中 文 摘 要 : 我們利用斑馬魚魚膘作為在活體模式(in vivo model),發現葉酸缺
乏會阻礙肺臟原始細胞的分化,並引發發炎反應及細胞外基質
(extracellular matrix; ECM)蛋白質酶/蛋白質酶抑制劑之間的失
衡,而導致肺部發育缺陷及類似慢性阻塞性肺病的組織病理特徵。
其主要是由於 cathepsin L和cystatin B (CSTB)-like
(zCSTB-like) proteins的表現失衡所導致。同時我們發現:在斑馬幼魚發育
過程中,cathepsin L 與 zCSTB-like 的表現在空間上剛好互補。
因葉酸缺乏而導致的cathepsin
L過量表現與其抑制蛋白cystatin-b的降低所導致的魚膘發育缺陷可經由補充5-CHO-THF而被成功預防
。
中 文 關 鍵 詞 : 葉酸、慢性阻塞性肺病、斑馬魚疾病模式、蛋白質酶蛋白質酶抑制
劑失衡
英 文 摘 要 : Lung injury is one of the pathological hallmarks of most
respiratory tract diseases including asthma, acute
respiratory distress syndrome (ARDS) and chronic
obstructive pulmonary disease (COPD). It involves
progressive pulmonary tissue damages which are usually
irreversible and incurable. Therefore, measures to
facilitate drug development against lung injury are needed.
Here, we characterized the zebrafish folate-deficiency (FD)
transgenic line that lacks a well-established swim bladder.
Whole-mount in-situ hybridization revealed comparable
distribution patterns of swim bladder tissue markers
between wild-type and FD larvae, suggesting a proper
development of swim bladder in early embryonic stages.
Unexpectedly, neutrophils infiltration was not observed in
the defective swim bladder. Microarray analysis revealed a
significant increase and decreased of the transcripts for
cathepsin L and a cystatin B (CSTB)-like (zCSTB-like)
proteins, respectively, in FD larvae. The distribution of
cathepsin L and the zCSTB-like transcripts was
spatio-temporally specific in developing wild-type embryos and, in
appropriate measure, correlated with their potential roles
in maintaining swim bladder integrity. Supplementing with
5-formyltetrahydrofolate successfully prevented the swim
bladder anomaly and the imbalanced expression of cathepsin
L and the zCSTB-like protein induced by folate deficiency.
Injecting the purified recombinant zebrafish zCSTB-like
protein alleviated FD-induced swim bladder anomaly. We
concluded that the imbalanced expression of cathepsin L and
the zCSTB-like protein contributed to the swim bladder
malformation induced by FD and suggested the potential
application of this transgenic line to model the lung
1
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
A novel zebrafish model to
emulate lung injury by folate
deficiency-induced swim bladder
defectiveness and protease/
antiprotease expression imbalance
Gang-Hui Lee
1,2, Nai-Wei cheng
1, Hsin-Hsuan Yu
1, Jen-Ning tsai
3,4, Tsunglin Liu
5,
Zhi-Hong Wen
6, Bing-Hung chen
7,8,9,10& Tzu-Fun fu
1,2Lung injury is one of the pathological hallmarks of most respiratory tract diseases including asthma, acute respiratory distress syndrome (ARDS) and chronic obstructive pulmonary disease (COPD). It involves progressive pulmonary tissue damages which are usually irreversible and incurable. Therefore, strategies to facilitate drug development against lung injury are needed. Here, we characterized the zebrafish folate-deficiency (FD) transgenic line that lacks a fully-developed swim bladder. Whole-mount in-situ hybridization revealed comparable distribution patterns of swim bladder tissue markers between wild-type and FD larvae, suggesting a proper development of swim bladder in early embryonic stages. Unexpectedly, neutrophils infiltration was not observed in the defective swim bladder. Microarray analysis revealed a significant increase and decrease of the transcripts for cathepsin L and a cystatin B (CSTB)-like (zCSTB-like) proteins, respectively, in FD larvae. The distribution of cathepsin L and the zCSTB-like transcripts was spatio-temporally specific in developing wild-type embryos and, in appropriate measure, correlated with their potential roles in maintaining swim bladder integrity. Supplementing with 5-formyltetrahydrofolate successfully prevented the swim bladder anomaly and the imbalanced expression of cathepsin L and the zCSTB-like protein induced by folate deficiency. Injecting the purified recombinant zebrafish zCSTB-like protein alleviated FD-induced swim bladder anomaly. We concluded that the imbalanced expression of cathepsin L and the zCSTB-like protein contributed to the swim bladder malformation induced by FD and suggested the potential application of this transgenic line to model the lung injury and ECM remodeling associated with protease/protease inhibitor imbalance.
Lung injury is one of the pathological hallmarks of most respiratory tract diseases including asthma, acute res-piratory distress syndrome (ARDS) and chronic obstructive pulmonary disease (COPD). It involves progres-sive pulmonary tissue damages which are usually irreversible and incurable. Patients usually receive drug or
1Department of Medical Laboratory Science and Biotechnology, National Cheng Kung University, Tainan, Taiwan. 2Institute of Basic Medical Science, National Cheng Kung University, Tainan, Taiwan. 3Department of Medical
Laboratory and Biotechnology, Chung Shan Medical University, Taichung, Taiwan. 4Clinical Laboratory, Chung Shan
Medical University Hospital, Chung Shan Medical University, Taichung, Taiwan. 5Department of Biotechnology and
Bioindusry Sciences, National Cheng Kung University, Tainan, Taiwan. 6Department of Marine Biotechnology and
Resources, Asia-Pacific Ocean Research Center, National Sun Yat-sen University, Kaohsiung, Taiwan. 7Department
of Biotechnology, Kaohsiung Medical University, Kaohsiung, Taiwan. 8Department of Medical Research, Kaohsiung
Medical University Hospital, Kaohsiung Medical University, Kaohsiung, Taiwan. 9Centers for Biomarkers and
Biotech Drugs, Kaohsiung Medical University, Kaohsiung, Taiwan. 10The Institute of Biomedical Sciences, National
Sun Yat-sen University, Kaohsiung, Taiwan. Gang-Hui Lee, Nai-Wei Cheng and Jen-Ning Tsai contributed equally. Correspondence and requests for materials should be addressed to B.-H.C. (email: [email protected]) or T.-F.F. (email: [email protected])
Received: 11 June 2019 Accepted: 19 August 2019 Published: xx xx xxxx
2
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
supportive therapies that relieve symptoms but do not halt or reverse lung damage1. Therefore, there is a need to understand the pathophysiological mechanisms so as to develop new preventive/therapeutic strategies and the tools that could expedite drug discovery.
Although the underlying molecular mechanism still remains not completely understood, the progressive lung tissue damage is likely, at least in part, due to the consequence of extracellular matrix (ECM) destruction owing to increased pulmonary proteases activity, which often results from an imbalanced expression of tissue-degrading enzyme and their inhibitors2–5. ECM is a supporting structure for morphogenesis and central to lung physiology for maintaining mechanical strength and elasticity, normal interstitial fluid dynamics, effective gas exchange, proper control of cell behavior and tissue repair and remodeling6,7. The composition and functions of ECM are regulated by the mutual effect on ECM protein synthesis and the proteolytic activity attributed to the abun-dant ECM proteases, such as matrix metalloproteinases and cathepsins. Elevated protease activity, either due to increased protease expression, decreased inhibitors or both, is often associated with excessive extracellular turn-over and defective bronchopulmonary development and worsening of respiratory diseases8. Therefore, strategies and drugs capable of maintaining the balance between prominent proteases and their corresponding inhibitors could lead to the development of an effective treatment9–11.
Studies have shown that folate status influences lung development and disease risk, albeit the effects are con-troversial and the underlying mechanisms remain largely unexplored. Folate, also known as vitamin B9, provides the one-carbon unit for the biosynthesis of numerous macromolecules, including amino acids, proteins, nucleo-tides, several neurotransmitters and S-adenosylmethionine. S-adenosylmethionine is the primary methyl-donor for DNA, histone, protein and lipid methylation and hence is capable of modulating gene expression in a tissue-, site-, gene- and even gender-specific manner12–14. Reduced folate also possesses strong anti-oxidative activity comparable to vitamin C and vitamin E, which is crucial to maintaining proper intracellular oxidative stress and embryogenesis15–17. Owing to its multi-activities, folate has been considered as a modifiable environmental factor and a nutraceutical that can modulate the “developmental origin of health and disease (DOHaD)”18. Nevertheless, folate metabolic disturbance and folate intake imbalance, either excessive or insufficient, can lead to harmful consequences. High serum folate levels were associated with a low risk of high total serum IgE concentrations, atopy, and wheezing19. Nevertheless, folic acid supplementation during pregnancy was reported to be associ-ated with an increased risk of infant bronchiolitis and childhood asthma20,21. These conflicting results raise the concern and debate on the policy of mandatory fortification of food and maternal folic acid supplementation, a highly prevalent maneuver world-wide. To date, studies pertaining to the pathophysiological effects between folate supplementation and lung injury in general still remain scarce. Therefore, it is imperative to elucidate how folate affects lung physiology and the pathogenesis of pulmonary diseases to help further discern the nutritional importance of folate.
With the combined advantages of in vivo complexity and in vitro convenience of high-throughput screening, zebrafish is a vertebrate model prominent for research on various fields including molecular function, diseases mechanism, developmental biology, toxicology and drug discovery22–27. Currently, efforts to delineate the path-omechanism of and treatment for diseases involving pulmonary tissue damage have been greatly assisted by rodent models28–30. Possessing the advantages of “ in vitro convenience” and “ in vivo complexity”, zebrafish is ideal to complement rodent for a real-time, dynamic and high-throughput observation and is especially powerful for drug discovery. Specifically to lung research, zebrafish swim bladder provides an ECM environment, which cannot be achieved by cultured cells, and has been suggested to be a simple in vivo platform for the study on lung elastin injury and repair31,32. Zebrafish swim bladder is a buoyancy organ located in the dorsal-anterior part of the body cavity. Similar to mammalian lung, teleost swim bladder originates from an outgrowth of the foregut endo-derm into an out-pocketing gas-filled sac during embryonic development33. Zebrafish swim bladder is mainly composed of three distinct tissue layers: endodermal epithelium, the middle mesenchymal layer and the outer mesothelium layer34. The gene products closely related to the structural and functional elements of lung tissue are also enriched in swim bladder35. Conserved expression of genes involved in Hedgehog (Hh) signaling was observed in developing swim bladder, as in early lung development34. Transcriptomic analysis suggested a role for ECM in the development and function of zebrafish swim bladder, as for the human bronchopulmonary system8,35. Examination on the zebrafish swim bladder wall revealed a simple structure resembling an inner layer of elastin fibers31. The resemblance of structural, ontogenic and molecular characteristics between zebrafish swim bladder and mammalian lung supports the use of the zebrafish swim bladder to model lung physiology and pathology.
Previously, we had established a fluorescent zebrafish transgenic line that displays folate deficiency (FD) upon induction in a timing-, duration- and extent-controllable manner17. We found that most of these FD larvae lacked successfully inflated swim bladder. In this study, we examined the histopathological characteristics of these FD larvae and the potential pathways that contributed to the impeded swim bladder formation. Additionally, we iden-tified a previously uncharacterized zebrafish protein, named zebrafish cystatin B (zCSTB)-like, which is structur-ally and functionstructur-ally comparable to its human cystatin B ortholog and whose expression was down-regulated in response to FD induction in our zebrafish model. The causal link between FD and the pathological phenotypes was confirmed by folate supplementation. The mechanisms underlying the FD-induced swim bladder malforma-tion and the potential of using this transgenic line to model lung injury are also proposed and discussed.
Materials and Methods
Materials.
The SMARTTM RACE amplification kit was purchased from Clontech. Takara Bio Co. (Mountain3
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
from Sigma-Aldrich Chemical Co. (Montana, US). The E. coli strains and vectors used for cloning and protein expression have been described previously36.
Fish maintenance.
Zebrafish (Danio rerio, AB strain) were maintained following the standard procedure37. The animal studies and all procedures were approved by Affidavit of Approval of Animal Use Protocol of National Cheng-Kung University (IACUC Approval NO. 96062). All procedures complied with the “Guidelines for the Use of Zebrafish” (Chapter 22) in the “A Guidebook for the Care and Use of Laboratory Animals” (3rd edition), published by Taiwan Society of Laboratory Animal Sciences, Council of Agriculture, Executive Yuan, Taiwan (https://animal.coa.gov.tw/html/index_04_3_1.html). The maintenance of FD transgenic lines, Tg(mpx:EGFP), Tg(lfabp:mCherry/hsp:EGFP-γGH) and Tg(hsp:EGFP-γGH), and the protocols used to induce folate deficiency were as previously described17.Whole mount in-situ hybridization (WISH).
WISH was performed following the stand-ard protocol37 with the probes prepared from the plasmids for Hb9, Acta2, Anax538, zebrafish cys-tatin B-like protein (zCSTB-like) and cathepsin L. Both zebrafish cathepsin L and CSTB-like cDNA encompassing partial 3′-coding sequence and 3′-UTR were cloned using the following primers: cathepsin L:5′-GGCATATGGCTCCATCTATAGAC-3′(forward) and 5′-CGCTCGAGCATTAGGGGATAG-3′(reverse); zCSTB-like:5′-CGAGAATTCATCAACAATGTCAGAG-3(forward) and 5′-CACGGATCCTGCAAATGCTC-3′(reverse). Digoxigenin-UTP-labeled antisense RNA probes were synthesized using DIG-RNA Labeling kit (SP6/T7) (Roche).Histochemical staining.
Cryosectioning and H&E staining of zebrafish tissue sections were performed fol-lowing the protocols in Zebrafish Book37. Histochemical staining for swim bladder was performed using elastin/ collagen staining kit following the manufacture’s instruction (Sigma-Aldrich).Oil red O (ORO) staining and Sudan black staining.
ORO staining was performed according to pub-lished protocol by Schlegel and Stainier39. Generally, zebrafish larvae were fixed by 4% PFA/PBS and soaked in 1, 2-propanediol. Then, larvae were stained with filtered 0.5% ORO in 1, 2-propanediol for 16 hours. ORO-stained larvae were stored in 80% glycerol/PBS after briefly washing in PBS. Sudan black staining were performed follow-ing the protocols previously described40.Cathepsin L and zCSTB-like expression.
Reverse-transcription PCR (RT-PCR) was per-formed using the following primers: cathepsin L: 5′-GCTGATGTTGCTGGGAATAG-3′(forward) and 5′-CCACTATCATAGCCCTAACAGC-3′(reverse); zCSTB-like: 5′-GTGTTTATTGCTGGAGATGAGTGTGC-3′(forward) and 5′-GAGAGTTTAATGTTGAGGAATGAAGCAG-3′(reverse) as previously described. For pro-tein analysis, Western blotting was performed using the polyclonal antibodies against cathepsin L and tubulin (GeneTax, Taiwan)17.Cloning for zebrafish CSTB-like.
The primers used to PCR clone the zCSTB-like coding sequence from zebrafish 5′-RACE-Ready cDNA libraries were designed based on the nucleotide sequence of the uncharacterized protein (GenBankTM accession number EH546362), whose transcript showed an 83-folddecrease in FD larvae. The primer sequences were: 5′-CATATGTCAGAGGCAAAGGCAGG-3′ (forward) and 5′-CGCTCGAGTTTCTTAGGAAGATTCAG-3′ (reverse) with introduced NdeI and XhoI restriction enzyme sites (underlined). The PCR fragments of 318 base-pairs were cloned into the pET43.1a expression vector between its NdeI and XhoI sites, generating zebrafish CSTB-like-His/pET43.1a. Successful cloning was confirmed by restriction enzyme digestion and DNA sequencing. The resulting constructs were transformed into Rosetta (DE3) E. coli for protein expression.
Expression and purification of recombinant zebrafish CSTB-like.
The expression and purification of recombinant zebrafish CSTB-like protein was performed using nickel-affinity column following the protocols described previously with minor modifications41. In brief, E. coli containing zebrafish CSTB-like-His/pET43.1a, that expressed recombinant zebrafish CSTB-like protein with C-terminal His-tag, was grown in enriched Luria broth (2-YT) and induced with IPTG at log phase. After 4 hrs induction, bacteria were pelleted and disrupted in lysis buffer (20 mM sodium phosphate, pH 7.4, 0.5 M NaCl, 5 mM 2-mercaptoethanol, 1 mM MgCl2 and 10%glycerol) with sonication. After removing the majority of chromatin with DNase I, cell lysates were subjected to affinity chromatography using a Nickel-Sepharose column (1.5 × 3 cm) for purifying zebrafish CSTB-like protein. Concentrated recombinant zebrafish CSTB-like protein was stored in phosphate buffer containing 10% glycerol at −80 °C.
Cystatin B activity assay.
The protease inhibitory property of recombinant zebrafish CSTB-like was exam-ined based on the inhibition on papain activity for hydrolyzing azo-casein42. In brief, papain (1 mg/ml in 0.1 M phosphate buffer, pH7.4) was incubated with various amounts of recombinant zebrafish CSTB-like (concentra-tions ranging from 5 to 500 ng/μl) and incubated at 25 °C for 10 min before adding 125 µl of 0.5% azo-casein pre-pared in PBS. After incubating for another 30 min at 37 °C, reactions were stopped with 10% trichloroacetic acid and centrifuged. The supernatant was measured for absorbance at 440 nm. The percentage of remaining papain activity was calculated from the ratio of absorbance with and without zebrafish CSTB-like using the formula: 100 × (A440nm with CSTB-like/A440 nm without CSTB-like).4
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
FD larvae at the region surrounding swim bladder. Larvae injected with PBS served as injection control. Injected larvae were grown in embryo water and observed for swim bladder inflation at 5 dpf.
Results
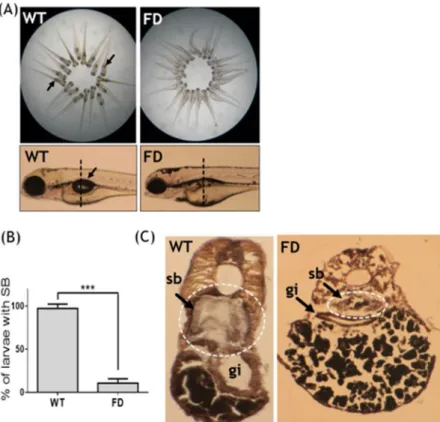
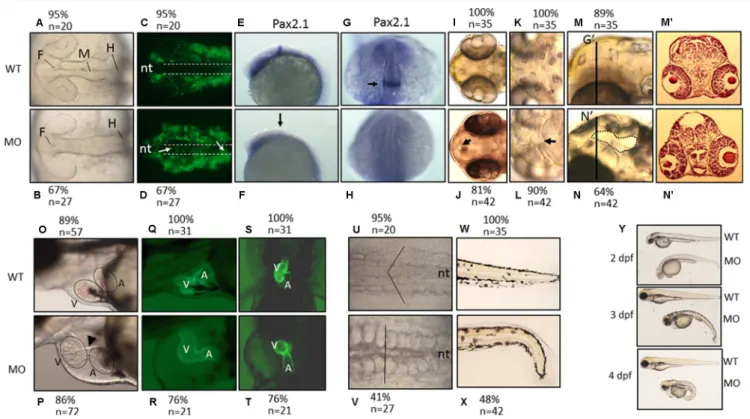
Defective swim bladder was observed in FD larvae.
The previously established fluorescent transgenic line Tg(hsp:eGFP-GGH) will overexpress a fusion of eGFP with γ-glutamylhydrolase (GGH) upon heat-shock, which facilitates the exportation of intracellular folate and leads to folate deficiency with the extent positively cor-related to its fluorescence intensity17. Normally, the inflated single-chambered swim bladder can be observed at 5 dpf in growing zebrafish larvae. However, more than 80% of FD larvae lacked a visible inflated swim bladder at 5 dpf (Fig. 1A,B). Cryo-sections prepared from 5-dpf wild-type larvae revealed an intact and properly inflated swim bladder (Fig. 1C). Contrarily, the swim bladder remnants resulting from unsuccessful inflation was observed in FD larvae. The absence of a successfully inflated swim bladder indicated impeded formation of swim bladder in FD larvae.The expression patterns of swim bladder tissue markers in early development were
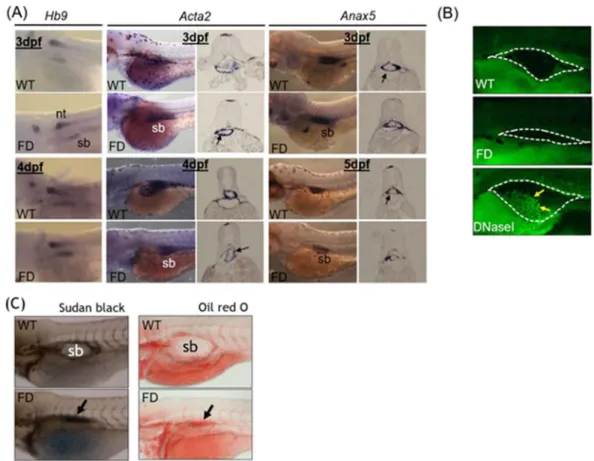
compara-ble between wild-type larvae and FD larvae.
Whole-mount in-situ hybridization (WISH) is a strategy commonly used to examine the organ development and tissue integrity in developing embryos, in which the tissue morphology is visualized by the synthetic riboprobes complementary to the transcripts expressed specifi-cally in the tissues of interest. Comparable expression patterns and intensity for all three probes (Hb9, Acta2 andAnax5) corresponding to the three major tissue layers of zebrafish swim bladder were observed for both wild-type
control and FD embryos at 3 dpf, nevertheless, the signals became weaker in FD larvae at later stages (Fig. 2A). Results from a TUNEL assay revealed no apparent signal for apoptosis in the swim bladder of both control and FD larvae (Fig. 2B). To investigate the integrity of swim bladder, larvae at 5 dpf were subjected to either Sudan black or Oil red O staining. With both staining methods, the swim bladder of the heat-shocked wild-type con-trol larvae was revealed as a clear outline surrounding a morphologically sound and inflated air chamber area (Fig. 2C). On the contrary, a rather shrunk and darker stain patch was observed in the 5-dpf FD larvae, indicating a failed inflation of the air chamber possibly due to a swim bladder leakage. These data suggested that the devel-opment of swim bladder before 3 dpf was not affected by FD status. However, the tissue integrity of swim bladder might have been damaged in FD larvae.
5
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
The transcripts of cathepsin L and a cystatin B-like (CSTB-like) protein were significantly
increased and decreased, respectively, in FD larvae.
To examine the impact of folate deficiency on the global gene expression profile during zebrafish embryogenesis, FD embryos at 14 hpf, 30 hpf and 120 hpf were subjected to microarray analysis. The expression profiles of both wild-type embryos and a transgenic line express-ing a non-functional γ-glutamylhydrolase were also examined to serve as comparison17. Of all the genes display-ing affected expression levels, cathepsin L and a CSTB-like protein (GenBankTM accession number EH546362)were among those with the most apparent changes at 120 hpf (Table 1). The alteration in the mRNA levels of cathepsin L and zCSTB-like in FD larvae was confirmed with RT-PCR (Fig. 3A). Additionally, the cathepsin L transcript expression levels were increased and exhibited a positive correlation with the FD severity of the larvae. Western blotting results showed that cathepsin L was abundant in wild-type embryos at 48 hpf but vanished after Figure 2. The tissue integrity and pathology of the swim bladder in wild-type and FD embryos and larvae. Heat-shocked larvae of both wild-type control and Tg(hsp:EGFP-GGH) were harvested at indicated stages post embryogenesis and analyzed for swim bladder development. (A) Embryos were subjected to WISH with tissues specific probes including Hb9 (epithelial layer), Acta2 (mesenchyme layer, smooth muscle) and Anax5 (outer mesothelium layer). Cross-sections of the embryos after WISH staining revealed comparable intensity and morphology for the tissue layers composed of swim bladders (black arrows) in the early embryos. (B) Larvae at 5 dpf examined with TUNEL assay for apoptotic cells (arrows) revealed no positive signal in the swim bladder area (circled by dash-line) of both wild-type control and FD larvae. Embryos pre-treated with DNase I served as a positive control. (C) Larvae at 5 dpf were stained with Sudan black (left) and Oil red O (right) to reveal outlines of swim bladder (black arrows) in larvae. nt, neural tube; sb, swim bladder; WT, wild-type larvae with heat-shock; FD, FD larvae with heat-shock.
Access. No. Description
Fold change in expression (FD/ SDM)*
14 hpf 31 hpf 120 hpf
NM_131198 Danio rerio cathepsin L, 1b −1.071 −1.119 +6.92 EH546362 Danio rerio cystatin B-like −1.487 1.058 −107.157
Table 1. The fold of expressional change for cathepsin L and CSTB-like obtained from microarray data.
*Reported is the averaged transcripts ratio between FD and SDM larvae of all probes (ranging from 1 to 5 probes
6
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
56 hpf. Contrarily, cathepsin L protein levels persisted in FD embryos until 120 hpf (Fig. 3B). The concurrent decrease of the zCSTB-like and increase of cathepsin L expression implied a possible elevation of cathepsin L activity in FD larvae.
The expression of cathepsin L and zCSTB-like was spatio-temporally specific in developing
embryos.
The WISH with riboprobes specific to cathepsin L and CSTB-like revealed a stringent restriction for their expression in developing embryos. The WISH signal for cathepsin L was not detected in early embryos until 12 hpf (Fig. 4A). An intense signal patch appeared at the anterior prechordal plate, the most likely origin of the rostral cranial mesoderm and an integral part of the roof of the foregut. The signal was focused in the hatch gland of embryos at 1 to 3 dpf but disappeared later. At 4 dpf, the cathespin L signal was observed in head region and swim bladder. Contrarily, the CSTB-like transcripts were distributed homogenously in early embryos, but focused in the region surrounding swim bladder at later stages (Fig. 4B). Abundant and homogenous distribu-tion of CSTB-like transcripts was observed in embryos before 4 hpf, even as early as the 4-cell stage, suggesting the deposition of maternal mRNA. Zebrafish CSTB-like signals became focused anteriorly in 1 dpf embryos and gradually diminished, as development proceeded. A signal of moderate intensity was observed in the trunk along the vessels at 3 dpf. Zebrafish CSTB-like transcripts were abundant in heart at 4 dpf but diminished at 5 dpf. Apparent signal was visible in the region surrounding the swim bladder at 4 dpf and became enriched at 5 dpf. The seemingly complementary expression patterns of cathepsin L and CSTB-like imply a potential mutual restraint in their functions between these two proteins during embryogenesis.The structure and activity of zebrafish CSTB-like are comparable to mammalian
ort-hologs.
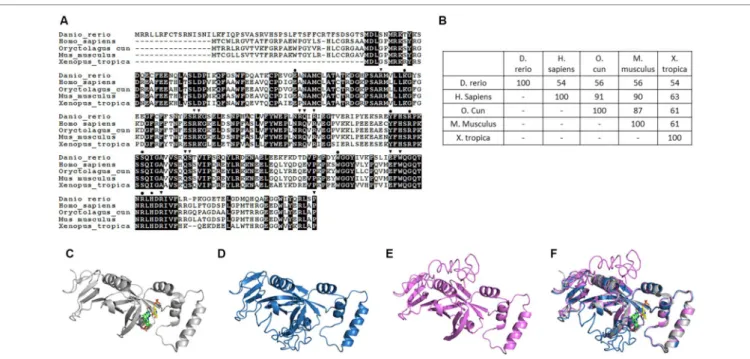
Amino acid sequence alignments reveal an approximate 45% homology and 32% identity between zebrafish CSTB-like and human cystatin B, which are the highest among the comparisons between zebrafish CSTB-like with other cystatins (Fig. 5A). Similarly, secondary structures, including a helix at the N-terminus followed by four consecutive β-strands, were predicted for both human cystatin B and zebrafish CSTB-like (Fig. 5B). The helix was situated right in front of the four-stranded β-sheet in the predicted 3D structure for both zCSTB-like and human CSTB (Fig. 5C). The structural motifs crucial for interacting with target enzymes, such as the Q-V-G motif, were also present in zebrafish CSTB-like43. The recombinant zebrafish CSTB-like was cloned, overexpressed in E. coli and purified to at least 90% purity (Fig. 5D). The purified recombinant zebrafish CSTB-like displayed inhibitory activity on the hydrolase activity of the cysteine protease papain. These results suggested structural and functional similarity between zebrafish CSTB-like and the proteins in the cystatin B family, supporting the cystatin B identity of zebrafish CSTB-like.7
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
the expression levels of cathepsin L and zCSTB-like in FD larvae (Fig. 6A, bottom panel). It should be noted that adding folate to wild-type embryos also increased the expression of both cathepsin L and zCSTB-like, yet the mRNA ratio between the two remains unchanged. The percentage of FD larvae with successfully inflated swim bladder was also increased when embryos were rescued with 5-FTHF (Fig. 6B). Unexpectedly, adding folic acid did not exert the same rescuing effect as with 5-FTHF. Adding N-acetylcysteine (NAC), a strong antioxidant, to embryo water did not prevent the impeded swim bladder formation induced by FD either (Fig. 6B). These results suggested that increased oxidative stress due to FD did not contribute to the defective swim bladder observed in FD larvae. These data supported the causal link between FD and the swim bladder anomaly, as well as the altered expression of cathepsin L and zCSTB-like observed in FD larvae. In addition, the expression of both cathepsin L and zCSTB-like was susceptible to folate modulation. Our results also revealed the diverse biological activities of different folate derivatives, at least between 5-FTHF and folic acid.
8
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
examine the contribution of zCSTB-like in swim bladder formation. The percentage of FD larvae with successfully inflated swim bladder was significantly and dose-dependently increased with regard to the injected zCSTB-like concentration (Fig. 6C,D). These results supported the causal role of imbalanced cathepsin L/zCSTB-like activity for the impeded swim bladder formation in FD larvae.
Discussion
Imbalanced protease/antiprotease activity is central to the current hypothesis for the pathomechanism of most respiratory tract diseases involving lung injury. In the current study, we showed cathepsin L and zCSTB-b com-plementarily express in developing zebrafish larvae at the antecedent of swim bladder and their expression pro-foundly modulate the integrity of swim bladder. Moreover, the increased cathepsin L and decreased zCSTB-b expression were observed in FD larvae. This imbalanced protease/antiprotease activity in response to FD contrib-uted to the impaired swim bladder formation. This disturbed protease/antiprotease balance is also analogous to the pathomechanism of lung injury observed in emphysema, supporting the use of this model for related studies and drug discovery.
9
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
10
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
(DHFR) inhibitor. DHFR converts folic acid and dihydrofolate to tetrahydrofolate, the active coenzyme. 5-FTHF is the reduced folate derivative often used in methotrexate combined therapy for treating cancers. Therefore, it is surmisable that FD increases an individual’s vulnerability to environmental toxins and/or other risk factors causing COPD. This speculation is supported by the increased mortality and the rate of malformation when FD embryos were exposed to caffeine and ethanol as compared to wild-type control (unpublished data). One other interesting observation is that both cathepsin L and zCSTB-like transcripts were increased in wild-type larvae exposed to 5-FTHF but the ratio between this protease/antiprotease pair remained unaffected. These results help further emphasize the importance of maintaining protease/antiprotease balance in order to sustain tissue integ-rity. Understanding how the expressions of these protease/antiprotease are regulated, and how folate functions in helping maintain ECM integrity and lung health shall be beneficial to optimizing disease management and improving healthcare regimens for COPD patients.
While adding 5-FTHF significantly prevented the FD-induced swim bladder anomaly, supplementing with folic acid failed to rescue was rather unexpected. Folic acid is the inactive and oxidized form of folate commonly included in the food and nutrient supplement available in the neighborhood drug stores. As aforementioned, folic acid needs to be further reduced by DHFR in order to be physiologically active. Unlike reduced folate, folic acid lacks appreciable antioxidant activity, raising the possibility that the defective swim bladder formation was due to increased oxidative stress caused by FD17. Nevertheless, the presence of antioxidant NAC did not prevent against swim bladder impairment observed in FD larvae in our current studies. These results indicated that the swim bladder anomaly was possibly not caused by the direct elevated oxidative stress in FD larvae. The other clue comes from the altered expression of cathepsin L and CSTB-like observed in FD larvae, which implies a dis-turbed intracellular gene activity. Gene activity is known to be modulated by intracellular methylation potential (i.e. the levels of S-adenosylmethionine and 5-methyltetrahydrofolate) via epigenetic control. We had previously shown that zebrafish DHFR is structurally and functionally comparable to mammalian orthologs and is abun-dant in developing embryos54,55. Therefore, the supplemented folic acid should have been converted to reduced folate and utilized by larvae. However, it should be noted that folic acid is a less effective substrate of DHFR. The presence of high-dose of folic acid had been shown to repress the conversion of dihydrofolate to tetrahydrofolate
in vitro54. Moreover, elevated cellular levels of dihydrofolate had been suggested to cause inhibition on the activ-ity of methylenetetrahydrofolate reductase, the enzyme catalyzing the production of 5-methyltetrahydrofolate56. Therefore, the disturbed methylation potential caused by FD and folic acid supplementation might have contrib-uted to the impaired expression of cathepsin L/cystatin B expression, as well as the unsuccessful rescues with folic acid, respectively. Previously, we had shown that supplementing with different folate derivatives exerted different physiological impacts on the receiving organisms, suggesting distinct and non-interchangeable activity between different folate derivatives55,57. The notion that folic acid failed to rescue FD-induced swim bladder malformation further supports the diverse activities among different folate derivatives. Further investigation, especially charac-terization on the distribution of different one-carbon derivatives in folate pool under the circumstance of FD and folate supplementation shall provide additional insights to the pathological mechanisms underlying FD and for the best and safe use of folate supplements.
In conclusion, we showed that the concurrent increase and decrease of cathepsin L and CSTB-like expres-sion, respectively, contributed to the impeded swim bladder formation observed in FD larvae. Local injection of recombinant zebrafish CSTB-like protein, which is structurally and functionally comparable to human cystatin B, successfully prevented the FD-induced swim bladder malformation. Diverse effectiveness of rescue mediated by different folate adducts were also observed, in which successful rescue was found when 5-FTHF supplementation, but not folic acid, was used. Our studies provide the evidence to surmise the connection between the individual’s folate status to ECM integrity, and potentially pulmonary function, by intervening protease/antiprotease balance.
References
1. Mercado, N., Ito, K. & Barnes, P. J. Accelerated ageing of the lung in COPD: new concepts. Thorax 70, 482–489, https://doi. org/10.1136/thoraxjnl-2014-206084 (2015).
2. Greene, C. M. & McElvaney, N. G. Proteases and antiproteases in chronic neutrophilic lung disease - relevance to drug discovery.
British journal of pharmacology 158, 1048–1058, https://doi.org/10.1111/j.1476-5381.2009.00448.x (2009).
3. Kulkarni, T., O’Reilly, P., Antony, V. B., Gaggar, A. & Thannickal, V. J. Matrix Remodeling in Pulmonary Fibrosis and Emphysema.
American journal of respiratory cell and molecular biology. https://doi.org/10.1165/rcmb.2015-0166PS (2016).
4. Haq, I., Lowrey, G. E., Kalsheker, N. & Johnson, S. R. Matrix metalloproteinase-12 (MMP-12) SNP affects MMP activity, lung macrophage infiltration and protects against emphysema in COPD. Thorax 66, 970–976, https://doi.org/10.1136/thx.2011.159087
(2011).
5. Ariestanti, D. M., Ando, H., Hirose, S. & Nakamura, N. Targeted Disruption of Ig-Hepta/Gpr116 Causes Emphysema-like Symptoms That Are Associated with Alveolar Macrophage Activation. Journal of biological chemistry 290, 11032–11040, https://doi. org/10.1074/jbc.M115.648311 (2015).
6. Shih, H. P., Panlasigui, D., Cirulli, V. & Sander, M. ECM Signaling Regulates Collective Cellular Dynamics to Control Pancreas Branching Morphogenesis. Cell reports 14, 169–179, https://doi.org/10.1016/j.celrep.2015.12.027 (2016).
7. Pelosi, P., Rocco, P. R., Negrini, D. & Passi, A. The extracellular matrix of the lung and its role in edema formation. Anais da
Academia Brasileira de Ciencias 79, 285–297, https://doi.org/10.1590/S0001-37652007000200010 (2007).
8. Mizikova, I. & Morty, R. E. The Extracellular Matrix in Bronchopulmonary Dysplasia: Target and Source. Frontiers in medicine 2, 91,
https://doi.org/10.3389/fmed.2015.00091 (2015).
9. Meyer, M. & Jaspers, I. Respiratory protease/antiprotease balance determines susceptibility to viral infection and can be modified by nutritional antioxidants. American journal of physiology. Lung cellular and molecular physiology 308, L1189–1201, https://doi. org/10.1152/ajplung.00028.2015 (2015).
10. Mohanka, M., Khemasuwan, D. & Stoller, J. K. A review of augmentation therapy for alpha-1 antitrypsin deficiency. Expert opinion
on biological therapy 12, 685–700, https://doi.org/10.1517/14712598.2012.676638 (2012).
11. Gardi, C., Stringa, B. & Martorana, P. A. Animal models for anti-emphysema drug discovery. Expert opinion on drug discovery 10, 399–410, https://doi.org/10.1517/17460441.2015.1016871 (2015).
11
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
13. Boeke, C. E. et al. Gestational intake of methyl donors and global LINE-1 DNA methylation in maternal and cord blood: prospective results from a folate-replete population. Epigenetics: official journal of the DNA Methylation Society 7, 253–260, https://doi. org/10.4161/epi.7.3.19082 (2012).
14. Barua, S., Kuizon, S. & Junaid, M. A. Folic acid supplementation in pregnancy and implications in health and disease. Journal of
biomedical science 21, 77, https://doi.org/10.1186/s12929-014-0077-z (2014).
15. Gliszczynska-Swiglo, A. & Muzolf, M. pH-dependent radical scavenging activity of folates. Journal of agriculture and food chemistry
55, 8237–8242, https://doi.org/10.1021/jf070986x (2007).
16. Rezk, B. M., Haenen, G. R., van der Vijgh, W. J. & Bast, A. Tetrahydrofolate and 5-methyltetrahydrofolate are folates with high antioxidant activity. Identification of the antioxidant pharmacophore. FEBS Letters 555, 601–605, S0014579303013589 [pii (2003). 17. Kao, T. T. et al. Folate deficiency-induced oxidative stress contributes to neuropathy in young and aged zebrafish–implication in
neural tube defects and Alzheimer’s diseases. Neurobiology of disease 71, 234–244, https://doi.org/10.1016/j.nbd.2014.08.004 (2014). 18. Rosenfeld, C. S. Animal models to study environmental epigenetics. Biology of reproduction 82, 473–488, https://doi.org/10.1095/
biolreprod.109.080952 (2010).
19. Matsui, E. C. & Matsui, W. Higher serum folate levels are associated with a lower risk of atopy and wheeze. Journal of allergy and
clinical immunology 123, 1253–1259 e1252, https://doi.org/10.1016/j.jaci.2009.03.007 (2009).
20. Whitrow, M. J., Moore, V. M., Rumbold, A. R. & Davies, M. J. Effect of supplemental folic acid in pregnancy on childhood asthma: a prospective birth cohort study. American journal of epidemiology 170, 1486–1493, https://doi.org/10.1093/aje/kwp315 (2009). 21. Veeranki, S. P. et al. Association of folic acid supplementation during pregnancy and infant bronchiolitis. American journal of
epidemiology 179, 938–946, https://doi.org/10.1093/aje/kwu019 (2014).
22. Kermi, C., Aze, A. & Maiorano, D. Preserving Genome Integrity During the Early Embryonic DNA Replication Cycles. Genes (Basel)
10, https://doi.org/10.3390/genes10050398 (2019).
23. Benchoula, K. et al. The promise of zebrafish as a model of metabolic syndrome. Exp Anim, https://doi.org/10.1538/expanim.18-0168
(2019).
24. Wrighton, P. J., Oderberg, I. M. & Goessling, W. There’s Something Fishy About Liver Cancer: Zebrafish Models of Hepatocellular Carcinoma. Cellular and molecular gastroenterology and hepatology, https://doi.org/10.1016/j.jcmgh.2019.05.002 (2019). 25. Fichi, G. et al. Fishing in the Cell Powerhouse: Zebrafish as A Tool for Exploration of Mitochondrial Defects Affecting the Nervous
System. International journal of molecular sciences 20, https://doi.org/10.3390/ijms20102409 (2019).
26. Oehlers, S. H. et al. Expression of zebrafish cxcl8 (interleukin-8) and its receptors during development and in response to immune stimulation. Developmental and comparative immunology 34, 352–359, https://doi.org/10.1016/j.dci.2009.11.007 (2010). 27. Geary, B. et al. Acute stress alters the rates of degradation of cardiac muscle proteins. Journal of proteomics 191, 124–130, https://doi.
org/10.1016/j.jprot.2018.03.015 (2019).
28. Wright, J. L. & Churg, A. Animal models of COPD: Barriers, successes, and challenges. Pulmonary pharmacology & therapeutics 21, 696–698, https://doi.org/10.1016/j.pupt.2008.01.007 (2008).
29. Beckett, E. L. et al. A new short-term mouse model of chronic obstructive pulmonary disease identifies a role for mast cell tryptase in pathogenesis. The Journal of allergy and clinical immunology 131, 752–762, https://doi.org/10.1016/j.jaci.2012.11.053 (2013). 30. Bastarache, J. A. & Blackwell, T. S. Development of animal models for the acute respiratory distress syndrome. Disease models &
mechanisms 2, 218–223, https://doi.org/10.1242/dmm.001677 (2009).
31. Perrin, S., Rich, C. B., Morris, S. M., Stone, P. J. & Foster, J. A. The zebrafish swimbladder: A simple model for lung elastin injury and repair. Connective tissue research 40, 105–112, https://doi.org/10.3109/03008209909029106 (1999).
32. Gratacap, R. L., Bergeron, A. C. & Wheeler, R. T. Modeling mucosal candidiasis in larval zebrafish by swimbladder injection. Journal
of visualized experiments: JoVE, e52182, https://doi.org/10.3791/52182 (2014).
33. Finney, J. L., Robertson, G. N., McGee, C. A., Smith, F. M. & Croll, R. P. Structure and autonomic innervation of the swim bladder in the zebrafish (Danio rerio). Journal of comparative neurology 495, 587–606, https://doi.org/10.1002/cne.20948 (2006). 34. Winata, C. L. et al. Development of zebrafish swimbladder: The requirement of Hedgehog signaling in specification and organization
of the three tissue layers. Developmental biology 331, 222–236, https://doi.org/10.1016/j.ydbio.2009.04.035 (2009).
35. Zheng, W. et al. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung.
PloS one 6, e24019, https://doi.org/10.1371/journal.pone.0024019 (2011).
36. Chang, W. N., Lin, H. C. & Fu, T. F. Zebrafish 10-formyltetrahydrofolate dehydrogenase is similar to its mammalian isozymes for its structural and catalytic properties. Protein expression and purification 72, 217–222, S1046-5928(10)00107-5 [pii, https://doi. org/10.1016/j.pep.2010.04.003 (2010).
37. Westerfield, M. THE ZEBRAFISH BOOK: A guide for the laboratory use of zebrafish (Danio rerio). 5 edn, (University of Oregon Press, 2007).
38. Yin, A., Korzh, S., Winata, C. L., Korzh, V. & Gong, Z. Wnt signaling is required for early development of zebrafish swimbladder.
PloS one 6, e18431, https://doi.org/10.1371/journal.pone.0018431 (2011).
39. Schlegel, A. & Stainier, D. Y. Microsomal triglyceride transfer protein is required for yolk lipid utilization and absorption of dietary lipids in zebrafish larvae. Biochemistry 45, 15179–15187, https://doi.org/10.1021/bi0619268 (2006).
40. Le Guyader, D. et al. Origins and unconventional behavior of neutrophils in developing zebrafish. Blood 111, 132–141, https://doi. org/10.1182/blood-2007-06-095398 (2008).
41. Kao, T. T., Chang, W. N., Wu, H. L., Shi, G. Y. & Fu, T. F. Recombinant zebrafish {gamma}-glutamyl hydrolase exhibits properties and catalytic activities comparable with those of mammalian enzyme. Drug metabolism and disposition: the biological fate of chemicals
37, 302–309, https://doi.org/10.1124/dmd.108.024042 (2009).
42. Xiao, P. P., Hu, Y. H. & Sun, L. Scophthalmus maximus cystatin B enhances head kidney macrophage-mediated bacterial killing.
Developmental and comparative immunology 34, 1237–1241, https://doi.org/10.1016/j.dci.2010.07.008 (2010).
43. Wassler, M. et al. Identification and characterization of cystatin-related epididymal spermatogenic protein in human spermatozoa: localization in the equatorial segment. Biology of reproduction 67, 795–803, https://doi.org/10.1095/biolreprod.102.003970 (2002). 44. Johnson, S. R. Untangling the protease web in COPD: metalloproteinases in the silent zone. Thorax 71, 105–106, https://doi.
org/10.1136/thoraxjnl-2015-208204 (2016).
45. Shapiro, S. D., Campbell, E. J., Welgus, H. G. & Senior, R. M. Elastin degradation by mononuclear phagocytes. Annals of the New
York Academy of Sciences 624, 69–80, https://doi.org/10.1111/j.1749-6632.1991.tb17007.x (1991).
46. Serveau-Avesque, C. et al. Active cathepsins B, H, K, L and S in human inflammatory bronchoalveolar lavage fluids. Biology of the
cell / under the auspices of the European Cell Biology Organization 98, 15–22, https://doi.org/10.1042/BC20040512 (2006). 47. Taggart, C. C. et al. Cathepsin B, L, and S cleave and inactivate secretory leucoprotease inhibitor. Journal of biological chemistry 276,
33345–33352, https://doi.org/10.1074/jbc.M103220200 (2001).
48. Turk, V., Stoka, V. & Turk, D. Cystatins: biochemical and structural properties, and medical relevance. Frontiers in bioscience: a
journal and virtual library 13, 5406–5420, https://doi.org/10.2741/3089 (2008).
49. Maher, K. et al. A role for stefin B (cystatin B) in inflammation and endotoxemia. Journal of biological chemistry 289, 31736–31750,
https://doi.org/10.1074/jbc.M114.609396 (2014).
50. Jevnikar, Z. K. J. CSTB(cystatin B (Stefin B)). Atlas of genetics and cytogenetics in oncology and haematology 13, 3 (2009).
51. Lee, M. J. et al. Identification of cystatin B as a potential serum marker in hepatocellular carcinoma. Clinical cancer research: an
official journal of the American Association for Cancer Research 14, 1080–1089, https://doi.org/10.1158/1078-0432.CCR-07-1615
12
Scientific RepoRtS | (2019) 9:12633 | https://doi.org/10.1038/s41598-019-49152-7www.nature.com/scientificreports
www.nature.com/scientificreports/
52. Veeranki, S. P. et al. Maternal Folic Acid Supplementation During Pregnancy and Early Childhood Asthma. Epidemiology 26, 934–941, https://doi.org/10.1097/EDE.0000000000000380 (2015).
53. Tchetina, E. V., Demidova, N. V., Karateev, D. E. & Nasonov, E. L. Rheumatoid factor positivity is associated with increased joint destruction and upregulation of matrix metalloproteinase 9 and cathepsin k gene expression in the peripheral blood in rheumatoid arthritic patients treated with methotrexate. International journal of rheumatology 2013, 457876, https://doi. org/10.1155/2013/457876 (2013).
54. Kao, T. T. et al. Characterization and comparative studies of zebrafish and human recombinant dihydrofolate reductases–inhibition by folic acid and polyphenols. Drug metabolism and disposition: the biological fate of chemicals 36, 508–516, https://doi.org/10.1124/ dmd.107.019299 (2008).
55. Kao, T. T. et al. Grape seed extract inhibits the growth and pathogenicity of Staphylococcus aureus by interfering with dihydrofolate reductase activity and folate-mediated one-carbon metabolism. Internation journal of food microbiology 141, 17–27, S0168-1605(10)00256-4 [pii], https://doi.org/10.1016/j.ijfoodmicro.2010.04.025 (2010).
56. Matthews, R. G. & Daubner, S. C. Modulation of methylenetetrahydrofolate reductase activity by S-adenosylmethionine and by dihydrofolate and its polyglutamate analogues. Advances in enzyme regulation 20, 123–131, https://doi.org/10.1016/0065-2571(82)90012-7 (1982).
57. Kao, T. T. et al. Methotrexate-induced decrease in embryonic 5-methyl-tetrahydrofolate is irreversible with leucovorin supplementation. Zebrafish 10, 326–337, https://doi.org/10.1089/zeb.2013.0876 (2013).
Acknowledgements
We would like to thank Dr. Verne Schirch/Virginia Commonwealth University for valuable advice and assistance. We also thank Taiwan Zebrafish Core Facility at ZeTH (supported by the grant MOST 104-2319-B-400-001-) for the material and technical support. This study was supported by grants from the Ministry of Science and Technology, Taiwan (MOST 103-2321-B-006-027, 103-2321-B-006-011, 106-2311-B-006-004-MY3, 107-2320-B-037-017, and 108-2320-B-037-004) and Kaohsiung Medical University, Taiwan (KMU-M107002, NSYSUKMU107-P013, and NSYSUKMU108-P019). This work was supported by grants (NSC 101-2321-B-006-023, MOST 103-2321-B-006-027, MOST 103-2321-B-006-011, MOST 106-2311-B-006 -004 -MY3 and MOST 107-2320-B-037-017) from the Ministry of Science and Technology, Taiwan, and grants (KMU-M107002, NSYSUKMU107-P013, NSYSUKMU108-P019) from Kaohsiung Medical University, Kaohsiung, Taiwan.
Author Contributions
T.F. Fu and B.H. Chen conceptualized this study. T.F. Fu, N.W. Cheng and J.N. Tsai designed the experiments. G.H. Lee, N.W. Cheng and H.H. Yu performed the experiments. T.F. Fu, T. Liu, Z.H. Wen and B.H. Chen analyzed the data, discussed layout of the manuscript and wrote the manuscript. B.H. Chen and T.F. Fu are responsible for funding acquisition.
Additional Information
Supplementary information accompanies this paper at https://doi.org/10.1038/s41598-019-49152-7. Competing Interests: The authors declare no competing interests.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Cre-ative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not per-mitted by statutory regulation or exceeds the perper-mitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
1 September 2019 | Volume 10 | Article 1086
ORIGINAL RESEARCH
doi: 10.3389/fphar.2019.01086published: 20 September 2019
Frontiers in Pharmacology | www.frontiersin.org
Edited by:
Rodrigo Pacheco, Fundación Ciencia and Vida, Chile
Reviewed by:
Carmen G. Feijoo, Universidad Andrés Bello, Chile Martino Luigi Di Salvo, Sapienza University of Rome, Italy
*Correspondence: Martin K. Safo [email protected] Tzu-Fun Fu [email protected] Specialty section:
This article was submitted to Neuropharmacology, a section of the journal Frontiers in Pharmacology
Received: 23 May 2019 Accepted: 26 August 2019 Published: 20 September 2019 Citation:
Chen P-Y, Tu H-C, Schirch V, Safo MK and Fu T-F (2019) Pyridoxamine Supplementation Effectively Reverses the Abnormal Phenotypes of Zebrafish Larvae With PNPO Deficiency. Front. Pharmacol. 10:1086. doi: 10.3389/fphar.2019.01086
Pyridoxamine Supplementation
Effectively Reverses the Abnormal
Phenotypes of Zebrafish Larvae With
PNPO Deficiency
Po-Yuan Chen 1, Hung-Chi Tu 1, Verne Schirch 2, Martin K. Safo 2* and Tzu-Fun Fu 1,3*
1 College of Medicine, Institute of Basic Medical Science, National Cheng Kung University, Tainan, Taiwan, 2 Department of
Medicinal Chemistry and Institute for Structural Biology, Drug Discovery and Development, School of Pharmacy, Virginia Commonwealth University, Richmond, VA, United States, 3 Department of Medical Laboratory Science and Biotechnology,
College of Medicine, National Cheng Kung University, Tainan, Taiwan
Neonatal epileptic encephalopathy (NEE), as a result of pyridoxine 5
′-phosphate oxidase
(PNPO) deficiency, is a rare neural disorder characterized by intractable seizures and
usually leads to early infant death. The clinical phenotypes do not respond to antiepileptic
drugs but are alleviated in most cases by giving large doses of pyridoxal 5
′-phosphate
(PLP). PLP is the active form of vitamin B6 participating in more than 100 enzymatic
pathways. One of the causes of NEE is pathogenic mutations in the gene for human
PNPO (hPNPO). PNPO is a key enzyme in converting pyridoxine (PN), the common
dietary form of vitamin B6, and some other B6 vitamers to PLP. More than 25 different
mutations in hPNPO, which result in reduced catalytic activity, have been described for
PNPO-deficiency NEE. To date, no animal model is available to test new therapeutic
strategies. In this report, we describe using zebrafish with reduced activity of Pnpo as
an animal model. Knocking down zPnpo resulted in developmental anomalies including
brain malformation and impaired locomotor activity, similar to the clinical features of
PNPO-deficiency NEE. Other anomalies include a defective circulation system. These
anomalies were significantly alleviated by co-injecting either zpnpo or hPNPO mRNAs.
As expected from clinical observations in humans, supplementing with PLP improved the
morphological and behavioral anomalies. PN only showed marginal positive effects, and
only in a few anomalies. Remarkably, pyridoxamine (PM), another dietary form of vitamin
B6, showed rescue effects even at a lower concentration than PLP, presenting a possible
new therapeutic treatment for PNPO-deficiency NEE. Finally, GABA, a neurotransmitter
whose biosynthesis depends on a PLP-dependent enzyme, showed some positive
rescue effect. These results suggest zebrafish to be a promising PNPO-deficiency model
for studying PLP homeostasis and drug therapy in vivo.
Pyridoxamine Effectively Reverses PLP Deficiency Chen et al.
2 September 2019 | Volume 10 | Article 1086 Frontiers in Pharmacology | www.frontiersin.org
INTRODUCTION
A severe neurological problem that presents in prenatal, neonatal, or infant children is neonatal epileptic encephalopathy (NEE) as a result of pyridoxine 5′-phosphate oxidase (PNPO) deficiency (Mills et al., 2005; Hoffmann et al., 2007; Ormazabal et al., 2008;
Musayev et al., 2009; Balasubramaniam et al., 2010; Bowling, 2011; Ghatge et al., 2012; Mills et al., 2014; di Salvo et al., 2017a;
di Salvo et al., 2017b; Wilson et al., 2019). Most patients with this condition are born prematurely and may die if not treated (Mills et al., 2005; Hoffmann et al., 2007). Apart from the typical severe and intractable seizures (tonic, myoclonic, and clonic), other expressed symptoms may include fetal distress, lactic acidosis, repetitive facial movement, hypoglycemia, stunted growth, anemia, increased blood lactate, electroencephalogram with burst suppression pattern, and asphyxia (Mills et al., 2005;
Panayiotopoulos, 2005; Hoffmann et al., 2007; Ormazabal et al., 2008; Ghatge et al., 2012; Mills et al., 2014). The symptoms are in most part non-responsive to conventional antiepileptic drugs; however, surviving infants show improvement when treated with large and repeated doses (60–100 mg/kg/day) of PLP (Mills et al., 2005; Hoffmann et al., 2007; Ormazabal et al., 2008; Ghatge et al., 2012; Mills et al., 2014). In some instances, patients respond to pyridoxine (Mills et al., 2014; Plecko et al., 2014). Although PLP therapy has led to normal developmental outcomes in several patients, high doses of PLP are known to cause seizures (Ishioka et al., 1995; Hammen et al., 1998) and/ or severe liver damage (Mills et al., 2012). This toxic effect of PLP is likely due to the reactive aldehyde forming complexes (aldimines) with non-PLP proteins and interfering with their function (Bartzatt and Beckmann, 1994; Ishioka et al., 1995;
Salazar and Tapia, 2001; Scott et al., 2008). PLP is a cofactor for more than 100 vitamin B6 (PLP-dependent) enzymes that are involved in a large number of metabolic pathways, for example, amino acid metabolism, glucose metabolism, and heme and lipid syntheses (Percudani and Peracchi, 2009; di Salvo et al., 2011; Ghatge et al., 2012; di Salvo et al., 2017a). The function and development of the central nervous system also require enzymes that use PLP as a cofactor (Riggs et al., 1996; Musayev et al., 2009; Percudani and Peracchi, 2009; di Salvo et al., 2011;
Garcia-Cazorla A and Clayton, 2012; Ghatge et al., 2012; Mills et al., 2012; Mills et al., 2014; di Salvo et al., 2017a). For example, the synthesis of many neurotransmitters, for example, GABA, serotonin, melatonin, dopamine, epinephrine, norepinephrine, and histamine, to mention a few, are dependent on several PLP-dependent enzymes (Riggs et al., 1996). Deficiency of PLP in cells is, therefore, likely to impact the development of many organs.
One form of NEE (PNPO-deficiency NEE) is caused by mutations in the human PNPO (hPNPO) gene encoding hPNPO. At present, about 25 different mutations have been reported (Mills et al., 2005; Hoffmann et al., 2007; Ormazabal et al., 2008; Musayev et al., 2009; Balasubramaniam et al., 2010; Bowling, 2011; Ghatge et al., 2012; Mills et al., 2014;
di Salvo et al., 2017a; Wilson et al., 2019). Humans, unlike most prokaryotes, rely on a B6 salvage pathway that includes hPNPO, pyridoxal kinase, and PLP phosphatase to synthesize
and/or recycle PLP during protein turnover (Figure 1) (di Salvo et al., 2011; di Salvo et al., 2012; Ghatge et al., 2012). The primary B6 forms, pyridoxine (PN), pyridoxamine (PM), and pyridoxal (PL), are phosphorylated to pyridoxine 5′-phosphate (PNP), pyridoxamine 5′-phosphate (PMP), and pyridoxal 5′-phosphate (PLP), respectively, by pyridoxal kinase (Figure 1). PNP and PMP are converted to PLP by PNPO. PLP is converted to PL by phosphatases (Figure 1) during protein turnover, which is then reconverted to PLP by pyridoxal kinase (Figure 1). Another type of B6-dependent epilepsy is due to biallelic variants in the enzyme antiquitin (ALDH7A1) and is referred to as PN-dependent epilepsy (Pena et al., 2017; Coughlin et al., 2019). Antiquitin catalyzes oxidation of α-aminoadipic semialdehyde to α-aminoadipic acid. Its inactivation by mutations leads to a metabolic buildup of piperideine-6-carboxylic acid that reacts with and depletes PLP from the cell. PN-dependent NEE is characterized by mild to severe clinical symptoms and usually responds to treatment with pyridoxine and/or conventional antiepileptic drugs (Pena et al., 2017; Coughlin et al., 2019; Wilson et al., 2019). Recently, mutations in the gene encoding a PLP-binding protein that is potentially crucial for B6 homeostasis (PLPHP or PROSC) have also been shown to cause a novel form of B6-dependent epilepsy (Johnstone et al., 2019).
Development of new therapies and animal models to study these various forms of vitamin B6 pathophysiology is warranted. This report focuses on using zebrafish as an animal model for studying PNPO-deficiency NEE. Zebrafish has prominently served as a vertebrate model for research in disease mechanisms and drug discovery. It shares about 70% similarity with human genetic and molecular pathways (Howe et al., 2013). Their low cost, easy maintenance, amenable genetic manipulation, high productivity, fast maturation, and transparent embryos have made these animals suitable for developmental studies, for example, for epilepsy or seizures (Baraban et al., 2005;
Hortopan et al., 2010; Teng et al., 2011; Baraban et al., 2013;
Grone et al., 2016; Sourbron et al., 2016; Griffin et al., 2017;
Ibhazehiebo et al., 2018; O’Connor et al., 2019). Two major ways to induce seizure and/or other anomalies in zebrafish are genetic and pharmacologic (Baraban et al., 2005; Alfaro et al., 2011; Vermoesen et al., 2011; Pena et al., 2017; Vada et al., 2017; Yang et al., 2017). One genetic approach uses CRISPR/ Cas9 gene editing to generate an aldh7a1-null zebrafish model that mimics the clinical and biochemical features of PN-dependent NEE (Pena et al., 2017). A recent report also showed that zebrafish larvae with PLPHP knockout displayed phenotypic signs of seizure and are responsive to treatment with pyridoxine (Johnstone et al., 2019).
Pyridoxamine Effectively Reverses PLP Deficiency Chen et al.
3 September 2019 | Volume 10 | Article 1086 Frontiers in Pharmacology | www.frontiersin.org
MATERIALS AND METHODS
Fish (Danio rerio) Care and Maintenance
Zebrafish (Danio rerio, AB strain) were purchased from NTHU-NHRI Zebrafish Core Facility, Taiwan. They were bred and maintained at 28.5°C in a 10- to 14-h light–dark diurnal cycle following standard procedure (Westerfield, 2000). Both transgenic lines Tg(sox10:eGFP) and Tg(cmlc2:eGFP) were purchased from Taiwan Zebrafish Core Facility at ZeTH (support by MOST 105-2319-B-400-001). All usage and experiments, including adult and larval zebrafish, were approved by the Institutional Animal Care and Use Committee, National Cheng Kung University, Tainan, Taiwan (IACUC Approval No. 106086).
Knockdown and Rescue of zPnpo
Expression With MO and mRNA
Knockdown of zPnpo expression was accomplished by injecting embryos with the MO, specifically targeting the translation start site of zpnpo to block protein translation. The sequence of zebrafish Pnpo MO designed by the manufacturer (Gene-Tools, LLC, Philomath, OR) based on the zebrafish pnpo coding sequence (NM_001256178.1) is 5′-ACGTCTCATGCTTGTTCCGCG-3. A scrambled MO mixture containing 425 different nucleotide
sequences was used as a standard control MO. For microinjection, approximately 4.6 nL of solution containing 5 to 10 ng of zPnpo MO or 10 ng of scrambled MO was injected into embryos at the one- to two-cell stage. For rescuing with mRNA, MOs were co-injected with 800 pg of zpnpo or hPNPO mRNA. All reagents for microinjection were dissolved in degassed and RNase-free Danieu’s buffer to make proper stock solutions.
zPnpo Protein Expression
The expression of zPnpo was examined with Western blotting as previously described, with minor modifications (Kao et al., 2008). In brief, 30 embryos at 24 hpf were homogenized in 30 μL of E buffer (40 mM of Tris–HCl, 2 mM of EDTA, 2 mM of EGTA, 500 mM of NaCl, and 4 M of urea) containing 10 mM of phenylmethanesulfonyl fluoride (Sigma, 78840) and proteinase inhibitor cocktail (Sigma, catalog No. P8340). After centrifugation, 30 µg of total protein in the supernatant was subjected to Western blot analysis with rabbit anti-zPnpo antibodies (Genetex, customized).
Whole-Mount In Situ Hybridization (WISH)
WISH was performed following the standard protocol as previously described, with minor modifications (Tu et al., 2017). In brief, riboprobes against zebrafish pnpo transcripts (anti-sense) and template strand (sense) pax2.1 transcripts were synthesized in vitro from the plasmid containing zpnpo coding sequence linearized with NcoI (using SP6 polymerase) and NdeI (using T7 polymerase) and the plasmid containing
pax2.1 coding sequence linearized with NdeI (using T7
polymerase), respectively, in the presence of digoxigenin (DIG) using the DIG-labeled RNA kit (Roche, catalog no. 11175025910).
Cryosection and Hematoxylin and Eosin
(H&E) Staining
The preparation and H&E staining for zebrafish tissue cryosections were performed following the protocols in The Zebrafish Book (Westerfield, 2000).
Pyridoxamine Effectively Reverses PLP Deficiency Chen et al.
4 September 2019 | Volume 10 | Article 1086 Frontiers in Pharmacology | www.frontiersin.org
RNA Isolation and Reverse Transcription–
Polymerase Chain Reaction (RT-PCR)
Total RNA was extracted from 30–40 embryos at indicated stages as previously described (Kao et al., 2014). The zpnpo transcript was analyzed with RT-PCR with forward primer 5′-CAGCATCAAGCAGAGGGAG-3′ and reverse primer 5′-AACGCAGGACATTGAGGA-3′. Actin was used as an internal control with forward primer 5′-AGACATCAA GGAGAAGCTGTG-3′ and reverse primer 5′-TCCAGACGGA GTATTTAC-3′.
Compound Treatment
B6 vitamers, including PLP, PN, PM, and PL, were freshly prepared in embryo water and were added to embryo water at 1 hpf to reach the highest concentrations without inducing apparent harmful effects to embryonic development for rescue. The concentrations used were 1 mM for PN and 0.1 mM for PM and PL for all trials. The neurotransmiter GABA was added to embryos water at 6 hpf to reach the concentration of 0.5 mM. Considering the toxicity, stability, and light sensitivity, the PLP concentrations used were sequentially decreased until the indicated stages: 1 mM of PLP at 0–1 dpf, 0.5 mM at 1–2 dpf, and 0.1 mM at 2–4 dpf. Embryos were continuously exposed to the tested B6 vitamers with larval morphology and survival recorded at the indicated times. Embryo water was replaced daily with water containing freshly prepared B6 vitamers.
Cloning, Expression, and Purification of
Recombinant
zPnpo
The recombinant zPnpo was obtained following the protocol as previously described (Kao et al., 2008). The forward primer (5′-GGATCCAGACGTCTGTTAAGGTTTTG-3′) and reverse primer (5′-CAAGCTTTCAAGGGGACAAGCG-3′), with design based on the zpnpo coding sequence (NM_001256178), were used for PCR cloning of the zpnpo coding sequence from the zebrafish cDNA library. The 847-bp PCR product was cloned into the expression vector pET43.1a via BamHI and HindIII restriction enzyme sites. The recombinant zPnpo was overexpressed with a Nus-His tag at the N-terminus in
Escherichia coli strain HMS174(DE3) and purified with a
nickel Sepharose column. The purified recombinant zPnpo fusion protein was digested with thrombin to remove the Nus-His tag before being subjected to antibody production. The specificity of the antibodies was confirmed by comparing the Western blotting signals generated from the E. coli extracts with/without containing the induced recombinant zPnpo that was used for antibody generation (Suppl. S1).
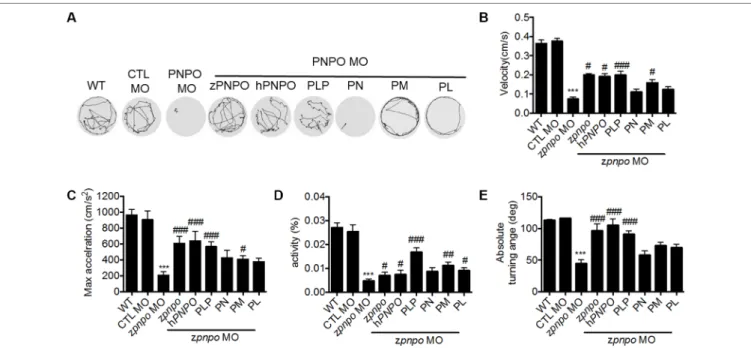
Spontaneous Movement
Larvae at 1 dpf were placed on a concave slide with embryo water under a transmitted-light stereomicroscope (Leica, MDG28) equipped with a digital single-lens reflex camera (Canon, EOS 550D). Larval spontaneous contraction was video-recorded in 60 frames per second and analyzed with EthoVision XT (Noldus, Wageningen, The Netherlands).
Larval Swimming Behavior Analysis
Larval swimming behavior was recorded and analyzed with a high-throughput image recording and analysis system from DanioVision™ (Noldus, Wageningen, The Netherlands). The
larva at 4 dpf was placed in a well on a 48-well plate. The plate was moved into the DanioVision™ device 15 min before analysis to
allow larvae to rest. Larval swimming behavior was video-tracked and recorded for 1 h at 9.94 frames per second. The velocity, maximum acceleration, relative turning angle, absolute turning angle, and body activity track of movement were analyzed with the built-in analytic software EthoVision XT.
Statistical Analysis
The statistical significance was calculated with one-way analysis of variance (ANOVA) (Kruskal–Wallis test) for behavior study and one-tailed Mann–Whitney nonparametric U test for phenotype analysis at 95% confidence intervals using the software GraphPad Prism 5.
RESULTS
Zebrafish Pnpo Displays Structural
Conservation During Evolution
Pnpo has been identified in many species, with zebrafish displaying conservation in both amino acid sequence and folding structure during evolution. To our knowledge, there is only one copy of zpnpo in the zebrafish genome, even though many genes in zebrafish exist with extra copies. The zpnpo gene contains seven exons located on chromosome 12 and encodes zPnpo, which has 267 amino acids (Figure 2A). The peptide sequence alignment and comparison show that zPnpo is 54% identical and 71% homologous to hPNPO (Figures 2A, B). In addition, the key residues in hPNPO for binding PNP/PLP (E77, W206, H227, K100, Y157, R161, R225, and S165) and FMN (Q174, Q139 W219, P261, S175, R116, R141, R95, W219, E217, K117, R138, and R229) are strictly conserved in zPnpo (Figure 2A). Our group had previously determined the crystal structure of hPNPO (Figure 2C) (Musayev et al., 2003). As expected from the high sequence similarity between hPNPO and zPnpo (Figures 2A, D, E), the two structures predicted by the online program I-TASSER (Iterative Threading ASSEmbly Refinement) showed almost identical predicted folds that are also close to the determined fold for hPNPO (Figure 2F) (Zhang, 2008; Roy et al., 2010; Yang et al., 2015). These data on the structural analysis of PNPO support the conservation among PNPO during evolution.