ON THE POSSIBLE FUNCTION OF COATED
VESICLES IN MELANOGENESIS OF
THE REGENERATING FOWL FEATHER
GERD G . MAUL and J . A . BRUMBAUGH
From the Department of Pathology, Temple University Health Sciences Center, Philadelphia, Pennsylvania 19140,and the Department of Zoology, University of Nebraska,
Lincoln, Nebraska68508
ABSTRACT
How tyrosinase becomes associated with the premelanosomes was investigated by histo-chemical demonstration of tyrosinase activity by the use of dihydroxyphenylalanine (DOPA) in melanocytes of regenerating fowl feathers . The reaction product of DOPA was localized in the anastomosing membrane tubules associated with the concave side of some dictyosomes of the Golgi apparatus and in coated vesicles most of which were in connection with the dictyosomes . No reaction product was found in early premelanosomes . In premelanosomes, the reaction product of DOPA appears first in vesicles approximately 400 A in diameter which are surrounded by a matrix with a characteristic periodicity . These observations seem to allow the speculation that the coated vesicles function in the transport of tyrosinase, and that the premelanosomes are formed in a process which is not necessarily dependent on the Golgi apparatus as was assumed earlier.
INTRODUCTION
The formation of the melanosome has been the subject of many investigations. (For reviews see 1, 5, and 20) . The presence of different stages of premelanosomes in the cytocentrum (20), coupled with biochemical results (22), suggested a direct involvement of the Golgi apparatus . It was hypothesized that tyrosinase is synthesized, like zymogen (8, 9), on the rough-surfaced endoplasmic reticulum (RER) and channeled through the intracisternal space to the Golgi apparatus . It is thought to be packaged there in small vesicles which enlarge by growth or fusion . Then the enzyme forms a complex matrix of helical fibers or folded sheets (2, 11, 22) . Oxidation and polym-erization of tyrosine onto this matrix would then complete the formation of the pigment granule . This hypothesis postulates tyrosinase to be present
THE JOURNAL OF CELL BIOLOGY • VOLUME 48, 1971 . pages4 1 -48
in an active or inactive form in the earliest pre-melanosomes.
A different sequence of events was suggested by Novikoff et al . (17), who found tyrosinase localized in Golgi-associated, tubular, smooth endoplasmic reticulum (SER) in mouse melanomas (17), and by Maul (13), who showed a direct membranous continuity between premelanosomes and tubular SER in human melanomas in vitro . In order to confirm these results in a normal system, we in-vestigated the melanocytes of the regenerating fowl feather after incubation in DOPA . The reac-tion product of tyrosinase and DOPA should reveal how tyrosinase becomes associated with the premelanosomes. Perotti (18) and Novikoff et al . (17) found tyrosinase activity also in Golgi-asso-ciated vesicles in the retinal epithelium of the
41
jcb.rupress.org
Downloaded from
embryonic rat and in mouse melanoma, respec- had been found to function in the transport of tively. Stanka et al. (23) were able to clearly show acid hydrolases (6) . In the present study, we tried that these vesicles were of the coated variety in to elucidate the transport of tyrosinase to the hamster melanoma in vitro . The coated vesicles melanosome in the normal melanocyte .
FIGURE 1 Survey micrograph of the Golgi region. The center contains tubular SER filled with the reaction product of tyrosinase and DOPA. In the right upper corner, there is a premelanosome (pm,) without reaction product . The premelanosome underneath (pmt)is partially filled with reaction product . Arrows point out coated vesicles (cv) which are connected to SER . Scale marker, 2000 A . X 50,000.
42
THE JOURNAL OF CELL BIOLOGY . VOLUME 48, 1971
jcb.rupress.org
Downloaded from
MATERIALS AND METHODS
Several black regenerating breast feathers from an adult fowl were plucked, trimmed, and split longi-tudinally. They were fixed in 3% glutaraldehyde in 0 .1Mphosphate buffer for 2 hr . Some of the specimens were then incubated in freshly prepared 5 mm dihy-droxyphenylalanine (DOPA, Mann Research Labo-ratories) in 0 .1 Mphosphate buffer (pH 6 .9) for 3 hr at 38°C (14, 15, 17, 18) . Control and incubated specimens were osmicated (2% OsO4) in 0 .1 M phos-phate buffer for
l3j
hr, block-stained with2% ura-nyl acetate in water (10), dehydrated, and embedded in Epon 812 (12) . Ultrathin sections were picked up on single hole grids coated with 1 % collodion in amylacetate and observed with a Siemens 101 electron microscope at 80 kv, unstained or stained with lead citrate (19) .OBSERVATIONS
Melanocyte differentiation in the regenerating fowl feather takes place in the collar and proximal barb- ridges . Melanocytes observed here were found where the collar had developed into dis-tinct barb ridges. The melanocytes were filled with melanosomes of different degrees of maturity . The reaction product of tyrosinase and DOPA was localized in the tubular membranes often associated with the dictyosomes at their concave surfaces (Fig. 1) . Surprisingly, there was no tyrosinase activity in early premelanosomes. These organelles were membrane delimited and con-tained numerous vesicles approximately 400 A in diameter (Fig . 1, pm l , and Fig . 2) . The vesicles are surrounded by a fibrous matrix which shows a distinct periodicity. A shorter, wider type of premelanosome was also observed (Fig. 1, pm 2) . The appearance and distribution of these different types of melanosomes will be discussed elsewhere .
The highest concentration of reaction product, as judged by electron opacity, was found in the coated vesicles . This can be seen in Fig . 3 as well as in the lower left corner of Fig . 1 where the arrows point to coated vesicles showing attachment to the tubular SER . However, the reaction prod-uct was also present at high concentrations in the internal vesicles of certain melanosomes (Fig. 4) . As already observed by Dürrer and Villiger (4) and by Stanka et al . (23), there were hardly any intermediate stages present . Wise (25) had found an electron opaque material within the fenes-trated cisternae of the Golgi apparatus . Such material was also found in our preparation, but it was never found in cisternae which contained DOPA reaction product (Fig . 5) . It is, therefore, assumed not to consist of tyrosinase-dependent formation of melanoprotein (25) . Tyrosinase activity, as judged by the presence of DOPA reaction product, was usually found in the anas-tomosing network of membranous tubules associ-ated with the concave face of the dictyosome and also in SER which could not clearly be related to a dictyosome. A face-on view of such a network is shown in Fig . 6 . Serial sections showed that these anastomosing tubules are not part of the fenes-trated cisternae of the dictyosome . No reaction product can be seen in the fenestrated cisternae pictured in Fig. 7 a. The tubular membranes in Fig . 7 b were separated from the fenestrated cisternae by three sections (approximately 2500 A) which included oblique views of the dictyosome. Thus, the two membrane systems are situated at opposite poles of this organelle .
A number of images were obtained which suggested a continuity of tyrosinase-containing membrane tubules with premelanosomes .
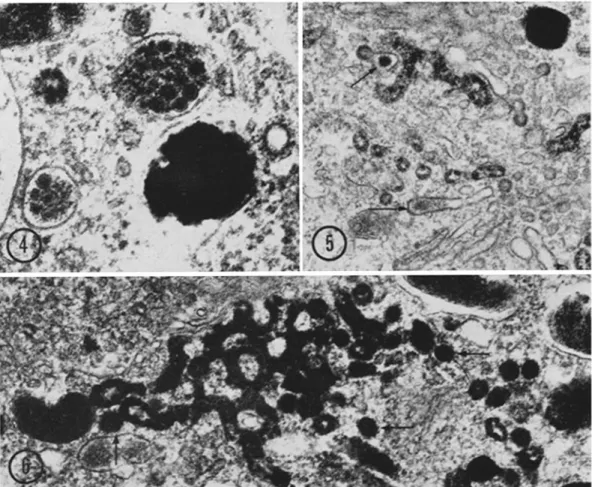
How-FIGURE 2 Longitudinal section of a premelanosome of the same cell as Fig . 1 . The premelanosome consists of a fibrous matrix with distinct periodicity . This matrix surrounds many densely packed vesicles some of which show reaction product (arrows) . Scale marker, 2000 A . X 80,000 .
FIGURE S Two coated vesicles and smooth-surfaced vesicle or cross-sectioned membrane tubule con-taining reaction product. X 86,000.
G. G . MAUL AND J . A . BRUMBAUGH Coated Vesicles in Melanogenesis
43
jcb.rupress.org
Downloaded from
FIGURE 4 Oblique sections through premelanosomes with advanced melanization . The internal small
vesicles are all filled with reaction product . Scale marker, 1000 A. X 100,000 .
FIGURE 5 Fibrous material localized in dictyosomal cisternae and associated membrane elements (arrows) . This material was not found in membrane tubules connected to reaction product-containing membrane elements . Scale marker,2000 A . X 50,000.
FIGURE 6 Tangential view of the anastomosing network of SER containing reaction product . It is situated parallel to the dictyosome . Coated vesicles are pointed out by arrows . Scale marker, 2000 A .
X 50,000 .
ever, in most cases one could not unequivocally demonstrate the uninterrupted nature of the unit membrane . Continuous serial sections of the lateral edge of a dictyosome are shown in Fig . 8 . The continuity of the unit membrane of a tubule with the obliquely sectioned premelanosome can be recognized (Figs. 8 d, e), but it is not clear whether the membrane tubule loops back into the organelle or passes "underneath ." In Figs. 8 aand b, a cross-sectioned premelanosome appears connected with tubular SE.R . Also, a direct con-nection of reaction product-containing SER with
44
THE JOURNAL OP CELL BIOLOGY . VOLUME 48, 1971
a membrane-delineated space comparable in diameter to a cross-sectioned premelanosome is shown in Figs. 8 a and b, but it does not contain the structural characteristics of a premelanosome . Coated vesicles in continuity with tubular SER contain reaction product or appear empty . An attempt was made to determine the ratio between reaction product-containing coated vesicles which are free in the cytoplasm and those which are connected to SER . The majority of coated vesicles seem to be connected with SER as judged from serial sections . The coated vesicles have a
diam-jcb.rupress.org
Downloaded from
FIGURE 7 Illustration of the absence of tyrosinase activity in the fenestrated cisterns at the outer
face of the dictyosome (a), but its presence in the tubular membranes associated with the dictyosome on its inner face (b) . The two micrographs show the same area separated by approximately 2500 A . Arrows point to structures which extend through depth of all serial sections . Scale marker, 1µ . X 32,000 . eter of approximately 700 A, excluding the
coat, and can, therefore, be contained in a single section . Only a few unattached coated vesicles containing reaction product could be demon-strated unequivocally. The findings, that a re-action product-containing tubule is in direct membranous continuity with a premelanosome as seen in Fig . 8a(arrow pointing right) might be interpreted as the fusion of a coated vesicle which was still attached to the Golgi-associated brane system .
The observation tubules or double
of mature melanosomes with membrane sheets filled with DOPA reaction product which loop back into the melanosome (Fig . 9) casts some doubt on the observation that the membrane tubule in Figs . 8 d and e (arrows) connects to the Golgi-associated membrane system .
DISCUSSION
The concept that melanosomes are developed from Golgi-derived vesicles (1, 2, 4, 5, 21, 22) was first questioned when tyrosinase activity was found in Golgi-associated SER (GERL, 17) and when membrane continuity between SER and premelanosomes was observed (13) . It was, therefore, postulated that premelanosomes develop as focal dilatations of SER .
mem-Tyrosinase was thought to contribute struc-turally as well as enzymatically to the melanosome so as to form the matrix onto which tyrosine after oxidation becomes polymerized (2, 11) . The dem-onstration that active tyrosinase is present at ribosomes (21, 22), in Golgi-associated SER and coated vesicles (see also reference 23), but not in early premelanosomes, as judged by the DOPA reaction, makes it seem unlikely that the char-acteristic matrix consisting of helical fibers or folded sheets is composed of aggregated tyrosinase molecules or that they contain active tryosinase as suggested by Novikoff et al . (17) ; but inhibited tyrosinase in early melanosomes cannot be ruled out .
The results of this investigation on normal melanocytes as well as similar observations on melanoma cells of mouse and hamster might allow the following speculation . The premelanosome with its complex matrix is not formed by enlarging vesicles of the Golgi apparatus, but rather within SER or RER as evidenced by the membranous connection to tubular membranes, especially in very early premelanosomes (13) . Tyrosinase is accumulated in Golgi-associated SER (GERL, 17) and then condensed in coated vesicles . Upon contact with a premelanosome, the coated vesicle releases its tyrosinase into the
premelan-G . premelan-G . MAUL AND J. A . BRUMBAUGH Coated Vesicles in Melanogenesis
4 5
jcb.rupress.org
Downloaded from
FIGURE 8 Consecutive serial sections show the direct continuity of premelanosome with membrane
tubules (arrows) containing reaction product . Coated vesicles (cv) with reaction product and "empty" ones are found close together . Scale marker, 2000 A. X 58,000 .
jcb.rupress.org
Downloaded from
FIGURE 9 Melanosomes with membrane exten-sions which are filled with reaction product but loop back into the organelle . Scale marker, 2000 A. X 58,000.
osome. The transport of tyrosinase by coated vesicles to premelanosomes seems to explain the presence of premelanosomes without tyrosinase activity, but is not proof of it since tyrosinase may be inhibited in the premelanosome .
Most startling was the observation that no tyrosinase activity was localized in early premelan-osomes ; i .e ., the ordered matrix, the small vesicle, and the space surrounded by the delimiting membrane were free of reaction product. If there was any reaction product present, it could be found in the small vesicles first, often only in one . There were very few intermediate stages in melanosome formation in our material ; an observation which was also reported by Stanka et al . (23) for hamster melanoma cells in vitro and by Durrer and Villiger (4) for the iridescent melanosome ofLamprotornis, a starling . Sometimes the center of large premelanosomes showed a dense
core of reaction product as in Fig . I (pm2), but none between the limiting membrane and the internal melanosomal structures . This is also true for the oblique sections of premelanosomes which contain reaction product in all small vesicles as well as on the matrix (Fig . 4) . The small vesicles were also observed in the melanosomes of hair (24) and in human melanoma cells in vitro (13) where they seem to be "empty" or appear nega-tively stained within the completely melanized melanosome . The significance of these small vesicles for melanogenesis is not clear . Durrer and Villiger (4) believe that they form the internal space for the so-called "gas filled" melanosomes of Lamprotornis . However, melanosomes with an "empty" center are also observed in our material (Fig. 8), making it seem possible that the small vesicles eventually break down . Despite an intensive search, no image was found which would indicate vesicle formation into premelanosomes . The absence of DOPA reaction product in small vesicles of early premelanosomes and its presence in more mature forms could be interpreted to mean that tyrosinase is absent or that it is inhib-ited . In order to reconcile the observation that coated vesicles are filled with reaction product of DOPA indicating active tyrosinase, and the appearance of DOPA reaction product first in the small vesicles of the premelanosomes, one may have to assume an inhibitor of tyrosinase to be present in the premelanosome . How this assumed inhibition of tyrosinase is released cannot be imagined at the present, but the con-tents of the small vesicles may eventually provide some clues.
The speculation that coated vesicles transport tyrosinase into melanosomes is based on the assumption that the dense reaction product is the result of the tyrosinase activity on DOPA, and on the hypothesis that coated vesicles transfer hydrolytic enzymes (6) . Nowhere in the literature nor in our material could we find unequivocal evidence for a coated vesicle's fusing with its receptor organelle or the plasma membrane, but it is well documented that vesicles forming at the plasma membrane for protein uptake or budding from Golgi membrane form a "coat ."
The function of coated vesicles in protein uptake has been established in various systems (summa-rized in reference 6) . Other investigators (3, 7, 16) postulated that coated vesicles function in the transport of soluble products of intracellular
G . G . MAUL AND J. A . BRUMBAUGH Coated Vesicles in Melanogenesis
4,
jcb.rupress.org
Downloaded from
digestion . Friend and Farquhar (6) could separate the two functions by a combined statistical and histochemical approach. The small coated vesicles function in the transport of hydrolases and were assumed to be primary lysosomes . Images were presented which showed fusion of large "coated" vesicles (heterophagosomes) filled with externally applied peroxidase with multivesicular bodies, but the coat of these heterophagosomes could not be discerned. This did not seem to be caused by problems of fixation since other coated heter-ophagosomes were present in the same image. It might be, therefore, that the coat is lost at the time of contact and membrane fusion . This may be the principal difficulty in proving that "coated" vesicles fuse with other organelles .
Friend and Farquhar emphasized the possibility that the transport functions of coated vesicles are multiple (6) . Coated vesicles devoid of DOPA reaction product but in the immediate vicinity of coated vesicles with reaction product and of the same diameter may possibly function in the transport of other enzymes . Also, tyrosinase seems to be the first enzyme demonstrated in Golgi-derived coated vesicles which does not belong to the intracellular digestive system. If the coated vesicles have transport function, which seems established for acid phosphatase and protein uptake and is likely for tyrosinase (see also ref-erence 23), one might infer a mechanism of selective fusing with the correct receptor organelle (multivesicular body or premelanosome) . This mechanism may reside in the coat.
This research was supported by grant CA-11654 from the National Institutes of Health and by a Public Health Research Career Development Award (1 K04 GM42355-01, Dr. Brumbaugh) from the National Institutes of General Medical Sciences . Received for publication 24 March 1970, and in revised form 20 July 1970.
REFERENCES
1 . BILLINGHAM, R . E ., and W . K . SILVERS . 1960 . Quart. Rev. Biol. 35 :1 .
48
THE JOURNAL OF CELL BIOLOGY . VOLUME 48, 1971
2 . BIRBECK, M . S . 1963 . Ann . N. Y . Acad. Sci. 100 : 540 .
3 . BRUNI, C ., and K . R. PORTER . 1965 . Amer . J. Pathol. 46 :691 .
4. DÜRRER, H ., and W . VILLIGER . 1967. Z. Zell forsch . Mikrosk . Anat. 81 :445 .
5 . FITZPATRICK, T . B ., M . MIYAMOTO, and K . ISHIKAwA . 1967 . Arch . Dermatol . 96 :305 . 6 . FRIEND, D . S ., and M . G. FARQUHAR . 1967 . J.
Cell Biol. 35 :357.
7 . HOLTZMAN, E., A. B. NOVIKOFF, and H . VILLA-VERDE . 1967 . J. Cell Biol. 33 :419 .
8 . JAMIESON, J . D ., and G. E . PALADE . 1968 . J. Cell Biol. 34 :577 .
9 . JAMIESON, J . D., and G. E . PALADE . 1968 . J. Cell Biol . 34 :597 .
10 . KELLENBERGER, E ., A . RYTER, and Y . SÉCHAUD . 1959 . J. Biophys. Biochem . Cytol . 4 :671 . 11 . KOECKE, H . W. 1966, In Beiträge zur Licht- und
Electronenmikroskopie . Carl Zeiss, Ober-kochen, Germany.
12 . LUFT, J . H . 1961 . J. Biophys . Biochem . Cytol. 9 : 409 .
13 . MAUL, G. G . 1969 . J. Ultrastruct. Res . 26 :163 . 14 . MISHIMA, Y . 1962 . J. Invest . Dermatol. 39 :369 . 15 . MISHMA, Y. 1964. J. Histochem . Cytochem . 12 :184. 16 . NOVIKOFF, A . B., P. S. ROHEIM, and N. QUINTANA.
1966 . Lab . Invest. 15 :27 .
17. NOVIKOFF, A. B ., A . ALBALA, and L . BIEMPICA . 1968 . J. Histochem . Cytochem . 16 :299 .
18 . PEROTTI, M . E . 1965 . Proceedings del V . Congresso Italiano di Microscopia Elletronica Bologna . Tipografia del Seminario di Tadova,
Italy 37.
19 . REYNOLDS, E . A. 1963 . J. Cell Biol. 17 :208 . 20 . SEIJI, M . 1967 . In Ultrastructure of Normal and
Abnormal Skin . Alvin Zelickson, editor . Lea Febiger, Philadelphia, Pa .
21 . SEIJI, M ., and S . IwASHITA. 1963 . J. Biochem . 54 :103 .
22 . SEIN, M ., K . SHIMAO, M . S . C. BIRBECK, and T . B. FITZPATRICK . 1963 . Ann . N. Y . Acad. Sci. 100 :497.
23 . STANKA, P ., V. KINZEL, and U. MOHR. 1969. Virchows Arch . Abt . B . Zellpathol. 2 :91 . 24 . TousIMIS, A . J . 1963. Ann. N. Y. Acad. Sci . 100 :
447.
25. WISE, G . E . 1969 . J. Ultrastruct. Res . 27 :472.