行政院國家科學委員會專題研究計畫 成果報告
Transglutaminase 於培養細胞對高溫及氧化壓力反應所扮
演之角色
研究成果報告(精簡版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 95-2311-B-002-020-
執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日
執 行 單 位 : 國立臺灣大學生化科學研究所
計 畫 主 持 人 : 張震東
計畫參與人員: 此計畫無參與人員:無
處 理 方 式 : 本計畫可公開查詢
中 華 民 國 96 年 12 月 05 日
行政院國家科學委員會補助專題研究計畫成果報告 Transglutaminase於培養細胞對高溫及氧化壓力反應所扮演之角色 計畫類別:X 個別型計畫 □整合型計畫 計畫編號:
: NSC 952311B002 020
-執行期間: 95年 08月 01日至 96年 07月 31日 計畫主持人:張震東 共同主持人: 本成果報告包括以下應繳交之附件: □赴國外出差或研習心得報告一份 □赴大陸地區出差或研習心得報告一份 □出席國際學術會議心得報告及發表之論文各一份 □國際合作研究計畫國外研究報告書一份 執行單位:國立台灣大學生命科學院生化科學研究所 中 華 民 國 96 年 10 月 22 日行政院國家科學委員會專題研究計畫成果報告 國科會專題研究計畫成果報告撰寫格式說明
Preparation of NSC Project Reports
計畫編號:
NSC 952311B002 020
-執行期限:95年8月1日至96年7月31日主持人:張震東 國立台灣大學生命科學院生化科學研究所 一、中文摘要
轉穀胺醯氨酶摧化之反應基本上是以酵素Cys 的thiol group 去攻打Gln 上的carbonyl group,NH3離開,形成Thioester bond。而此Thioester bond 大多是接受Lys 的ε-amino group 攻擊形成Isopeptide bond 而酵素離開。此Isopeptide bond 亦可接受另一個一 級氨、水分子或者醇類的攻擊。因此此酵素作用的反應為Transadmidation 造成蛋白
質之聚合、Amine incorporation 造成polyamines共價結合上蛋白質、Esterification 造成Gln 轉換 成酸酯、Deamidation 造成Gln 轉換成Glu、及Isopeptide cleavage 造成聚合蛋白質的分開。其主要 功能在於凝血、瘍口癒口、調控細胞自戕死亡、神導傳導物質釋放、訊息傳導及細胞間質形成等。轉穀 胺醯氨酶之功能必需透過受質之鑑定以及Transamidation 造成之影響オ得以瞭解。 本實驗室以新穎方法來純化及鑑定轉穀胺醯氨酶之受質,小鼠(>10 周齡)之肝臟及睪丸其轉穀胺醯氨酶 酵素活性,並純化出受質,通過質譜分析鑑定受質的身分,最後以免疫轉漬法以確認受質之真正身分。 在超過一白個鑑定受質中,大概屬下列性質蛋白;細胞骨架及其調節蛋白、Chaperones 及 Co-chaperones、內質網蛋白、細胞Detoxification用蛋白、蛋白質轉譯調節蛋白等,大多是與細胞壓 カ反應有關之蛋白。更重要的是其中多個受質屬新的發現。由於轉穀胺醯氨酶本身是氧化壓力所活化的 酵素,而且多數受質屬壓力反應有關之蛋白,因此我們認為轉穀胺醯氨酶及其受質参與細胞壓力反 應,其扮演之角色則是本計畫探討的主題。 關鍵詞:轉穀胺醯氨酶、質譜分析、受質 Abstract
Transglutaminases (TG) are Ca2+-dependent enzymes which catalyze a post-translational modification of
proteins. The enzyme reaction leads to the formation of an isopeptide bond either within or between polypeptide chains. The γ-glutamyl-ε-lysine crosslinks are formed between the γ-carboxamide group of peptide-bound glutamine residues and the ε-amino group of peptide-bound lysine residues. Polyamines can replace lysine residue in the transamidation reaction in vitro and in vivo. Transglutaminases are abundant enzymes which are involved in a number of different physiologic processes, e.g., plasma transglutaminase factor XIII stabilizes the fibrin clot during hemostasis, and keratinocyte and epidermal transglutaminase contribute to the formation of the cornified envelope in skin. Several physiological roles for tissue transglutaminases have been demonstrated, such as wound healing, fibrosis, apoptosis, and matrix formation.
In order to understand the physiological functions of tissue transglutaminase (tTG or TG2), one needs to identify the acyl donor and the acyl acceptor substrates in the transamidation reaction. To this end, we have identified 29 potential transglutaminase substrates from mouse liver extract by substrate purification and tandem Mass Spectrometry analysis. Interestingly, chaperones and co-chaperones are the most abundant tTG substrates found. Other transglutaminase substrates include intermediate filament protein, β-actin, proteasome proteins, peroxiredoxin, 14-3-3 proteins, valosin-containing protein, nucleolin, glyoxalase 1. Many of the tTG substrates are cellular proteins with functions related to cellular response to stress. More importantly, some of the tTG substrates have not been reported. In light of the fact that oxidative stress or UV irradiation elevates in situ tTG activity, we seek to study the role of tTG in HepG2 cells in response to heat shock and oxidative stress. More specifically, we will determine whether tTG is up-regulated or activated in HepG2 cells in response to heat shock and oxidative stress and further to determine the consequences of tTG activation. In addition, we will examine whether the tTG substrates that we have identified in mouse liver extract are indeed modified in vivo in HepG2 cells. In the end, we wish to know the roles of tTG and heat shock proteins and their interplay in HepG2 cells in response to heat shock and oxidative stress.
Keywords: transglutaminase、substrates、proteomics、Mass spectrometry 二、緣由與目的
轉穀胺醯氨酶家族有三組成員:Papain-like transglutaminases、protein disulfide isomerase-like transglutaminases及bacterial toxin transglutaminases。Papain-like transglutaminases屬於Cysteine protease
superfamily的一員,其它次家族成員有papain、calpain、foot and mouth virus protease、deubiquitylating enzymes、及N-acetyltransferases (Laszlo and Graham, 2003)。這些酵素具有與摧化反應相關的結構如 Cys-His-Asn或Cys-His-Glu,稱catalytic triad。
人類全基因含九個transglutaminase基因,其中有一個是erythrocyte band4.2巳不具摧化能力,其詳細 特性如表一所列。巳知的蛋白質均無醣化修飾,也缺乏雙硫鍵,摧化機制需要鈣離子(mM)的存在。Type 2 transglutaminase(TG2)是第一個被發現的成員,幾乎表現在所有的器官,也表現在細胞的不同部位如 cytosol (80%)、plasma membrane (含extracellular matrix,10-15%)、nuclear membrane (5%)。雖然一級結構 缺乏Signal peptide,TG2卻可以分泌到細胞表面及細胞間質上。最奇特的是它又是個GTP-binding protein, 結合GTP會抑制transglutaminase酵素活性,但是它又具GTPase活性,GTP終究水解為GDP。不過若TG2 先結合了鈣離子,鈣離子可阻斷GTP的結合。因此TG2與高分子量G protein (Ghα)是同一個蛋白質,可媒 介αadrenergic receptor活化所造成之phospholipase C的活化(Nakaoka et al., 1994)。
轉穀胺醯氨酶之受質鑑定有一種較傳統的方法,是把純化蛋白加入酵素看蛋白質是否聚合,或再加 入5-(biotinamido)pentylamine為探針去標示蛋白質,不過這種作法之生理意義較受質疑,實驗靠內生性酵 素作反應較有意義。轉穀胺醯氨酶之受質鑑定有二個對象;一稱Acyl donor指提供Gln參與反應的蛋白 質,另一稱Acyl acceptor指提供Lys參與反應的蛋白質,此二對象一般是二個不同之蛋白質亦可以是同一 個蛋白質,形成Intramolecular isopeptide bond,或是Heterodimer。所以在受質鑑定就有二群對象;以 5-(biotinamido)pentylamine為探針(或3[H]-putrescine),經摧化反應後可標示到Acyl donor,此標示之蛋白
質可轉漬至PVDF後以Streptavidin-peroxidase呈色(或以X光片)顯示,或經Streptavidin-agarose親和力管柱 純化。至於Acyl acceptor之鑑定文獻上則有不同的biotinylated peptide的設計,其中序列中含Gln氨基酸; 如LGLGQGKVLG (Gorman and Folk, 1984)、TVQQEL (Ruoppolo et al., 2003)、GQQQLG (Hu and Messersmith, 2003)、QQIV (Lorand et al., 1992)等。Pastor等(1999)曾對此設計作過詳細探討,結論如下: 1. Gln不可在N-或C-terminus、2. Gln不可位在二個鹼性胺基酸中間或Pro中間。在文獻上報導的Acyl donor 較多,可能是因為5-(biotinamido)pentylamine及3[H]-putrescine較易取得,同時也不必作設計。針對這問題, 作者巳設計合成了一個新的探計Biotin-RSGQQQLGSS,反應極佳而且水溶解度非常好,因此本計畫將以 此胜肽及5-(biotinamido)pentylamine為探針,尋找及鑑定轉穀胺醯氨酶之受質。 在本計畫中,我們利用小鼠的睪丸為材料,以蛋白質體學的方法去純化以及鑑定小鼠睪丸內TG的受 質,希望在一個像testis這樣持續進行細胞生長與分化的組織裡,了解TG所扮演的角色為何。經由純化 及MS/MS鑑定過後,我們發現了數十種蛋白質在小鼠睪丸內可能為TG的受質,在經由重組TG受質蛋白的 in vitro transamidation反應以及TG substrates pull-down的immunoblotting assay,我們也證實了 這些蛋白在in vitro的確為TG的受質。未來我們希望從中選取了幾個可能為TG受質的蛋白,觀察其TG進 行transamidation過後,會對這些蛋白的酵素活性、protein-protein interaction或是protein translocation會有怎樣的影響,進而能更加了解TG在testis內的角色與功能。
三、材料與方法 蛋白質膠體電泳
電泳配方及條件參考Schägger & Jagow (1987)所著Tricine-SDS方法,Running gel之濃度為7.5%, stacking gel濃度為4%,Cathode buffer組成是0.1 M Tris、0.1M Tricine、0.1% SDS,pH 8.25。Anode buffer 組成是0.2 M Tris, pH 8.9。Gel buffer組成是1 M Tris、0.1% SDS,pH 8.45。電泳條件是五十伏特二十分鐘 及一百伏特七十分鐘。之後膠體以水洗六十分鐘後以Colloidal Coomassie blue G-250 染色六十分鐘,再 以水洗淨二十分鐘。銀染色則採取Amersham-Pharmacia Plus One Silver Stain Kit﹐並依原廠商提供步驟進 行染色﹐但是退染改為5% 醋酸﹒
近免疫轉漬法 (Near-immunoblotting)
將欲分析之材料進行膠體電泳分析,利用Diffusion blotting (Chen and Chang, 2001)將蛋白質轉 印至PVDF濾紙上。以Phosphate buffered saline (PBS)洗滌十分鐘,加入Blocking solution (5% skim milk in PBS, 0.05% Tween 20)室溫反應一小時,再以PBS洗滌三次。接著加入Rabbit IgG-streptavidin complex (1 ug/ul於PBS,含0.05% Tween 20、3 mg/ml BSA)於室溫反應一小時,再以PBS-T洗滌三次,然後加入 Peroxidae-conjugated anti-rabbit IgG抗體(溶於PBS含0.05% Tween、3 mg/l BSA)反應於室溫一小時,再以 PBS洗滌三次。最後以DAB (0.6 mg/ml)、0.01% 氯化鎳、1 ul/ml 雙氧水呈色。
轉穀胺醯氨酶摧化反應
我們取成鼠Testis以10重量體積之20 mM Tris, pH 8, and 2% Triton X-100溶液研磨後,以20,000 xg離 心三十分鐘分成分上清液及不溶物二部分。不溶物再以20 mM Tris, pH8, and 0.15M NaCl (TBS)清洗一 次。轉穀胺醯氨酶摧化反應含0.2 ug/ul 5-(biotinamido)pentylamine (bPA) 或 biotin-RSGQQQLGSS (bPQ) 以及5 mM CaCl2, and 0.1 mM DTT及適量之組織材料。反應於室溫進行三十分鐘,有時加入20 mM of
Urea到8M濃度,混合十分鐘後離心取得可溶蛋白。 轉穀胺醯氨酶受質純化
因為要作純化所以上清液中止反應改成加入Urea到8M、DTT到50 mM,於室溫一小時後加入 Iodoacetamide到100 mM,再置於室溫一小時。之後材料以Amersham Biosciences出產之2-D clean-up kit 作蛋白質沈澱,後溶於8M urea/TBS中,與等體積之Streptavidin-argarose beads (以8 M urea/TBS平衡)結 合,後以五倍體積8 M urea/1.0 M NaCl/TBS, 0.1% SDS/TBS, 及20mM Tris buffer清洗Streptavidin-agarose beads各一次,最後以二倍體積4% SDS並加熱將受質沖洗出來。
西方墨點法(Western blotting)
將欲分析之材料進行膠體電泳分析,利用Semi-dry blotter (Hoeffer Semi-Phor)將蛋白質轉印至 PVDF濾紙上。以Phosphate buffered saline (PBS)洗滌十分鐘,加入Blocking solution (5% skim milk in PBS) 室溫反應一小時,再以PBS洗滌三次。接著加入抗血清(一比一千稀釋於PBS,含1 mM EDTA、3 mg/ml BSA)於攝氏四度反應十六小時,再以PBS洗滌三次,然後加入Peroxidase-conjugated anti-guinea pig IgG 抗體(溶於PBS含1 mM EDTA、3 mg/l BSA)反應於室溫二小時,再以PBS洗滌三次。最後以DAB (0.6 mg/ml)、0.01% 氯化鎳、1 ul/ml 雙氧水呈色。
質譜分析 (Mass spectrophotometry analysis and protein in-gel digestion)
將純化得到之蛋白質作SDS電泳,以Coomassie blue染色,待腿色後,以刀片將色帶切下,放入微 量管中,加入100 µl之DTT/25 mM碳酸氫氨。於37ºC反應一小時,加入100 µl Acetonitrile,振盪數分鐘, 離心除去上清液。加入100 µl 65 mM Iodoacetamide/25 mM 碳酸氫氨,於室溫反應一小時,加入100 µl Acetonitrile,振盪數分鐘,離心除去上清液。加入200 µl 50% Acetonitrile/25 mM 碳酸氨,振盪數分鐘, 離心除去上清液。加入200 µl 100% Acetonitrile,振盪數分鐘,離心除去上清液。加入10-15 µl trypsin 溶 液(0.1-0.15 µg配於25 mM碳酸氨),於37ºC反應16小時,加入10 µl 100% Acetinitrile,振盪數分鐘,離心 收集上清液。殘餘膠體加入20 µl 100% Acetonitrile/0.1% TFA萃取,振盪數分鐘,離心收集上清液。殘 餘膠體加入20 µl 50% Acetonitrile/0.1% TFA萃取,振盪數分鐘,離心收集上清液。殘餘膠體加入20 µl 100% Acetonitrile/0.1% TFA萃取,振盪數分鐘,離心收集上清液。把各次收集之上清液混合,並以 SpeedVac抽至殘留體積約10 µl,送中央研究院基因體/蛋白質體中心,以LC-Mass-Mass作蛋白質定序。 四、結果與討論
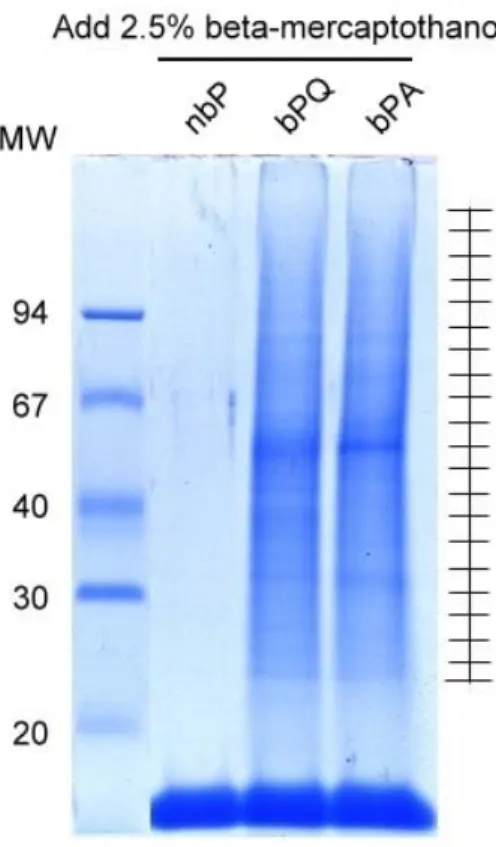
我們不將mouse cytosol直接進行in vitro transamidation的反應,再利用ammonium sulphate做沉澱, 以8 M urea/20 mM Tris-HCl 回溶沉澱後的蛋白,並利用streptavidin argerose進行純化步驟。最後 純化到的binding proteins,經電泳及Coomassie blue染色解析後,將SDS-PAGE上的lane(包含bPQ和 bPA)對分為二十等分,操作in-gel digestion,將tryptic peptides由膠片中萃取出,接著進行MS/MS 的分析(圖一),鑑定出的蛋白身分詳列於表一。經由蛋白質質譜鑑定,最後鎖定12個我們有興趣研究 的TG substrates,進行重組蛋白的製備。所選用的表現載體為pET21b,使用E. coli BL21作為宿主, 表現我們所挑選出蛋白的部分片段。預測這幾個蛋白的分子量理論值:LDH testis isoform為36 kDa、 NSFL1 cofactorp47為41 kDa、nudC為38 kDa、Peroxiredoxin-1為22 kDa、Retinal dehydrogenase為 54 kDa、TOM34為34 kDa、Glutathione S-transferase Mu 5為27 kDa、HSP70為70 kDa、APG-1為94 kDa、 Poly(A)-binding protein 1為71 kDa、Heterogeneous nuclear ribonucleoprotein K為51 kDa、hnRNP A2/B1為36 kDa。 在這裡我們盡量去表現出全長的蛋白,但其中HSP70與APG-1僅表現出1/3與1/2的長度, 這些表現蛋白在經過induction以及純化之後,接著進行in vitro transamidation以及biotin overlay assay。由結果顯示(圖二),在有外加TG的條件下,這些重組蛋白會有bPA incorporation的情形發生, 有些甚至會有高分子量的complex形成,懷疑是由於TG的cross-linking作用而將其重組蛋白本身作了 cross-linking所導致。將純化到之binding proteins,進行SDS-PAGE及轉印後,以各分子的抗體血清 操作西方轉印法(圖三)。由圖三結果我們可以發現,albumin, nudc, hsp70, tubulin, apg1及hsp90 在沒有加入任何的biotin probe sample裡,看不到訊號的產生,而在加入bPQ或是bPA的sampl裡,可以 看到有蛋白存在的訊號。
在此次的TG substrate純化與鑑定中,我們一共使用了兩種不同的probes(bPA:
5-biotin-amidopentylamine、bPQ:biotin-SGQQQLGSS ),其中bPA作為一個K donor,提供TG反應時 所需的amino group,bPQ作為一個Q donor,提供TG反應時所需的carboxamide group。所以本次實驗經 由MS/MS的分析結果,有些蛋白是TG利用bPA所鑑定到的受質,有些則是TG利用bPQ所鑑定到的受質,代 表這些蛋白有些是利用本身的帶有的carboxamide group與bPA反應,有些則是利用本身的amino group 與bPQ進行反應。而有些蛋白同時被鑑定為bPA和bPQ都可反應的分子時,代表這類的分子本身應該同時 具有Q donor和K donor,而這樣的分子本身就容易被TG作用,產生oligomer。像是Poly A-binding protein 1、Heat shock protein 70.2、Retinal dehydrogenase、Glutathione S-transferase 1、Peroxiredoxin
等。
TG substrates
bPA in testis
Molecula
r size
Function Mascot
score
Fragments
found
Accession #
Serum albumin
67 kD
Serum protein
407
14
P07724
Alpha-2-macroglobulin
220 kD
Protease inhibitor
218
4
Q61838
Pyruvate carboxylase
130 kD
Endogenous
biotin-conjugated protein
197 6
Z14044
Clathrin heavy chain
90 kD
Endocytosis
116
5
Q68FD5
(HLA-B-associated
transcript 3)
83 kD
Contains ubiquitin-like
domain
87 1 Q6ZQ38
Poly(A)-binding protein1
71 kD
Pre-mRNA splicing
592
17
P29341
Heterogeneous nuclear
ribonucleoprotein U-like
protein 1
96 kD
mRNA processing,
transcription repressor
124 2
Q8VDM6
Glial fibrillary acidic
protein,
50 kD
Glial intermediate filament
61
5
P03995
Tubulin alpha-6 chain
50 kD
Microtubule component
111
3
P68373
Heat shock 70 kDa protein
4
94kD
HSP 70 family chaperone
151
5
Q61316
Hexokinase-1
108 kD
Glycolysis enzyme
56
2
P17710
Protein KIAA1967
homolog
102 kD
Unknown
67
2
Q8VDP4
Heat shock protein HSP
90-alpha
86 kD
Molecular chaperone
769
17
P07901
Heat shock protein HSP
90-beta
86 kD
Molecular chaperone
408
11
P11499
Piwi-like protein 1
98 kD
RNA-binding protein
183
6
Q9JMB7
Microtubule-associated
protein 4
117 kD
Promotes microtubule
assembly
133 3
P27546
Polypyrimidine
tract-binding
protein-associated-splicin
g factor)
75 kD
Pre-mRNA splicing
73
2
Q8VIJ6
Sperm equatorial segment
protein 1
45 kD
Acrosome protein
67
3
Q9D5A0
Heat shock-related 70 kDa
protein 2
70 kD
Molecular chaperone
435
10
P17156
TG substrates
bPA in testis
Molecula
r size
Function Mascot
score
Fragments
found
Accession #
RNA-binding protein
EWS
68 kD
Possible transcription
repressor
79 3
Q61545
L-amino acid oxidase
70 kD
Lysosome enzyme
70
2
O09046
Matricin
70 kD
TCP-1 gamma chaperone
112
3
P80318
RNA-binding protein
FUS
53 kD
Nuclear riboprotein
92
3
P56959
Elongation factor Tu
50 kD
Protein translation
128
5
P10126
Alpha-2-antiplasmin
55 kD
Serine protease inhibitor
84
4
Q61247
Propionyl-CoA
carboxylase alpha chain
80 kD
Endogenous
biotin-conjugated protein
70 3
Q91ZA3
Retinal dehydrogenase 1
54 kD
Aldehyde dehydrogenase
153
5
P24549
UV excision repair
protein RAD23 homolog
B
43 kD
DNA repair
115
3
P54728
Heterogeneous nuclear
ribonucleoprotein K
51 kD
Pre-mRNA splicing
241
7
P61979
Y-box-binding protein 2 38 kD
Pre-mRNA splicing
166
4
Q9Z2C8
T-complex protein 1
subunit alpha A
60 kD
Molecular chaperone
73
3
P11984
Nucleoporin-like protein
RIP
58 kD
Acrosome biogenesis
68
3
Q8K2K6
Alpha-tubulin 3/7
50 kD
Microtubule component
415
9
P05214
GPI-anchored protein
p137
73 kD
Unknown
116
1
Q60865
Drebrin-like protein
49 kD
F-actin binding
60
2
Q62418
NSFL1 cofactor p47
41 kD
Golgi fragmentation and
reassembly
457 8
Q9CZ44
ATP-dependent RNA
helicase eIF4A-1
46 kD
Protein translation
106
2
P60843
Septin-2 (Protein
NEDD5)
41 kD
Cytokinesis
72
2
P42208
AU-rich element
RNA-binding protein 1
38 kD
mRNA turnover
56
3
Q60668
Beta-actin
42 kDa
Microfilament component
435
8
P60710
40S ribosomal protein SA 33 kDa
Ribosomal protein
113
3
P14206
LDH testis isoform
36 kDa
Glucose metabolism
936
31
P00342
APC-binding protein EB1 30 kDa
Microtubule and spindle
formaiton
121 3
Q61166
14-3-3 protein epsilon
29 kDa
Phosphoprotein binding
202
4
P62259
TG substrates
bPA in testis
Molecula
r size
Function Mascot
score
Fragments
found
Accession #
14-3-3 protein theta
28 kDa
Phosphoprotein binding
139
3
P68254
14-3-3 protein zeta/delta
28 kDa
Phosphoprotein binding
135
4
P63101
Tubulin beta-2C
50 kDa
Microtubule component
134
4
P68372
LDH-A
36 kDa
Glucose metabolism
128
9
P06151
DEAD box RNA helicase
DEAD1
69 kDa
Pre-mRNA splicing
114
2
Q61656
Peroxiredoxin-4
31 kDa
Redox regulation
146
4
O08807
Glutathione S-transferase
Mu 5
27 kDa
Testis GST
220
8
P48774
hnRNP-E1
37 kDa
RNA binding
117
2
P60335
snRNP-B
24 kDa
Pre-mRNA splicing
83
2
P27048
hnRNP F
45 kDa
Pre-mRNA splicing
80
4
Q9Z2X1
Glutathione S-transferase
A4
26 kDa
Cell detoxification
158
6
P24472
Peroxiredoxin-2
21 kDa
Redox regulation
112
3
Q61171
hnRNP H
49 kDa
Pre-mRNA splicing
112
3
O35737
TG substrates
bPQ in testis
Molecula
r size
Function Mascot
score
Fragments
found
Accession
#
Serum albumin
67 kD
Serum protein
418
9
P07724
Pyruvate carboxylase
129 kD
Endogenous
biotin-conjugated protein
140 6
Q05920
Alpha-tubulin 3/7
50 kD
Microtubule component
291
7
P05214
Clathrin heavy chain
191 kD
Endocytosis
98
2
Q68FD5
MAP 4
118 kD
Microtubule assembly
65
2
P27546
Elongation factor Tu
50 kD
Protein translation
371
12
P10126
GRP94; HSP 90 beta
92 kD
Molecular chaperone
336
9
P08113
HSP 90 alpha
85 kD
Molecular chaperone
708
15
P07901
Hsp 75
80 kD
Mitochondria chaperone
96
2
Q9CQN1
Beta-actin
42 kD
Microfilament component 415
10
P60710
Nucleolin
77 kD
Nucleolar protein
100, 72
2, 2
P09405
Spermatid-specific
thioredoxin-1
58 kD
Spermatogenesis
89
2
Q6P902
Transferrin
77 kD
Fe (III) binding
63
2
Q921I1
GFAP
50 kD
Glial intermediate filament 63
2
P03995
HSP 70.2
70 kD
Molecular chaperone
564
14
P17156
Heat shock cognate 71
kDa protein
71 kD
Molecular chaperone
364
7
P63017
Cortactin
61 kD
Src substrate
227
6
Q60598
Propionyl-CoA
carboxylase alpha chain
80 kD
Endogenous
biotin-conjugated protein
166 4
Q91ZA3
DEAD box RNA helicase
DEAD1
69 kD
Pre-mRNA splicing
118
3
Q61656
Heat shock 70 kDa
protein 4L
94 kD
Molecular chaperone
induced by osmotic stress
86 2
P48722
Kininogen-1
73 kD
Cys protease inhibitor
128
4
O08677
Y-box-binding protein 2
38 kD
mRNA processing
62, 95
2, 1
Q9Z2C8
Retinal dehydrogenase 1
54 kD
Aldehyde dehydrogenase
330
7
P24549
Tubulin beta-2C
49 kD
Microtubule component
245
7
P68372
ALDH class 2
56 kD
Aldehyde dehydrogenase
183
6
P47738
PAI mRNA-binding
protein 1
45 kD
mRNA stability
94
1
Q9CY58
Unc-33-like
phosphoprotein)
62 kD
Synaptic vesicle protein
66
2
Q62188
hnRNP F
46 kD
Pre-mRNA splicing
165
6
Q9Z2X1
TG substrates
bPQ in testis
Molecula
r size
Function Mascot
score
Fragments
found
Accession #
Proacrosin-binding
protein sp32
61 kD
Acrosome packaging
137
3
Q3V140
Argininosuccinate
synthase
46 kD
Urea cycle enzyme
110
3
P16460
hnRNP H
49 kD
Pre-mRNA splicing
80
2
O35737
Testis-specific gene A2
34 kD
Male miosis
196
4
Q8VIG3
hnRNP A3
40 kD
Pre-mRNA splicing
109
1
Q8BG05
Septin-2 (NEDD5)
42 kD
Cytokinesis
96
2
P42208
Leucine-rich
repeat-containing protein
46
nudC
38 kD
Mitotic spindle formation
191
6
O35685
GAPDH
36 kD
Glycolysis enzyme
397
8
P16858
Suppressor of G2 allele of
SKP1 homolog
38 kD
SCF complex component
127
3
Q9CX34
hnRNP A2/B1
36 kD
Pre-mRNA splicing
202
4
O88569
hnRNP C1 / C2
34 kD
Pre-mRNA splicing
89
2
Q9Z204
TOM34
34 kD
Mitochondria protein
import
494 14
Q9CYG7
Protein phosphatase 1C
catalytic subunit
37 kD
Signal transduction
84
2
P63087
LDH testis subunit
36 kD
Glycolysis
837
29
P00342
Swiprosin-1
27 kD
Calcium binding
146
4
Q9D8Y0
Nuclear protein Hcc-1
23 kD
Nucleic acid binding
92
2
Q9D1J3
TCP-1-theta
59 kD
Molecular chaperone
88
2
P42932
eIF4E
25 kD
Protein translation
61
2
P63073
Heat shock 70 kDa
protein 4L
94 kD
Molecular chaperone
86
2
P48722
14-3-3 protein epsilon
29 kD
Phosphoprotein binding
285
5
P62259
14-3-3 protein zeta/delta
28 kD
Phosphoprotein binding
182
4
P63101
eIF-4H
27 kD
Protein translation
92
1
Q9WUK2
Peroxiredoxin-1
22 kD
Redox regulation
405
13
P35700
HSP 27
23 kD
Molecular chaperone
124
2
P14602
AKAP 82
93 kD
Sperm motility
83
2
Q60662
14-3-3 protein beta/alpha 28 kD
Phosphoprotein binding
73
2
Q9CQV8
14-3-3 protein sigma
28 kD
Phosphoprotein binding
73
2
O70456
TG substrates
bPQ in testis
Molecula
r size
Function Mascot
score
Fragments
found
Accession #
Peroxiredoxin-2
21 kD
Redox regulation
235
5
Q61171
Glutathione S-transferase
Mu 1
26 kD
Cell detoxification
189
7
P10649
Peroxiredoxin-4
31 kD
Redox regulation
118
3
O08807
GST class-mu 5
26 kD
Cell detoxification
393
9
P48774
GST A4-4
25 kD
Cell detoxification
152
6
P24472
GST class-mu 6
25 kD
Cell detoxificatin
108
6
O35660
Poly(A)-binding protein 1 70 kD
Pre-mRMA splicing
107
3
P29341
Stathmin 18
kD
Microtubule
destabilization
hnRNP F
50 kD
Pre-mRNA processing
128
4
Q9Z2X1
五、參考文獻
Lorand, L. and Graham, R.M. (2003), Transglutaminases: crosslinking enzymes with pleiotropic functions, Nature Reviews Molecular Cell Biology 4, 140-156
Nakaoka, H., et al. (1994), Gh, a GTP-binding protein with transglutaminase activity and receptor signaling function, Science 264, 1593-1596
Fesus, L. and Piacentini, M. (2002), Transglutaminase 2: an enigmatic enzyme with diverse functions, Trends Biochem. Sci., 27, 534-539
Hu, B.H. and Messersmith, P.B. (2003), Rational design of translgutaminase substrate peptides for rapid enzymatic formation of hydrogels, J. Am. Chem. Soc., 125, 14298-14299
Ruoppolo, M., Orru, S., D’smato, A., Francese, S., Rovero, P., Marino, G. and Esposito, C. (2003), Analysis of translgutaminase protein substrates by functional proteomics, Protein Sci., 12, 1290-1297
Orru, S., Caputa. I., D’Amato, A., Ruoppolo, M. and Esposito, C. (2003), Proteomics identification of acyl-acceptor and acyl-donor substrates for transglutaminase in a human intestinal epithelial cell line, J. Biol. Chem., 278, 31766-31773
Gorman, J.J. and Folk, J.E. (1984), Structural features of glutamine substrates for transglutaminases, J. Biol. Chem., 259, 9007-9010
Figure 1. Purification of bPQ- and bPA- incorporated TG substrates from mouse testis cytosol.
Mouse testis crude extract was processed by in vitro transamidation. The biotin-labeled TG substrates were purified by streptavidin beads and separated by SDS-PAGE. Polyacrylamide gel of each lane was divided equally into 20 parts and subjected to in-gel digestion and LC-MS/MS analysis.