國立台灣大學獸醫專業學院獸醫學研究所 碩士論文
Graduate Institute of Veterinary Medicine College of Veterinary Medicine National Taiwan University
Master Thesis
犬 YKL-40 重組蛋白之表現 及其在犬乳腺腫瘤細胞之作用
Recombinant Canine YKL-40 Expression and Its Effects on Canine Mammary Gland Tumor Cells
林均豫 Chun-Yu Lin
指導教授:廖泰慶 博士
Advisor: Albert Taiching Liao, D.V.M., Ph.D.
中華民國 102 年 7 月
July 2013
獻給 林健彥 我的父親 楊淑慧 我的母親 感謝這一路支持我走來的你們
謝謝廖泰慶老師一直以來的指導,總是殷殷期盼不算認真的我能有所表現,
我確實在這兩年學到了相當豐富的知識和技術,這是當初選擇讀研究所時未曾想 過的意外收穫,這樣的學習使我充實,也許對於未來的人生旅程亦是受益良多。
謝謝林辰栖老師提點關於實驗的寶貴經驗與訣竅,讓我能不局限於某一角度思考。
謝謝實驗室的各位夥伴,這實驗室實在有著非常熱於分享經驗的學長姊,非常能 共同努力的同學,以及非常認真幫忙的學弟妹,這樣的相處氛圍好到讓人會以身 為這大家庭的一份子而驕傲,除了感謝之外,希望大家能把這種感動持續下去,
持續保持這麼好的研究環境。
最要感謝的還是我的父母親,我的求學歷程確是不盡順利,若沒有他們全心 全力的支持與理解,我無法走到今天,爸媽,謝謝你們。
最後謝謝為我加油的朋友們,謝謝讓我選擇進入臺大讀研究所的你。
Contents
中文摘要 ... I
Abstract ... II
Chapter 1 Background and Literatures Review ... 1
1.1 YKL-40 ... 1
1.1.1 Expression and regulation of YKL-40 ... 2
1.1.2 Biological properties of YKL-40 ... 6
1.1.3 YKL-40 in infectious, chronic inflammation and joint diseases ... 9
1.1.4 YKL-40 in cancer diseases ... 10
1.1.5 YKL-40 may be an autoantigen ... 11
1.1.6 YKL-40 in cell migration ... 12
1.1.7 Possible cell signal pathway of YKL-40 ... 13
1.2 Canine mammary gland tumor ... 13
1.3 Conclusion ... 14
Chapter 2 Introduction ... 16
Chapter 3 Material and Method ... 19
3.1 Experimental animals ... 19
3.2 Cell lines ... 19
3.3 Anitbodies ... 20
3.4 Total RNA extraction ... 20
3.5 RT-PCR ... 21
3.6 Preparation and propagation of YKL-40/pcDNA 3.1 plasmid... 22
3.7 Transfection of canine YKL-40 gene into BALB/3T3 cells ... 22
3.8 Expression of canine YKL-40 protein in BALB/3T3 cells ... 23
3.9 Concentration of the recombinant canine YKL-40 protein ... 23
3.10 Purification of the recombinant canine YKL-40 protein... 24
3.11 Quantification of the recombinant canine YKL-40 protein ... 24
3.12 Identification of the purified recombinant canine YKL-40 protein ... 25
3.13 Western blot analysis ... 25
3.14 Proliferation assay ... 26
3.15 Migration assay ... 26
3.16 Invasion assay ... 27
3.17 Autoantigen analysis ... 27
3.18 Cell signal pathway analysis ... 28
Chapter 4 Results ... 30
4.1 YKL-40 protein expression by stable cell line establishment ... 30
4.2 YKL-40 mRNA expression examination ... 30
4.3 Purification of the recombinant canine YKL-40 ... 31
4.4 Proliferation assay ... 32
4.5 Migration assay ... 32
4.6 Invasion assay ... 33
4.7 Cell signal pathway analysis ... 33
Chapter 5 Discussion ... 35
Tables ... 40
Table 1. Reverse transcription protocol ... 40
Table 2. YKL-40 primer list ... 41
Figures ... 42
Figure 1. The construct of canine YKL-40/pcDNA 3.1/V5-HIS-TOPO. ... 42
Figure 2. Expression of the YKL-40 mRNA in different tumor cell lines. ... 43
Figure 3. The transfection of YKL-40 into BALB/3T3 cells ... 44
Figure 4. Purification of the recombinant YKL-40 protein with His-column and Heparin-column ... 45
Figure 5. YKL-40 increases the proliferation rate of CMT-1 and MPG cells. ... 46
Figure 6. The result of cell migration assay on CMT-1 cells ... 47
Figure 7. The result of cell migration assay on MPG cells ... 48
Figure 8. YKL-40 stimulates the invasion ability of CMT-1 cells. ... 50
Figure 9. YKL-40 stimulates the invasion ability of CMT-1 cells. ... 52
Figure 10. Autoantigenicity of YKL-40 ... 53
Figure 11. The PI3K/AKT and MAPK pathway analysis on CMT-1 and MPG cells ... 54
References ... 55
I
中文摘要
YKL-40 首次被發現於人類的骨瘤細胞株 MG63 中,為一分泌型之醣蛋白。
其命名來自其蛋白胺基酸序列 N 端之前三個胺基酸,分別是酪胺酸(Y)、賴胺酸
(K)以及白胺酸(L),取其縮寫並加上其分子重量約為 40 kDa,因此命名為 YKL-40。
此 外 由 於 YKL-40 具 有 類 似 幾 丁 酶 可 結 合 幾 丁 質 之 能 力 而 有 別 名 CHI3L1
(chitinase-3-like-1),但其主要作用區域之胺基酸由白胺酸(leucine)所取代,因 而失去幾丁酶之活性。過往關於 YKL-40 的研究皆以人類疾病及實驗動物為主,與 犬隻相關的研究發表屈指可數。而在 YKL-40 與癌症相關的研究中,一般認為血清 中增量之 YKL-40 與許多種類之癌症及預後不良有關。近期的研究也指出 YKL-40 具有調控血管新生的能力。然而 YKL-40 確切的功用與作用機制仍尚未明瞭,更遑 論犬隻 YKL-40 的研究仍屬稀少。本研究將以 BALB/3T3 細胞進行轉染帶有犬隻 YKL-40 基因的載體,透過真核系統之細胞株穩定表現此分泌型醣蛋白。合成後的 蛋白會被分泌至培養液中,收集後經濃縮後進行純化。之後利用犬隻乳腺癌細胞 株 CMT-1 及 MPG 來進行 YKL-40 生物功能的測試與評估,包括:細胞增生試驗、
細胞移行試驗以及細胞侵入能力試驗等。在最低 100 ng/mL 的濃度下,YKL-40 對 於促進細胞移行及侵入具有一定的效果,然而在細胞增生的能力並不顯著。而在 細胞訊息傳遞路徑的探索上,發現 YKL-40 具有活化 AKT 的能力,推測其主要透 過此一路徑進行訊息傳遞。
關鍵字:YKL-40、CHI3L1、犬乳腺腫瘤
II
Abstract
YKL-40 was first identified as a secretory glycoprotein from a human osteosarcoma cell line, MG63. This protein is named based on its molecular weight of 40 kDa and N-terminal amino acids tyrosine (Y), lysine (K), and leucine (L). It is also known as CHI3L1 (chitinase-3-like-1), which has the chitin-binding affinity but lacks the chitinase activity due to the substitution of an essential glutamic acid with leucine in the chitinase-3-like catalytic domain. During the past decade, most of YKL-40 studies were focused on human diseases and experimental animal models such as mice, and little information of canine research was presented so far. Elevated serum YKL-40 has been associated with worse prognosis in a variety of human cancers. Recent studies suggested that YKL-40 plays a role in regulation of tumor angiogenesis. However, the real function and molecular mechanism of YKL-40 still remained unknown, and the investigation on canine source YKL-40 protein was also rare. We have reported previously the molecular cloning of the canine YKL-40 gene and a predicted YKL-40 protein structure. In this study, a transfection was performed on BALB/3T3 cell to establish a stable cell line which can overexpress canine YKL-40 protein. Recombinant canine YKL-40 protein was then concentrated and purified from the cultured medium of
III
the transfected cell line. The biological functions of this protein were evaluated on the canine mammary gland tumor (MGT) cells including proliferation, migration and invasion assays, etc.. YKL-40 at a concentration as low as 100 ng/mL seem to has effect in cell migration and invasion, yet little effect in cell proliferation. The alterations of molecular signaling induced by addition of YKL-40 protein will be also evaluated on canine MGT cells.
Keyword: YKL-40, CHI3L1, canine mammary gland tumor
1
Chapter 1 Background and Literatures Review
1.1 YKL-40
This glycoprotein was identified secreted in human osteosarcoma cell line MG63 (Johansen et al. 1992), and also discovered in differentiated swine smooth muscle cells from swine explants of the thoracic aorta (Millis et al. 1985), human synovial cells and chondrocytes (Hakala et al. 1993), bovine mammary secretions during the non-lactating period (Rejman et al. 1988), murine breast tumor cells (Morrison et al.
1994), and caprine mammary gland secretion with known crystal 3D structure and nucleotide sequence (Mohanty et al. 2003). The protein was named YKL-40 based on its three N-terminal amino acids Tyrosine (Y), Lysine (K) and Leucine (L) and its molecular mass of 40 kDa (Johansen et al. 1992). The protein also has several different names: YKL-40 (Johansen et al. 1992), Human Cartilage glycoprotein-39 (HC gp39) (Hakala et al. 1993), Breast regressing protein 39 Kd (brp-39) (Morrison et al. 1994), 38-kDa heparin-binding glycoprotein (gp38k) (Shakelton et al. 1995), Chitinase-3-like-1 (CHI3L1) (Rehli et al. 1997), Chondrex (Harvey et al. 1998), and 40 kDa mammary gland protein (MGP-40) (Mohanty et al. 2003). The protein is categorized in glycosyl hydrolase family 18 as a chitolectin (Henrissat et al. 1993)
2
without chitinase activites (Hakala et al. 1993; Renkema et al. 1998). Most of the YKL-40 studies are based on human researches, from inflammatory diseases to cancer diseases, and some involved the mouse as a laboratory animal (Johansen 2006). Since rarely investigation of canine YKL-40 was presented so far, we’re eagerly to known further about this protein and hope there will in the future be more research in this field.
1.1.1 Expression and regulation of YKL-40
YKL-40 protein and mRNA are expressed in vitro and in vivo in several different cells, including non-malignant cells and malignant cancer cells.
1.1.1.1 Expression in non-malignant cells
Different cells originated from ectoderm, mesoderm, and endoderm express YKL-40 protein and mRNA in certain conditions (Johansen 2006).
1.1.1.1.1 Macrophages
YKL-40 mRNA is not expressed in normal human monocytes, but strongly expressed in the late stage of macrophage differentiation (Krause et al. 1996; Rehli et al. 1997;
Renkema et al. 1998; Rehli et al. 2003). In vivo YKL-40 mRNA and protein expression are found by a subpopulation of macrophages in different tissues with inflammation and extracellular matrix (ECM) remodeling (Kirkpatrick et al. 1997; Boot et al. 1999;
3
Johansen et al; 1999; Johansen et al. 2005; Junker et al. 2005). Another “mammalian chitinase-like protein”, chitotriosidase, is also produced by activated macrophages but not by the same sub-population as YKL-40 (Boot et al. 1999).
1.1.1.1.2 Neutrophil granulocytes
Neutrophil granulocytes share a common progenitor cell with macrophages and neutrophil precursors begin to synthesize YKL-40 at the myelocyte-metalocyte stage (Volck et al. 1998). These granulocytes store YKL-40 in the specific granules, and release them after full activation (Volck et al. 1998; Boussac et al. 2000).
1.1.1.1.3 Chondrocytes
YKL-40 mRNA expression is high in cartilage from rheumatoid arthritis (RA) patients and undetectable in normal cartilage (Hakala et al. 1993). The cartilage explant- or monolayer chondrocyte cultures isolated from RA cartilage (Hakala et al. 1993) and osteoarthritis (OA) cartilage (Johansen et al.
2001) secreted YKL-40 in vitro. Spontaneous YKL-40 release was higher in OA than in normal cartilage explant cultures (Johansen et al. 2001).
1.1.1.1.4 Vascular smooth muscle cells
YKL-40 is synthesized in vitro by vascular smooth muscle cells isolated from
4
explants of swine thoracic aorta during the time of transition from a proliferating monolayer culture to a non-proliferation differentiated multilayer culture (Millis et al. 1985; Millis et al. 1986).
1.1.1.1.5 Bone cells
No YKL-40 was found in monolayer cultures of osteoblasts from adult and fetal bone (Johansen et al. 1992), but the YKL-40 mRNA expression was found in end-stage osteoblasts in osteophytic tissue and in primary osteocytes and osteoblasts at sites of endochondral and intramembranous bone formation (Connor et al. 2000).
1.1.1.1.6 Fibroblast-like synovial cells
Human synovial cell monolayers from RA patients secrete YKL-40 in vitro (Nyirkos et al. 1990). YKL-40 mRNA expression is found in inflamed synovial membrane from RA patients but not in non-inflamed synovial membrane (Hakala et al. 1993).
1.1.1.1.7 Liver cells
Strong YKL-40 mRNA expression was found in human liver (Hakala et al.
1993). The liver tissue may originated from a fibrotic liver since the result could not be reproduced by Hu et al. (1996). Immunohistochemical studies of
5
liver biopsies have shown YKL-40 protein expression in areas of the liver with fibrosis and no expression in hepatocytes (Johansen et al. 1997; Johansen et al. 2001)
1.1.1.1.8 Mammary epithelial cells
YKL-40 was isolated from bovine mammary secretions by heparin-agarose affinity chromatography and gel filtration (Rejman et al. 1988)
1.1.1.2 Expression in cancer cells
YKL-40 was discovered to be secreted in vitro in large amount by a human osteosarcoma cell line MG63 (Johansen et al. 1992). Studies based on different cancer diseases also indicated that YKL-40 was expressed in many different types of human solid cancer cells, including osteosarcoma (Johansen et al. 1992), breast cancer (Jensen et al. 2003; Shao et al. 2009), small cell lung cancer (Junker et al. 2005), cervical adenocarcinoma (Mitsuhashi et al. 2009), colon cancer (Shao et al. 2009), colorectal cancer (Kawada et al. 2011), glioblastoma (Horbinski et al. 2010; Francescone et al.
2011). The YKL-40 gene was identified by microarray gene analysis to be one of the most highly over-expressed genes in high-grade malignant gliomas (Lal et al. 1999; Marker et al. 2002; Tanwar et al. 2002). Nevertheless, not all
6
the cancer cells express YKL-40. Those cancer cells that express YKL-40 may have a different phenotype compared to cancer cells without YKL-40 expression, and the protein may reflect differences in the biology of various cancer cells.
1.1.1.3 Regulation of YKL-40
Some studies indicated that changes in extracellular matrix (ECM) are related to changes in YKL-40 synthesis, since the changes in the ECM environment of chondrocytes seem to affect YKL-40 production by these cell (Johansen 2006). Some cytokines were found to affect YKL-40 expression.
Interleukin-1-beta (IL-1ß) inhibited YKL-40 mRNA expression and secretion by human monolayer chondrocyte and cartilage explant cultures (Johansen et al. 2001). IL-6 can stimulate YKL-40 secretion in freshly isolated chondrocytes (Xing et al. 1998)
1.1.2 Biological properties of YKL-40
Thought the real biological function and mechanism of YKL-40 still remain unknown, there’re some properties have been proposed, including the growth
stimulating property, and some bind properties that the YKL-40 is proved to bind heparin, chitin and some type of collagen.
7
1.1.2.1 Growth properties
YKL-40 protein purified from the MG63 osteosarcoma cell line has the growth factor activity that increases growth rates of three fibroblastic cell lines derived from human osteoarthritic synovium, fetal lung and adult skin.
This property was related to the mitogen-activated protein kinase (MAPK) and protein kinase B (AKT) signaling pathways, suggests that YKL-40 initiates a signaling cascade in connective-tissue cells which leads to increased cell proliferation. YKL-40 stimulated proliferation of chondrocytes and synoviocytes, and total proteoglycan synthesis by guinea pig and rabbit chondrocytes in a dose-dependent manner (De Ceuninck et al. 2001). The observation of the activation of the MAP kinase and PI-3K signaling pathways by YKL-40 was anaylysed in comparison with IGF-1. A dose-dependent phosphorylation of ERK1 and ERK2 was observed for synovial cells, human fetal lung fibroblasts, and skin fibroblasts. Signaling through the PI-3K pathway has been associated with cell survival, and activation of AKT pivotal in this aspect. YKL-40 induced a dose-dependent AKT phosphorylation in human synovial cells, with an effective range similar to that of IGF-1 (Recklies et al. 2002). YKL-40 induces VEGF expression
8
and angiogenesis through signaling activation of Integrin ανβ5, Syndecan-1, FAK, and ERK
1.1.2.2 Heparin binding properties
The deduced sequence of YKL-40 reveals a protein of 383 amino acids containing a single consensus sequence for N-linked glycosylation and heparin binding (Shackelton et al. 1995). In vitro studies reveal that it is glycosylated and has affinity for heparin-Sepharose (Millis et al. 1985).
YKL-40 has a single putative heparin binding site (location 144-147) (Malinda et al. 1999; Nishikawa et al. 2003). With this binding site, YKL-40 may interact with heparin-like molecules in the extra-cellular matrix or on the cell surfaces.
1.1.2.3 Chitin binding properties
The existence of a human chitinolytic enzyme was noted and named chitotriosidase on the basis of its capacity to hydrolyse the artificial substrate 4-methylumbelliferyl-β-chitotrioside (Hollak et al. 1994). Chitotriosidase is enzymatically active towards natural chitin. Activated human macrophages, derived from peripheral blood monocytes, can also secrete large quantities of chitotriosidase (Renkema et al. 1998). YKL-40 has high amino acid sequence
9
homology to bacterial chitinases (Hakala et al. 1993; Johansen et al. 1993) and strong binding affinity for chitin (Hakala et al. 1993; Renkema et al. 1998;
Houston et al. 2003). YKL-40 is also homologous (53% sequence identity) to the human macrophage chitinase and other family 18 chitinases, although it lacks the chitinase activity (Hakala et al. 1993; Renkema et al. 1998), as expected from sequence conservation it is able to bind chitin and therefore it can be defined as a chitinase-like lectin (Houston et al. 2003).
1.1.2.4 Collagen binding properties
Affinity chromatography experiments with purified YKL-40 demonstrated specific binding of all three forms to collagen types I, II, and III, thus identifying this important extracellular matrix (ECM) macromolecule as a novel, non-carbohydrate ligand (Bigg et al. 2006).
1.1.3 YKL-40 in infectious, chronic inflammation and joint diseases
YKL-40 is expressed and secreted by macrophages, neutrophils, fibroblast-like synovial cells, chondrocytes, vascular smooth muscle cells and hepatic stellate cells. It has been hypothesized that YKL-40 has a role in acute and chronic inflammation and in pathological conditions leading to tissue fibrosis.YKL-40 can be regarded as an acute phase protein, since its serum concentration increased
10
by more than 25% following an inflammatory stimulus (Kushner 1982).Activated macrophages and neutrophils are probably the major source of circulating YKL- 40 in infectious diseases. However, the function of YKL-40 in the inflammatory foci of a bacterial infection is unknown.YKL-40 is an autoantigen in RA and may play a pathogenic role in the inflammatory process and joint destruction of RA patients. Increased YKL-40 synthesis in the joints of RA patients may lead to an increased YKL-40-derived peptide presentation, which could amplify the local autoimmune response (Johansen 2006).
1.1.4 YKL-40 in cancer diseases
The biological function of YKL-40 in cancer diseases is still unknown, and it is hypothesized that YKL-40 is a growth factor of cancer cells or protects them from undergoing apoptosis (Johansen 2006). Neoplasms are biologically heterogeneous and contain subpopulations of cancer cells with different angiogenic, invasive and metastatic properties. Metastases can have a clonal origin, and different metastases can originate from the proliferation of different single cells. The process of metastases is selective for cells that succeed in promoting angiogenesis, invasion, embolization, survival in the circulation, arrest in distant capillary beds and extravasation into and multiplication within the organ
11
parenchyma (Johansen 2006). There’re some properties of YKL-40 have been proposed (section 1.1.2), all of which may have played an important role in helping cancer cell proliferation, metastasis and survive. Therefore YKL-40 is considered a protein which has a growth factor-like ability, and involved in proliferation of cancer cells, the surrounding tissue remodeling processes and angiogenesis. Furthermore, the serum concentration of YKL-40 has been suggested as a potential biomarker which is associated with a worse prognosis in cancer diseases (Jensen et al. 2003; Mitsuhashi et al. 2009; Shao et al. 2009; Allin et al. 2012) and the protein has been found to be a contributory factor in tumor-associated angiogenesis (Shao et al. 2009).
1.1.5 YKL-40 may be an autoantigen
Chitin is the second most abundant natural polysaccharide (after cellulose), produced by fungi, crustaceans, helminths, insects, and many others, to more than one billion tons per year. It is not produced by animals; however, degradation of this stable and insoluble polymer is important for immune defense against chitin-containing organisms. Active chitinases are induced in diseases involving chitin-containing organisms but have also been implicated in several disorders in which a role for such pathogens has not been demonstrated
12
(Qureshi et al. 2011). There is increasing evidence that several chitinase-like proteins, notably YKL-40, have an association with chronic inflammation and tissue remodeling, with overexpression observed in autoimmune diseases, allergy, wound healing, infection, and several cancers and indeed can be prognostic indicators for some cancers (Coffman 2008).
1.1.6 YKL-40 in cell migration
YKL-40 has the effect with a dose-dependent manner in stimulating human umbilical vein endothelial cells (HUVEC) migration, and reached a maximal level similar to that produced in response to the control chemoattractant basic fibroblast growth factor (bFGF). The expression of YKL-40 during a period of smooth muscle cell migration into nodules suggests that it might have a role in modulating migration of vascular cells (Malinda et al. 1999). YKL-40 also stimulates vascular smooth muscle cell (VSMC) migration to a level approximately 10 to 15 times that of the BSA-containing negative control and 10 times that of the known chemoattractant platelet-derived growth factor (PDGF).
The migration effect of YKL-40 on nonvascular cell migration was less than that observed with BSA and considerably less than the PDGF effect on these cells, suggested that the migration response to YKL-40 may be specific for vascular
13
cells (Nishikawa et al. 2003).
1.1.7 Possible cell signal pathway of YKL-40
The use of selective inhibitors of the mitogen-activated protein kinase (MAPK) and the protein kinase B (AKT) signaling pathways indicates that both are involved in mediating the mitogenic response to YKL-40. Phosphorylation of both extracellular signal-regulated kinases 1/2 and AKT occurred in a dose- and time-dependent fashion upon addition of YKL-40 (Recklies et al. 2002).
YKL-40 also induced coordination of membrane receptor syndecan-1 (Syn-1) and integrin ανβ5, and triggered a signaling cascade through FAK397 to MAPK (ERK1/2), leading to elevated vascular endothelial growth factor (VEGF) and enhanced angiogenesis (Shao et al. 2009; Francescone et al. 2011).
1.2 Canine mammary gland tumor
Canine mammary gland tumors (cMGT) are the most common neoplasm in female dogs, comprising 52% of all neoplasms (Benjamin et al. 1999). cMGT can be multiple and may vary in histology within or among different tumor sites in an individual. The tumors often have a complex morphology consisting of epithelial, mesenchymal or mixed-type cells. All type of the tumors can exist both as benign or malignant, with about 50% of the tumors being benign, with complex adenoma and
14
benign mixed tumor as the dominant types, whereas pure benign mesenchymal tumors are rare (Hellmén et al. 2000). The first histologic classification system for canine mammary gland tumor have been published in 1974, and later a modification
in 1999 (Goldschmidt et al. 2011). The World Health Organization published the first
“International Histological Classification of Tumours of Domestic Animal” in 1974,
which included tumors and dysplasia of the mammary gland. Spontaneous tumors in dogs are appropriate and valid model tumor systems available for testing cancer therapeutic agents or studying cancer biology. Dogs develop spontaneous tumors with histopathologic and biologic behavior similar to tumors that occur in humans (MacEwen 1990).
1.3 Conclusion
The YKL-40 certainly plays a role in the disease development, whether inflammatory, chronic, or neoplastic. It’s interesting that this protein has the ability similar to the growth factor, and has the ability to stimulate tumor angiogenesis. The actual mechanism of its biological function and the real cell signal pathway it’s involved in appears to be more attractive and important. Dr. Johansen said that the study of YKL-40 has just started and several biological questions regarding this protein remain to be answered. Since there’s little research related to canine diseases so far,
15
we’re eagerly to find out more information of this protein in the field of veterinary medicine, and hope to contribute to the cancer research of animals. In the study to the cancer research, spontaneous tumor in dog such as mammary gland tumor will be an appropriate model for the study of cancer biology. Therefore, we chose the cMGT cell lines CMT-1 and MPG to test the biological function of YKL-40 and also the cell signaling pathway that YKL-40 is involved in.
16
Chapter 2 Introduction
YKL-40 is a glycoprotein which was identified to be secreted in human osteosarcoma cell line MG63, so far most of the YKL-40 studies were based on human researches (Nishikawa et al. 2003; Johansen et al. 2006), and rare data related to canine is presented. The YKL-40 protein is a secretory glycoprotein with heparin-binding activity (Millis et al. 1985; Shackelton et al. 1995; Nishikawa et al. 2003; Bigg et al. 2006). Both well-differentiated macrophages and activated neutrophils are found to express YKL-40 (Krause et al. 1996; Rehli et al. 1997; Renkema et al. 1998). Some studies also indicated that human articular chondrocytes express YKL-40 when stimulated with interleukin-6 (IL-6) and IL-17, while transforming growth factor-beta (TGF-ß) reduces YKL-40 expression (Johansen et al. 2001).
However, the biological function of YKL-40 in cancer diseases is still unknown. It has been hypothesized that YKL-40 is a growth factor of cancer cells or protects them from undergoing apoptosis (Johansen 2006). Some properties of YKL-40 have been proposed, including the ability to stimulate cell growth, heparin binding ability, chitin binding ability, and collagen binding ability. It is believed that with these properties YKL-40 is related to the proliferation of connective tissues and activation of vascular endothelial cells, therefore promotes tumor angiogenesis and malignancy (Johansen
17
2006; Shao et al. 2009; Francescone et al. 2011). This made us wonder the biological function of the homologous canine YKL-40 protein, whether if it has the same properties in canine neoplasms.
Spontaneous tumors in dogs are appropriate and valid model tumor systems available for testing cancer therapeutic agents or studying cancer biology. Dogs develop spontaneous tumors with histopathologic and biologic behavior similar to tumors that occur in humans (MacEwen 1990). Therefore we chose the canine mammary gland tumor cell lines, CMT-1 and MPG, to investigate the biological function of our recombinant canine YKL-40 protein. Based on the properties that have been proposed, we will test the ability of YKL-40 in stimulation of cell migration, proliferation and invasion, and evaluate the effects of the recombinant YKL-40 protein. Furthermore, as an advanced research in the growth property of YKL-40, we will investigate the cell signal pathway that YKL-40 may be involved with its growth factor similarity.
There are also studies indicated that YKL-40 may be an autoantigen in some autoimmune diseases (Patil et al. 2001; Qureshi et al. 2011). This draws our attention and makes us interested in the detection of YKL-40 by its autoantibody. We wonder that whether the recombinant YKL-40 will be recognized by the autoantigen in the dog. Out of the available antibodies, especially the monoclonal antibody, that against canine
18
YKL-40, it seems the potential autoantibody in the dogs could be our best chance.
Therefore we tried applying the dog serum as a primary antibody to probe our recombinant canine YKL-40 in the Western blot, and the preliminary result showed the recombinant canine YKL-40 indeed recognized by the antibody in the dog serum. This result suggested the possibility of this theory of autoantigenicity of YKL-40.
19
Chapter 3 Material and Method
3.1 Experimental animals
All experimental beagle dogs had be de-wormed and vaccinated with distemper, adenovirus, parainfluenza virus, leptospirosis, parvovirus, coronavirus, infectious hepatitis virus, and hemorrhagic icterus virus in the eight in one shot vaccine. All experiments were performed according to the National Taiwan University Animal Experimental Ethics Committee guidelines.
3.2 Cell lines
In this study, we used two different canine mammary gland tumor cell lines, CMT-1 and MPG, which were gifts from Dr. Lin’s laboratory (Chang et al. 2010), to perform biological functional assays. Both CMT-1 and MPG were cultured in the complete Dulbecco’s Modified Eagle Medium (DMEM, Caisson, USA) supplemented with 10% fetal bovine serum (FBS, Caisson, USA) and 1% antibiotics (Casisson, USA).
These two cell lines don’t spontaneously express YKL-40 mRNA and protein. In expression of the YKL-40 protein, we applied the BALB/3T3 cells in the transfection. The BALB/3T3 (ATCC®CCL-163.2TM) cell line was from the American Type Culture Collection (ATCC). After transfection and selection, we established the stable cell line CL7208 which can stably, continuously, and
20
spontaneously express the recombinant canine YKL-40 protein.
3.3 Anitbodies
In this study, we used the mouse source anti-6x-Histidine monoclonal antibody (mAb) (Novus Biologicals, CO, USA) to probe the recombinant canine YKL-40 protein in Western blot analysis, dilution 1:2000. In the cell signal pathway analysis, we used the following antibodies to probe the cell signal pathway proteins in the Western blot: rabbit source anti-phospho-Akt mAb (#4060, Cell Signaling, MA, USA), dilution 1:2000; rabbit source anti-Akt antibody (#9272, Cell Signaling, MA, USA), dilution 1:1000; rabbit source anti-phospho-MAPK (Erk1/2) mAb (#4370, Cell Signaling, MA, USA), dilution 1:2000; mouse source anti-MAPK (ERK1/2) mAb (#4696, Cell Signaling, MA, USA), dilution 1:2000; rabbit source anti-phospho-Stat3 mAb (#9145, Cell Signaling, MA, USA), dilution 1:2000;
mouse source anti-Stat3 mAb (#9139, Cell Signaling, MA, USA), dilution 1:1000.
3.4 Total RNA extraction
In the YKL-40 cloning portion, we used TRIzolTM (Invitrogen) to extract total RNA from canine macrophages, and followed the procedure suggested by the manufacturer. While in the YKL-40 mRNA expression examination for the canine mammary gland tumor cells, we used RNAzol® RT (Molecular Research Center,
21
Inc., OH, USA) to extract total RNA from the cMGT cells.
3.5 RT-PCR
The RNA was subsequently reverse-transcribed into cDNA with a Super-Script II reverse transcriptase kit (Invitrogen) according to the manufacturer’s suggestion.
Because that so far only partial canine YKL-40 sequence was predicted (Accession number: XM_547343, GenBank database: http://www.ncbi.nlm.nih.gov/genbank/), we obtained the full length of the canine YKL-40 gene for the plasmid insertion. The total RNA will first be quantified and qualified by using a spectrometer, then 1 μg of RNA was proceeded with a de-genomic-DNA procedure: using RQ1 RNase-free DNase (Proméga, Madison, WI, USA), and RQ1 DNase 10X Reaction Buffer (Proméga, Madison, WI, USA). After incubation at 37℃ for 30 minutes, the RQ1 Stop Solution (Proméga, Madison, WI, USA) was added to stop the DNase reaction at 65 ℃ for 10 minutes. The RNA without genomic-DNA was then reverse-transcribed into cDNA by using the HiScript I Reverse Transcriptase (MmLV) (Bionovas, Toronto, Ontario, Canada), 5X Strand Buffer (Bionovas, Toronto, Ontario, Canada), 100 mM DTT (Bionovas, Toronto, Ontario, Canada), Fermentas RiboLock RNase Inhibitor (Thermo Scientific, Burlington, Ontario, Canada), 1μM oligo dT primer and random primer. The PCR protocol as following:
22
30℃,10 min; 42℃, 60 min; 70℃ 15 min.
3.6 Preparation and propagation of YKL-40/pcDNA 3.1 plasmid
The canine YKL-40 gene without stop codon (InC) was obtained from RT-PCR using primer pair F and InR, and the product InC was subsequently cloned into pcDNA3.1/V5-His-TOPO plasmid using the pcDNATM3.1/V5-His-TOPO®TA Expression Kit (Invitrogen, CA, USA), and followed the procedure suggested by the manufacturer. Once the canine YKL-40 gene is cloned into the pcDNA 3.1/V5-His-TOPO plasmid, it is transformed into E.coli for propagation. The multiplied plasmid is then extract with the Plasmid Midi Kit (Qiagen), and follow the procedure suggested by the manufacturer.
3.7 Transfection of canine YKL-40 gene into BALB/3T3 cells
This extract canine YKL-40/pcDNA 3.1 plasmid was then transfected into BALB/3T3 cells by using Lipofectamine 2000 (Invitrogen, CA, USA); the plasmid pcDNA 3.1 was used as a mock control, while the eGFP plasmid was used as a positive control and representative of the transfection rate. The transfected cells were harvested after 24, 36 and 72 hours. 1000 cells from each time point were transferred to 96-well plate and G418 (Amresco, OH, USA) at the concentration of 800 ng/μL was added into each well to select the successfully transfected cell. Since
23
the pcDNA3.1 plasmid contains an anti-neomycin gene, successfully transfected cell will survive in the presence of G418. By using this method, we obtained several cell lines which were developed from single cell. After 2 weeks of treatment of G418, the concentration was decreased to 400 ng/μL, for the maintenance growth of the cell line.
3.8 Expression of canine YKL-40 protein in BALB/3T3 cells
The transfected cell lines were named by the harvesting time, There were totally 15 cell lines acquired, after confirmation and quantification, we chose the cell line CL7208 to produce our recombinant YKL-40 protein. The protein was overexpressed by this cell line, and secreted into the medium. The 0% FBS medium was used to collect the secreted protein.
3.9 Concentration of the recombinant canine YKL-40 protein
The recombinant canine YKL-40 is a secreted glycoprotein, which will be secreted into the culture medium. We collect the FBS-free DMEM cultured with CL7208 cells after 24, 48 and 72 hours of incubation. Once the cultured medium is collected, the proteinase inhibitor (Bionovas, Toronto, Ontario, Canada) is added follow the manufacturer’s suggestion. The medium will be concentrated with the centrifugal concentrator tubes (Vivaspin 20, pore size: 10 kDa MWCO, GE Healthcare, Little
24
Chalfont Buckinghamshire, UK), and centrifuged with 2330 xg at 4℃ (Kubota 5910 centrifuge, Japan) first. The centrifuge time depends on the volume of the medium needs to be concentrated, 30 minutes for one centrifugation, mostly 3 to 5 hours at total. The protein was concentrated about 20-40 folds, and the medium will be replaced by phosphate buffered saline (PBS). The concentrated protein will be quantified after concentration is completed. After protein quantification, we found that the most appropriate time for collection is 48 to 72 hours, because that the protein will reach a certain concentration and the condition of cells still remains well.
3.10 Purification of the recombinant canine YKL-40 protein
The solution was then purified by His-column (IMAC FF 5 mL, GE Healthcare Bio-Sciences, NJ, USA), and Heparin-column (HiTrap Heparin HP 1 mL, GE Healthcare Bio-Sciences, NJ, USA). The different proportion of the purification, including supernatant, flow-through, wash, and elute were collected for protein detection.
3.11 Quantification of the recombinant canine YKL-40 protein
After purification, the eluted and concentrated recombinant canine YkL-40 protein will be quantified with the Coomassie Plus – The Better Bradford AssayTM Kit
25
(Thermo Scientific, IL, USA), and followed the procedure suggested by the manufacturer. The bovine serum albumin (BSA) standard (2 mg/mL, Thermo Scientific Pierce, IL, USA) was used to generate a standard curve. Each concentration of protein is triplicate.
3.12 Identification of the purified recombinant canine YKL-40 protein
The purified and concentrated recombinant canine YKL-40 protein will be separated by 10% SDS-PAGE and western blotted. The membrane was probed with mouse source anti-His-tag monoclonal antibody.
3.13 Western blot analysis
The protein samples were boiled with 5X protein loading dye for 10 minutes. The samples are separated by 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) with 80 volt for 120 to 150 minutes, and then Western blotted to the polyvinylidene difluoride (PVDF) membranes, the condition is 300 mA for 1 hour. 20 mL of skim milk is used to block the membrane on a rotator shaker at 4℃ for 2 hours. Then the primary antibody will be diluted with the skim milk at a proper dilution ratio and began to probe with the incubation on a rotator shaker at 4℃ overnight. Before adding the secondary antibody, the membrane will be washed 3 times with Tris buffered saline and Tween 20 (TBST), the Tween concentration is
26
1% and each washing time is 10 minutes. Then the secondary antibody will be diluted with the skim milk at a proper dilution ratio and began incubation on a rotator shaker at room temperature for 6 hours. The membrane will be washed with TBST three times, each time for 10 minutes. After washing, the membrane will be put on a plastic board and add the enhanced chemiluminescence (ECL, RapidStep™
ECL Reagent) (Millipore, MA, USA). The membrane will be photographed by the image system Geliance 600 (Perkin Elmer, MA, USA).
3.14 Proliferation assay
The cMGT cell lines CMT-1 and MPG were seeded into 96-well plates at 3000 to 5000 cells per well density, and allowed to adhere for 24 hours in DMEM containing 10% FBS. The medium was then discarded and washed with FBS-free DMEM. The cells were then divided into six groups, treated with DMEM plus 10, 100, and 1000 ng/mL of YKL-40. The medium DMEM without FBS cultured with BALB/3T3 and CL7208 cells were also used to this test. DMEM without FBS was used as the negative control. Each condition was assayed in triplicate wells.
3.15 Migration assay
The cMGT cell lines CMT-1 and MPG were seeded into 6-well plates at 2x105 cells per well density. After 24 hours of incubation with 10% FBS DMEM, each well was
27
grooved with 1 mL pipette tip. The cultured medium was discarded and the wells were then washed by DMEM without FBS. The purified YKL-40 protein in different concentration was added to each well. The result was observed at 18, 24, 36, and 48 hours with microscope. The results were then calculated by computer software and also analyzed.
3.16 Invasion assay
Invasion assays were conducted in cell culture inserts (BD Falcon, NJ, USA) with a transparent PET track-etched membrane (pore size 8.0 μm). 100 μL of Matrigel (BD, NJ, USA) was added into the insert at 4℃ and incubate at 37℃ for gelling, then 4x105 cells in 200 μL of DMEM without FBS were added onto the gel. This prepared insert was then placed into the 24-well plate, each well contains different medium to be tested.
3.17 Autoantigen analysis
The recombinant canine YKL-40 protein was separated by 10% SDS-PAGE and western blotted. The membrane was then probed with the dog serum from the healthy dog without any known disease of our experimental Beagles. The sera were diluted 500 times for the use as primary antibody in the Western blot analysis. Then the anti-canine antibody conjugated with horseradish peroxidase (HRP) was used a
28
secondary antibody to probe the bound serum antibodies. The membrane was observed with ECL and photographed by the image system as well as the Western blot analysis.
3.18 Cell signal pathway analysis
This analysis applied the canine mammary gland tumor cell lines CMT-1 and MPG to the test. After trypsinized from the 175 cm2 flask, the cells are distributed into 5 eppendorfs per 5x106 for each tube. Then centrifuge with 1500 xg for 10 minutes (Prism Microcentrifuge) (Labnet International, Inc., NJ, USA), and discard the medium. 1 mL of DMEM without FBS will added into each eppendorf and incubate at 37℃ for 1 hour serum starvation. The cells will be treated with 10% FBS DMEM, 10, 100, 1000 ng/mL of recombinant YKL-40 protein, respectively, the DMEM without FBS will be the negative control. After 1 hour of treatment, the cells will be centrifuged with 1500 xg for 10 minutes, and discard the medium. Then 100 μm of RIPA lysis buffer (Millipore, MA, USA), which contains 1% of proteinase inhibitor (Bionovas, Toronto, Ontario, Canada), 2 % of phosphatase inhibitor (Sigma-Aldrich, MO, USA), and 1% of Na3VO4, will be added into each eppendorf. After 30 minutes of incubation on ice, with vortex thoroughly every 5 minutes, and centrifuge with 3000 xg for 10 minutes. The supernatant will be transfer to a new eppendorf and the
29
total protein will be quantified. 100 μg of protein will be applied to the Western blot, the membrane will be probed with anti-phospho-AKT, anti-phospho-MAPK (p-ERK1/2) and anti-phospho-STAT3 antibodies. Then the membrane will be stripped and probed with anti-AkT, anti-MAPK, anti-STAT3 and anti-actin antibodies again.
30
Chapter 4 Results
4.1 YKL-40 protein expression by stable cell line establishment
The cloned YKL-40/pcDNA 3.1 (Fig 1) was sequenced and multiplied by E.coli. The plasmid was then extract with the plasmid extraction kit, and then transfected into BALB/3T3 cells. Because we use the eGFP plasmid was as positive control and representative of transfection rate, it can be detected by cytofluorescence assay (Fig 3). Different groups of 24 hours, 48 hours, and 72 hours was then selected by treatment with G418 at the concentration of 800 ng/μL. There were totally 15 cell lines survived the selection, but not everyone expression the recombinant YKL-40 protein correctly and successfully. And we discovered that there were two types of the transfected cells (Fig 4), one will scatter evenly and the other will overlapped.
After confirmation and quantification, CL2436, CL4805 and CL7208 were the cell lines that can correctly express the YKL-40 protein. However, CL7208 expresses the YKL-40 protein more rapidly and massively. Therefore it is chosen to be our primary protein expression cell line.
4.2 YKL-40 mRNA expression examination
In order to confirm that the cell lines we used in this study don’t express YKL-40 mRNA or protein spontaneously. We examined all the cell lines for the expression of
31
YKL-40 mRNA. The mRNA was examined by extract the total RNA and transcribed into cDNA, and performed polymerase chain reaction (PCR) to check the specific sequence product of YKL-40 mRNA. We tested BALB/3T3, CL2436, CL7208, CMT-1, MPG, and the other canine mammary gland tumor cell line CF41, the result showed no mRNA expressed in these cell lines (Fig 2A). For further investigation, we examined the osteosarcoma cell line D17, lymphoma cell lines CLBL-1, UL-1, CLC, and melanoma cell lines CM-01-01, M1, M2, M3, M4, and M5. The result showed that UL-1, CLC, and M1 don’t express YKL-40 mRNA, while CM-01-01, M2 to M5 expressed YKL-40 mRNA spontaneously and CLBL-1 expressed sparsely (Fig 2B).
4.3 Purification of the recombinant canine YKL-40
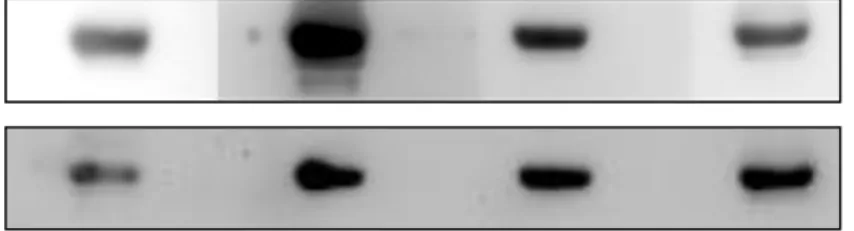
The recombinant canine YKL-40 protein was secreted into the cultured medium by CL7208 cells. The cultured medium were collected and concentrated by centrifugal concentrator tubes, the concentration was about 20 folds. We first used the His-column to purify the concentrated recombinant canine YKL-40 protein, the different fluid portion of purification was collected and separated by 10% SDS-PAGE and western blotted, and membrane was probed with anti-His-tag antibody (Fig. 5A).
The result showed that the His-column didn’t bind the recombinant canine YKL-40
32
protein successfully, most of the protein was presented in the flow-through fluid and the rest of the protein was washed away after washing buffer applied. No protein was detected in the elute fluid, even after concentration with centrifugal concentrator.
Because YKL-40 has the ability to bind heparin, we used the heparin-column to purify this protein instead (Fig. 5B). No protein was detected in the flow-through fluid and the washed fluid. Most of the protein was successfully bound by the heparin-column and eluted. The protein can still be detected after concentration by using the centrifugal concentrator.
4.4 Proliferation assay
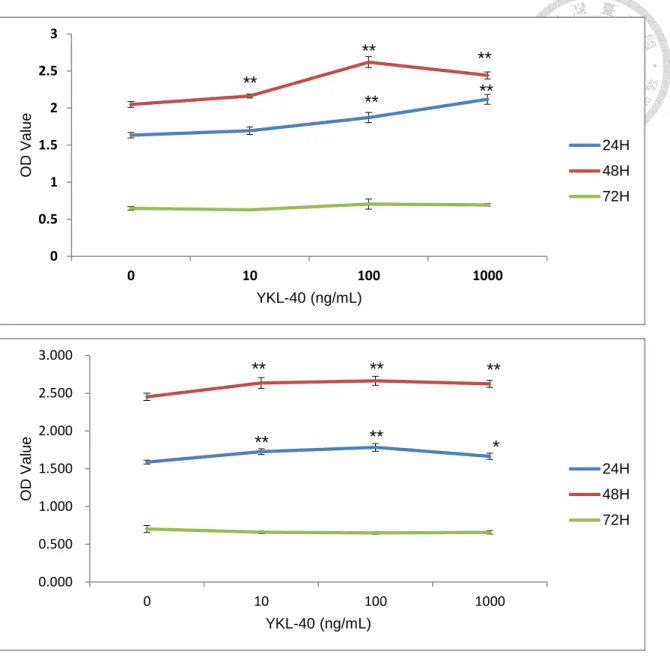
To test if the recombinant canine YKL-40 has the ability to stimulate cell to proliferate, we evaluate the effect of the recombinant canine YKL-40 in CMT-1 (Fig 6A) and MPG cells (Fig 6B). After several times of repeat, the recombinant canine YKL-40 didn’t show any significant effect in the stimulation of cell proliferation.
There seems to be a trend in the effect of the recombinant canine YKL-40, and the cell treated with the recombinant canine YKL-40 do proliferate better than the ones which were not treated. And the result seems to have a dose-dependent pattern in the treatment.
4.5 Migration assay
33
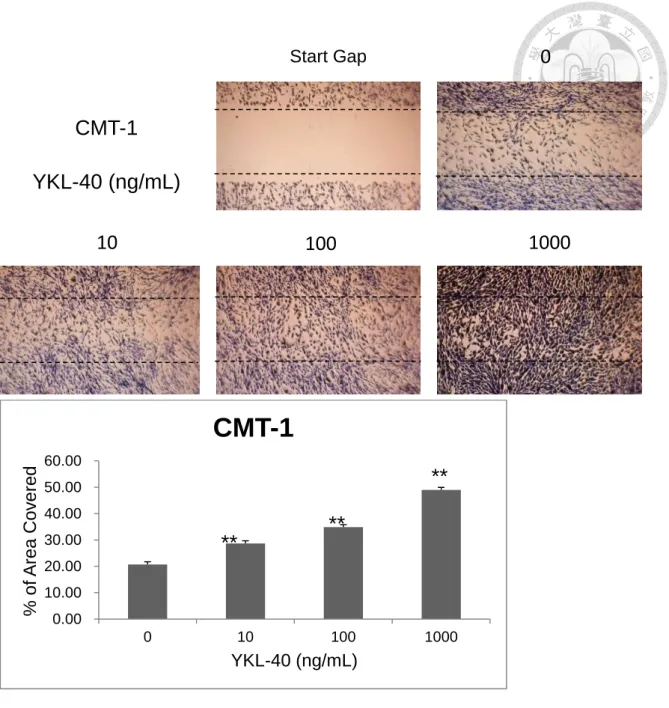
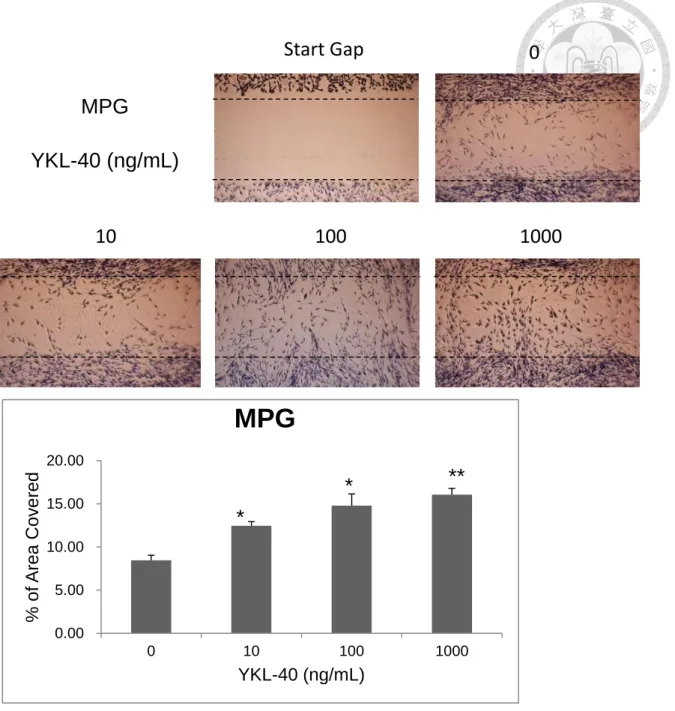
To test if the recombinant canine YKL-40 has the ability in stimulation of cell migration, we applied the CMT-1 and MPG cells to the test. After different time of treatment, the difference became apparent after 24 hours. In the group treated with the recombinant canine YKL-40, the cells migrate and recover the gap faster than the control group in a dose-dependent pattern. This effect was more obvious in CMT-1 cells (Fig. 7A) than in MPG cells (Fig 8A), as the result being quantified by computer software (Fig. 7B, 8B). *P<0.05 and **P<0.01 compared with corresponding control cells treated with serum-free DMEM.
4.6 Invasion assay
To examine whether the YKL-40 protein has the ability to stimulate cancer cell invade into the extracellular matrix (ECM), we applied the matrigel to mimic the property of ECM. By observation of the opposite side of the membrane of the insert, we found that the YKL-40 stimulates the invasiveness of the cancer cells in a dose-dependent manner (Fig 9).
4.7 Cell signal pathway analysis
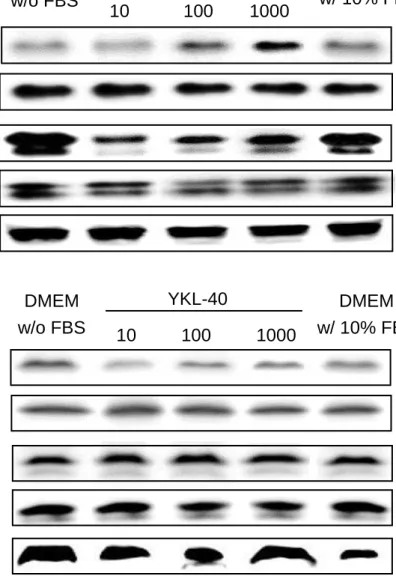
In the result of cell signal pathway investigation, YKL-40 was found to induce the PI3K/AKT pathway both in CMT-1 and MPG cells (Fig 11, 12). The activated form of AKT was increased by YKL-40 in as low as 10 ng/mL, the result in CMT-1 and
34
MPG cells both showed a dose-dependent manner. In CMT-1 cells, the activated form of MAPK (ERK1/2) were also induced by YKL-40 in as low as 10 ng/mL and in a dose-dependent manner. However, this result wasn’t showed in MPG cells.
35
Chapter 5 Discussion
In this study we want to investigate the biological function of canine YKL-40 protein, for this reason, we need a source that can stably provide this protein for us to apply in different assays. Most of the studies rely on the human or mouse cartilage chondrocyte (Hakala et al. 1993; De Ceuninck et al. 2001; Faibish et al. 2011), vascular smooth muscle cells (Nishikawa et al. 2003), to express the YKL-40 protein. So far we don’t have any canine cell line that can spontaneously express canine YKL-40, this means that we must produce the canine YKL-40 with a bioengineering method. In addition, there’s no available antibody can recognize the canine YKL-40 protein for us to use, a 6x-histidine tag attached to the recombinant protein will be an appropriate method for detection. Because that the YKL-40 protein is a secreted glycoprotein, for the consideration of the proper glycosylation, we choose the eukaryotic system instead of prokaryotic system to express this recombinant protein.
There’s few study information about using eukaryotic cells to generate recombinant YKL-40, only Scully et al. applied 293T retroviral packaging cells to express recombinant YKL-40 (Scully et al. 2011). Now we want to use the BALB/3T3 mouse embryo cells to express the recombinant canine YKL-40 protein, there’s some uncertainty.
36
One of the advantages of the secreted glycoprotein is that the protein will be secreted out of the cell. We can easily collect the medium which cultured with the CL7208 cells, and no need to lyse the cell. Nevertheless, the phenol red in the DMEM makes the quantification of the protein inaccurate. The protein quantification kit that we used is functioning based on the reaction between the protein and the coomassie G-250 dye, and the detection of the absorbance at wavelength of 595nm. In concern with the effect that the color of the phenol red, we replaced the DMEM with PBS in the end stage of using centrifugal concentrator. Once the solution become transparent, the protein can be quantified correctly.
The pcDNA 3.1/YKL-40 plasmid contains a 6x histidine tag, which should provide a binding site for the his-column. However, the his-column we used somehow failed to purify the recombinant canine YKL-40 protein. One of the most possible reasons should be the folding conformation of the protein, which may be false folded in the expression procedure of the BALB/3T3 cells. Since the 6x histidine tag is aligned in the tail of the amino acid sequence, it may be enwrapped inside the protein and failed to bind the his-column. Fortunately, the YKL-40 protein has the ability to bind heparin. By using this advantage, we’re able to purify the recombinant canine YKL-40 protein with the heparin-column. Though the recycle rate of the heparin-column is considerably high,
37
there’s still elute buffer used in the purification procedure, and thus added the volume of the protein solution. This dramatically decreased the concentration of the purified recombinant canine YKL-40 protein. In the meantime, the protein will slowly degrade as time passes, even the proteinase inhibitor was added and the procedure is in the low temperature condition. Therefore we will again concentrate the purified protein to make sure the concentration of the protein can be in the acceptable range, mostly between 10 to 100 ng/mL.
In the confirmation of the recombinant canine YKL-40 protein, we applied the Western blot analysis to detect the his-tag on the protein. The his-tag of the protein can be probed by the anti-6x-histidine monoclonal antibody correctly, and this can also be used as a double confirmation in the autoantigen analysis. Since there are no available antibody can be used to recognize the canine YKL-40 protein so far, we rely mainly on the anti-his-tag antibody to detect the YKL-40 protein.
In cell proliferation assay, the result showed no significance between different dosages of YKL-40. This was tested several times,
In the investigation of the cell signal pathways that YKL-40 was involved in, we chose to see the PI3K/AKT and MAPK (ERK1/2) pathways. YKL-40 was found to induce the PI3K/AKT pathway in human synovial cells in a dose-dependent manner
38
(Recklies et al. 2002), and also the MAPK pathway in human synovial cells and human microvascular endothelial cells (Recklies et al. 2002; Shao et al. 2009). In this study, we found that YKL-40 indeed induced the PI3K/AKT pathway in both CMT-1 and MPG cells and in a dose-dependent manner (Fig. 11). However, in the MAPK pathway, the result of CMT-1 showed difference in a dose-dependent manner. While in the result of MPG, the activated MAPK (ERK1/2) showed no difference between different YKL-40 concentrations (Fig. 12). Considering the cells applied in this analysis are different, and there’re no mammary gland tumor used in other research, we can only presume that there are several innate differences between the CMT-1 and MPG cells. This can also be verified in the experiment of cell migration assay, in which the CMT-1 showed more obvious response to YKL-40 than the MPG cells.
YKL-40 has many biological functions related to tumorigenesis, such as fibroblast proliferation stimulation and angiogenesis promotion (Recklies et al. 2002; Francescone et al. 2011). However, the specific receptors for YKL-40 have not yet been identified (Hakala et al. 1993; Francescone et al. 2011). For investigation of the role of canine YKL-40 in cancer progression, discovering the specific receptors will be an priority in the future.
In conclusion, the recombinant canine YKL-40 protein that we produced indeed
39
has the properties and abilities in cell growth, including the effects in cell migration and cell invasion. So far we only observed the effects in cell migration and invasion, and mostly obvious on CMT-1 cells. This may be the result of the difference of the canine mammary cell lines. Since most studies investigating the biological function of YKL-40 were based on the effects on human vascular endothelial cells and human synovial cells, there are little information about the YKL-40 in canine study can be referred to. The canine endothelial cells seem to be an appropriate cell in the next step of YKL-40 biological function investigation.
40
Tables
Table 1. Reverse transcription protocol
Solution Volume
5x First strand buffer (Bionovas, Canada) 4 μL HiScript I Reverse transcriptase (Bionovas, Canada) 2 μL
100 mM DTT (Bionovas, Canada) 2 μL
100 mM dNTPs 1 μL
RiboLocks RNase inhibitor (Thermo Scientific, Canada) 1 μL
1 μM Oligo dT primer 1 μL
1 μM Random primer 1 μL
RNA suspension (1 μg total RNA) 8 μL
41
Table 2. YKL-40 primer list
Primer Name Sequence (5'->3')
cYKL-40-F137 CATCTACAGCTTCGCCAACA
cYKL-40-R156 TGTTGGCGAAGCTGTAGATG
cYKL-40-R GGATGGAGCTTTGGTTCTCA
cYKL-40-R968 GTCATCATACCCCACCCACT
oYKL-40-CF741 CTGATGGGCATCCCCACCTT
oYKL-40-CR760 AAGGTGGGGATGCCCATCAG
oYKL-40-CR955 ACCCACTGGTTGCCCTTGGT
CF-YKL-40 TCTGCTGCAGCCAGGATGCT
42
Figures
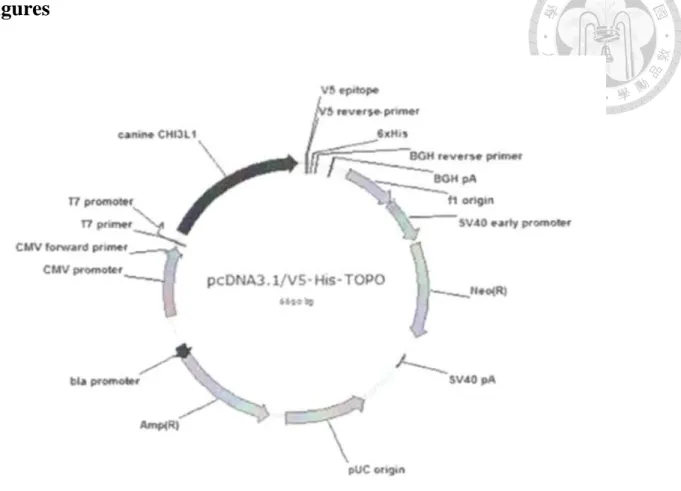
Figure 1. The construct of canine YKL-40/pcDNA 3.1/V5-HIS-TOPO.
The YKL-40 gene was inserted into the vector pcDNA3.1/V5-HIS-TOPO plasmid, which contains 6-histidine tag and anti-neomycin gene.
43
Figure 2. Expression of the YKL-40 mRNA in different tumor cell lines.
YKL-40 mRNA expression in different canine tumor cell lines were analyzed using RT-PCR, including canine mammary gland tumor (MGT) cell lines, osteosarcoma (OSA) cell line, lymphoma cell lines, and melanoma cell lines. No YKL-40 mRNA was expressed in MGT, OSA, lymphomas, and M1 melanoma cell line. NC stands for negative control and PC stands for positive control.
YKL-40
GAPDH
CLBL-1 UL-1 CLC
Lymphomas
CMT-1 MPG CF4 D17
MGTs OSA
YKL-40 GAPDH
CM-01-01 M1 M2 M3 M4 M5 NC PC
Melanomas
44
Figure 3. The transfection of YKL-40 into BALB/3T3 cells
Transfection of canine YKL-40 gene into BALB/3T3 cells. (A)The transfection of eGFP as a positive control, and (B) the pcDNA3.1 as a mock control. (C) The transfection successful rate is more than 50%.
A B
C
45
(A)
(B)
Figure 4. Purification of the recombinant YKL-40 protein with His-column and Heparin-column
(A) The recombinant canine YKL-40 protein purified with His-column and separated by 10%
SDS-PAGE and western blotted, the membrane was probed with anti-His-tag antibody. (B) The recombinant canine YKL-40 protein purified with heparin-column and separated by 10%
SDS-PAGE and western blotted, the membrane was probed with anti-His-tag antibody.
YKL-40 (His-tag)
Supernatant Flow-through Wash Elute
His-column
Elute Concentrated
Supernatant Flow-through Wash Elute
Heparin-column
Elute Concentrated
YKL-40 (His-tag)
A B
46
(A)
(B)
Figure 5. YKL-40 increases the proliferation rate of CMT-1 and MPG cells.
(A) CMT-1 cells were treated with YKL-40 protein, concentration from 10 to 1000 ng/mL, and the treatment time from 24 to 72 hours. The result shows significant difference and is also dose-dependent. (B) MPG cells have similar reaction to YKL-40 protein.
0 0.5 1 1.5 2 2.5 3
0 10 100 1000
OD Value
YKL-40 (ng/mL)
24H 48H 72H
** **
**
** **
0.000 0.500 1.000 1.500 2.000 2.500 3.000
0 10 100 1000
OD Value
YKL-40 (ng/mL)
24H 48H 72H
** ** *
** ** **
47
(A)
(B)
Figure 6. The result of cell migration assay on CMT-1 cells
(A) The result of the cell migration assay on CMT-1 cells, the cells have a better efficiency in migration to recover a gap in a dose-dependent manner. (B) The results in Fig 7A were calculated and analyzed, *P<0.05 and **P<0.01 compared with corresponding control cells treated with serum-free DMEM.
0.00 10.00 20.00 30.00 40.00 50.00 60.00
0 10 100 1000
% of Area Covered
YKL-40 (ng/mL)
CMT-1
**
**
**
CMT-1 YKL-40 (ng/mL)
10 100 1000
Start Gap
0
48
(A)
(B)
Figure 7. The result of cell migration assay on MPG cells
The result of the cell migration assay on MPG cells, the cells appear to have better migration efficacy in higher concentration. (B) The results in Fig 8A were calculated and analyzed by computer software. *P<0.05 and **P<0.01 compared with corresponding control cells treated with serum-free DMEM.
0.00 5.00 10.00 15.00 20.00
0 10 100 1000
% of Area Covered
YKL-40 (ng/mL)
MPG
* **
* MPG
YKL-40 (ng/mL)
10 100 1000
Start Gap 0
49
(A)
(B)
(C)
0 10 20 30 40 50 60
0 10% FBS YKL-40 100
ng/mL
YKL-40 1000 ng/mL
% of Area Covered
**
**
60 80 100 120 140 160 180
0 10% FBS YKL-40 100
ng/mL
YKL-40 1000 ng/mL
% of Control
**
**
0 10% FBS
100 1000
50
Figure 8. YKL-40 stimulates the invasion ability of CMT-1 cells.
(A) The insert with CMT-1 cells treated with YKL-40 showed obvious invasion ability compare to the control group treated with serum-free DMEM. The invasion ability will not be activated by the FBS. (B) The results of cell invasion in Fig. 9A are calculated by computer software, significant differences are presented in the groups with YKL-40 added. *P<0.05 and
**P<0.01 compared with corresponding control cells treated with serum-free DMEM.
51
(A)
(B)
(C)
0 5 10 15 20 25
0 10% FBS YKL-40 100
ng/mL
YKL-40 1000 ng/mL
% of Area Covered
**
**
60 70 80 90 100 110 120 130 140 150 160
0 10% FBS YKL-40 100
ng/mL
YKL-40 1000 ng/mL
% of Control
**
**
0 10% FBS
100 1000
52
Figure 9. YKL-40 stimulates the invasion ability of CMT-1 cells.
(A) The insert with CMT-1 cells treated with YKL-40 showed obvious invasion ability compare to the control group treated with serum-free DMEM. The invasion ability will not be activated by the FBS. (B) The results of cell invasion in Fig. 9A are calculated by computer software, significant differences are presented in the groups with YKL-40 added. *P<0.05 and
**P<0.01 compared with corresponding control cells treated with serum-free DMEM.
53
Figure 10. Autoantigenicity of YKL-40
The recombinant canine YKL-40 protein was separated with 10% SDS-PAGE and Western blotted, the membrane was probed with dog serum (dog A, B, C, D). The membrane was then stripped and confirmed by anti-His-tag antibody again.
A B C D
YKL-40 (His-tag)
YKL-40 (Serum)
Dog Sera
54
(A)
(B)
Figure 11. The PI3K/AKT and MAPK pathway analysis on CMT-1 and MPG cells
(A) CMT-1 cells pretreated with serum-free DMEM were stimulated with YKL-40 from 10 to 1000 ng/mL and DMEM with 10% FBS for 1 hour. The result showed that pAKT and pERK were induced by YKL-40 in a dose-dependent manner. (B) MPG cells pretreated with serum-free DMEM were stimulated with YKL-40 from 10 to 1000 ng/mL and DMEM with 10% FBS for 1 hour. The result showed that pAKT and pERK were induced by YKL-40, and pAKT was induced in a dose-dependent manner.
Actin
10 100 1000 YKL-40
DMEM w/o FBS
DMEM w/ 10% FBS pAKT
pERK1/2 ERK1/2
AKT
10 100 1000 YKL-40
DMEM w/o FBS
DMEM w/ 10% FBS pAKT
pERK1/2 AKT
ERK1/2 Actin
55
References
Allin KH, Bojesen SE, Johansen JS, Nordestgaard BG. Cancer risk by combined levels of YKL-40 and C-reactive protein in the general population. Br J Cancer 2012;106:199-205.
Baeten D, Boots AMH, Steenbakkers PGA, Elewaut D, Bos E, Verheijden GFM, Verbruggen G, Miltenburg AMM, Rijnders AWM, Veys EM, De Keyser F. Human cartilage gp-39+, CD16+ monocytes in peripheral blood and synovium. Arthritis Rheum 2000;43(6):1233-1243.
Benjamin SA, Lee AC, Saunders WJ. Classification and behavior of canine mammary epithelial neoplasms based on life-span observations in Beagles. Vet Pathol 1999;36:423-436.
Bigg HF, Wait R, Rowan AD, Cawston TE. The mammalian Chitinase-like lectin, YKL-40, binds specifically to type I collagen and modulates the rate of type I collagen fibril formation. J Biol Chem 2006;281(30):21082-21095.
Boots AMH, Hubers H, Kouwijzer M, den Hoed-van Zandbrink L, Westrek-Esselink BM, van Doorn C, Stenger R, Bos ES, van Lierop MC, Verheijden GF, Timmers CM, van Staveren CJ. Identification of an altered peptide ligand based on the endogenously presented, rheumatoid arthritis-associated, human cartilage glycoprotein-39(263-275) epitope: an MHC anchor variant peptide for immune modulation. Arthritis Res Ther 2007;9(4):R71.