國 立 臺 中 教 育 大 學 教 育 學 院 體 育 學 系 碩 士 論 文
指 導 教 授 : 程 一 雄 教 授
協 同 指 導 : 呂 香 珠 教 授
八 週 綠 茶 萃 取 物 補 充 對 人 體 運 動
後 骨 骼 肌 肉 能 量 代 謝 之 影 響
Effect of Eight Weeks Oral Green Tea Extracts Supplementation on
Energy Metabolism Involved in Glycogen Synthesis and
Intramuscular Triglyceride in Exercised Human Skeletal Muscl e
研 究 生 : 賴 信 同 撰
I 謝 辭 終 於 到 了 這 一 天,回 想 當 初 大 學 時 因 為 當 實 驗 的 受 試 者 所 以 認 識 了 程 一 雄 老 師 , 每 次 實 驗 參 與 都 讓 我 覺 得 很 有 趣 , 也 從 不 想 讀 碩 士 變 成 想 讀 碩 士。在 這 三 年 不 長 也 不 短 的 碩 士 生 涯 裡 我 學 會 了 很 多 東 西 , 非 常 感 謝 程 一 雄 老 師 在 這 三 年 間 教 會 了 我 如 何 獨 立 思 考,並 告 訴 我 在 學 術 上 任 何 事 都 沒 有 所 謂 的 正 確 答 案 , 只 有 最 合 理 的 解 釋 。 感 謝 張 振 崗 老 師 提 供 了 完 善 的 實 驗 室 資 源 和 實 驗 上 許 多 的 問 題 解 答 以 及 新 知 識。感 謝 體 育 學 系 呂 香 珠 主 任 在 實 驗 過 程 以 及 論 文 寫 作 上 給 我 很 多 的 指 導。感 謝 體 育 學 系 系 辦 詹 淑 芬 小 姐 於 實 驗 期 間 與 口 試 過 程 中 協 助 我 處 理 許 多 事 務 。 更 感 謝 程 一 雄 老 師 、 彰 化 基 督 教 醫 院 廖 淑 芬 醫 生 以 及 中 國 醫 藥 大 學 蔡 岑 瑋 副 主 任 犧 牲 假 日 幫 助 我 完 成 實 驗,還 有 實 驗 室 的 嘉 珍 學 姊 、 顥 文 、 霖 鉉 、 鎮 陽 、 偉 駿 與 受 試 者 們 排 除 萬 難 配 合 實 驗 。 特 別 感 謝 榆 家 學 姐 以 及 榮 鑣 學 長 在 實 驗 上 教 會 我 許 多 細 節,以 及 學 術 上 幫 助 我 解 決 許 多 疑 問。感 謝 臺 北 市 立 大 學 運 動 科 學 所 運 動 生 化 實 驗 室 的 侯 建 文 老 師 的 耐 心 教 導 , 使 我 學 會 許 多 新 的 實 驗 技 術 。 感 謝 臺 中 教 育 大 學 生 理 實 驗 室、臺 灣 體 育 運 動 大 學 運 科 中 心 與 臺 北 市 立 大 學 運 動 科 學 所 運 動 生 化 實 驗 室 提 供 許 多 儀 器 以 及 良 好 的 實 驗 環 境。感 謝 這 些 實 驗 室 的 學 長 姐 、 同 學 以 及 學 弟 妹 們 , 在 我 忙 碌 時 幫 助 我 處 理 一 些 瑣 碎 的 事 情 。 最 後 感 謝 一 路 上 無 怨 無 悔 地 在 後 面 支 持 我 的 父 母 親 和 在 我 身 邊 不 離 不 棄 的 女 朋 友 悅 伶 以 及 台 北 的 摯 友 純 志、耀 弘 有 你 們 的 支 持 和 激 勵 我 才 能 從 許 多 次 低 潮 走 出 來 。 如 今 我 要 畢 業 了 , 接 下 來 換 我 在 你 們 身 邊 以 及 身 後 支 持 你 們 , 陪 伴 你 們 。 再 次 感 謝 這 三 年 來 身 邊 的 所 有 人 , 因 為 有 你 們 我 才 會 有 今 天 。 賴 信 同 謹 誌 中 華 民 國 1 0 4 年 1 月
II 摘 要 老 鼠 試 驗 顯 示 綠 茶 萃 取 物 ( G r e e n t e a e xt r a c t , G T E ) 能 有 效 分 解 脂 質 並 增 進 耐 力 運 動 表 現 。 肌 肉 肝 醣 ( mu s c le g l yc o g e n) , 是 骨 骼 肌 肉 重 要 能 量 來 源 , 另 外 長 時 間 耐 力 運 動 期 間 骨 骼 肌 肉 細 胞 三 酸 甘 油 脂 ( I n t r a m yo c e l l u l a r Tr i g l y c e r i d e , I M T G ) 和 肌 肉 肝 醣 一 樣 是 扮 演 能 量 重 要 提 供 來 源。目 前 沒 有 人 體 研 究 探 討 綠 茶 萃 取 物 補 充 是 否 能 增 加 運 動 恢 復 期 肌 肉 肝 醣 與 I M T G 的 濃 度,因 此 本 研 究 探 討 八 週 綠 茶 萃 取 物 補 充 對 運 動 後 骨 骼 肌 肉 能 量 代 謝 的 影 響 。 研 究 方 法 : 8 名 自 願 且 健 康 的 受 試 者 ( 平 均 年 齡 2 1 . 9 ± 0 . 7 7 歲 ) 補 充 8 週 綠 茶 萃 取 物 ( 5 0 0 mg / d a y) , 八 週 綠 茶 萃 取 物 補 充 前 後 , 受 試 者 分 別 接 受 7 5 % V‧O2 m a x 固 定 式 腳 踏 車 運 動 6 0 分 鐘 。 運 動 後 立 即 給 予 碳 水 化 合 物 飲 食 補 充 ( 每 公 斤 體 重 2 克 的 碳 水 化 合 物 , 8 0 % 碳 水 化 合 物 , 8 % 脂 肪 , 1 2 % 蛋 白 質 ), 觀 察 運 動 後 恢 復 期 0 和 3 小 時 , 收 集 股 外 側 肌 肉 樣 本 , 分 析 肌 肉 肝 醣 與 I M T G 濃 度 。 研 究 結 果 顯 示 , 8 週 綠 茶 萃 取 物 補 充 後 單 次 運 動 恢 復 期 的 0 至 3 小 時 肌 肉 肝 醣 增 加 量 1 7 . 0 3 ± 2 . 8 7 μ mo l/ g w e t t is s u e , 顯 著 性 高 於 綠 茶 萃 取 物 補 充 前 11 . 5 0 ±3 . 4 5 μ mo l/ g w e t t is su e ( p < . 0 5 )。 另 外 八 週 綠 茶 萃 取 物 補 充 後 顯 著 提 升 運 動 後 G L U T 4 蛋 白 質 表 現 。 然 而 綠 茶 萃 取 物 補 充 並 不 會 影 響 運 動 後 I M T G 濃 度 。 結 論 : 本 研 究 發 現 八 週 綠 茶 萃 取 物 補 充 能 使 運 動 後 肌 肉 肝 醣 再 合 成 增 加 , 提 高 肌 肉 中 的 G L U T 4 蛋 白 質 表 現 , 但 I M T G 無 顯 著 改 變 。 因 此 , 這 次 研 究 顯 示 8 週 綠 茶 萃 取 物 補 充 可 能 可 作 為 運 動 後 肌 肉 肝 醣 增 補 劑 。
關 鍵 字 : 綠 茶 萃 取 物 、 肌 肉 肝 醣 、 肌 肉 三 酸 甘 油 脂
III Abstract
Previous studies showed that green tea extracts (GTE) are able to reduce body fat mass in mice and humans. Skeletal muscle is the most important tissue involved in the regulation of glucose and fatty acids metabolism in human. Glycogen content and Intramyocellular Triglyceride (IMGT) in skeletal muscle influences the individuals’ endurance performance. The mechanism be hind this phenomenon and GTE impact on metabolic consequences in human skeletal muscle remains unclear. Therefore, the present study investigated whether eight weeks oral GTE supplementation enhanced fatty acid oxidation and attenuated glucose utilization resulting in enough glycogen reserves in exercised human skeletal muscle.
Eight healthy volunteer subjects (average age 21.9 ± 0.77 years old) were given 500mg of GTE per day for consecutive eight weeks. Before and after GTE supplementation, every subject was asked to perform cycling exercise for 60 minutes at 75% VO2 max and consumed a carbohydrate meal (2 g carbohydrate/kg body weight, 80 % carbohydrate, 8 % fat, 12 % protein) immediately after cycling exercise. Biopsied muscle samples were taken from va stus lateralis immediately and 3-h following exercise.
We found significantly increased muscle glycogen synthesis after GTE administration compared to pre-GTE supplement under condition after a single bout of exercise ( p < .05). Muscle GLUT4 protein expres sion was significantly elevated immediately after exercise in GTE Administration. However, IMTG level was not significantly altered with GTE administration. In contrast, evidences from our previous lab data about gaseous exchanges analyses showed that GTE had advantage effects on enhancing the fat oxidation during exercise recovery period
IV
after GTE supplementation. The phenomenon of IMTG and gaseous exchange after GTE supplementation implied that GTE enhanced fat oxidation not IMTG during exercise recovery period. However, The present study suggested that eight weeks oral GTE supplementation can be used as ergogenic aids, evidences from our results showed the beneficial effect of GTE on glycogen synthesis in e xercised human skeletal muscle.
V
目 錄
謝 辭 . . . Ⅰ 中 文 摘 要 . . . Ⅱ A b s t r a c e . . . Ⅲ 目 錄 . . . Ⅴ 表 目 錄 . . . Ⅷ 圖 目 錄 . . . Ⅸ 第 壹 章 緒 論 . . . 1 第 一 節 研 究 背 景 . . . 1 第 二 節 研 究 目 的 . . . 3 第 三 節 研 究 假 設 . . . 3 第 四 節 研 究 範 圍 與 限 制 . . . 4 第 五 節 名 詞 操 作 型 定 義 . . . 4 第 貳 章 文 獻 探 討 . . . 6 第 一 節 綠 茶 萃 取 物 . . . 6 第 二 節 肌 肉 肝 醣 與 運 動 的 關 係 . . . 8 第 三 節 葡 萄 糖 轉 運 蛋 白 4 與 肌 肉 中 三 酸 甘 油 脂 . . . 1 0 一 、 葡 萄 糖 轉 運 蛋 白 4 ( G l u c o s e t r a n s p o r t e r s - 4 , G L U T - 4 ) . . . 1 0 二 、 肌 肉 中 三 酸 甘 油 脂 ( I n t r a m y o c e l l u l a r T r i g l y c e r i d e , I M T G ) . . . 1 3VI 第 參 章 研 究 方 法 與 步 驟 . . . 1 5 第 一 節 研 究 對 象 . . . 1 5 第 二 節 實 驗 設 計 與 流 程 . . . 1 5 一 、 實 驗 設 計 . . . 1 6 二 、 最 大 攝 氧 量 ( V‧O2 m a x ) 測 量 與 運 動 強 度 設 定 . . . 1 6 三 、 碳 水 化 合 物 飲 食 內 容 . . . 1 8 四 、 綠 茶 萃 取 物 來 源 與 劑 量 . . . 1 8 五 、 實 驗 流 程 圖 . . . 1 9 第 三 節 實 驗 肌 肉 收 集 方 法 . . . 2 0 第 四 節 資 料 處 理 與 統 計 分 析 . . . 2 1 第 肆 章 結 果 . . . 2 2 第 一 節 受 試 者 基 本 資 料 . . . 2 2 第 二 節 骨 骼 肌 肝 醣 合 成 相 關 反 應 . . . 2 4 一 、 肌 肉 肝 糖 ( m u s c l e g l y c o g e n ) . . . 2 4 二 、 肌 肉 肝 醣 增 加 量 ( I n c r e a s e M u s c l e G l y c o g e n ) . . . 2 5 三 、 肌 肉 肝 醣 合 成 速 率 ( G l y c o g e n s y n t h e s i s r a t e ) . . . 2 6 第 三 節 骨 骼 肌 肝 醣 合 成 相 關 蛋 白 反 應 . . . 2 7 一 、 葡 萄 糖 轉 運 蛋 白 4 ( g l u c o s e t r a n s p o r t e r 4 , G L U T 4 ) . . . 2 7 二 、 肌 肉 中 三 酸 甘 油 脂 ( I n t r a m y o c e l l u l a r T r i g l y c e r i d e , I M T G ) . . . 2 9 三 、 肌 肉 中 三 酸 甘 油 脂 增 加 量 ( I n c r e a s e d I M T G ) . . . 3 0
VII 第 伍 章 討 論 . . . 3 1 第 一 節 主 要 發 現 . . . 3 1 第 二 節 綠 茶 萃 取 物 與 脂 肪 代 謝 . . . 3 1 第 三 節 骨 骼 肌 肝 醣 合 成 相 關 反 應 . . . 3 2 第 四 節 骨 骼 肌 中 三 酸 甘 油 脂 相 關 反 應 . . . 3 2 第 陸 章 結 論 . . . 3 3 參 考 文 獻 . . . 3 4 附 錄 . . . 3 9 附 錄 一 受 試 者 須 知 . . . 3 9 附 錄 二 受 試 者 同 意 書 . . . 4 0 附 錄 三 人 體 試 驗 委 員 會 通 過 證 明 . . . 4 2 附 錄 四 混 合 餐 點 營 養 成 分 表 . . . 4 3 附 錄 五 肌 肉 肝 醣 分 析 ( S k e l e t a l M u s c l e G l y c o g e n C o n t e n t A s s a y ) . . . 4 4 附 錄 六 西 方 點 墨 法 分 析 ( W e s t e r n B l o t t i n g A n a l y s i s ) . . . 4 7 附 錄 七 肌 肉 三 酸 甘 油 脂 分 析 ( I n t r a m y o c e l l u l a r T r i g l y c e r i d e , I M T G ) . . . 5 3
VIII
表 目 錄
表 3 - 1 測 量 受 試 者 個 別 攝 氧 峰 值 之 運 動 設 計 . . . 1 7 表 4 - 1 受 試 者 基 本 資 料 . . . 2 3
IX
圖 目 錄
圖 2 - 1 兒 茶 素 結 構 圖 . . . 6 圖 2 - 2 胰 島 素 影 響 G L U T - 4 的 路 徑 示 意 圖 . . . 1 2 圖 2 - 3 I M T G 與 運 動 生 理 反 應 . . . 1 3 圖 3 - 1 碳 水 化 合 物 餐 點 . . . 1 8 圖 3 - 2 實 驗 流 程 圖 . . . 1 9 圖 3 - 3 肌 肉 穿 刺 步 驟 . . . 2 0 圖 4 - 1 P r e 與 G T E 之 肌 肉 肝 醣 濃 度 . . . 2 4 圖 4 - 2 P r e 與 G T E 0 至 3 小 時 肌 肉 肝 醣 增 加 量 . . . 2 5 圖 4 - 3 P r e 與 G T E 0 至 3 小 時 肌 肉 肝 醣 合 成 速 率 . . . 2 6 圖 4 - 4 P r e 與 G T E 之 骨 骼 肌 G L U T 4 蛋 白 量 . . . 2 8 圖 4 - 5 P r e 與 G T E 之 I M T G 含 量 . . . 2 9 圖 4 - 6 P r e 與 G T E 0 至 3 小 時 I M T G 增 加 量 . . . 3 01
第壹章
緒論
第一節
研究背景
人 體 進 行 中 、 高 強 度 運 動 時 , 肌 肉 肝 醣 ( mu s c le g l yc o g e n) 是 主 要 的 能 量 來 源 ( R o m i jn e t a l. , 1 9 9 3 ) , 若 肌 肉 肝 醣 的 儲 存 量 降 低 , 則 運 動 表 現 也 會 跟 著 下 降 ( H e r ma n s e n, H u lt ma n , & S a lt in , 1 9 6 7 )。在 每 次 比 賽 只 有 約 3 - 4 小 時 的 休 息 時 間 下,前 一 場 比 賽 後 的 肌 肉 肝 醣 恢 復 速 率 若 不 夠 , 可 能 使 之 後 比 賽 中 的 運 動 表 現 不 佳 。 因 此 能 夠 提 高 運 動 時 能 量 的 轉 換 , 使 骨 骼 肌 肌 肉 肝 醣 合 成 量 增 加 的 營 養 增 補 劑 , 對 運 動 員 來 說 是 非 常 重 要 的 。 肌 肉 肝 醣 合 成 時 細 胞 內 部 至 少 需 要 碳 水 化 合 物 0 . 5 g / k g / h r, 當 碳 水 化 合 物 達 到 1 . 5 g / k g / hr 時 ( I v y, 1 9 9 8 ) , 肌 肉 肝 醣 回 補 量 為 最 高 ( D o y l e , S h e r m a n , & S t r a u s s , 1 9 9 3 ) 。 在 肌 肉 肝 醣 相 關 研 究 中 , 已 經 證 實 了 碳 水 化 合 物 的 攝 取 確 實 可 以 提 升 肝 醣 含 量,進 而 增 加 耐 力 運 動 表 現 與 持 續 時 間 ( B e r g s t r ö m, 1 9 6 2 ) 。 因 此 如 何 在 運 動 過 程 中 利 用 脂 肪 做 為 能 量 來 源,或 者 擁 有 較 多 的 肌 肉 肝 醣 含 量 , 皆 可 使 耐 力 運 動 表 現 時 間 維 持 更 長 。 在 長 時 間 運 動 表 現 中 , 肌 肉 肝 醣 會 不 斷 地 提 供 能 量 , 而 已 經 耗 盡 的 肌 肉 肝 醣 需 要 外 部 補 充 , 此 時 若 由 脂 肪 產 生 能 量 , 並 能 使 肌 肉 肝 醣 充 分 回 補 , 以 完 成 接 下 來 的 運 動 。 運 動 員 在 比 賽 中 除 了 技 術 的 表 現 , 更 重 要 的 是 在 比 賽 的 休 息 時 間 能 使 幾 乎 快 要 耗 竭 的 肌 肉 肝 醣 快 速 恢 復,並 保 持 良 好 身 體 狀 態。故 增 加 肌 肉 肝 醣 恢 復 量 以 及 能 量 的 轉 換 為 提 升 耐 力 運 動 表 現 的 方 法 之 一 。2 目 前 在 市 面 上 常 見 的 脂 肪 燃 燒 劑 有 羥 基 檸 檬 酸 ( h yd r o x yc it r i c a c i d , H C A ) 、 肉 鹼 ( c a r n it i n e ) 、 咖 啡 因 ( c a f f e i n e ) 、 共 軛 亞 麻 油 酸 ( c o n j u g a t e d l i n o l e i c a c i d , C L A ) 、 綠 茶 ( g r e e n t e a ) 等 ( J e u k e n d r u p , 2 0 1 0 ) 。 而 脂 肪 燃 燒 劑 與 運 動 表 現 相 關 的 研 究 有 , C h e ng 等 ( 2 0 1 2 ) 發 現 單 次 運 動 後 補 充 5 0 0 mg H C A 會 使 肌 肉 肝 醣 合 成 速 率 上 升 , 但 葡 萄 糖 轉 運 蛋 白 4 ( G L U T- 4 ) 蛋 白 表 現 下 降。Ts a o 等 ( 2 0 1 4 ) 給 予 1 2 名 健 康 男 性 混 合 C L A ( 3 . 8 g / d ) 8 週 , 發 現 肌 肉 肝 醣 和 G L U T- 4 蛋 白 表 現 皆 有 顯 著 性 上 升。P e d e r s e n 等 ( 2 0 0 8 ) 招 募 自 行 車 和 鐵 人 三 項 選 手 共 7 名, 在 運 動 後 給 予 c a f f e in e ( 8 mg / k g ) 發 現 肌 肉 肝 醣 和 磷 酸 化 蛋 白 質 激 酶 B ( p - A k t ) 蛋 白 表 現 皆 有 顯 著 性 上 升 。 有 動 物 研 究 顯 示,綠 茶 萃 取 物 它 具 有 分 解 脂 質 並 增 進 耐 力 運 動 表 現 的 效 果 ( M u r a s e , H a r a m iz u , S h i mo t o yo d o me , To k im it s u , & H a s e , 2 0 0 6 ) , 以 及 具 有 改 善 心 血 管 疾 病 、 抗 氧 化 、 增 加 脂 肪 代 謝 等 多 種 功 效 ( B o s c h m a n n & T h ie le c k e , 2 0 0 7 ) 。 有 研 究 指 出 2 4 0 名 肥 胖 受 試 者 ( 年 齡 介 於 2 5 ~ 5 0 歲 ) , 1 2 週 內 每 天 給 予 3 4 0 毫 升 兒 茶 素 飲 料 ( 實 驗 組 為 5 8 3 m g ; 控 制 組 為 9 6 m g ) , 結 果 顯 示 體 重 、 體 脂 肪 和 B M I 皆 顯 著 低 於 控 制 組 ( N a g a o , H a s e , & To k im it s u , 2 0 0 7 ) 。 另 有 研 究 指 出 單 次 補 充 綠 茶 萃 取 物 會 使 健 康 的 年 輕 男 性 在 中 等 強 度 的 運 動 過 程 中 增 加 脂 肪 的 氧 化,並 提 高 胰 島 素 敏 感 性 和 葡 萄 糖 的 耐 受 度 ( Ve n a b le s , H u ls t o n, C o x, & J e u k e n d r u p , 2 0 0 8 ) 。 M u r a s e 等 人 ( 2 0 0 6 ) 的 實 驗 觀 察 到 給 予 小 鼠 補 充 綠 茶 萃 取 物 加 上 運 動 , 發 現 耐 力 運 動 表 現 、 游 離 脂 肪 酸 以 及 脂 肪 酸 β 氧 化 有 顯 著 性 的 提 升 , 而 呼 吸 交 換 率 和 丙 二 醯 輔 酶 A ( M a lo n y l - C o A) 則 有 顯 著 性 下 降 , 這 些 結 果 顯 示 脂 肪 氧 化 有 提 升 。
3 目 前 尚 未 有 研 究 證 明 綠 茶 萃 取 物 能 否 作 為 肌 肉 肝 醣 增 補 劑,因 此 本 研 究 依 此 論 點 延 伸 , 探 討 補 充 8 週 綠 茶 萃 取 物 後 , 能 否 使 能 量 利 用 偏 向 脂 肪 氧 化 , 並 提 高 運 動 後 骨 骼 肌 肌 肉 肝 醣 再 合 成 率 , 進 而 使 綠 茶 萃 取 物 做 為 營 養 增 補 劑 是 否 有 功 效,對 於 增 加 運 動 後 人 體 骨 骼 肌 肌 肉 肝 醣 恢 復 率 增 加 的 可 能 性 。
第 二 節 研 究 目 的
探 討 人 體 補 充 八 週 綠 茶 萃 取 物 後 , 進 行 單 一 次 7 5 % V‧O2 m a x 腳 踏 車 運 動 6 0 分 鐘 後 , 立 即 攝 取 碳 水 化 合 物 是 否 可 以 改 變 肌 肉 肝 醣 、 G L U T- 4 和 肌 肉 三 酸 甘 油 脂 ( I n t r a m yo c e l l u l a r Tr i g l y c e r i d e , I M T G ) 的 濃 度 , 觀 察 能 量 代 謝 的 轉 換 以 及 肌 肉 肝 醣 再 合 成 效 果 。第 三 節 研 究 假 設
本 研 究 假 設 : 補 充 八 週 綠 茶 萃 取 物 對 於 單 次 7 5 % V O 2 ma x 運 動 挑 戰 後 給 予 碳 水 化 合 物 補 充 , 對 身 體 中 的 肌 肉 肝 醣 、 G L U T- 4 的 濃 度 皆 有 較 高 的 表 現 , 而 I M T G 呈 現 較 低 的 濃 度 。 這 會 使 運 動 能 量 偏 向 脂 肪 代 謝 以 及 肌 肉 肝 醣 回 補 率 有 較 高 的 表 現,因 此 骨 骼 肌 對 葡 萄 糖 的 利 用 率 提 升 , 進 而 促 進 肌 肉 肝 醣 再 合 成 率 增 加 。 意 味 著 補 充 八 週 綠 茶 萃 取 物 , 在 單 次 7 5 % V O 2 ma x 腳 踏 車 運 動 6 0 分 鐘 後 , 恢 復 期 間 補 充 碳 水 化 合 物 能 增 加 肌 肉 肝 醣 再 合 成 率 。4
第 四 節 研 究 範 圍 與 限 制
一 、 本 研 究 招 募 的 受 試 者 對 象 為 8 名 有 運 動 習 慣 的 健 康 男 性 , 年 齡 2 1 - 2 5 歲 的 大 專 生 。 二 、 本 研 究 無 法 證 實 透 過 肌 肉 穿 刺 取 得 部 分 肌 肉 樣 本 的 肌 纖 維 型 態 類 型 是 否 相 同 。 三 、 本 研 究 所 測 量 之 肝 醣 濃 度 為 運 動 後 0 及 3 小 時 之 變 化 , 無 法 推 估 運 動 前 以 及 運 動 完 3 小 時 之 後 的 肝 醣 濃 度 變 化 。第 五 節 名 詞 操 作 型 定 義
一 、 最 大 攝 氧 量 ( m a xi m u m o xy g e n c o n s u m p t i o n , V‧O2 m a x ) 測 量 攝 氧 量 的 過 程 中 出 現 之 最 大 值,並 非 個 體 攝 氧 量 的 絕 對 最 大 值 。 本 次 研 究 利 用 固 定 式 腳 踏 車 ( M o n a r k 8 7 4 E , Va n s b r o S w e d e n ) 以 及 運 動 心 肺 功 能 測 試 系 統 ( V ma x E n o r e 2 9 c S ys t e m) ( C a r e F u s io n, C a l i fo r n i a , U S A ) 測 量 受 試 者 攝 氧 量 , 腳 踏 車 功 率 保 持 6 0 r p m 阻 力 從 1 . 0 k g 開 始 , 4 分 鐘 後 每 2 分 鐘 增 加 0 . 5 k g , 當 攝 氧 量 不 再 隨 著 增 加 或 增 加 的 值 小 於 2 m l/ k g / m in 時 , 並 且 呼 吸 交 換 率 大 於 1 . 1 5 , 即 判 定 為 受 試 者 的 最 大 攝 氧 量 。 二 、 綠 茶 萃 取 物 ( G re e n Te a E xt r a c t G T E ) 本 研 究 增 補 劑 以 每 日 5 0 0 mg 綠 茶 萃 取 物 , 內 含 3 5 0 mg 的 茶 多 酚 ( p o l y p h e n o l s ) 與 1 5 0 m g 的 沒 食 子 兒 茶 素 沒 食 子 酸 酯 ( e p i g a l lo c a t e c h i n5 - 3 - g a l l a t e , E G C G ) 以 膠 囊 的 形 式 提 供 給 受 試 者 服 用 。 三 、 第 四 型 葡 萄 糖 轉 運 子 ( g lu c o s e t ra n s p o rt e r 4 , G L U T- 4 ) G L U T- 4 是 肌 肉 組 織 細 胞 中 最 主 要 的 葡 萄 糖 轉 運 體 , 肌 肉 細 胞 處 於 一 般 狀 態 下 時 , 僅 有 少 量 的 G L U T- 4 在 肌 肉 細 胞 膜 上 ; 而 當 肌 肉 呈 現 收 縮 狀 態 時 ,原 本 存 在 細 胞 內 囊 泡 中 的 G L U T- 4 會 被 大 量 轉 位 到 細 胞 膜 上 開 始 運 作 , 讓 葡 萄 糖 能 夠 進 入 細 胞 內 儲 存 利 用 。 而 這 樣 的 轉 移 的 動 作 分 別 受 到 胰 島 素 以 及 肌 肉 收 縮 兩 種 不 同 的 刺 激 所 控 制,葡 萄 糖 從 細 胞 外 運 送 至 細 胞 內 是 由 兩 種 不 同 的 機 制 所 調 控 ( S u h, P a ik , & J a c o b s , 2 0 0 7 ) 。 四 、 肌 肉 三 酸 甘 油 脂 ( I n t ra m y o c e l l u la r Tr i g ly c e ri d e , I M T G ) I M T G 和 肌 肉 肝 醣 在 長 時 間 耐 力 運 動 期 間 都 是 扮 演 能 量 提 供 者 的 角 色 。 有 學 者 提 出 , I M T G 在 身 體 處 於 不 同 生 理 情 境 下 所 扮 演 的 生 理 生 化 角 色 是 呈 現 兩 種 完 全 不 同 的 現 象 ( va n L o o n & G o o d p a s t e r, 2 0 0 6 ) 。
6
第貳章
文獻探討
第 一 節
綠 茶 萃 取 物
綠 茶 擁 有 豐 富 的 茶 多 酚 ( p o lyp h e no ls ) , 而 其 中 一 項 物 質 黃 烷 醇 ( f l a v a n o s ), 又 稱 作 兒 茶 素 ( c a t e c h i n s )。 兒 茶 素 為 綠 茶 眾 多 的 萃 取 物 之 一 , 包 含 了 沒 食 子 兒 茶 素 沒 食 子 酸 酯 ( e p ig a l lo c a t e c h i n - 3 - g a l la t e , E G C G )、兒 茶 素 沒 食 子 酸 酯 ( e p i c a t e c h i n - 3 - g a l l a t e , E C G ) 以 及 沒 食 子 兒 茶 素 ( e p ig a l lo c a t e c h in , E G C ) 等 物 質 ( Wa lt ne r - L a w e t a l. , 2 0 0 2 ) 。 圖 2 - 1 兒 茶 素 結 構 圖 ( K a o , H iip a k k a , & L ia o , 2 0 0 0 )7 有 研 究 顯 示,綠 茶 萃 取 物 和 相 等 量 的 咖 啡 因 相 比 之 下 對 於 刺 激 組 織 細 胞 產 生 熱 能 更 有 效 果 ( D u l lo o , S e y d o u x, & G ir a r d ie r, 1 9 9 6 ) 。 我 們 知 道 在 綠 茶 中 是 含 有 非 常 豐 富 的 黃 酮,而 在 許 多 的 多 酚 類 中 最 特 別 的 黃 酮 混 合 物 被 稱 為 兒 茶 酚 胺。兒 茶 酚 胺 已 經 被 研 究 出 能 夠 抑 制 兒 茶 酚 - O - 甲 基 轉 移 酶 ( c a t e c h o l O - m e t h y lt r a n s f e r a s e , C O M T ) , 能 夠 降 低 正 腎 上 腺 素 的 酵 素 ( B o r c h a r d t & H u b e r, 1 9 7 5 ) 。交 感 神 經 系 統 ( s y mp a t h e t ic n e r v o u s s y s t e m , S N S ) 在 神 經 傳 送 至 正 腎 上 腺 素 分 泌 是 一 個 很 重 要 的 角 色 , 它 能 夠 控 制 脂 肪 氧 化 以 及 生 熱 效 應 , 所 以 我 們 能 夠 想 像 如 果 兒 茶 酚 胺 抑 制 C O M T, 就 會 造 成 正 腎 上 腺 素 分 泌 更 多 , 進 而 促 使 脂 肪 氧 代 謝 和 生 熱 效 應 ( D u l lo o e t a l. , 1 9 9 9 ) 。 有 研 究 指 出 2 4 0 名 肥 胖 受 試 者 ( 年 齡 介 於 2 5 ~ 5 0 歲 ),1 2 週 內 每 天 給 予 3 4 0 毫 升 兒 茶 素 飲 料 ( 實 驗 組 為 5 8 3 mg ; 控 制 組 為 9 6 mg ) , 結 果 顯 示 體 重、體 脂 肪 和 B M I 皆 顯 著 低 於 控 制 組 ( N a g a o e t a l. , 2 0 0 7 )。M u r a s e 等 ( 2 0 0 6 ) 的 研 究 顯 示 給 予 小 鼠 補 充 綠 茶 萃 取 物 加 上 運 動 , 發 現 運 動 耐 力 表 現 、 游 離 脂 肪 酸 以 及 脂 肪 酸 β 氧 化 有 顯 著 性 的 提 升 , 而 呼 吸 交 換 率 和 丙 二 醯 輔 酶 A ( M a lo n y l - C o A) 則 有 顯 著 性 下 降 , 這 些 結 果 顯 示 脂 肪 氧 化 有 提 升 。 研 究 指 出 3 T 3 - L 1 脂 肪 細 胞 培 養 在 不 同 濃 度 的 E G C G 1 5 分 鐘 後 , 觀 察 到 乙 醯 輔 酶 A 羧 化 酶 ( Ac e t yl - C o A c a r bo x y la s e ) 的 抑 制 酶 長 鏈 醯 基 輔 酶 A( lo ng c h a in a c y l - C o A ) 出 現 。 因 此 , E G C G 可 以 改 變 脂 質 代 謝 的 路 徑 , 從 脂 肪 儲 存 轉 化 成 脂 肪 氧 化 ( Wa t a na b e , Ka w a b a t a , & N i k i , 1 9 9 8 ) 。 研 究 發 現 補 充 的 綠 茶 萃 取 物 中 含 有 8 1 . 3 % 茶 多 酚 的 肥 胖 小 鼠,在 無 運 動 和 有 運 動 下 皆 會 刺 激 全 身 脂 肪 利 用 率 ( S h i mo t o yo d o me , H a r a m i z u , I n a b a , M u r a s e , & To k i m i t s u , 2 0 0 5 )
8
第 二 節
肌 肉 肝 醣 與 運 動 的 關 係
肝 醣 在 人 體 中 儲 存 在 肌 肉 以 及 肝 臟 當 中,儲 存 在 肝 臟 中 的 我 們 稱 之 為 肝 臟 肝 醣 ( l iv e r g l yc o g e n),主 要 作 用 是 調 節 血 液 中 的 葡 萄 糖 濃 度 ; 而 儲 存 在 肌 肉 中 的 肝 醣 稱 之 為 肌 肉 肝 醣 ( mu s c le g l yc o g e n ) , 主 要 作 用 是 提 供 能 量 給 肌 肉 群 使 用 。 運 動 時 能 量 供 給 的 方 式 有 兩 種,主 要 是 取 決 於 運 動 強 度 和 時 間 長 短 。 當 運 動 強 度 在 低 強 度 以 下 時 , 偏 向 於 脂 肪 氧 化 分 解 提 供 運 動 所 需 之 能 量 ; 而 當 運 動 強 度 到 達 中 高 強 度 以 上 時 , 會 偏 向 於 分 解 肝 醣 而 提 供 能 量 。 隨 著 運 動 時 間 的 增 加 , 肌 肉 肝 醣 也 會 被 消 耗 殆 盡 , 當 肝 醣 耗 竭 時 疲 勞 感 就 會 產 生 。 因 此 在 運 動 比 賽 中 , 對 於 高 強 度 且 連 續 長 時 間 的 項 目,肌 肉 肝 醣 的 儲 存 量 以 及 休 息 時 的 肝 醣 再 合 成 量 會 是 勝 負 的 重 要 關 鍵 。 有 研 究 已 經 證 實 , 當 運 動 達 到 中 、 高 強 度 時 , 肌 肉 肝 醣 會 成 為 主 要 的 能 量 來 源 ( R o m i jn e t a l. , 1 9 9 3 )。B e r g s t r ö m 和 H u lt ma n ( 1 9 6 7 ) 的 研 究 中 發 現 , 當 運 動 強 度 越 大 時 , 肌 肉 肝 醣 能 源 的 需 求 也 越 大 。 在 運 動 過 程 中 , 疲 勞 感 與 肌 肉 肝 醣 耗 竭 的 程 度 是 有 相 關 的 , 當 肌 肉 肝 醣 耗 竭 時 , 肌 肉 就 無 法 維 持 高 強 度 耐 力 運 動 。 在 運 動 恢 復 期 時 肌 肉 肝 醣 回 補 至 正 常 含 量 是 很 重 要 的,而 運 動 時 骨 骼 肌 收 縮 及 胰 島 素 的 刺 激 是 影 響 肌 肉 中 葡 萄 糖 運 輸 的 主 要 原 因 ( G o o d y e a r, H i r s h m a n , S m it h , & H o r t o n , 1 9 9 1 ) 。 運 動 恢 復 期 的 肝 醣 再 合 成 分 為 快 速 合 成 期 與 慢 速 合 成 期 兩 個 不 同 的 階 段。在 快 速 合 成 期 時 , 不 需 要 依 賴 胰 島 素 就 可 以 使 肝 醣 快 速 合 成,主 要 是 因 為 運 動 會 引 起 葡 萄 糖 轉 運 蛋 白 4 ( g lu c o s e t r a n s p o r t e r, G L U T 4 ) 從 肌 肉 細 胞 中 轉 位 至 膜9 上 , 使 肌 肉 細 胞 對 於 血 液 中 葡 萄 糖 的 通 透 性 增 加 , 造 成 細 胞 內 葡 萄 糖 - 6 - 磷 酸 ( g l o u s e - 6 - p h o s p a t e, G - 6 - P ) 濃 度 上 升 , 以 及 肌 肉 細 胞 對 於 胰 島 素 的 敏 感 性 增 加 ( I v y, 1 9 9 8 ) , 所 以 這 一 階 段 也 可 稱 作 非 胰 島 素 依 賴 期 ( i n s u l i n - i n d e p e n d e n t p h a s e ) 。 另 一 階 段 為 慢 速 合 成 期 , 這 一 階 段 中 大 部 分 是 依 賴 胰 島 素 促 進 葡 萄 糖 運 送 至 肌 肉 細 胞 中,使 肝 醣 合 成 活 化 , 達 到 肝 醣 合 成 的 作 用 ( Wo jt a s z e w s k i e t a l. , 2 0 0 0 ) , 這 一 階 段 可 以 持 續 2 4 小 時 或 直 到 肌 肉 肝 醣 完 全 回 補 為 止 。 在 運 動 後 的 肝 醣 合 成 期 當 中,第 二 階 段 的 慢 速 合 成 期 之 肝 醣 合 成 速 率 只 有 第 一 階 段 快 速 合 成 期 的 1 0 - 3 0 % ( P r ic e e t a l. , 1 9 9 4 ) , 但 如 果 在 運 動 後 立 即 補 充 碳 水 化 合 物 , 又 會 高 出 慢 速 合 成 期 的 數 倍 , 之 後 持 續 補 充 碳 水 化 合 物 會 導 致 肝 醣 的 回 補 高 於 正 常 含 量 (Ivy, 1991)。
10
第 三 節
葡 萄 糖 轉 運 蛋 白 4 與 肌 肉 中 三 酸 甘 油 脂
一 、 葡 萄 糖 轉 運 蛋 白 4 ( G lu c o s e t r a n s p o rt e r s - 4 , G L U T- 4 ) ( 一 ) G L U T- 4 與 葡 萄 糖 之 關 係 絕 大 多 數 的 哺 乳 類 動 物 皆 須 要 葡 萄 糖 當 作 產 生 AT P 或 做 為 能 量 儲 存 。 因 為 葡 萄 糖 為 親 水 性 物 質 , 故 無 法 自 行 通 過 細 胞 膜 的 雙 層 脂 質 結 構 , 需 要 一 種 特 殊 的 蛋 白 攜 帶 , 透 過 促 進 擴 散 的 方 式 運 送 進 雙 層 膜 內 , 此 特 殊 蛋 白 稱 為 葡 萄 糖 轉 運 子 ( g lu c o s e t r a n s p o r t e r s ) , 簡 稱 為 G L U Ts。至 目 前 為 止 被 發 現 的 G L U Ts 共 有 1 3 種。其 中 又 以 G L U T- 4 是 主 要 在 骨 骼 肌 肉 細 胞 表 面 負 責 運 送 葡 萄 糖,是 骨 骼 肌 中 數 種 葡 萄 糖 轉 運 蛋 白 中 表 現 量 最 多 ( Ts a o , B u r c e l in , K a t z , H u a ng , & C h a r r o n, 1996 ) , 所 以 本 篇 主 要 探 討 G L U T- 4 。 研 究 顯 示 肌 肉 肝 醣 合 成 是 由 葡 萄 糖 的 運 送 速 率 限 制,而 且 肝 醣 合 成 增 加 被 認 為 是 與 葡 萄 糖 轉 運 子 第 四 型 ( g lu c o s e t r a ns p o r t e r - 4 G L U T- 4 ) 的 蛋 白 表 現 量 有 關 ( I v y & K U O , 1 9 9 8 ; R e n e t a l. , 1 9 9 3 ) 。 G L U T- 4 是 肌 肉 組 織 細 胞 中 最 主 要 的 葡 萄 糖 轉 運 體 , 當 肌 肉 細 胞 處 在 一 般 狀 態 下 , 僅 有 少 量 G L U T- 4 存 在 於 肌 肉 細 胞 膜 上 , 然 而 當 肌 肉 收 縮 時 ,原 本 存 在 細 胞 內 的 G L U T- 4 被 大 量 轉 位 到 細 胞 膜 上 用 來 運 送 葡 萄 糖 進 入 細 胞 內 以 供 儲 存 或 利 用。這 樣 轉 位 的 動 作 分 別 受 到 胰 島 素 與 肌 肉 收 縮 兩 種 不 同 刺 激 來 源 所 啟 動,意 味 著 葡 萄 糖 運 送 的 路 徑 是 由 兩 種 不 同 的 機 制 所 調 控 ( S u h e t a l. , 2 0 0 7 ) 。11 ( 二 ) G L U T- 4 與 胰 島 素 之 關 係 G L U T- 4 對 於 胰 島 素 是 屬 於 高 敏 感 性 的 , G L U T- 4 在 細 胞 膜 上 的 濃 度 會 受 到 胰 島 素 的 刺 激 而 上 升。當 胰 島 素 濃 度 增 加 或 肌 肉 收 縮 時 , 才 會 由 細 胞 内 轉 位 至 細 胞 膜 的 表 面 , 並 且 發 揮 運 送 葡 萄 糖 的 功 能 ( P r ic e e t a l. , 1 9 9 4 ) 。 胰 島 素 作 用 於 G L U T- 4 路 徑 如 下 :胰 島 素 會 先 與 細 胞 膜 上 的 胰 島 素 接 收 器 結 合 , 然 後 引 發 一 連 串 的 自 體 磷 酸 化 作 用 , 有 胰 島 素 接 受 器 受 質 1 ( i n s u l i n r e c e p t o r s u b s t r a t e - 1 , I R S - 1 ) 、 磷 脂 醯 醇 3 激 酶 ( p h o s p h a t i d y l i n o s it o 3 - k i n a s e , P I 3 - K ) , 形 成 磷 脂 醯 3 磷 酸 ( p h o s p h a t i d y l i n o s it o l 3 - p h o s p h a t e , P I 3 - P ) , 進 一 步 去 調 控 磷 酸 肌 醇 依 賴 激 酶 ( p ho s p h i no s it id e - d e p e nd e nt k in a s e , P D K) , 接 著 P D K 會 活 化 活 化 下 游 蛋 白 質 激 酶 B ( p r o t o t yp e s p r o t e in k in a s e B , P K B ), 而 P K B 更 進 一 步 活 化 Ak t s u b s t r a t e 1 6 0 ( AS 1 6 0 ) 蛋 白 。 有 研 究 發 現 , A S 1 6 0 是 一 種 小 型 的 G 蛋 白 , 它 能 協 助 G L U T- 4 轉 位 至 細 胞 膜 上 , 並 完 成 葡 萄 糖 運 輸 ( H e nr ik s e n, 2 0 0 2 ; Kr a me r e t a l. , 2 0 0 6 ) 。 目 前 我 們 知 道 胰 島 素 會 使 肌 肉 細 胞 利 用 葡 萄 糖 能 力 增 加 是 因 為 胰 島 素 會 刺 激 G L U T- 4 轉 位 到 膜 上 並 發 揮 葡 萄 糖 運 輸 的 功 能 路 徑 有 兩 條 。 其 中 一 條 路 徑 是 胰 島 素 影 響 胰 島 素 調 節 氨 肽 酶 ( in s u l i n - r e g u la t e d a m i n o p e p t i d a s e , I R A P ) , 進 而 影 響 G L U T- 4 。
12 圖 2 - 2 胰 島 素 影 響 G L U T- 4 的 路 徑 示 意 圖 ( R o w l a n d , F a z a k e r l e y, & J a m e s , 2 0 1 1 ) ( 三 ) G L U T- 4 與 運 動 之 關 係 運 動 時 會 影 響 骨 骼 肌 中 許 多 代 謝 產 物 或 離 子 濃 度 的 改 變。而 這 些 物 質 對 於 葡 萄 糖 的 利 用 與 運 輸 是 有 一 些 潛 在 的 影 響,這 些 影 響 與 胰 島 素 無 關 , 皆 發 生 於 運 動 期 間 ( C a r t e e & H o llo s z y, 1 9 9 0 )。 運 動 能 刺 激 活 化 一 系 列 的 訊 息 傳 遞 、 調 控 , 與 葡 萄 糖 的 吸 收 、 肝 醣 合 成 的 相 關 基 因 表 現 以 及 G L U T- 4 蛋 白 合 成 ( Kr a n io u , C a me r o n - S m it h, & H a r g r e a v e s , 2 0 0 4 ; Y. K r a n io u , C a m e r o n - S m it h , M i s s o , C o l l i e r, & H a r g r e a v e s , 2 0 0 0 ) 。 研 究 指 出,人 類 和 大 鼠 的 骨 骼 肌 經 過 單 次 激 烈 運 動 或 短 暫 的 運 動 訓 練 之 後 G L U T- 4 mR N A 的 含 量 會 顯 著 增 加 ( Kr a n io u e t a l. , 2 0 0 4 ; Kr a n io u e t a l. , 2 0 0 0 ; K u o , H u n t , D i n g , & I v y, 1 9 9 9 ) 。 另 一 個 人 體 研 究 證 明 , 運 動 可 以 向 上 調 節 G L U T- 4 mR N A 的 含 量,於 運 動 後 3 小 時 仍 維 持 在 高 含 量 。 運 動 誘 發 的 mR N A 含 量 提 高 通 常 會 伴 隨 相 對 應 蛋 白 質 的 快 速 增 加 ( P i le g a a r d e t a l. , 2 0 0 5 ) 。
13 二 、 肌 肉 中 三 酸 甘 油 脂 ( I n t r a m y o c e l lu l a r Tri g ly c e r i d e , I M T G ) ( 一 ) I M T G 與 能 量 及 生 理 的 關 係 體 內 儲 存 能 量 來 源 以 醣 類 、 脂 肪 為 主 。 全 身 脂 肪 分 布 極 廣 , 除 脂 肪 組 織 外 , 骨 骼 肌 肉 細 胞 內 含 I M T G 。 長 時 間 耐 力 運 動 期 間 , I M T G 和 肌 肉 肝 醣 一 樣 地 扮 演 能 量 提 供 者 。 有 學 者 提 出 看 法 , I M T G 在 身 體 處 於 不 同 生 理 情 境 下 所 扮 演 的 生 理 生 化 角 色 是 呈 現 南 轅 北 轍 的 現 象。 同 樣 是 高 I M T G 含 量,耐 力 運 動 者 生 理 反 應 是 呈 現 出 有 利 於 脂 肪 氧 化 , 而 且 有 利 於 提 升 胰 島 素 敏 感 度 。 相 反 地 , 不 運 動 者 或 沒 有 運 動 習 慣 的 人 高 I M T G 時 , 生 理 反 應 呈 現 出 不 利 於 脂 肪 氧 化 , 並 且 容 易 引 起 胰 島 素 阻 抗 ( 如 圖 2 - 3 ) ( M o r o , B a j p e y i , & S m it h , 2 0 0 8 ; v a n L o o n & G o o d p a s t e r, 2 0 0 6 ) 。 圖 2 - 3 I M T G 與 運 動 生 理 反 應 ( M o r o et a l. , 2 0 0 8 ) In tra m yo ce llu la r Li pi d (IM C L) co nt en t
14 肝 醣 濃 度 是 耐 力 運 動 表 現 限 制 因 素 之 一,然 而 當 肌 肉 肝 醣 耗 竭 時 , 身 體 的 能 量 利 用 會 從 碳 水 化 合 物 轉 換 成 脂 肪 氧 化。因 此 在 運 動 期 間 肝 醣 耗 竭 發 生 時 , 身 體 脂 肪 ( 包 含 I M T G ) 扮 演 能 量 提 供 的 重 要 角 色 , 以 利 維 持 身 體 運 動 的 延 續 ( Kr s s a k e t a l. , 2 0 0 0 ) 。 然 而 I M T G 分 解 和 合 成 的 機 制,明 顯 受 到 許 多 種 不 同 生 理 因 素 的 影 響,有 肌 肉 纖 維 的 型 態 ( D e B o c k e t a l. , 2 0 0 5 ) 、 運 動 訓 練 狀 態 、 性 別 ( Ta r n o p o ls k y e t a l . , 2 0 0 7 ) 和 飲 食 型 態 等 ( B a c h ma n n e t a l. , 2 0 0 1 ) 。 ( 二 ) I M T G 與 飲 食 的 關 係 D e c o m b a z , F l e it h , H o p p e l e r, K r e i s , & B o e s c h , ( 2 0 0 0 ) 的 研 究 中 指 出 運 動 恢 復 期 間 會 因 不 同 的 飲 食 內 容 物 ( 碳 水 化 合 物 、 脂 肪 比 例 ) 的 攝 取 , 對 於 身 體 肌 肉 肝 醣 含 量 和 肌 肉 脂 質 合 成 呈 現 反 比 例 的 現 象 , 因 此 不 同 的 飲 食 型 態 對 於 骨 骼 肌 肉 合 成 的 影 響 程 度 是 非 常 明 顯。然 而 , 文 獻 顯 示 飲 食 型 態 對 運 動 恢 復 期 I M T G 合 成 的 討 論 大 部 分 都 是 著 眼 於 不 同 脂 肪 比 例 餐 點 對 I M T G 含 量 的 影 響。少 有 研 究 證 明 運 動 後 脂 肪 燃 燒 劑 合 併 正 常 飲 食 攝 取 對 運 動 後 肌 肉 肝 醣、肌 肉 脂 肪 合 成 的 影 響 。 但 是,在 過 去 的 研 究 文 獻 中 所 提 到 的 飲 食 內 容 大 部 份 在 於 了 解 不 同 脂 肪 比 例 的 餐 點 ( 極 高 或 偏 低 的 脂 肪 比 例 ) 對 運 動 恢 復 期 I M T G 合 成 的 影 響,極 少 有 研 究 探 討 運 動 後 攝 取 正 常 飲 食 餐 點 ( 碳 水 化 合 物 6 0% 、 脂 肪 2 5 % 、 蛋 白 質 1 5 % ) 並 加 上 脂 肪 燃 燒 劑 , 在 運 動 恢 復 期 間 是 否 影 響 肌 肉 肝 醣 及 肌 肉 脂 肪 的 合 成 效 果 。
15
第參章
研究方法與步驟
第 一 節
研 究 對 象
受 試 者 為 8 名 有 規 律 性 運 動 的 健 康 男 性,自 願 且 有 告 知 實 驗 相 關 步 驟 及 注 意 事 項 。第 二 節
實 驗 設 計 與 流 程
本 研 究 實 驗 流 程 經 國 立 臺 灣 體 育 運 動 大 學 人 體 試 驗 委 員 會 審 查 同 意 , 每 位 受 測 者 在 參 與 實 驗 前 , 需 經 告 知 實 驗 目 的 與 內 容 取 得 受 測 者 同 意 後 簽 署 參 與 研 究 同 意 書。正 式 的 實 驗 開 始 之 前 會 先 進 行 一 次 的 前 置 試 驗,測 得 每 位 受 測 者 最 大 攝 氧 量 ( ma x i mu m o x yg e n c o ns u mp t io n, V ‧ O2 m a x ) , 做 為 受 測 者 實 驗 之 7 5 % V ‧ O2 ma x 運 動 強 度 換 算 使 用 。 此 研 究 設 計 以 前 後 測 方 式 進 行, 前 測 組 別 ( P r e - s u p p le me n) 為 補 充 八 週 綠 茶 萃 取 物 前 一 天,後 測 組 別 ( G T E ) 為 補 充 八 週 綠 茶 萃 取 物 ( 4 g / d ) 後 一 天 。 受 試 者 進 行 實 驗 的 前 一 晚 會 要 求 受 試 者 禁 食 8 小 時 , 直 到 實 驗 當 天 早 上 8 點 至 實 驗 室 接 受 採 血,接 著 熱 身 5 分 鐘 後 進 行 7 5 % V‧O2 m a x 腳 踏 車 運 動 6 0 分 鐘 , 運 動 期 間 可 以 自 由 飲 水 。 在 兩 次 的 試 驗 中 , 受 試 者 完 成 運 動 後 ( 0 h) 立 即 進 行 採 血 及 肌 肉 穿 刺 採 取 肌 肉 樣 本,第 一 次 採 取 肌 肉 樣 本 後 立 即 給 予 碳 水 化 合 物 飲 食,接 著 在 補 充 碳 水 化 合 物 後 的 第 3 小 時 進 行 第 2 次 肌 肉 穿 刺。實 驗 過 程 中 所 採 集 之 肌 肉 依 不 同 時 間 點 分 別 進 行 樣 本 分 析 。16 一 、 實 驗 設 計 實 驗 處 理 : 8 名 受 測 者 分 別 各 參 與 兩 次 試 驗 。 分 別 有 補 充 綠 茶 萃 取 物 前 及 補 充 八 週 綠 茶 萃 取 物 後 兩 組,實 驗 前 一 天 晚 上 晚 餐 過 後 禁 食 8 小 時 後 至 實 驗 室 立 即 進 行 單 次 7 5 % V‧O2 m a x 腳 踏 車 運 動 6 0 分 鐘 , 運 動 結 束 立 即 進 行 肌 肉 穿 刺 , 隨 後 立 即 補 充 碳 水 化 合 物 ( 每 公 斤 體 重 兩 克 ) 後 的 第 三 小 時 進 行 第 二 次 肌 肉 穿 刺 。 二 、 最 大 攝 氧 量 ( V‧O2 m a x ) 測 量 與 運 動 強 度 設 定 受 試 者 執 行 腳 踏 車 運 動 ( M o na r k 8 3 4 E , Va n s br o , S w e d e n), 並 戴 上 集 氧 式 面 罩 與 運 動 心 肺 功 能 測 試 系 統 ( V ma x E no r e 2 9 c S ys t e m, C a r e F u s io n , C a l i fo r n i a , U S A ) 。 運 動 開 始 0 ~ 4 分 鐘 , 受 試 者 維 持 每 分 鐘 6 0 轉 速 ( r e vo lu t io ns p e r m i n u t e , R P M ) 、 阻 力 功 率 為 0 . 5 k g 的 運 動 , 之 後 每 2 分 鐘 腳 踏 車 功 率 增 加 0 . 5 k g ( 表 3 - 1 ) , 直 到 受 試 者 攝 氧 量 達 穩 定 或 開 始 下 降 。 受 試 者 在 功 率 增 加 的 情 況 下,攝 氧 量 不 再 隨 著 增 加 或 增 加 的 值 小 於 2 ml/ k g / m i n; 且 呼 吸 交 換 率 ( r e s p i r a t o r y e x c ha n g e r a t io , R E R ) 大 於 1 . 1 5 ( Wi l l i a m s , P o w e r s , & S t u a r t , 1 9 8 6 ) 時 , 會 請 受 試 者 結 束 測 量 。 並 利 用 接 著 利 用 氣 體 分 析 儀 分 析 收 集 到 數 據 , 計 算 出 線 性 迴 歸 公 式 y = a x + b ( y = 公 斤 、 x = V O2) , 求 得 a ( 斜 率 ) 和 b ( 截 距 ) , 並 藉 由 上 述 公 式 求 得 每 位 受 試 者 在 最 大 攝 氧 量 7 5 % V‧O2 ma x 的 公 斤 數 , 即 為 受 試 者 在 正 式 實 驗 中 進 行 6 0 分 鐘 的 運 動 強 度 。
17 表 3 - 1 測 量 受 試 者 個 別 攝 氧 峰 值 之 運 動 設 計 階 段 持 續 時 間 ( m i n ) 負 荷 ( k p ) 轉 速 ( r p m) 阻 力 ( w a t t ) ( k g ) 1 0 ~ 4 0 . 5 6 0 3 0 0 . 5 2 4 ~ 6 1 . 0 6 0 6 0 1 . 0 3 6 ~ 8 1 . 5 6 0 9 0 1 . 5 4 8 ~ 1 0 2 . 0 6 0 1 2 0 2 . 0 5 1 0 ~ 1 2 2 . 5 6 0 1 5 0 2 . 5 6 1 2 ~ 1 4 3 . 0 6 0 1 8 0 3 . 0 7 1 4 ~ 1 6 3 . 5 6 0 2 1 0 3 . 5 8 1 6 ~ 1 8 4 . 0 6 0 2 4 0 4 . 0 9 1 8 ~ 2 0 5 . 0 6 0 3 0 0 5 . 0
18 三 、 碳 水 化 合 物 飲 食 內 容 每 位 參 與 實 驗 的 受 試 者 皆 給 予 每 公 斤 體 重 2 克 碳 水 化 合 物 飲 食 , 內 容 物 包 括 有 玉 米 片 ( 0 . 8 6 g / k g ) 、 低 脂 鮮 奶 ( 3 . 5 0 g / k g ) 、 白 土 司 ( 1 . 1 5 g / k g ) 、 草 莓 果 醬 ( 0 . 2 9 g / k g ) 、 葡 萄 糖 粉 ( 1 . 8 0 g / k g ) 、 水 ( 8 g / k g ) ( 如 圖 3 - 1 ) ( K a l l e n & L a z a r, 1 9 9 6 ; Wu , N i c h o l a s , Wi l l i a m s , To o k , & H a r d y, 2 0 0 3 ) 。 圖 3 - 1 碳 水 化 合 物 餐 點 四 、 綠 茶 萃 取 物 來 源 與 劑 量 本 研 究 使 用 補 充 劑 為 綠 茶 萃 取 物 ( 5 0 0 mg / d ) , 內 含 3 5 0 mg 的 p o l y p h e n o l s ( 多 酚 類 ) 與 1 5 0 m g 的 E G C G ( 兒 茶 素 ) 為 膠 囊 形 式 ( G e n e r a l N u t r it io n C e n t e r s , P it t s b u r g h , P e n n s y l v a n i a , U . S . ) 。
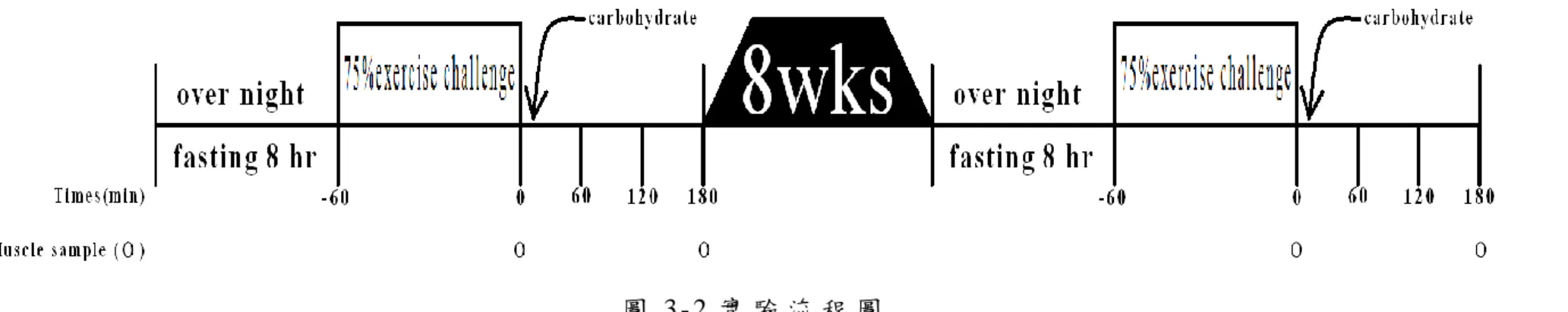
19 五 、 實 驗 流 程 圖
20
第 三 節
實 驗 肌 肉 收 集
本 實 驗 肌 肉 穿 刺 樣 本 取 樣 由 具 有 醫 師 執 照 之 合 格 醫 生 執 行,執 行 過 程 會 請 運 動 後 的 受 試 者 坐 在 長 椅 上 , 在 大 腿 股 外 側 肌 ( va s t u s l a t e r a l i s ) 上 做 一 個 簡 單 的 記 號 , 接 著 以 優 碘 與 酒 精 進 行 消 毒 三 次 後 , 鋪 蓋 上 無 菌 之 洞 巾 , 隨 後 注 入 1 - 2 m l 麻 醉 劑 ( 2 % L ig no c a in e ) 進 行 局 部 麻 醉 1 - 2 分 鐘 後,以 1 4 號 肌 肉 穿 刺 針 ( Te mno , M c G a w P a r k , I L , U S A) 於 股 外 側 肌 的 記 號 上 進 行 穿 刺 , 取 得 肌 肉 樣 本 。 樣 本 採 集 後 以 人 工 方 式 進 行 簡 單 的 去 血 和 去 脂 之 動 作,隨 即 置 入 液 態 氮 中 直 到 分 析 為 止 。 圖 3 - 3 肌 肉 穿 刺 步 驟21
第 四 節
資 料 處 理 與 統 計 分 析
實 驗 所 得 資 料 以 S P S S fo r WI N D O W S 1 2 . 0 版 統 計 套 裝 軟 體 進 行 分 析 : 一 、 以 相 依 樣 本 t 檢 定,比 較 2 個 試 驗 受 試 者 前 側 與 後 測 運 動 後 第 0、 3 小 時 肌 肉 樣 本 肝 醣 含 量、肝 醣 回 補 率、肌 肉 中 的 G L U T- 4 濃 度 、 以 及 I M T G 濃 度 之 差 異 。 二 、 所 有 數 據 以 平 均 值 ± 標 準 誤 ( me a n ± S E ) 表 示 , p < . 0 5 達 顯 著 水 準 。22
第肆章
結果
本 研 究 資 料 經 由 統 計 軟 體 處 理 後,將 所 得 結 果 分 為 受 試 者 基 本 資 料、骨 骼 肌 肝 醣 合 成 相 關 反 應 以 及 骨 骼 肌 肝 醣 合 成 相 關 蛋 白 反 應 等 三 部 份 , 分 別 敘 述 。第 一 節
受 試 者 基 本 資 料
8 名 受 試 者 平 均 年 齡 為 2 2 . 4 ± 0 . 5 6 歲 、 身 高 為 1 7 5 . 6 3 ± 1 . 1 8 公 分 , 補 充 G T E 前 體 重 為 7 5 . 4 3 ± 3 . 0 3 公 斤 、 身 體 質 量 指 數 為 2 4 . 4 9 ± 0 . 7 6 k g / m2, 補 充 後 體 重 為 7 7 . 3 9 ± 2 . 9 4 公 斤 、 身 體 質 量 指 數 為 2 5 . 1 3 ± 1 . 0 8 k g / m2, 最 大 攝 氧 量 為 4 3 . 2 3 ± 2 . 4 1 m l/ k g / m i n ( 如 表 4 - 1 ) 。23 表 4 - 1 受 試 者 基 本 資 料 項 目 P r e G T E 年 齡 ( ye a r s ) 2 2 . 4 ± 0 . 5 6 2 2 . 4 ± 0 . 5 6 身 高 ( c m) 1 7 5 . 6 3 ± 1 . 1 8 1 7 5 . 6 3 ± 1 . 1 8 體 重 ( k g ) 7 5 . 4 3 ± 3 . 0 3 7 7 . 3 9 ± 2 . 9 4 身 體 質 量 指 數 ( B M I ) ( k g / m2 ) 2 4 . 4 9 ± 0 . 7 6 2 5 . 1 3 ± 1 . 0 8 最 大 攝 氧 量 ( V‧O2 m a x ) ( m l / k g / m i n ) 4 3 . 2 3 ± 2 . 4 1 所 有 數 據 以 me a n ± S E 表 示 , ( N = 8 )
24
第 二 節
骨 骼 肌 肝 醣 合 成 相 關 反 應
一 、 肌 肉 肝 糖 ( m u s c l e g ly c o g e n ) 補 充 G T E 前 運 動 後 3 小 時 肌 肉 肝 醣 含 量 為 6 0 . 8 8 ± 5 . 6 9 μ mo l/ g 顯 著 高 於 運 動 後 0 小 時 4 9 . 3 9 ± 5 . 5 9 μ mo l/ g , 經 過 8 週 補 充 G T E 後 運 動 後 3 小 時 肌 肉 肝 醣 含 量 為 6 5 . 1 0 ± 7 . 9 7 μ mo l/ g 顯 著 高 於 運 動 後 0 小 時 4 8 . 0 7 ± 8 . 8 9 μ mo l / 。 如 圖 4 - 1 所 示 。 圖 4 - 1 P r e 與 G T E 之 肌 肉 肝 醣 濃 度※
P r e 組 為 補 充 八 週 綠 茶 萃 取 物 前 ; G T E 組 為 補 充 八 週 綠 茶 萃 取 物 後 。” * ” 表 示 與 0 小 時 比 較 達 顯 著 差 異 ( p < . 0 5 )。 數 值 以 me a n ± S E 呈 現 ( n = 8 ) 。25 二 、 肌 肉 肝 醣 增 加 量 ( M u s c le G l y c o g e n S y n t h e s i s ) 經 過 8 週 補 充 G T E 後 運 動 後 肌 肉 肝 醣 增 加 量 為 1 7 . 0 3 ± 2 . 8 7 μ mo l/ g 顯 著 高 於 補 充 前 11 . 5 0 ± 3 . 4 5 μ mo l/ g 。 如 圖 4 - 2 所 示 。 圖 4 - 2 P r e 與 G T E 0 至 3 小 時 肌 肉 肝 醣 增 加 量 ※ P r e 組 為 補 充 八 週 綠 茶 萃 取 物 前 ; G T E 組 為 補 充 八 週 綠 茶 萃 取 物 後 。” * ” 表 示 與 。 P r e 比 較 達 顯 著 差 異 ( p < . 0 5 )。 數 值 以 me a n ± S E 呈 現 ( n = 8 ) 。
26 三 、 肌 肉 肝 醣 合 成 速 率 ( G ly c o g e n s y n t h e s i s ra t e ) 經 過 8 週 補 充 G T E 後 運 動 後 肌 肉 肝 醣 合 成 速 率 為 5 . 6 8 ± 0 . 9 6 μ mo l/ g 顯 著 高 於 補 充 前 3 . 8 3 ± 1 . 1 5 μ mo l/ g 。 如 圖 4 - 3 所 示 。 圖 4 - 3 P r e 與 G T E 0 至 3 小 時 肌 肉 肝 醣 合 成 速 率 ※ P r e 組 為 補 充 八 週 綠 茶 萃 取 物 前 ; G T E 組 為 補 充 八 週 綠 茶 萃 取 物 後 。” * ” 表 示 與 P r e 比 較 達 顯 著 差 異 ( p < . 0 5 ) 。 數 值 以 me a n ± S E 呈 現 ( n = 8 ) 。
27
第 三 節
骨 骼 肌 肝 醣 合 成 相 關 蛋 白 反 應
一 、 葡 萄 糖 轉 運 蛋 白 4 ( g lu c o s e t r a n s p o rt e r 4 , G L U T 4 ) 比 較 P r e 與 G T E 兩 組 分 別 於 運 動 後 與 運 動 後 補 充 碳 水 化 合 物 第 3 小 時 採 集 肌 肉 樣 本 , 比 較 兩 試 驗 骨 骼 肌 G L U T 4 蛋 白 量 之 差 異 。 結 果 顯 示 補 充 8 週 G T E 後 , 運 動 後 3 小 時 顯 著 高 於 補 充 8 週 G T E 後 , 運 動 後 0 小 時 和 G T E 補 充 前,運 動 後 3 小 時( p < . 0 5 )。如 圖 4 - 4 所 示 。28 圖 4 - 4 P r e 與 G T E 之 骨 骼 肌 G L U T 4 蛋 白 量 ※ P r e 組 為 補 充 八 週 綠 茶 萃 取 物 前 ; G T E 組 為 補 充 八 週 綠 茶 萃 取 物 後 。” * ” 表 示 與 P r e 比 較 達 顯 著 差 異 ( p < . 0 5 ) 。 ” # ” 表 示 同 組 0 小 時 比 較 達 顯 著 差 異 ( p < . 0 5 ) 。 數 值 以 me a n ± S E 呈 現 ( n = 8 ) 。 * #
29 二 、 肌 肉 中 三 酸 甘 油 脂 ( I n t r a m y o c e l lu l a r Tri g ly c e r i d e , I M T G ) 比 較 P r e 與 G T E 兩 組 分 別 於 運 動 後 與 運 動 後 補 充 碳 水 化 合 物 第 3 小 時 採 集 肌 肉 樣 本 , 比 較 兩 試 驗 骨 骼 肌 I M T G 濃 度 之 差 異 。 結 果 顯 示 G T E 與 P r e 比 較 後 皆 無 顯 著 性 差 異 , 如 圖 4 - 5 所 示 。 圖 4 - 5 P r e 與 G T E 之 I M T G 含 量 ※ P r e 組 為 補 充 八 週 綠 茶 萃 取 物 前 ; G T E 組 為 補 充 八 週 綠 茶 萃 取 物 後 。 數 值 以 me a n ± S E 呈 現 ( n = 8 ) 。
30 三 、 肌 肉 中 三 酸 甘 油 脂 增 加 量 ( I n c re a s e d I M T G ) 比 較 P r e 與 G T E 兩 組 分 別 於 運 動 後 與 運 動 後 補 充 碳 水 化 合 物 第 3 小 時 採 集 肌 肉 樣 本 , 比 較 兩 試 驗 骨 骼 肌 I M T G 濃 度 之 差 異 。 結 果 顯 示 G T E 與 P r e 比 較 後 皆 無 顯 著 性 差 異 , 如 圖 4 - 6 所 示 。 圖 4 - 6 P r e 與 G T E 0 至 3 小 時 I M T G 增 加 量 ※ P r e 組 為 補 充 八 週 綠 茶 萃 取 物 前 ; G T E 組 為 補 充 八 週 綠 茶 萃 取 物 後 。 數 值 以 me a n ± S E 呈 現 ( n = 8 ) 。
31
第伍章
討論
第 一 節
主 要 發 現
本 實 驗 主 要 發 現:補 充 八 周 綠 茶 萃 取 物 會 使 單 次 固 定 式 腳 踏 車 在 7 5 % V‧O2 m a x 的 運 動 強 度 下 運 動 6 0 分 鐘 運 動 後 補 充 碳 水 化 合 物 肝 醣 合 成 速 率 較 高 , 而 I M T G 的 增 加 並 無 顯 著 差 異 , 顯 示 運 動 能 量 代 謝 可 能 偏 向 脂 肪 , 使 較 多 的 外 源 性 醣 類 用 於 肌 肉 肝 醣 合 成 。第 二 節
綠 茶 萃 取 物 與 脂 肪 代 謝
補 充 八 周 綠 茶 萃 取 物 後 並 無 明 顯 改 變 身 體 質 量 指 數,但 在 運 動 恢 復 期 的 呼 吸 交 換 率 ( R e s p ir a t o r y e x c ha ng e r a t io , R E R ) 會 偏 向 脂 肪 氧 化 , 這 證 據 來 自 先 前 本 實 驗 室 所 做 的 研 究 結 果 ( 陳 顥 文 , 民 1 0 3 ) 。 而 文 獻 顯 示 G T E 中 的 E G C G 會 抑 制 兒 茶 酚 氧 位 甲 基 轉 移 酶 ( c a t e c h o l - O - m e t h y l t r a n s f e r a s e , C O M T ) , 並 促 使 正 腎 上 腺 素 刺 激 脂 肪 細 胞 並 提 升 脂 肪 分 解 的 效 果 ( D u l lo o e t a l. , 1 9 9 9 ) 。 M a k i 等 人 ( 2 0 0 9 ) 發 現 G T E 加 上 運 動 可 以 提 高 腹 部 的 脂 肪 消 耗 。 Ve n a b le s 等 人 ( 2 0 0 8 ) 在 一 項 人 體 實 驗 中 發 現 , 在 實 驗 前 一 晚 給 於 1 2 名 受 試 者 補 充 G T E , 實 驗 當 天 1 2 名 受 試 者 皆 完 成 3 0 分 鐘 6 0 % 最 大 作 功 的 腳 踏 車 運 動,觀 察 運 動 3 0 分 鐘 內 的 能 量 消 耗,研 究 結 果 發 現 G T E 組 的 能 量 是 偏 向 於 脂 肪 氧 化 , 顯 著 性 高 於 控 制 組 1 7 %。 然 而 , 本 次 實 驗 結 果 顯 示 補 充 八 週 綠 茶 萃 取 物 後 單 次 運 動 後 恢 復 期 能 使 脂 肪 氧 化 提 升 , 但 是 不 會 改 變 I M T G 。32
第 三 節
骨 骼 肌 肝 醣 合 成 相 關 反 應
本 研 究 結 果 顯 示 , G T E 在 運 動 後 恢 復 期 肌 肉 肝 醣 含 量 、 增 加 量 以 及 合 成 速 率 , 均 顯 著 性 高 於 P r e 。 這 樣 的 結 果 顯 示 , 8 週 綠 茶 萃 取 物 補 充,對 於 運 動 後 恢 復 期 能 夠 增 加 肝 醣 合 成 速 率,且 文 獻 指 出 G T E 中 的 E G C G 可 增 加 G L U T 4 轉 位 至 膜 上 , 並 提 高 葡 萄 糖 吸 收 與 合 成 ( U e d a e t a l. , 2 0 0 8 ) , 最 後 轉 化 成 肌 肉 肝 醣 儲 存 。 骨 骼 肌 為 餐 後 吸 收 血 液 中 葡 萄 糖 的 重 要 器 官 之 一。運 動 後 補 充 碳 水 化 合 物 飲 食 會 促 進 肌 肉 肝 醣 的 再 合 成,主 要 的 機 制 是 因 為 胰 島 素 可 以 刺 激 G L U T 4 蛋 白 表 現 , 並 且 增 加 肌 肉 組 織 對 血 液 中 葡 萄 糖 吸 收 的 能 力 , 進 而 將 血 液 中 葡 萄 糖 運 輸 至 相 關 組 織 提 供 能 量 利 用 , 或 轉 變 成 肌 肉 肝 醣 儲 存 ( Ku o , H w a ng , L e e , C a s t le , & I v y, 2 0 0 4 ) 。 本 次 實 驗 補 充 8 週 綠 茶 萃 取 物 後 , 於 運 動 恢 復 期 3 小 時 內 , 肌 肉 中 肝 醣 含 量 、 肝 醣 增 加 量 以 及 合 成 速 率 皆 顯 著 高 於 補 充 前 , 以 上 結 果 可 以 印 證 G T E 在 動 物 實 驗 與 肌 肉 肝 醣 的 結 果 , M u r a s e 等 人 ( 2 0 0 5 ) 的 研 究 中 發 現 餵 食 綠 茶 萃 取 物 的 小 鼠 會 對 肌 肉 的 β 氧 化 作 用 產 生 變 化,有 助 於 提 高 耐 力 運 動 表 現 。第 四 節
骨 骼 肌 中 三 酸 甘 油 脂 相 關 反 應
H o r o w it z ( 2 0 0 3 ) 在 研 究 中 提 到 , 當 長 時 間 耐 力 運 動 進 行 時 , 脂 肪 組 織 的 三 酸 甘 油 酯 是 主 要 能 量 來 源 , 而 儲 存 在 脂 肪 組 織 中 的 T G 會 先 被 分 解 成 脂 肪 酸 , 以 利 運 動 中 的 肌 肉 使 用 。 有 動 物 研 究 指 出 補 充 E G C G 的 可 以 降 低 血 糖 和 三 酸 甘 油 酯 ( t r ig l y c e r i d e s , T G ) ( K a o e t a l. ,33 2 0 0 0 ) 。 另 外 一 個 動 物 實 驗 發 現 給 予 老 鼠 自 由 飲 用 六 週 綠 茶 ( 1 0 0 m l 水 0 . 5 g 綠 茶 粉 ) , 血 清 中 的 游 離 脂 肪 酸 ( f r e e f a t t y a c i d , F FA ) 和 T G 濃 度 有 顯 著 性 降 低 ( Wu , J u a n, H o , H s u , & H w a n g , 2 0 0 4 ) 。 另 外 有 人 體 實 驗 指 出 飲 用 含 有 綠 茶 兒 茶 素 的 飲 料 ( 6 2 5 mg / d a y) 可 以 提 升 運 動 造 成 的 腹 部 脂 肪 的 消 耗 以 及 減 少 F FA 和 T G 的 濃 度 ( M a k i e t a l. , 2 0 0 9 )。 此 次 研 究 顯 示 補 充 八 週 綠 茶 萃 取 物 後 不 會 引 響 單 次 運 動 後 恢 復 期 I M T G 的 濃 度 , 但 是 本 實 驗 室 先 前 G T E 補 充 運 動 恢 復 期 R E R 卻 明 顯 下 降 , 只 能 推 論 運 動 恢 復 期 脂 肪 氧 化 的 來 源 不 是 I M T G, 但 是 八 週 綠 茶 萃 取 物 補 充 可 以 節 省 單 次 運 動 後 碳 水 化 合 物 使 用 增 加,有 利 於 增 加 骨 骼 肌 肉 肝 醣 的 合 成 速 率 。
第陸章
結論
補 充 八 週 綠 茶 萃 取 物 後 , 經 單 次 7 5 % V‧O2 m a x 固 定 式 腳 踏 車 運 動 6 0 分 鐘 後 的 恢 復 期 , 立 刻 補 充 碳 水 化 合 物 飲 食 , 發 現 運 動 後 肌 肉 肝 醣 的 在 合 成 增 加 , 因 為 肌 肉 中 G L U T 4 蛋 白 表 現 增 高 使 得 骨 骼 肌 肉 對 葡 萄 糖 利 用 增 加 提 升 肝 醣 儲 存 , 然 而 I M T G 濃 度 並 無 明 顯 改 變 。 本 研 究 顯 示 八 週 綠 茶 萃 取 物 的 補 充 對 單 次 運 動 恢 復 期 骨 骼 肌 肉 能 量 儲 存 形 式 是 肌 肉 肝 醣 , 而 不 是 T G , 因 此 這 次 研 究 證 明 可 做 為 運 動 營 養 增 補 劑 , 可 利 用 於 肌 肉 肝 醣 的 回 補 而 非 I M T G 的 回 補 。 八 週 綠 茶 萃 取 物 補 充 可 作 為 提 升 運 動 恢 復 期 肌 肉 肝 醣 營 養 增 補 劑 。34
參 考 文 獻
Bachmann, O. P., Dahl, D. B., Brechtel, K., Machann, J., Haap, M., Maier, T., Schick, F. (2001). Effects of intravenous and dietary lipid challenge on intramyocellular lipid content and the relation with insulin sensitivity in humans. Diabetes, 50(11), 2579-2584.
Bergström, J., & Hultman, E. (1967). A study of the glycogen metabolism during exercise in man. Scandinavian Journal of Clinical & Laboratory Investigation, 19(3), 218-228. Bergstrom, J. (1962). Muscle electrolytes in man determined by neutron activation analysis
on needle biopsy specimens. Scandinavian Journal of Clinical and Laboratory
Investigation (England), 14(Suppl 68).
Borchardt, R. T., & Huber, J. A. (1975). Catechol O-methyltransferase. 5. Structure-activity relations for inhibition by flavonoids. Journal of medicinal chemistry, 18(1),
120-122.
Boschmann, M., & Thielecke, F. (2007). The effects of epigallocatechin-3-gallate on
thermogenesis and fat oxidation in obese men: a pilot study. Journal of the American
College of Nutrition, 26(4), 389S-395S.
Cartee, G., & Holloszy, J. (1990). Exercise increases susceptibility of muscle glucose transport to activation by various stimuli. American Journal of
Physiology-Endocrinology and Metabolism, 258(2), E390-E393.
Cheng, I., Huang, S.-W., Lu, H.-C., Wu, C.-L., Chu, Y.-C., Lee, S.-D., Kuo, C.-H. (2012). Oral hydroxycitrate supplementation enhances glycogen synthesis in exercised human skeletal muscle. British Journal of Nutrition, 107(07), 1048-1055.
De Bock, K., Richter, E. A., Russell, A., Eijnde, B. O., Derave, W., Ramaekers, M., Hespel, P. (2005). Exercise in the fasted state facilitates fibre type-specific intramyocellular lipid breakdown and stimulates glycogen resynthesis in humans. The Journal of
physiology, 564(2), 649-660.
Decombaz, J., Fleith, M., Hoppeler, H., Kreis, R., & Boesch, C. (2000). Effect of diet on the replenishment of intramyocellular lipids after exercise. European journal of
nutrition, 39(6), 244-247.
Doyle, J. A., Sherman, W. M., & Strauss, R. L. (1993). Effects of eccentric and concentric exercise on muscle glycogen replenishment. Journal of Applied Physiology, 74, 1848-1848.
Dulloo, A., Seydoux, J., & Girardier, L. (1996). Tealine and thermogenesis: interactions between polyphenols, caffeine and sympathetic activity. Int J Obes Relat Metab
35
Dulloo, A. G., Duret, C., Rohrer, D., Girardier, L., Mensi, N., Fathi, M., Vandermander, J. (1999). Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. The American
journal of clinical nutrition, 70(6), 1040-1045.
Goodyear, L. J., Hirshman, M. F., Smith, R. J., & Horton, E. S. (1991). Glucose transporter number, activity, and isoform content in plasma membranes of red and white skeletal muscle. Am J Physiol, 261(pt 1), E556-E561.
Henriksen, E. J. (2002). Invited review: Effects of acute exercise and exercise training on insulin resistance. Journal of Applied Physiology, 93(2), 788-796.
Hermansen, L., Hultman, E., & Saltin, B. (1967). Muscle glycogen during prolonged severe exercise. Acta Physiologica Scandinavica, 71(2‐3), 129-139.
Horowitz, J. F. (2003). Fatty acid mobilization from adipose tissue during exercise. Trends
in Endocrinology & Metabolism, 14(8), 386-392.
Ivy, J. (1998). Glycogen resynthesis after exercise: effect of carbohydrate intake.
International journal of sports medicine, 19(S 2), S142-S145.
Ivy, J., & KUO, C. H. (1998). Regulation of GLUT4 protein and glycogen synthase during muscle glycogen synthesis after exercise. Acta Physiologica Scandinavica, 162(3), 295-304.
Ivy, J. L. (1991). Muscle glycogen synthesis before and after exercise. Sports Medicine,
11(1), 6-19.
Jeukendrup, A. E. (2010). Sports Nutrition: From Lab to Kitchen. United Kingdom: Meyer & Meyer Sport.
Kallen, C. B., & Lazar, M. A. (1996). Antidiabetic thiazolidinediones inhibit leptin (ob) gene expression in 3T3-L1 adipocytes. Proceedings of the National Academy of
Sciences, 93(12), 5793-5796.
Kao, Y.-H., Hiipakka, R. A., & Liao, S. (2000). Modulation of endocrine systems and food intake by green tea epigallocatechin gallate 1. Endocrinology, 141(3), 980-987. Kramer, H. F., Witczak, C. A., Taylor, E. B., Fujii, N., Hirshman, M. F., & Goodyear, L. J.
(2006). AS160 regulates insulin-and contraction-stimulated glucose uptake in mouse skeletal muscle. Journal of Biological Chemistry, 281(42), 31478-31485.
Kraniou, G. N., Cameron-Smith, D., & Hargreaves, M. (2004). Effect of short-term training on GLUT-4 mRNA and protein expression in human skeletal muscle. Experimental
physiology, 89(5), 559-563.
Kraniou, Y., Cameron-Smith, D., Misso, M., Collier, G., & Hargreaves, M. (2000). Effects of exercise on GLUT-4 and glycogenin gene expression in human skeletal muscle.
36
Krssak, M., Petersen, K. F., Bergeron, R., Price, T., Laurent, D., Rothman, D. L., Shulman, G. I. (2000). Intramuscular Glycogen and Intramyocellular Lipid Utilization during Prolonged Exercise and Recovery in Man: A 13C and 1H Nuclear Magnetic
Resonance Spectroscopy Study 1. The Journal of Clinical Endocrinology &
Metabolism, 85(2), 748-754.
Kuo, C.-H., Hunt, D. G., Ding, Z., & Ivy, J. L. (1999). Effect of carbohydrate
supplementation on postexercise GLUT-4 protein expression in skeletal muscle.
Journal of Applied Physiology, 87(6), 2290-2295.
Kuo, C.-H., Hwang, H., Lee, M.-C., Castle, A. L., & Ivy, J. L. (2004). Role of insulin on exercise-induced GLUT-4 protein expression and glycogen supercompensation in rat skeletal muscle. Journal of Applied Physiology, 96(2), 621-627.
Li, M., Paran, C., Wolins, N. E., & Horowitz, J. F. (2011). High muscle lipid content in obesity is not due to enhanced activation of key triglyceride esterification enzymes or the suppression of lipolytic proteins. American Journal of
Physiology-Endocrinology and Metabolism, 300(4), E699-E707.
Maki, K. C., Reeves, M. S., Farmer, M., Yasunaga, K., Matsuo, N., Katsuragi, Y., Jones, F. (2009). Green tea catechin consumption enhances exercise-induced abdominal fat loss in overweight and obese adults. The Journal of nutrition, 139(2), 264-270. Moro, C., Bajpeyi, S., & Smith, S. R. (2008). Determinants of intramyocellular triglyceride
turnover: implications for insulin sensitivity. American Journal of
Physiology-Endocrinology and Metabolism, 294(2), E203-E213.
Murase, T., Haramizu, S., Shimotoyodome, A., Nagasawa, A., & Tokimitsu, I. (2005). Green tea extract improves endurance capacity and increases muscle lipid oxidation in mice. American Journal of Physiology-Regulatory, Integrative and Comparative
Physiology, 288(3), R708-R715.
Murase, T., Haramizu, S., Shimotoyodome, A., Tokimitsu, I., & Hase, T. (2006). Green tea extract improves running endurance in mice by stimulating lipid utilization during exercise. American Journal of Physiology-Regulatory, Integrative and Comparative
Physiology, 290(6), R1550-R1556.
Nagao, T., Hase, T., & Tokimitsu, I. (2007). A green tea extract high in catechins reduces body fat and cardiovascular risks in humans. Obesity, 15(6), 1473-1483.
Pedersen, D. J., Lessard, S. J., Coffey, V. G., Churchley, E. G., Wootton, A. M., Watt, M. J., & Hawley, J. A. (2008). High rates of muscle glycogen resynthesis after exhaustive exercise when carbohydrate is coingested with caffeine. Journal of Applied
Physiology, 105(1), 7-13.
37
Substrate availability and transcriptional regulation of metabolic genes in human skeletal muscle during recovery from exercise. Metabolism, 54(8), 1048-1055. Price, T., Rothman, D., Taylor, R., Avison, M., Shulman, G., & Shulman, R. (1994). Human
muscle glycogen resynthesis after exercise: insulin-dependent and-independent phases. Journal of Applied Physiology, 76(1), 104-111.
Ren, J.-M., Marshall, B., Gulve, E., Gao, J., Johnson, D., Holloszy, J., & Mueckler, M. (1993). Evidence from transgenic mice that glucose transport is rate-limiting for glycogen deposition and glycolysis in skeletal muscle. Journal of Biological
Chemistry, 268(22), 16113-16115.
Romijn, J., Coyle, E., Sidossis, L., Gastaldelli, A., Horowitz, J., Endert, E., & Wolfe, R. (1993). Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. American Journal of Physiology, 265, E380-E380. Rowland, A. F., Fazakerley, D. J., & James, D. E. (2011). Mapping insulin/GLUT4 circuitry.
Traffic, 12(6), 672-681.
Shimotoyodome, A., Haramizu, S., Inaba, M., Murase, T., & Tokimitsu, I. (2005). Exercise and green tea extract stimulate fat oxidation and prevent obesity in mice. Medicine
and science in sports and exercise, 37(11), 1884-1892.
Suh, S.-H., Paik, I.-Y., & Jacobs, K. (2007). Regulation of blood glucose homeostasis during prolonged. Mol Cells, 23, 272-279.
Tarnopolsky, M. A., Rennie, C. D., Robertshaw, H. A., Fedak-Tarnopolsky, S. N., Devries, M. C., & Hamadeh, M. J. (2007). Influence of endurance exercise training and sex on intramyocellular lipid and mitochondrial ultrastructure, substrate use, and mitochondrial enzyme activity. American Journal of Physiology-Regulatory,
Integrative and Comparative Physiology, 292(3), R1271-R1278.
Tsao, J.-P., Liao, S.-F., Korivi, M., Hou, C.-W., Kuo, C.-H., Wang, H.-F., & Cheng, I.-S. (2014). Oral conjugated linoleic acid supplementation enhanced glycogen resynthesis in exercised human skeletal muscle. Journal of sports
sciences(ahead-of-print), 1-9.
Tsao, T.-S., Burcelin, R., Katz, E. B., Huang, L., & Charron, M. J. (1996). Enhanced insulin action due to targeted GLUT4 overexpression exclusively in muscle. Diabetes, 45(1), 28-36.
Ueda, M., Nishiumi, S., Nagayasu, H., Fukuda, I., Yoshida, K.-i., & Ashida, H. (2008). Epigallocatechin gallate promotes GLUT4 translocation in skeletal muscle.
Biochemical and biophysical research communications, 377(1), 286-290.
van Loon, L. J., & Goodpaster, B. H. (2006). Increased intramuscular lipid storage in the insulin-resistant and endurance-trained state. Pflügers Archiv, 451(5), 606-616.
38
Venables, M. C., Hulston, C. J., Cox, H. R., & Jeukendrup, A. E. (2008). Green tea extract ingestion, fat oxidation, and glucose tolerance in healthy humans. The American
journal of clinical nutrition, 87(3), 778-784.
Waltner-Law, M. E., Wang, X. L., Law, B. K., Hall, R. K., Nawano, M., & Granner, D. K. (2002). Epigallocatechin gallate, a constituent of green tea, represses hepatic glucose production. Journal of Biological Chemistry, 277(38), 34933-34940.
Watanabe, J., Kawabata, J., & Niki, R. (1998). Isolation and identification of acetyl-CoA carboxylase inhibitors from green tea (Camellia sinensis). Bioscience, biotechnology,
and biochemistry, 62(3), 532-534.
Williams, J. H., Powers, S. K., & Stuart, M. K. (1986). Hemoglobin desaturation in highly trained athletes during heavy exercise. Medicine and science in sports and exercise,
18(2), 168-173.
Wojtaszewski, J., Hansen, B. F., Kiens, B., Markuns, J., Goodyear, L., & Richter, E. (2000). Insulin signaling and insulin sensitivity after exercise in human skeletal muscle.
Diabetes, 49(3), 325-331.
Wu, C.-L., Nicholas, C., Williams, C., Took, A., & Hardy, L. (2003). The influence of high-carbohydrate meals with different glycaemic indices on substrate utilisation during subsequent exercise. British Journal of Nutrition, 90(06), 1049-1056. Wu, L.-Y., Juan, C.-C., Ho, L.-T., Hsu, Y.-P., & Hwang, L. S. (2004). Effect of green tea
supplementation on insulin sensitivity in Sprague-Dawley rats. Journal of
Agricultural and Food Chemistry, 52(3), 643-648.
陳顥文. (民 103). 補充八週綠茶萃取物對人體肌肉肝醣之影響(碩士論文). 取自臺灣博 碩士論文系統.
39