases with major histomorphometric abnormalities: a decreased amount of bone mass and a reduced bone formation rate. Therapy of these diseases is usually inefficiency and requires lone-term treatments. Thus, discovery and establishment of new procedures and methods are urgent. Recent studies suggest that BMPs and flow-induced mechanical forces play important roles in functional modulation of different types of cells, including bone and tumor cells. Studies on the BMP effects demonstrated that BMPs are potential inducers for stimulating osteoblast differentiation and increasing bone mass. BMP-4 has been used clinically to treat the bone fracture healing in patients. Moreover, flow-induced mechanical forces have been proposed to modulate the process of bone formation and remodeling. Interstitial fluid flow in bone, in particular flow-induced shear stress, is required for the maintenance of bone integrity and
serves as a mediator in signal transduction and gene expression during mechanical loading-induced bone remodeling. In physiology, tissues and cells are affected by the interplay between chemical and mechanical stimuli. However, whether combination of BMPs and flow-induced shear stress provides new strategy to treat bone diseases remains to be determined. In this thesis, we investigated the effects of BMPs and flow-induced shear stress on signal transduction and gene expression in bone or tumor cells and the consequent modulation of their functions.
In the present study, we used human MG63 osteoblast-like cells to test our hypothesis. Human MG63 cells have been identified to possess osteoprogenitor potentials of differentiation in response to differentiation signals, tumor cell charactistics with rapid proliferative rate and no aging. In addition to investigate the effects of BMPs and flow-induced shear stress on osteoblast signaling, gene expression and function, we also investigate the effect of shear stress on tumor cell growth. Understanding the detail mechanisms underlying the effects of chemical and mechanical stimuli on bone cell differentiation and tumor cell growth may provide insignts that may be taken into account for therapeutic implication.
CHAPTER I
1.1 Bone morphogenetic proteins (BMPs)
Bone morphogenetic proteins (BMPs) belong to a large family of structurally related proteins known as the transforming growth factor-β (TGF-β) superfamily, to which TGF-βs, activins, nodal and Mullerian inhibiting substance (MIS)/anti-Mullerian hormone (AMH) also belong [1]. BMPs were originally identified as proteins capable of inducing ectopic cartilage and bone formation when implanted subcutaneously or in muscle pouches [2-3]. This ectopic cartilage/bone formation recapitulates the entire sequence of events that occurs during endochondral bone development in limb buds [4-5]. The activity of BMPs was discovered in 1965, but the BMP proteins were purified and sequenced in the late 1980s [2-3, 6-7]. After that, recombinant BMP proteins were expressed [6, 8]. To date, over 20 BMP family members have been identified and characterized. BMPs were found to have important roles in directing the cell fate choices of mesenchymal cells in vitro. They stimulate osteoblast differentiation and inhibit the differentiation of mesenchymal cells into other cell lineages [9-11]. Subsequent studies showed that BMPs are not only involved in bone formation, but, like other members of the TGF-β superfamily, are multifunctional proteins with effects not related to the formation of bone [12-13]. Correspondingly, BMPs were found to be expressed not only in skeletal tissues, but also in many soft tissues [14]. Consistent with these results, the gain or loss of function studies of BMPs in mice demonstrated that, besides their critical roles in cartilage and bone formation [15-19], BMPs are also important during mouse development and in the adult functions of cardiovascular, pulmonary, reproductive and urogenital organs and in the nervous system [20]. Recent studies suggest that BMPs act as inflammatory cytokines in systemic arteries, promoting endothelial activation, and confirm a striking upregulation of BMPs in atherosclerotic lesions [21-22]. Furthermore, it has been demonstrated that BMPs play important roles during tumor cell growth and metastasis [23]. In light of all these experimental results, it is
suggested that the pleiotropic functions of BMPs implicate a need for the further investigation of their significance in different cell types, including normal and tumor cells.
BMP signaling is mediated by means of the heteromeric complex formation of cognate type I and II transmembrane serine/threonine kinase receptors. Three type I receptors have been shown to bind BMP ligands, including type IA activin receptor (ActRIA or Alk2) and type IA (BMPRIA or Alk3) and IB (BMPRIB or Alk6) BMP receptors [24-26]. Three type II receptors have also been identified, consisting of type II BMP receptor (BMPRII) and type IIA (ActRIIA) and type IIB (ActRIIB) activin receptors [13, 27-28]. Type I and type II BMP receptors are both indispensable for signal transduction. The type II receptors are constitutively active kinases, whereas the type I receptors activate intracellular substrates and thus determine the specificity of the intracellular signals [29-32]. For example, BMP-6 binds exclusively to Alk2, which only activates specific substrates (receptor-regulated Smad proteins, R-Smad, i.e., Smad1 and 5). In contrast, BMP-7 preferentially binds to Alk2, yet has an affinity for Alk3 and 6; thus all three substrates (Smad1, 5, and 8) relay signals. Upon BMP binding, two type receptors form a complex consisting of two pairs of type I and type II receptor complexes [33]. The activation of BMPRII phosphorylates BMPRI, which then leads to the phosphorylation of R-Smads, i.e., Smad1, 5, and 8. The activated R-Smads interact with the common partner Smad4 (Co-Smad) and accumulate in the nucleus to regulate the transcription of different target genes depending on the transcriptional co-modulators recruited [29-35]. R-Smads can directly bind to DNA, however, the affinity is relatively low and an interaction with sequence-specific DNA binding proteins is critical for the formation of a stable DNA-binding complex [36]. The first demonstration that Smads can directly bind to DNA was reported in drosophila [37]. After that, it was also suggested that R-Smads may interact with bone-specific transcription factor Runx2 and activate the transcription of target genes such as alkaline phosphatase (ALP), osteocalcin (OCN), or
COX-2 and type X collagen (Col-X) in osteoblasts or in chondrocytes [38-42]. Like other signal transduction pathways, BMP signaling is subjected to modulation at multiple levels. Extracellularly, secreted BMP antagonists, such as noggin, chordin, follistatin, and sclerostin, control the local concentration of active BMPs by sequestration [34, 43]. Within the cytoplasm, inhibitory Smads (I-Smads, i.e., Smad 6 and 7) interfere with R-Smad and Co-Smad complexes [43], and Smurfs (Smad ubiquitination regulatory factors) target activated receptors and R-Smads for degradation [43-44].
Although abundant evidence demonstrates that Smads are critical for BMP signaling, accumulating data suggests that Smad-independent pathways may also exist. Recently, an alternative BMP-mitogen-activated protein kinase (MAPK) signaling pathway [45-47], including extracellular signal-regulated kinase (ERK), has been described. In vitro and cell culture studies have revealed that ERK may phosphorylate serine residues of Smad1 [48]. As a consequence of this phosphorylation, the nuclear accumulation of Smad1 is inhibited in cultured cells, although the mechanism for this inhibition is unclear. Thus, it appears that ERK can crosstalk with BMP signaling by differentially phosphorylating Smad1 to affect its nuclear localization. In addition, recent studies in osteoblasts also reported that ERK may mediate the signaling pathway of BMP-4 in regulating the synthesis of fibronectin and extracellular fibronectin fibrillogenesis, which is involved in the process of bone mineralization [49]. The importance of ERK in BMP signaling effects has already been demonstrated, however, the detailed mechanisms and the correlation between Smad and ERK pathways in response to BMPs remain unclear. Membrane integrins are the other BMP signaling mediators. Integrins, as the main receptors that connect the cytoskeleton and the extracellular matrix (ECM), have been shown to play important roles in modulating gene expression and cellular functions in a wide variety of cells seeded on the ECM [50]. Osteoblasts express several types of integrins, including αvβ3 and those containing the β1
subunit (dimerized with α subunits, including α1, α3, α5, and α6) [51-53], which have been
shown to play important roles in osteoblast commitment and differentiation [54]. The administration of the RGD (Arg-Gly-Asp) peptide, which is a non-specific inhibitor of integrins, has been shown to inhibit bone formation and resorption of fetal rat parietal bones [55]. The interaction between integrins and fibronectin has been shown to be required for calvarial osteoblast differentiation [56-57]. Recent studies using function-perturbing antibodies against integrins showed that αvβ and αβ1 integrins play an essential role in
BMP-2-induction of osteoblast differentiation [54]. It is likely that integrins cooperate with BMP receptors to mediate the BMP-eliciting signaling pathways in osteoblasts and hence modulate their gene expression and cellular functions.
1.2 BMP signaling in osteogenesis
During Osteogenesis, bone is formed in two different manners: intramembranous ossification and endochondral ossification [30, 58]. In the case of intramembranous ossification, osteogenesis occurs directly through the differentiation of mesenchymal stem cells to mature osteoblasts. Ossification generated in this fashion is responsible for forming the flat bones of the skull, part of the clavical, and the additional bone on the periosteal surface of long bones. In the process of endochondral ossification, mesenchymal stem cells first condense to form a cartilage model, and then bone formation occurs by replacing this cartilage. This type of ossification forms most of the bones, including the axial and appendicular skeletons. A fundamental function of BMP signaling is to induce the differentiation of mesenchymal stem cells toward cells of the osteoblast lineage to promote osteoblast maturation and function. To determine the importance of BMP ligands, receptors and signaling proteins in embryonic development and bone formation, null mutations affecting BMP signaling have been created and the phenotypic changes in the animals have
of mesodermal differentiation and pattern defects [59]. BMP-2/-4 conditional knockout (CKO) mice show severe defects in bone development [60]. Similarly BMP-4 CKO mice also show defects in bone development and postnatal bone formation [61]. Transgenic mice that overexpress noggin, BMPs’ antagonist, in osteoblasts have reduced bone mineral densities and bone formation rates [62-63]. Transgenic mice expressing truncated dominant-negative BMP receptors have been shown to develop postnatal osteopenia [64]. Deleting the Smad1 gene specifically in osteoblasts causes a reduction in bone mass [65] and mice with disruption of Smad5 develop multiple embryonic defects [66].
Osteogenesis and bone remodeling are complex process that begin in the embryo and continue in the adult to maintain the balance between bone formation and resorption. These processes include osteoblast proliferation, differentiation, and apoptosis [67-68]. Evidence is accumulating that shows mammalian progenitor cell proliferation depends on the cell cycle progression and that they must exit the cell cycle to differentiate into mature cells or program to death [69]. Therefore, these cell functions may be closely linked and all regulated by cell cycle distribution. BMPs have the unique function of inducing the differentiation of the osteoblast lineage, leading to an increase in the pool of mature cells. Recently, conventional gene knockout experiments have shown that BMPs have diverse biological activities in a variety of cell types, mediated by their ability to regulate cell proliferation and apoptosis [70-71]. Therefore, scientists suggest that the effects of BMPs on osteogenesis may be global, including the modulation of osteoblast proliferation, differentiation, and apoptosis, and that these effects and correlations need to be further clarified.
The proliferation of eukaryotic cells depends on their progression through the cell cycle, and cell cycle arrest at the G0/G1 phase is thought to be a prerequisite for cell differentiation
and apoptosis [69]. Cell cycle is controlled by regulatory proteins, including cyclin-dependent protein kinases (Cdks) and their regulatory subunits, cyclins, as well as
inhibitors such as p21CIP1 and p27KIP1 [72]. Recent studies demonstrated that p21CIP1 and p27KIP1 inhibit the activities of all Cdks and regulate cell proliferation and differentiation [73]. Halevy et al. [74] demonstrated that MyoD, a skeletal muscle-specific transcription regulator, induces cell cycle arrest during skeletal muscle differentiation by increasing the expression of p21CIP1. Using osteoprogenitor cells derived from the bone marrow of p27-/- mice, Drissi et al. [75] demonstrated that p27KIP1 plays a key role in the regulation of osteoblast differentiation by controlling proliferation-related events. Although p21CIP1 and p27KIP1 have been shown to play roles in regulating cell proliferation and differentiation, the functional significance of p21CIP1 and p27KIP1 in modulating the response of osteoblasts to BMP signaling have not been reported. Moreover, the mechanisms that regulate the expression of cell cycle regulatory proteins in osetoblasts in response to BMP signaling have not been fully clarified.
1.3 BMP signaling in tumor biology
In addition to normal cell regulation, some studies have suggested a role for BMP signaling in tumor growth and metastasis. The most striking indication that BMP signaling contributes to tumorigenesis comes from genetic studies of familial tumor syndromes. Mutations of Smad4 and BMPRIA are genetically responsible for familial juvenile polyposis [76-77]. Germ line mutations in BMPRIA have been identified in a subset of families with Cowden syndrome, an inherited breast tumor syndrome [78]. Recent studies even demonstrate that BMP signaling may inhibit the tumorigenic potential of tumor stem cells, including brain and glioma [79-80]. In addition, in prostate tumor tissues, increased levels of expression of BMP-6 and -7 have been correlated with bone metastasis [81-82], while another laboratory reported an inverse correlation between the abnormal expression of BMPRs and the pathological grade of the prostate tumor [83]. The loss of expression of
recurrence, and a decreased rate of survival after surgery [84]. In breast tumor tissues, the expression of BMPRIB has been proposed to be a major marker of progression and de-differentiation [85], while the overexpression of BMP-4 has been shown in colorectal adenomas [86]. Likewise, the coexpression of BMP-2/-4 and BMPRII has been reported to be associated with a poor prognosis in osteosarcoma patients [87]. It has been shown that various sporadic human tumors also exhibit aberrations in BMP signaling (summarized in Table 1-1) [88].
In tissue culture, the effect of BMP signaling varies between cell types and culture conditions. For example, among prostate tumor cell lines, BMP-2 inhibited the proliferation of the androgen-sensitive LNCaP but not the androgen-insensitive PC3 and DU145 cells [89]. Similarly, BMP-4 treatment decreased the rate of proliferation of LNCaP but not PC3 cells [90]. In contrast, BMP-6 inhibited the proliferation of both androgen-sensitive and –insensitive cell lines [84]. More recently, BMP signaling has been shown to enhance bone invasion by prostate tumor cells [91]. Despite the tremendous progress achieved in delineating the functional significance of BMP signaling in tumorigenesis during the last decade, the mechanism of BMP signaling in tumorigenesis is still not very clear. Compelling evidence indicates that the regulation of BMP signaling in tumorigenesis is cell-specific, and both pro-tumor and anti-tumor effects have been described. Thus, the precise effects and mechanisms of BMP signaling in malignant cells must be further demonstrated, and must be interpreted in the context of cell types and experimental conditions.
1.4 Mechanical flow force biology
The importance of microenvironmental dynamic mechanical stresses in tissue development, maintenance, function and pathogenesis has been well established for several decades [92-93]. Fluid flows and pressure gradients that are present in all living tissues
drive blood, lymph, or interstitial flow through blood vessels, lymphatic tube, or extracellular matrix (ECM), respectively [94]. These fluid flows not only help to transport nutrients, drugs, and wastes, but also have important roles in tissue homeostasis and cell biology, including embryonic development, tissue morphogenesis and remodeling, inflammation, lymphedema, tumor biology and immune cell trafficking [94-95]. The mechanisms of mechanical flow forces are extensive studied in vascular cells. Endothelial cells (ECs) are constantly subjected to blood flow-induced shear stress [96-99]. The nature and magnitude of the shear stress plays a significant role in the homeostasis of the structure and function of the blood vessel. Laminar blood flow with a high shear stress modulates cellular signaling and EC functions, and is protective against inflammatory reactions. In contrast, oscillatory blood flow and low shear stress stimulate the expression of EC genes that promote inflammation. This suggests that mechanical flow forces may modulate the intracellular signaling and gene expressions that regulate the ECs’ activities [96-99]. During this process, integrins may be the most likely membrane mechanosensors [100]. Integrins have been shown to play an important role in transmitting mechanical flow forces into chemical signals in a variety of cells seeded on an ECM [100-101]. In several systems, including ECs, shear-activation of integrins leads to an increase in their association with Shc and focal adhesion kinase (FAK), which subsequently activate several intracellular signaling cascades, including ERK signaling [100]. In addition, there is evidence that integrins may be cooperative with the receptors of several growth factors, including insulin receptor and platelet-derived growth factor-β receptor, to form integrin-receptor heteromeric complexes that mediate downstream signaling cascades under shearing [102]. However, the detailed mechanisms of the association between integrins and growth factor receptors under flow are not clear. Scientists also suggest that mechanical flow forces may modulate the association between integrins and membrane receptors, and hence modulate the downstream signaling, in
all cell types, including tumor cells. Recent studies demonstrated that mechanical flow forces may stimulate the Smad pathway to mediate EC differentiation [103-104]. Brown JD et al. demonstrated that shear stress induced Smad2 activation through the MAPK pathway in ECs [103]. However, Wang H et al. reported that shear stress down-regulated Smad2, 3, and 4, and up-regulated Smad7 activities, to induce the differentiation of mesenchymal stem cells into ECs [104]. The detailed mechanisms and correlations between mechanical flow forces and BMP signaling in endothelial cells, or even others cells, is still unclear.
1.5 Mechanical flow forces and tumors
Solid tumors consist of a porous interstitium and a neoplastic vasculature composed of a network of capillaries with highly permeable walls [105-107]. Fluid flows across the vasculature and enters the tumor by convective and diffusive extravasation through the permeable capillary walls. Tumor fluid flows are characterized by spatial and temporal heterogeneities, and can significantly affect tumor growth, metastasis, and therapy [105]. The characteristics of tumor fluid flows are such that (1) fluid flows are diverted away from the center of the tumor toward a more peripheral path, and (2) at a fixed location, the fluid flows are not uniform with time, showing an intermittent flow pattern and flow rate and even periodic inversions of the direction of flow. Despite the important role of tumor fluid flows in tumor physiopathology and treatment, to date there are no complete explanations for the observed tumor fluid flow anomalies.
Although millions of cells are shed from a tumor every day, metastasis is believed to be very inefficient [108]. This inefficiency is widely assumed to be a result of the destruction of cells by both fluid flow and the immune system. The circulation itself represents a highly toxic environment for disseminating tumor cells. The mechanical destruction of circulating tumor cells is the first line of defense in the host microenvironment acting against tumor spreading. Tumor cells circulating in the blood are subjected to intense mechanical stresses
by the shear stress caused by blood flow. Especially in narrow capillaries, the required sphere-to-cylinder shape-transformation is lethal to a majority of tumor cells. These shear forces acting on tumor cells are very intense in small capillaries. At the same time, this deformation of circulating cells within the narrow capillaries enables an intense contact of the tumor cell’s surface adhesion molecules with potential ligands at the capillary walls, and tumor cell adhesion may be initiated. This is caused by the enlarged contacting surface areas of deformed cells, resulting in the increased availability of cell adhesion molecules, and a longer time for the establishment of adhesive bonds. Therefore, the balance between the mechanical destruction and the adhesion initiation by mechanical forces on tumor cells appears to be a regulatory mechanism of tumor spreading [108].
Despite the influence of mechanical flow forces on tumor pathobiology and drug delivery having been studied, the effect of the flow-induced shear force on tumor cells has not been explored much. Compressive forces have been shown to inhibit tumor cell growth [109] and up-regulate adhesion molecules [110]. A recent study reported that tumor cell proliferation is affected by intratumoral pressure, and that the activations of mitogen-activated protein kinases and nuclear antigen Ki-67 are involved in this mechanical modulation [111]. While these results show that mechanical forces can modulate tumor cell responses, the detailed mechanisms by which mechanical stimuli are transduced into cellular signaling to regulate the tumor cell gene expression and functions remain unclear.
Table 1-1 Aberrations of BMP signaling in human tumor, the functional relevance and clinical correlations. (From Hsu MY. et. al., Cancer
CHAPTER II
HYPOTHESIS AND SPECIFIC
AIMS
BMP signaling is a multifunctional mechanism. It is important for bone formation, including osteoblast maturation (i.e., differentiation and proliferation) and tumor cell growth and metastasis inhibition. The BMP-4 synthesized by osteoblasts is one of the most potent inducers of bone formation through its stimulation of osteoblast differentiation. BMP-4 was also demonstrated as a potential growth factor for modulating other cell types’ functions, including normal and tumor cells’ growth or apoptosis. Mechanical flow forces were originally extensively studied in blood biology. Recent studies demonstrate that mechanical flow forces also affect the biophysics and physiology of other cells or tissues, including bone cells and tumor cells. Moreover, it is also suggested that mechanical flow forces may mediate the Smad signaling pathway. In mammals, the regulation of the cell cycle distribution can mediate cell functions; cell proliferation depends on the cell cycle progression, but the cell cycle must be arrested prior to cell differentiation and apoptosis. In light of all this information, we propose that BMP signaling may regulate the cell cycle distribution to mediate osteoblast proliferation and differentiation under BMP-4 treatment, and to mediate tumor cell growth under mechanical flow force stimulation. To test our hypothesis, these seven specific aims were proposed.
1. To observe if BMP-4 regulates the cell cycle distribution in order to induce differentiation in osteoblasts.
2. To investigate if BMP-4 signaling mediates the expression of cell cycle regulators and hence modulates this process.
3. To demonstrate if membrane integrins interact with BMP-4 receptors to optimize the BMP-4 effects.
4. To observe if mechanical flow forces regulate the cell cycle distribution in order to affect tumor cell growth.
regulators through BMP signaling and hence modulate this process.
6. To observe if the mechanical flow force-stimulated BMP signaling comes from BMP autocrine effects.
7. To demonstrate if the mechanosensor, i.e., integrin, response to mechanical flow forces induces the intracellular BMP signaling.
CHAPTER III
3.1 Cell lines and cell culture
Human MG63 osteoblast-like cells were osteosarcoma cells that were isolated in 1977 [112]. The cell line has been identified to possess osteoprogenitor potential for differentiation in response to differentiation signals, tumor cell character with rapid proliferation, and no aging [112-113]. Therefore, the MG63 cells were cultured to study the BMP-4 and mechanical flow force effects in the present thesis. Mouse MC3T3-E1 osteoblast-like cells were a positive control for the BMP-4 treatment experiments. Saos2 osteosarcoma cells, SCC25 oral carcinoma cells, and SW1353 chondrosarcoma cells were positive controls for the mechanical flow force experiments. All cell lines were obtained from American Type Culture Collection (Rockville, MD) and cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, Gaithersburg, MD) supplemented with 10% fetal bovine serum (FBS; Gibco). In the BMP-4 treatment experiments, cells (~1-2×105 cells/cm2)
were trypsinized and seeded onto the 60 mm culture dish. The medium was exchanged with a medium that was identical except that it contained only 0.5% FBS, and the cells were further incubated for 24 h before treatment with BMP-4. In the mechanical flow force experiments, cells (~1-2×105 cells/cm2) were trypsinized and seeded onto glass slides (75x38
mm; Corning, NY) pre-coated with type I collagen (30 μg/ml). The medium was then exchanged with DMEM containing only 2% FBS for the 24 h incubation of the cells prior to the experiment.
3.2 Materials
Mouse monoclonal antibodies (mAbs) against cyclin E (sc-25303), Cdk2 (sc-748), ERK2 (sc-1647), phospho-ERK (sc-7383), and β3 integrin (sc-46655), and goat polyclonal
antibodies (pAbs) against Smad1/5 (sc-6031), Runx2 (sc-12488) and αv integrin (sc-10719),
cyclins A (#4656), B1 (#4135), and D1 (#2926), Cdk-4 (#2906) and -6 (#3136), and p21CIP1 (#2946), and rabbit pAbs against p27KIP1 (#2552), Cdk1 (#9112), phospho-Cdk1 (Y15) (#9111), and phospho-Smad1/5 (#9511), were purchased from Cell Signaling Technology (Beverly, MA). Mouse mAbs against αvβ3 (MAB1976) andβ1 (MAB2253) integrins were
purchased from Chemicon (Temecula, CA). The dominant-negative mutants of Shc (Shc-SH2) and FAK [FAK (F397Y)] were previously described [114-115]. The OCN promoter construct was a gift from Dr. Leland WK Chung (Department of Urology, Emory University School of Medicine). The control siRNA and specific siRNAs of BMPRIA, BMPRIB, Smad1, Smad5, p21CIP1, p27KIP1, ERK, and αv, β1, and β3 integrins were purchased
from Invitrogen (Carlsbad, CA). Recombinant human BMP-4 was purchased from R & D Systems (Minneapolis, MN). All other chemicals of reagent grade were obtained from Sigma (St Louis, MO), unless otherwise noted.
3.3 Flow apparatus.
The glass slide with cultured tumor cells was mounted in a parallel-plate flow chamber characterized and described in detail elsewhere (Figure 3-1) [116]. The chamber was connected to a perfusion loop system, kept in a constant-temperature controlled enclosure, and maintained at pH 7.4 by continuous gassing with a humidified mixture of 5% CO2 in air.
The fluid shear stress (τ) generated on the cells by the flow was estimated to be 12 dynes/cm2, unless otherwise noted, using the formula τ = 6μQ/wh2, where μ is the dynamic viscosity of
the perfusate, Q is the flow rate, and h and w are the channel height and width, respectively.
3.4 Flow cytometric analysis.
The cells were harvested in PBS containing 2 mM ethylenediaminetetraacetic acid, washed once with PBS, and fixed for 30 min in cold ethanol (70%). Fixed cells were washed and permeabilized with 0.1% Triton X-100 in PBS. They were then stained with 50
μg/ml of propidium iodide (Roche, Basel, Switzerland) and 1 mg/ml RNase A for 30 min. Stained cells were analyzed with a fluorescence-activated cell sorter (FACS) Calibur (Becton-Dickinson, Franklin Lakes, NJ), and the data were analyzed using a mod-fit cell cycle analysis program.
3.5 ALP specific activity assay.
The cell extract was prepared with 0.1% Triton X-100 after the shear stress experiments. Cellular ALP activity was assayed at the end of the incubation with 10 mM p-nitrophenyl phosphate in 0.15 M sodium carbonate buffer (pH 10.3) and 1 mM MgCl2, as previously
described [117], and was normalized against the cellular protein determined by the Bio-Rad protein assay.
3.6 RNA isolation and quantitative real-time PCR.
The total RNA was isolated by the guanidium isothiocyanate/phenochloroform method and converted to cDNA as described [97]. The cDNA was amplified through PCR on a LightCycler (Roche Diagnostics, East Sussex, United Kingdom) using LightCycler FastStart DNA MasterPlus SYBR Green I (Roche Diagnostics) with 0.5 µM primers of OCN (sense: 5′-TGAGAGCCCTCACACTCCTC-3′; antisense: 5′-ACCTTTGCTGGACTCTGCAC-3′; product length, 98 bp), ALP (sense: 5′- CAACCCTGGGGAGGAGAC-3′; antisense: 5′-GCATTGGTGTTGTACGTCTTG-3′; product length, 78 bp), and β-actin (sense: 5′-AAATCGTCCGTGACATCAAG-3′; antisense: 5′- GGAAGGAAGGCTGGAAGAGA -3′; product length, 180 bp) genes. PCR was performed in triplicate at 95°C for 10 min followed by 45 cycles of denaturation at 95°C for 10 sec, annealing at 60°C for 5 sec, extension at 72°C for 8 sec, and single signal acquisition for 10 sec. β-actin gene expression was used as an internal control. The PCR conditions were optimized to obtain a PCR product with a
the LightCycler were analyzed using LightCycler Software Version 3.5 (Roche Diagnostics). The gene expression levels were normalized with the β-actin gene expression levels in the same sample.
3.7 Western blot analysis.
The cells were collected by scraping and lysed with a buffer containing 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, and a protease inhibitor mixture (PMSF, aprotinin, and sodium orthovanadate). The total cell lysate (100 µg of protein) was separated by SDS-polyacrylamide gel electrophoresis (PAGE) (10% running, 4% stacking) and transferred onto a polyvinylidene fluoride membrane (Immobilon P, 0.45-µm pore size). The membrane was then incubated with the designated antibodies. Immunodetection was performed using the Western-Light chemiluminescent detection system (Applied Biosystems, Foster City, CA).
3.8 Immunoprecipitation.
The cells were scraped and lysed with a buffer containing 25 mM HEPES, pH 7.4, 1% Triton X-100, 1% deoxycholate, 0.1% SDS, 0.125 M NaCl, 5 mM EDTA, 50 mM NaF, 1 mM Na3VO4, 1 mM PMSF, 10 mg/mL leupeptin, and 2 mM BGP. The cells were disrupted on
ice by repeated aspiration through a 21-gauge needle. The same amount of protein from each sample was incubated with a designated antibody for 2 h at 4°C with gentle shaking. The immune complex was then incubated with protein A/G plus agarose for 1 h and collected by centrifugation. These agarose-bound immunoprecipitates were washed and incubated with boiling sample buffer containing 62 mM Tris-HCl, pH 6.7, 1.25% (w/v) SDS, 10% (v/v) glycerol, 3.75% (v/v) mercaptoethanol, and 0.05% (w/v) bromphenol blue. The samples were then subjected to SDS-PAGE and Western blotting.
3.9 Treatments with RGD peptides and mAbs.
Type I collagen contains the integrin-recognition tripeptide RGD (Arg-Gly-Asp) sequence [118]. To block specific integrin-collagen interactions, the cells were pre-incubated with the tetrapeptide RGDS (Arg-Gly-Asp-Ser; 500 μg/mL), which blocks cell adhesion through the RGD sequence on ECM proteins, or the antibodies (10 μg/mL) against αvβ3 and β1 integrins for 2 h before seeding onto the glass slides pre-coated with type I
collagen and during the application of fluid flow.
3.10 Reporter gene construct, DNA plasmids, siRNA, transfection, and luciferase assay. The OCN promoter construct (OCN-Luc) contains 800 bp of OCN 5′-flanking DNA linked to the firefly luciferase reporter gene of the plasmid pGL3 (Promega Inc.) [119]. This fragment of the OCN promoter contains Runx2 binding sites. DNA plasmids at a concentration of 1 μg/mL were transfected into MG63 cells at 60% confluence by using lipofectamine (Gibco). The pSV-β-galactosidase plasmid was co-transfected to normalize the transfection efficiency. The cells were kept as static controls or subjected to shear stress experiments 48 h after transfection. For siRNA transfection, MG63 cells at 70-80% confluence were transfected with the designated siRNA at various concentrations (5, 15, 30, and 40 nM) using the RNAiMAX transfection kit (Invitrogen, Carlsbad, CA).
3.11 Electrophoretic mobility shift assay (EMSA).
The cells were collected by scraping in PBS. After centrifugation of the cell suspension at 2000 rpm, the cell pellets were resuspended in cold buffer A (containing, in mmol/L, KCl 10, ethylenediamine tetraacetate [EDTA] 0.1, dithiothreitol [DTT] 1, and phenyl methylsulfonyl fluoride [PMSF] 1) for 15 min. The cells were lysed by adding 10% NP-40 and then centrifuged at 6000 rpm to obtain pellets of nuclei. The nuclear pellets were
ethane-sulfonic acid [HEPES] 20, EDTA 1, DTT 1, PMSF 1, and NaCl 400), vigorously agitated, and then centrifuged. The supernatant containing the nuclear proteins was used for the EMSA or stored at -70°C until used. A sequence of 20 bp-oligonucleotides containing the human OCN Runx2 site was synthesized (5′-CGTATTAACCACAATACTCG-3′ and 5’- AATTGGTGTTATGAGCATGC-3’) [120]. The oligonucleotides were end-labeled with [γ-32P]ATP. The extracted nuclear proteins (10 µg) were incubated with 0.1 ng 32P-labeled
DNA for 15 min at room temperature in 25 µL binding buffer containing 1 µg poly(dI-dC). In the antibody supershift assay, an antibody against Runx2 (1 μg each; Cell Signaling Technology) was incubated with the mixture for 10 min at room temperature, followed by the addition of the labeled probe. The mixtures were electrophoresed on 5% nondenaturing polyacrylamide gels. The gels were dried and imaged by autoradiography.
3.12 Statistical analysis.
The results are given as the mean±SEM. Statistical tests were performed with an independent Student t-test for two groups of data and the analysis of variance (ANOVA) followed by Scheffe’s test for multiple comparisons. A P value < 0.05 was considered significant.
Cell culture Figure 3-1
Figure 3-1 Schematic diagram of the cell culture flow model. Tumor cells were seeded on the type I collagen gel on the glass slide.
CHAPTER IV
RESULTS
4.1 Bone Morphogenetic Protein-4 Induces Osteoblast G0/G1 Arrest and Differentiation
via Increased Expression of p21CIP1 and p27KIP1: Roles of integrins, extracellular signal-regulated kinase, and Smad
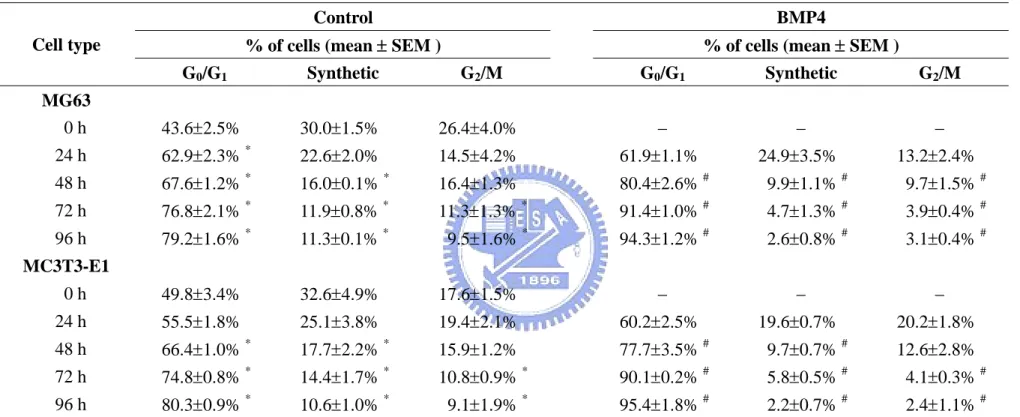
4.1.1 BMP-4 induces G0/G1 arrest and increased expression of p21CIP1 and p27KIP1 in osteoblast-like cells. To determine the regulatory effect of BMP-4 on the cell cycle in osteoblasts, human MG63 and mouse MC3T3-E1 osteoblast-like cells were kept as controls or treated with BMP-4 (25 ng/ml) for 24, 48, 72, and 96 h, and their cell cycle distributions were analyzed using flow cytometry. The incubation of these cells under static conditions for 48, 72, or 96 h led to an increase in the cell percentage in G0/G1 phases and a decrease in
synthetic and/or G2/M phases (Table 4-1). The treatment of these cells with BMP-4 caused a
significant increase in the cell percentage in the G0/G1 phases and decreases in the synthetic
and/or G2/M phases compared with the untreated control cells for the same periods. These
results suggest that BMP-4 induces G0/G1 arrest in these osteoblast-like cells. The changes
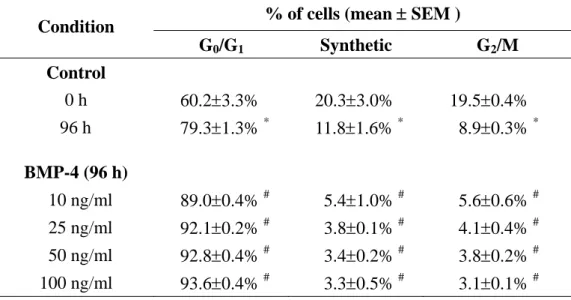
of the cell cycle distribution in MG63 cells induced by BMP-4 were consistent with the concentration range of the BMP-4 used (i.e., 10, 25, 50, and 100 ng/ml), indicating that the BMP-4-induced G0/G1 arrest was dose-independent over the range tested (Table 4-2).
We investigated the molecular basis of this BMP-4 effect, and the presentation is focused on the human MG63 cells. The treatment of the MG63 cells with BMP-4 (25 ng/ml) for 24, 48, 72, or 96 h resulted in increases in p21CIP1 and p27KIP1 expression in these cells (Figure 4-1). In contrast, BMP-4 did not have effects on the expression of cyclins A, B1, D1, and E, and Cdk-2, -4, and -6 in the MG63 cells.
4.1.2 BMP-4-induced MG63 cell differentiation is mediated by p21CIP1 and p27KIP1. Since cell cycle regulator-led inhibition of the G0/G1-to-synthetic phase transition has been shown to
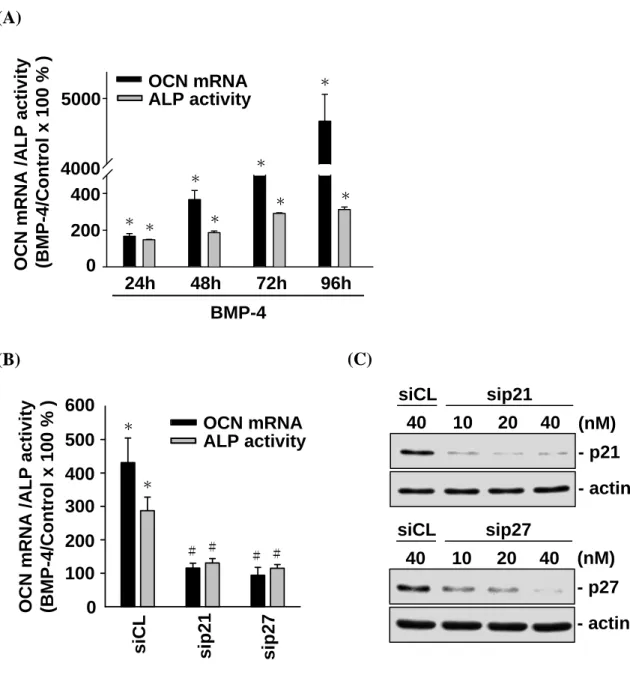
G0/G1 arrest in MG63 cells with concomitant increases in p21CIP1 and p27KIP1 expression, we
investigated whether the BMP-4-induced MG63 cell differentiation is mediated by p21CIP1 and p27KIP1. MG63 cells were kept as controls or treated with BMP-4 (25 ng/ml) for 24, 48, 72, and 96 h, and their expression or activity levels for differentiation markers, i.e., osteocalcin (OCN) and alkaline phosphatase (ALP), were examined. Treatment with BMP-4 for 24, 48, 72, and 96 h resulted in significant increases in OCN gene expression and ALP activity in MG63 cells (Figure 4-2A). These BMP-4-induced increases in OCN expression and ALP activity were abolished by the transfection of cells with p21CIP1- and p27KIP1-specific siRNAs (40 nM for each) (Figure 4-2B), which had 80-90% blocking effects on their respective protein expressions (Figure 4-2C). These results suggest that the BMP-4-induced MG63 cell differentiation is mediated by p21CIP1 and p27KIP1.
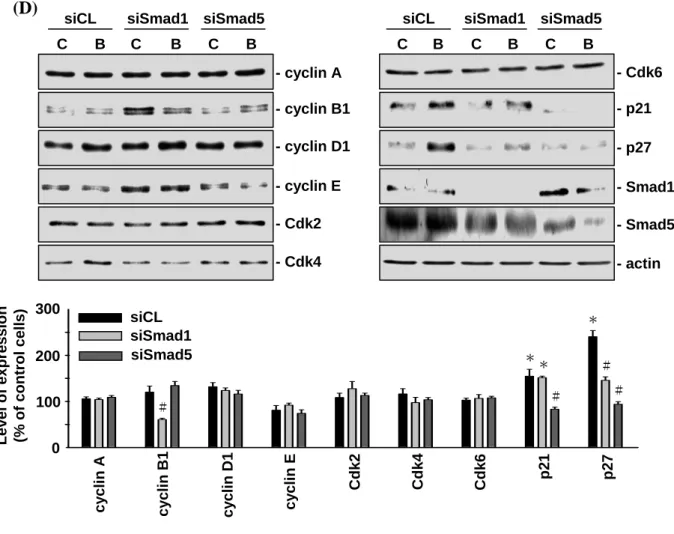
4.1.3 The BMP-4-induced p21CIP1 and p27KIP1 expression and G0/G1 arrest in MG63 cells is mediated by the BMPRIA/Smad5 pathway. The treatment of MG63 cells with BMP-4 (25 ng/ml) induced a rapid increase (within 30 min) in Smad1/5 phosphorylation, which reached a maximal level ≈5 times that of untreated controls within 1 h, and then declined but remained elevated after 24 h of treatment (Figure 4-3A). The increases in Smad1/5 phosphorylation were similar with BMP-4 concentrations of 25, 50, and 100 ng/ml, indicating that the BMP-4-induced Smad1/5 activation was dose-independent over the range tested (Figure 4-3B). The transfection of MG63 cells with BMPRIA-specific siRNA (40 nM, compared with control siRNA) abolished the BMP-4-induced Smad1/5 phosphorylation (Figure 4-3C). In contrast, BMPRIB-specific siRNA did not have inhibitory effects on the BMP-4-induced Smad1/5 phosphorylation. These results suggest that the BMP-4-induced Smad1/5 activation is mediated by BMPRIA, but not BMPRIB. BMPRIA- and BMPRIB-specific siRNAs almost totally abolished their respective receptor protein expressions (Figure 4-3C). To investigate the role of Smad1/5 in the BMP-4-modulation of cell cycle regulatory
protein expression in the MG63 cells, the cells were transfected with Smad1- or Smad5-specific siRNA (40 nM), which reduced the expressions of the corresponding Smad proteins by ≈80% compared to that with control siRNA (Figure 4-3D), and the cells were then kept as controls or treated with BMP-4 for 48 h. The transfection with Smad5-specific siRNA (compared with control siRNA) resulted in a significant inhibition of the BMP-4-induced up-regulation of p21CIP1 and p27KIP1 expression (Figure 4-3D). In contrast, the transfection with Smad1-specific siRNA only inhibited the BMP-4-induced up-regulation of p21CIP1 expression. To elucidate the importance of BMPRIA and Smad5 in modulating the BMP-4-induced G0/G1 arrest, MG63 cells were transfected with BMPRIA- or
Smad5-specific siRNA (40 nM) and then kept as controls or treated with BMP-4 for 72 h. For unstimulated cells, the transfection with BMPRIA- or Smad5-specific siRNA (compared with control siRNA) did not alter their cell cycle distribution (Table 4-3). After BMP-4 treatment, the MG63 cells transfected with either BMPRIA- or Smad5-specific siRNA had a significantly lower cell percentage in the G0/G1 phases and a higher cell percentage in
synthetic and G2/M phases, as compared with cells transfected with the control siRNA (Table
4-3). Taken together, these results suggest that the BMP-4-induced p21CIP1 and p27KIP1 expression and G0/G1 arrest in MG63 cells is mediated through the BMPRIA/Smad5 pathway.
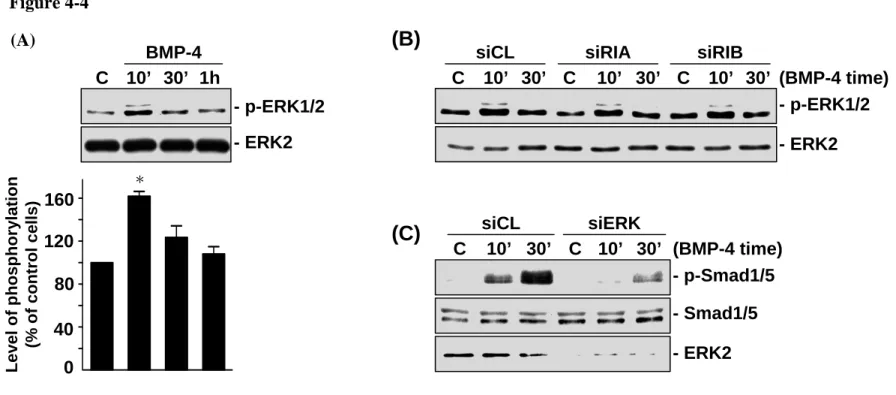
4.1.4 ERK mediates the BMP-4-induced Smad phosphorylation in MG63 cells. The ERK pathway is known to regulate gene expression and cellular functions, notably cell proliferation and differentiation [121]. We investigated the role of ERK in the BMP-induced Smad signaling in MG63 cells. The MG63 cells stimulated with BMP-4 (25 ng/ml) induced a rapid increase in ERK phosphorylation within 10 min after stimulation (Figure 4-4A). This increased level of phosphorylation decreased to nearly the basal level after 30 min post- BMP-4 stimulation. The transfection of MG63 cells with BMPRIA- and BMPRIB-specific
that the BMP-4-induced ERK activation is not mediated by the BMP receptor-signaling pathway. However, when MG63 cells were transfected with ERK-specific siRNA (40 nM), which caused 89-90% reduction in the protein expression of ERK, their BMP-4-induced Smad1/5 phosphorylation was significantly reduced, as compared with the cells transfected with control siRNA (Figure 4-4C). These results suggest that ERK mediates the BMP-4-induced Smad1/5 activation in MG63 cells.
4.1.5 The BMP-4-induced ERK and Smad phosphorylations in MG63 cells are mediated by
β3, but not αv and β1, integrins. To elucidate the role of αv, β1, and β3 integrins in the
BMP-4-induced ERK and Smad1/5 activation in MG63 cells, the cells were transfected with αv-, β1-, and β3-specific siRNAs (40 nM for each), and then kept as controls or stimulated
with BMP-4 (25 ng/ml) for 10 and 30 min. The BMP-4-induced ERK and Smad1/5 phosphorylations were inhibited by transfections of MG63 cells with β3-specific siRNA
(compared with control siRNA), but not αv- and β1-specific siRNAs, suggesting that the
BMP-4-induced ERK and Smad1/5 activations are mediated by β3, but not αv and β1,
integrins (Figure 4-5A). The αv-specific siRNA had 60-70% blocking effects on the αv
integrin expression; the β1- and β3-specific siRNAs almost totally abolished their respective
integrin expressions (Figure 4-5A). We further investigated the interaction between the integrins and the BMP receptors under the unstimulated control condition or in response to BMP-4. MG63 cells were kept as controls or treated with BMP-4 (25 ng/ml) for 10 min, and their extracts were immunoprecipitated with an antibody against BMPRIA or BMPRIB, followed by Western blot analysis with antibodies against the αv, β1, and β3 integrins. In the
unstimulated cells, αv, β1, and β3 integrins showed constitutive associations with BMPRIA
and BMPRIB (Figure 4-5B). The BMP-4 stimulation resulted in a significant decrease in the association of BMPRIA with the β1 integrin, as well as BMPRIB with the β1 and β3
integrins. BMP-4 did not affect the association of BMPRIA with the αv and β3 integrins, as
well as BMPRIB with the αv integrin.
4.2 Tumor cell cycle arrest induced by shear stress: role of integrins and Smad
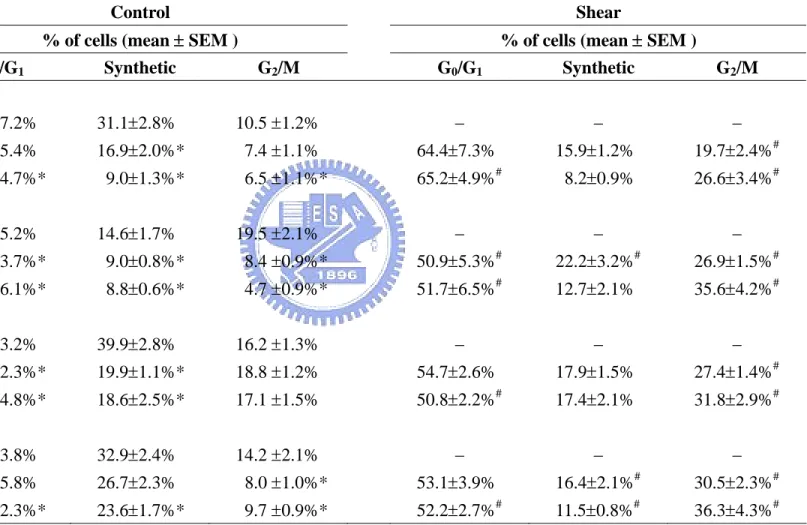
4.2.1 Shear stress induces G2/M arrest and the corresponding changes in cell cycle regulatory protein expression in tumor cells. The effects of shear stress on cell cycle distribution were studied in four tumor cell lines (human MG63 and Saos2 osteosarcoma cells, SCC25 oral squamous carcinoma cells, and SW1353 chondrosarcoma cells). Cells were kept as controls or subjected to shear stress (12 dynes/cm2 for 24 and 48 h). Flow cytometry showed that the incubation of these tumor cells under static conditions for 24 or 48 h led to increases in the cell % in G0/G1 phases and decreases in the synthetic and/or G2/M phases
(Table 4-4), indicating a G0/G1 arrest. The application of shear stress to these tumor cells
caused significant increases in the cell % in the G2/M phases and decreases in the G0/G1
phases, as compared with cells under static conditions for the same periods.
We investigated the molecular basis of this shear effect, and the presentation is focused on human MG63 cells. The application of shear stress to MG63 cells for 24 or 48 h increased cyclin B1 and p21CIP1 expression and decreased Cdk1 expression (Figure 4-6). The decrease in Cdk1 expression was accompanied by an increase in its tyrosine 15 phosphorylation, indicating a shear-induced decrease in Cdk1 activity [122]. Shear stress also decreased the expression of cyclins A, D1, and E, Cdk-2, -4, and -6, and p27KIP1.
4.2.2 Shear stress induces sustained phosphorylation of Smad1/5 in MG63 cells through BMPRIA. The application of shear stress to MG63 induced a rapid increase (within 10 min) in Smad1/5 phosphorylation, which reached a maximal level of ~10 times static controls within 1 h, and then declined but remained elevatedafter 24 h of shearing (Figure 4-7A).
The increases in Smad1/5 phosphorylation were similar with shear stresses of 2, 12, and 20 dynes/cm2, indicating that the shear-induced Smad1/5 activation was shear dose-independent over the range tested (Figure 4-7B). Pre-treating MG63 cells with Noggin, a specific antagonist that binds BMPs to block their binding to the BMP receptors, did not inhibit the shear-induced Smad1/5 phosphorylation; hence, the shear-activation of Smad1/5 was not mediated by BMPs (Figure 4-7C). As a positive control, the pre-treatment of MG63 cells with Noggin did cause an inhibition of the Smad1/5 phosphorylation induced by BMP-4.
To study the types of BMP receptors responsible for the shear-activation of Smad1/5, MG63 cells were transfected with BMPRIA- or BMPRIB-specific small interfering RNA (siRNA, 40 nM), which reduced the expressions of the corresponding receptor proteins by ~2/3 of those with control siRNA (Figure 4-7D), and the cells were then exposed to shear stress for 30 min. The shear-induced Smad1/5 phosphorylation was abolished by the BMPRIA-specific siRNA, but was not inhibited by the BMPRIB-specific siRNA (Figure 4-7E).
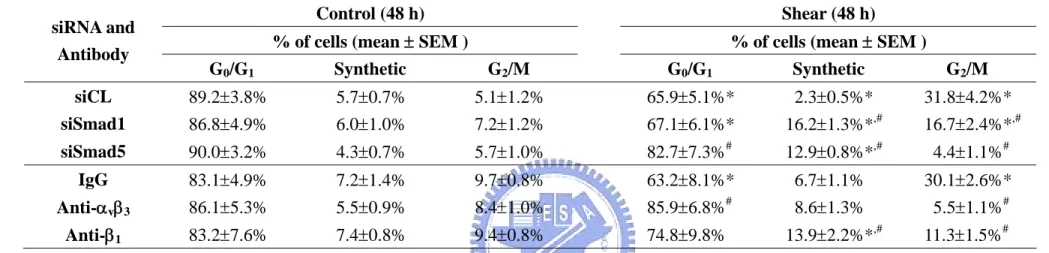
4.2.3 Shear-induced G2/M arrest in tumor cells is mediated by Smad1/5. MG63 cells were transfected with Smad1- or Smad5-specific siRNA (40 nM), which caused 80-90% reductions in expressions of the corresponding Smads (Figure 4-8A), and the cells were then kept under static conditions or exposed to flow for 48 h. Under static conditions, the MG63 cells transfected with Smad1- or Smad5-specific siRNA (compared to control siRNA, 40 nM) did not alter their cell cycle distribution (Table 4-5). Following shear stress, the MG63 cells transfected with either Smad5- or Smad1-specific siRNA had a significantly higher cell % in the G0/G1 and synthetic phases and a lower cell % in the G2/M phases, as compared to cells
transfected with control siRNA (Table 4-5). The transfection with Smad1- or Smad5-specific siRNA (compared to control siRNA) resulted in a significant inhibition of the shear-induced up-regulation of Cdk1 tyrosine 15 phosphorylation (Figure 4-8B).
Smad1-specific and Smad5-specific siRNAs have some differential actions, with the former inhibiting the shear-induced cyclin A down-regulation and cyclin B1 up-regulation, and the latter inhibiting the shear-induced p21CIP1 up-regulation. Neither Smad-specific siRNA had significant effects on the shear-induced changes of the other cell cycle regulatory proteins (Figure 4-8B).
4.2.4 αvβ3 and β1 integrins mediate shear-induced Smad1/5 phosphorylation and G2/M arrest in tumor cells. MG63 cells were pre-treated with RGDS (Arg-Gly-Asp-Ser), which blocks the cell-ECM interaction mediated by the integrin-recognition sequence RGD (Arg-Gly-Asp) on ECM proteins, or with specific antibodies against the αvβ3 and β1 integrins,
and were then kept under static condition or exposed to flow for 30 min. Pre-treatment with RGDS and integrin antibodies significantly inhibited the shear-induced Smad1/5 phosphorylation, as compared to cells pre-treated with control RGES or IgG (Figure 4-9A). The inhibition of shear-induced Smad1/5 phosphorylation by blocking the αvβ3 and β1
integrins was substantiated by the transfection of cells with αv-, β1-, and β3-specific siRNAs
(40 nM for each), which also showed significant inhibitory effects on the shear-induced Smad1/5 phosphorylation (Figure 4-9B). The αv- and β3-specific siRNAs had 50-60%
blocking effects on their respective integrin expressions; the β1-specific siRNA almost totally
abolished the β1 expression (Figure 4-9C). The transfection with dominant-negative mutants
of Shc and focal adhesion kinase (FAK) (compared to empty vector pcDNA3) did not have significant effects on the shear-induced Smad1/5 phosphorylation (Figure 4-9D). The pre-treatment of MG63 cells with specific antibodies against αvβ3 and β1 did not affect the
cell cycle distribution in static control cells, but under shear stress they caused a significant increase in the cell % in G0/G1 or synthetic phases and a decrease in the G2/M phases (Table