Purification,
Characterization,
and Molecular Cloning of
Gonadotropin
Subunits of Silver Carp (Hypophthalmichthys
molitrix)
Y.
S. CHANG,~.J.
HUANG,F. L.
HUANG,C.
S.LIU,
ANDT. B. Lo
Institute of Biological Chemistry, Academia Sinica and Znstitute of Biochemical Science, National Taiwan University, P.O. Box 23-106, Taipei, Taiwan
Accepted May 17, 1989
The a and i3 subunit of silver carp gonadotropin (scGTH-a and scGTH-g) were isolated by high-performance liquid chromatography. Heterogeneity of N-terminal amino acid sequence was observed in scGTH-a but not in scGTH-g. For determining the complete primary structures of scGTH-a and scGTH-g, their cDNAs were cloned. Combiig the data of N- and C-terminal sequences determined from proteins and the amino acid sequences deduced from cDNAs, we infer that scGTH-a consists of 95 and/or 93 residues and scGTH+ consists of 115 residues. Both scGTH-a and scGTH-g are glycoprotein. Their carbohydrate content is about 20 g per 100 g protein. The molecular weights of scGTH-a and scGTH-g were calculated to be 12,700 and 15,700 Da, respectively. The amino acid sequences of scGTH-a and scGTH-l3 are very similar to those of the corresponding subunit of carp GTH, diierent in only 2 and 4 residues, respectively. In addition, a high extent of homology (70%) was also observed between the CL subunits of fish and mammalian GTHs. In the case of g subunit, homology among various species of fish (75 to 98%) is much higher than that between fish and mammal (40%). These data suggest that the u subunit is conserved while the f3 subunit is diversified during the molecular evolution of vertebrate GTH. o 1990 Academic press, ~nc.
Mammalian
anterior pituitary gland syn-
thesizes and secretes three glycoprotein
hormones.
They are luteinizing
hormone
(LH), follicle-stimulating
hormone (FSH),
and thyroid-stimulating
hormone
(TSH).
The former two are collectively
designated
as gonadotropin
(GTH). All of them consist
of two nonidentical
subunits, the tx and l3
subunit. Upon nonconvalent
association of
these two subunits, hormonal
function
is
achieved.
Within
a given species, all of
these glycoprotein
hormones
share the
same a subunit while the p subunit is hor-
mone specific (see review of Pierce and
Parsons, 1981). Like mammalian
GTH, te-
leostean GTH is also synthesized and se-
creted from anterior
pituitary
gland and
composed
of two nonidentical
subunits.
The function of teleostean GTH on game-
togenesis and steroidogenesis has been well
documented
(see review of Idler and Ng,
1983).
The primary
structures of the a and l3
subunits of mammalian
GTHs from several
species had been reported (see review of
Pierce and Parsons, 1981). Comparison
of
known sequences of a subunits indicates
that they are highly conserved from species
to species. Similarly,
homology between l3
subunit is also apparent. There are regions
of striking conservation,
not only for the
same hormone
between species, but also
between hormones.
The a and p subunits of GTH from sev-
eral species of teleost have been purified
and characterized
(Burzawa-Gerard,
1974;
Chang et
al.,1988b; Huang et
al.,1982;
Itoh et
al.,1988; Suzuki et
al.,1988; Trinh
et al.,
1986). Compared to those of mamma-
lian GTHs, the primary structures of the a
and p subunits of fish GTH are not so ex-
tensively studied. Up to now, only the com-
plete sequence of the
asubunit of carp
GTH (cGTH-a)
and the p subunit of cGTH
23
0016-64W9O $1.50
Copyri&t 8 1990 by Academic Press. Inc. All rights of npmduction in any form reserved.
24 CHANG ET AL.
(cGTH+)
(Chang et al., 1988b) and salmon
GTH (sGTH+)
(Itoh et al., 1988; Trinh et
al., 1986) have been determined or deduced
from the nucleotide
sequence of cDNA.
The comparative
data show that there is a
high extent of homology
(70%) in amino
acid sequence existing among the cx subunit
of carp and mammalian
GTHs. As for the l3
subunit, the homology
between cGTH and
sGTH is high (75%) while that between fish
GTH and mammalian
GTH is low (40%).
Such findings suggest that the OL subunit is
conserved while the p subunit is diversified
during the molecular
evolution
of verte-
brate GTH.
The above findings are only based on the
data of two species of fish. In order to pro-
vide a wider basis for comparative
study,
we are attempting
to determine the primary
structures of the o and p subunit of GTHs
from more species of fish. This paper is one
of a series of such studies. Here, we report
the purification
and characterization
of the
[Y and /3 subunit of GTH (scGTH-(Y and
scGTH+)
from silver carp (Hypophthal-
michthys molitrix)
and the determination
of
their primary structures by molecular clon-
ing.
MATERIALS
AND METHODS
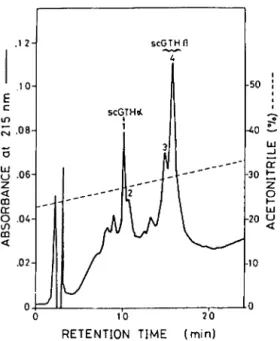
Purification of scGTH-a and scGTH+. The scGTH was purified as previously described (Chang et al., 1988a). Purillcation of scGTH-a and scGTH-8 was performed by high-performance liquid chromatogra- phy (HPLC). The scGTH was dissolved in H,O and chromatographed through a Nucleosil C,, column (4.6 X 250 mm, 7 pm) equilibrated with solvent A (22.5% acetonitrile-0.07% trifluoroacetic acid, TFA) and eluted by a linear gradient of solvent B (40% acetonitrile-O.07% TFA) in which 70% of solvent B was achieved at 60 min. The flow rate was 1 ml/mm. Operation of HPLC was performed at 40”. Individual peak was collected and recovered by lyophilization.
Bioassay of scGTH-a and scGTH-p. The GTH ac- tivities of scGTH-a and scGTH-B and their recombi- nant were assayed by the method of androgen produc- tion by carp testis in vitro as previously described (Huang and Chang, 1980). Reassociation of scGTH-a and scGTH-B was performed by mixing them at a 1: 1 molar ratio (Iinal concentration, 1 mg/ml) in 50 mjl4
phosphate buffer, pH 7.4, and incubated at 25” for 2 hr. Chemical analyses. The amino acid composition was determined by the method as described by Chang and Liu (1988). The N-terminal amino acid sequence was determined by a 477A protein sequencer and an on-line 120A phenylthiohydantoin analyzer of Applied Biosystems, Foster City, California (Hewick et al., 1981). For determination of C-terminal amino acid se- quence, scGTH-a and scGTH-B were digested with carboxypeptidase Y (Mill&e Corp., Bedford, MA) (substrate:enzyme, 30:1, w/w) in 50 n&f pyridine acetate, pH 5.4, at 37” for 0, 10, and 20 min by the method of Hayashi (1977). The enzymatic digestion was stopped by the addition of acetic acid. After dry- ing, released amino acids were converted to dimeth- ylaminoazobenzenesulfonyl(DABSYL)aminoacidand analyzed (Knecht and Chang, 1986). The carbohydrate content was estimated by using the thiobarbituric acid method for sialic acid (Warren, 1959), the orcinol re- action for neutral sugar (Winzler, 1955), and the p- dimethylaminobenzaldehyde reaction for amino sugar (Rondle and Morgan, 1955). The sodium dodecyl sul- fate (SDS)-polyacrylamide gel electrophoresis (PAGE) was performed as described by Laemmli (1970). The scGTH and its subunits were quantified by their protein content determined by the method of Lowry et al. (1951) with bovine serum albumin as ref- erence.
Radioimmunoassay. The scGTH-a and scGTH-$ were used as antigen to induce antisera in rabbit by multiple-site injection (Vaitukaitis et al., 1971). Iodin- ation of protein was performed by the &lot-amine-T method (Greenwoods et al., 1%3). For assay, 10,080 cpm of labeled antigen and a proper dilution of antise- rum which bound 30% of labeled antigen in the ab- sence of unlabeled antigen were used. The assay con- ditions were essentially the same as those described by Roser et al. (1984).
Construction of cDNA library. The polyadenylated mRNA of silver carp pituitary glands was prepared from liquid nitrogen frozen tissue by the guanidinium/ CsCl method (Ulhich er al., 1977) followed by oligo- dT cellulose column chromatography. Double- stranded cDNA, synthesized by the method of Gubler and Hoffman (1983), was ligated with EcoRI linker and subsequently inserted into the EcoRI site of pUC 19. The Escherichia coli strain JM 101 was used as host.
Screening of cDNA library. The cDNAs encoding cGTH-a and cGTH-B (Chang et al., 1988b) were used as probes to screen the cDNA coding for scGTH-a and scGTH-8, respectively. The cDNAs used for probes were labeled with ‘*P-dATP by nick translation.
Bacteria containing recombinant plasmids were fust grown on nitrocellulose filter, lysed with NaOH, baked, and then hybridized by the method of Grun- stein and Hogness (1975). For further confirmation the plasmid DNAs of positive clones were extracted and
digested with EC&I, subsequently subjected to elec-
trophoresis in agarose, and then hybridized in situ with
two electrophoretic bands. As described
labeled probes (Ridd et al., 1983).below, the N-terminal amino acid sequence
DNA sequence analysis. In order to analyze the
analysis indicated that peak 1 had a similar
complete nucleotide sequences of the cloned cDNAs,sequence to that of cGTH-a while peaks 3
they were further subcloned. The cDNAs encondingand 4 had a similar sequence to that of
scGTHa and scGTH-E were cleaved with PsrI andcGTH-B (Table 1). Consequently, peak 1
AvaII, respectively. After separation by electrophore-sis in agarose and electroelution, the resulted DNA
was designated as scGTH-a and peaks 3
fragments were subcloned into pUC 19. The nucle-and 4 were collectively designated as
otide sequences were determined by the dideoxynu-
scGTH-B.
cleotide chain termination method (Sanger et al.,1977).
GTH Acitivity
of scGTH-a,
scGTH-P,
and
Their Recombinant
RESULTS
As shown in Fig. 2, either scGTH-a or
Isolation
of scGTH-a and scGTH+
scGTH+ expressed very low GTH activity
By the HPLC system used in this study, while the recombinant of scGTH-a and
scGTH could be resolved into four major scGTH+ expressed high GTH activity
peaks (Fig. 1). When they were analyzed by fully comparable to native scGTH when as-
SDS-PAGE, peaks 1,3, and 4 showed one sayed by androgen production by carp tes-
electrophoretic band while peak 2 showed tis
in vitro.
These results further indicate
that scGTH-a and scGTH-B isolated by
HPLC are the two different subunits of
scGTH.
scGTH0 -
5
3
l;il”_
Itl
scGTH-a (AS-a) and that against scGTH$
E
(AS-g) were induced in rabbit. Both AS-a
tj .06- __-*
__---
30 tand As-l3 reacted strongly to the homolo-
5__--
__--
_--- 25
gous antigen but weakly to the heterolo-
2 --
L
0 .04- 2
20 2
gous antigen. Calculated at
B/B0 = 50%,
AS-a
showed 3.7% cross-reactivity toward
u
scGTH-B while As-B showed 5.6% cross-
.02- 10
reactivity
toward scGTH-a (data not
shown).
0 0
0 10 20
Chemical Properties
of scGTH-a
RETENTION TIME (mid
and scGTH$
FIG. 1. Reverse-phase HPLC fractionation of the
subunits of silver carp GTH. Ten micrograms of
The amino acid composition, carbohy-
scGTH dissolved in HZ0 was loaded onto a Nucleosildrate content, and N- and C-terminal amino
C,s column (4.6 x 250 mm, 7 pm) equilibrated withacid sequence of scGTH-a and scGTH-B
solvent A (22.5% acetonitrile-0.07% trifluoroaceticacid, TFA). The sample was eluted with a linear gra-
are presented in Table 1. Both scGTH-a
dient in which 70% of solvent B (40% acetoni-and scGTH-B had high content of Asp/Asn,
trile-O.O7% TFA) was reached at 60 min at a flow rateGlu/Gln, Pro, and half Cys. In comparison,
of 1 ml/mm. Operation of HPLC was performed at 40”.the scGTH-a contains more basic but less
Radioimmunoassay
In order to further characterize scGTH-a
and scGTH+, an immunological approach
was also undertaken. Antiserum against
CHANG ET AL.
26
30 I-
DOSE (pg/tube)
FIG. 2. The GTH activities of native scGTH, its Q and p subunits, and the recombinant of u and p subunits. The method of androgen production by carp testis in vitro (Huang and Chang, 1980) was used for assay of GTH activity. Each point is the mean of three incubations. Vertical line indicates standard deviation.
acidic amino acid residues than scGTH-P.
In addition, scGTH-a had a lower carbohy-
drate content than scGTH-P,
mainly attrib-
utable to amino sugar.
The amino acid sequence analysis indi-
cated heterogeneity
in N-terminal
sequence
was observed
in scGTH-a
but not in
scGTH+.
Two forms of scGTH-a
were
found, one with Tyr-Pro-kg-Asn-Asp-
while the other one had Arg-Asn-Asp
as
their N-terminal
sequence. For scGTHj3,
the N-terminal
sequence was identified
to
be Ser-Phs-Leu-Pro-Pro-.
The C-terminal
sequences of scGTH-a and scGTH+
were
determined
to be -Lys-Ser
and -Phe-Pro,
respectively.
The molecular
weights of
scGTH-a and scGTH-P
estimated by SDS-
PAGE were 17,400 and 22,670 Da, respec-
tively. However, these values are different
from those calculated by summing up the
molecular
weights of constituting
amino
acid residues and carbohydrate
content of
the molecule,
which yielded 12,700 Da for
scGTH-a and 15,700 Da for scGTH+.
Nucleotide
Sequencing of cDNA
Encoding scGTH-a
From 548 recombinant
cDNA clones, 4
positive clones were obtained when cDNA
encoding cGTH-a was used as a probe. The
nucleotide sequence and the deduced ami-
no acid sequence of one cDNA encoding
scGTH-a are presented in Fig. 3. As shown
in Fig. 3, it has 869 base pairs (bp) in length,
consisting of 31 bp of the 5’ untranslated
region, 354 bp of the open reading frame,
and 481 bp of the 3’ untranslated
region.
The open reading frame encodes a polypep-
tide of 118 residues which contains a 95-
residue protein with the same N- and C-
terminal
sequences as those determined
from scGTH-a.
The amino acid composi-
tion predicted from cDNA is very close to
that determined
from scGTH-a
(Table 1).
TABLE 1
THECHEMICALPROPERTIESOF scGTH-a AND scGTH-B
a subunit B subunit
Protein” cDNAb Protein” cDNAb
Amino acid composition”
LYS 9.2 (9) 10 4.0 (4) 4 His 3.2 (3) 3 3.0 (3) 3 ‘b 3.9 (4) 4 4.1 (4) 4 Asx 10.2 (10) 10 9.1 (9) 9 Thr 6.6 (7) 7 10.3 (10) 11 Ser 5.8 (6) 6 7.6 (8) 9 Glx 6.3 (6) 6 11.6 (12) 12 pro 6.4 (6) 6 13.2 (13) 13 GUY 3.7 (4) 3 4.1 (4) 3 Ala 4.7 (5) 4 2.7 (3) 2 ll2Cys 9.6 (10) 10 12.1 (12) 12 Val 8.4 (8) 8 11.4 (11) 11 Met 1.9 (2) 2 2.0 (2) 2 Ile 2.9 (3) 3 3.2 (3) 3 Leu 5.4 (5) 5 7.1 (7) 7 ‘b 4.2 (4) 5 5.6 (6) 6 Phe 3.0 (3) 3 4.0 (4) 4 Total (95) 95 (115) 115
Amino acid sequence
N-terminal (Tyr-Pro)-Arg-Asn-Asp- Ser-Phe-Leu-Pr+Pr+
C-terminal -LysSer -Phe-Pro
Carbohydrate contentd
Neutral sugar 10.2 10.6
Amino sugar 6.6 11.2
Siahc acid 1.3 2.1
0 Determined from protein by amino acid analysis. b Predicted from nucleotide sequence of cDNA.
c Number of residues. The integrated number is shown in parentheses. d g per 100 g protein.
Nucleotide
Sequencing of cDNA
Encoding scGTH+
There are three positive clones obtained
when cDNA encoding cGTH+
was used as
a probe for screening of the silver carp pi-
tuitary cDNA library. The nucleotide
and
deduced amino acid sequence of one cDNA
encoding scGTH+l
are presented in Fig. 4.
It has 554 bp in length, consisting of 18 bp
of the 5’ untranslated
region, 423 bp of the
open reading frame, and 110 bp of the 3’
untranslated
region.
The open reading
frame encodes a 115residue
protein with
the same N- and C-terminal
sequences as
those determined from scGTH-P.
The data
of Table 1 also indicated that the amino acid
composition
predicted from cDNA is very
close to that determined
from scGTH+.
DISCUSSION
The scGTH has been purified and char-
acterized (Chang et al., 1988a). In this pa-
per, we continued our previous work to pu-
rify and characterize the subunits of scGTH
for the purpose of getting a better under-
standing of scGTH and of providing a wider
basis for comparative
study of vertebrate
GTHs.
The two subunits of scGTH prepared in
this study seem to be highly purified,
as
28
CHANG ET AL.CATCTCACTGGAAGTCAAGAACAAAGCCATC ATG TTT TGG ACA AGA TAT GCT GGA 55 Met Phe Trp Thr Arq Tyr Ala Gly
-23 -20
GCA AGT ATA TTA TTG TTT TTA ATG CTT ATT CAT CTT GGA CAA GTA TAT 103 Ala Ser Ile Leu Leu Phe Leu Met Leu Ile His Leu Gly Gin Val Tyr
-10 -1 1
CCA AGA AAT GAT ATT ACT AAC TTT GGA TGT GAG GAG TGC AAA CTC AAG 151 Pro Arg Asn Asp Ile Thr Asn Phe Gly Cys Glu Glu Cys Lys Leu Lys
10
GAG AAC AAC ATT TTC TCA AAA CCC GGC GCT CCC GTC TAT CAG TGT ATG 199 Glu Asn Am Ile Phe Ser Lys Pro Gly Ala Pro Val Tyr Gln Cys Met
20 30
GGA TGC TGC TTT TCC AGG GCT TAC CCC ACA CCC CTG AGG TCC AAG AAA 247 Gly Cys Cys Phe Ser Arg Ala Tyr Pro Thr Pro Leu Arq Ser Lys Lys
40
ACC ATG CT'I' GTT CCC AAA AAT ATC ACA TCA GAA GCT ACA TGC TGT GTA 295 Thr Met Leu Val Pro Lys Asn Ile Thr Ser Glu Ala Thr Cys Cys Val
50 60
GCC AAA GAA GTT AAA CGG GTA CTT GTC AAT GAT GTC AAA CTA GTG AAC 343 Ala Lys Glu Val Lys Arq Val Leu Val Asn Asp Val Lys Leu Val Asn
70 80
CAC ACA GAC TGC CAC TGT AGC ACC TGT TAC TAT CAC AAA TCT TAA AAA 391 His Thr Asp Cys His Cys Ser Thr Cys Tyr Tyr His Lys Ser ***
90 95 CACTATGACATTTCAAATTTACTTGTGTTTGCTTACTTACTTATATTCCTGTACCTATTTTTT 454 CTGCAGTGCTTATTTTCTGTTCTTGATTCACAATGACTTACATATTTAAAGTGAAAACATACT 517 GTTAGAAGTTTGCTATATACCCGTACTGTGCAAATTTTCTTCATACTGTCGATTGTTTTAAAC 580 AATTCTTTTTTAAAAGGTTGTCATAATTGTTGTAATTTGTGCCCTACTTCCATAAATAGCTTA 643 AAATGCAATATTTTATCATTTATAAAATGCAAGATAACTTCATTACTATGCTCTGCTTGGTAT 706 TATTTTAAACCCTCTTTTGTTGGTAATTGCTGACTTGTTTTGTCGTCGCTGCTATACGATTGT 769 TCAAATACATTAATTAAAAACAAGATTGCTTATGA (n=66) 869
FIG. 3. The nucleotide sequence and the deduced amino acid sequence of cDNA encoding scGTH-a.
evidenced
by the following
criteria:
(1)
Both scGTH-a and scGTH-B have only one
electrophoretic
band in SDS-PAGE;
(2) re-
association
of scGTH-a
and scGTH-B
could fully restore the hormonal
function
(Fig. 2); (3) the antiserum against scGTH-a
and that against scGTH+
had very low
cross-reactivity
toward their counterpart
subunits;
and (4) both
scGTH-a
and
scGTH-B have their own unique N- and C-
terminal
sequence (Table 1). However,
it
should be noted that heterogeneity
is found
in the N-terminal
sequence of scGTH-a.
Such phenomenon
was also observed in the
case of ovine LH-a (Liu et al., 1972).
Molecular
cloning has been widely used
to determine the primary structure of pro-
tein. Combining
the data of N- and C-
terminal
sequences determined
from pro-
tein and the amino acid sequence deduced
from cDNA, we infer that scGTH-a
has 95
and/or 93 residues while scGTH-B
has 115
residues.
The C-terminal
residue
of
scGTH-a is the same one just preceding the
stop codon while that of scGTH-B
is the
one which is two residues ahead of the stop
AGAGGGACACCTGTCAAG ATG TTA GCT GTT CGA AAC AAC ATC CTC CTT CTC 51
Met Leu Ala Val Arg Asn Am Ile Leu Leu Leu
-24 -20
TTA TTC TGT TTA GTT GTT CTG CTA GTC TTT GCT CAA AGC TCT TTT CTT 99 Leu Phe Cys Leu Val Val Leu Leu Val Phe Ala Gln Ser Ser Phe Leu
-10 -1 1
CCA CCA TGT GAG CCA GTT AAT GAG ACT GTT GCA GTG GAG AAA GAG GGC 147 Pro Pro Cys Glu Pro Val Asn Glu Thr Val Ala Val Glu Lys Glu Gly
10
TGT CCA AAA TGT CTG GTG TTT CAG ACC ACC ATC TGC AGT GGC CAC TGC 795 Cys Pro Lys Cys Leu Val Phe Gln Thr Thr Ile Cys Ser Gly His Cys
20 30
CTA ACA AAG GAG CCT GTA TAC AAG AGC CCA TTT TCC ACT GTC TAC CAA 243 Leu Thr Lys Glu Pro*Val Tyr Lys Ser Pro Phe Ser Thr Val Tyr Gln
40 50
CAC GTG TGC ACT TAC CGG GAC GTC CGC TAT GAG ACA GTC CGC TTG CCA 291 His Val Cys Thr Tyr Arg Asp Val Arg Tyr Glu Thr Val Arg Leu Pro
60
GAC TGT CCT CCC GGG GTG GAC CCC CAT ATC ACT TAC CCG GTG GCT CTC 339 Asp Cys Pro Pro Gly Val Asp Pro His Ile Thr Tyr Pro Val Ala Leu
70 80
AGC TGC GAC TGC AGC CTC TGC ACC ATG GAC ACG TCC GAC TGT ACC ATC 387 Ser Cys Asp Cys Ser Leu Cys Thr Met Asp Thr Ser Asp Cys Thr Ile
90
GAA AGC CTG CAG CCT GAT TAC TGC ATG TCT CAG AGG GAG GAT TTC CCT 435 Glu Ser Leu Gin Pro Asp Tyr Cys Met Ser Gln Arg Glu Asp Phe Pro
100 110
GTG TAT TAG CCTACAGGAGTACTGTCCTGTCATCAAACCACAAAGCCCACTCTAAATCAG 494 Val Tyr ***
117
ATAAATGTCACATAGATGTATATCAATAAAAAACTACATACTTCATA~n=,3~ 554
FIG. 4. The nucleotide sequence and the deduced amino acid sequence of cDNA encoding scGTH-8.
codon. Consequently,
there must be a post-
translational
modification
of scGTH-8
by
proteolytic
cleavage of two residues (-Va&
Tyr) from the precursor. Post-translational
modification
at the C-terminal
part is also
observed in cGTH-8
(Chang ef al., 1988b)
and mammalian
LH-B (Maurer,
1985) and
TSH-l3 (Maurer
et al., 1984), but not in
sGTH-B
(Itoh er al., 1988; Trinh et al.,
1986) and mammalian
FSH-8 (Esch et al.,
1986).
Recently, two distinct GTHs, designated
as GTH I and II, from chum salmon pitu-
itary glands were isolated by Suzuki et al.
(1988). They share a common OL subunit but
have a different B subunit. The homology of
the amino acid sequence between these two
subunits is very low, only about 30% (Itoh
et al., 1988). Therefore,
duality
of GTH
was hypothesized.
However, only one type
of GTH was isolated
and characterized
from other species of fish, including
silver
carp (Burzawa-Gerard,
1974; Chang et al.,
1988a,b; Donaldson
er al., 1972; Farmer
and Papkoff,
1977; Huang et al., 1981;
Pierce et al., 1976). Such discrepancy may
be due to either species differences or differ-
ent methods used for GTH extraction
and
purification.
If two types of B subunits of
GTH are present in silver carp, then two
types of corresponding
cDNAs should be
expected. Because the probe we used for
30
CHANG ET AL.screening scGTHj3
was the cDNA encod-
ing cGTH+,
which is highly homologous to
the p subunit of salmon GTH II, therefore
only one type of cDNA encoding scGTH-P
was obtained.
In order to investigate
whether duality of GTH is also present in
silver carp or not, we are currently
re-
screening the cDNA library of silver carp
with other probes.
The amino acid sequence of scGTH-cr is
very similar to that of cGTH-ol, with a dif-
ference of 2 residues. In addition, scGTH-ol
bears a high extent of homology
(72%) to
that of the (Y subunit of mammalian
glyco-
protein hormones (Tables 2 and 4). The ex-
tent of homology between the cx subunit of
fish and mammalian
GTH is close to that
between the OL subunit of pituitary
glyco-
protein hormones from different species of
mammal.
Like the (Y subunit, homology
of
amino acid sequences between scGTH-P
and cGTH+
is extremely high, only differ-
ent in 4 residues. When scGTH$
is com-
pared with sGTH-l3,
the homology
in the
amino acid sequence decreased to 75% (Ta-
ble 4). The p subunit of Cyprinidae
GTH
(scGTH and cGTH) and Salmonidae
GTH
(sGTH) share a long homologous
region in
the middle part of the molecule
(Table 3).
However,
their N- and C-terminal
se-
quences are different
from each other.
These results reveal the phylogenetic
rela-
TABLE 2
THE AMINO ACID SEQUENCES OF THE a SUBUNIT OF SILVER CARP AND CARP GTH AND HUMAN AND BOVINE LH
scGTH cGTH hLH bLH scGTH cGTH hLH bLH scGTH cGTH hLH bLH scGTH cGTH hLH bLH scGTH cGTH hLH bLH scGTH cGTH hLH bLH scGTH cGTH hLH bLH Tyr-Pro-Arg-Asn-Asp- Be -Thr-Asn-Phe-Gly-Cys-Glu-Glu-Cys- - - - - _ Met Asn - - _ _ _ _ _ Val Gln Asp - Pro - - Phe - Asp Gly Glx Phe - Met Glu - - Pro Gin - Lys-Leu-Lys-Glu-Asn-Asn- Ile -Phe-Ser-Lys-Pro-Gly-Ala-Pro-
---
Thr - Gin - - Pro Phe - - Gin - - - - - - - Lys Tyr - - - - Asp - - Val-Tyr-Gln-Cys-Met-Gly-Cys-Cys-Phe-Ser-Arg-Ala-Tyr-Pro- -_-_--- Ile -L.g _ - - _ _ - - _ _ _ _ _ Ile _ - _ _ - - - _ - - - Thr-Pro-Leu-Arg-Ser-Lys-Lys-Thr-Met-Leu-Val-Pro-Lys-Asn- ---_--- --- Gln - - - - ~a--- Ile -Thr-Ser-Glu-Ala-Thr-Cys-Cys-Val-Ala-Lys-Glu-Val-Lys- ---
Val - - - Ser - - - Ser Tyr Asn - - - Glx - - - Ala Phe Thr Arg-Val-Leu-Val-Asn-Asp-Val-Lys-Leu-Val- X -Asn-His-Thr-
---
- - Thr - Met Gly Gly Phe Lys - Glu - - - Lys Ala Thr - Met Gly Asn Val Arg - Glx - - - Asp-Cys-His-Cys-Ser-Thr-Cys-Tyr-Tyr-His-Lys-Ser
---_-_-___
&a _-__ -___--_ Glx---e-m
Note. SC, silver carp; c, carp; h, human; b, bovine; -, residue identical to that of scGTH; X, gap insertion for half cystine alignment. References: cGTH from Chang et al. (19t38b); hLH from Fiddes and Talmadge (1984); bLH from Cornell and Pierce (1974).
TABLE 3
THE AMINO ACID SEQUENCES OF THE /3 SUBUNIT OF SILVER CARP, CARP, AND SALMON GTH AND BOVINE LH AND FSH
scGTH cGTH SGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH scGTH cGTH sGTH bLH bFSH Ser-Phe-Leu-Pro-Pro-Cys-Glu-Pro-Val-Asn-Glu- - Tyr - _ - _ - - - - _
- Leu Met Gln - - Gln - Ile - Gln Ser-Arg-Gly-Pro Leu Arg - Leu - Gln - Ile - Ala - - Leu Thr - Be Thr-Val-Ala-Val-Glu-Lys-Glu-Gly-Cys-Pro-Lys-Cys-Leu-Val-
---_---
- - Ser Leu - _ - _ - _ ti _ _ - - Leu - Ala - - - Ala - - Val - Be Thr - Ile Thr - - - - Glu - Gly Phe - Ile Ser Phe-Gln-Thr-Thr- Ile -Cys-Ser-Gly-His-Cys-Leu-Thr-Lys-Glu-
Leu---
Be Arg Ala Pro - - - Val - - - - Thr - Ser - - Ala - Tyr - Pro Ser Met Lys Ile Asn - - Trp - Ala - Tyr - Tyr - Arg Asp Pro-Val-Tyr-Lys-Ser-Pro-Phe-Ser-Thr-Val-Tyr-Gln-His-Val-
---
- _ phe _ - - - -
kc - Leu Pro Val Ile Leu Pro Pro Met Pro - Arg - Leu - - Arg Asp - Ala Arg Pro Asn Ile - Lys Thr Cys-Thr-Tyr-Arg-Asp-Val-Arg-Tyr-Glu-Thr-V~-~g-Leu-~o-
---
- - - Met Ile - - -
- - - His Glu Leu - Phe Ala Ser - - - - - - Phe Lys Glu Leu Val - - - - Lys Val - Asp-Cys-Pro-Pro-Gly-Val-Asp-Pro-His- Ile -Thr-Tyr-Pro-Val-
---
- - - - Trp Ser - - - Val - - - -
Gly - - - Met Val Ser Phe - - Gly - Ala His His Ala - Ser Leu Tyr - - - - Ala-Leu-Ser-Cys-Asp-Cys-Ser-Leu-Cys-Thr-Met-Asp-Thr-Ser-
- - - -
- - - - His -
- Thr Glu - His - Asp-Cys-Thr- Ile -Glu-Ser
- - - - _ - - - - - - - Gly Gly Pro Arg - - - Val Arg Gly Gln - Arg - Glu -Asp- Phe - Pro
- - - Asn - - - -
Gly Pro - Arg Leu Ser Ser Thr - Lys - Asp Ser Asp Ser Thr -Leu-Gln-Pro-Asp-Tyr-Cys-Met-Ser-
- _ - - Phe - - - - - - - Phe - Ile Thr Thr - - Leu Ala - Asp His - Gly - Ser Tyr - Ser Phe
- - - Leu
- - Val Leu Thr Asp-Gly-Asp-Met-Trp Pro Pro Leu Pro Asp Ile Leu Arg Glu Ile Lys Glu
Note. SC, silver carp; c, carp; s, salmon; b, bovine; -, residue identical to that of scGTH. References: cGTH from Chang et al. (1988b); sGTH from Trinh et al. (1986); bLH from Maurer (1985); bFSH from Esch et al. (1986).
32
CHANG ET AL.TABLE 4
HOMOLOGY ANALYSIS OF VERTEBRATE GTHs Subunit Type of GTH a scGTH vs cGTH scGTH vs bLH bLH vs hLH B scGTH vs cGTH scGTH vs sGTH scGTH vs bLH scGTH vs bFSH sGTH vs bLH sGTH vs bFSH bLH vs bFSH Homology (%) 98 72 74 97 75 42 40 41 35 38 Note. References: cGTH-a and cGTH+ from Chang et al. (1988b); bLH-a from Cornell and Pierce (1974); bLH+ from Maurer (1985); hLH-a from Fid- des and Tahnadge (1984); sGTH-B from Trinh et al. (1986); bFSH-B from Esch et al. (1986).
tionship of the (3 subunit of fish GTH. The
data of Tables 3 and 4 indicate that homol-
ogy between the (3 subunit of fish and mam-
malian GTH is around 40%, which is close
to that between the p subunit of LH and
FSH of a given species of mammal.
Al-
though the amino acid sequences of the p
subunit of fish and mammalian
GTHs are
diversified,
all of them contain
12 half-
cystines and these residues can be aligned
at the same positions.
The homology
analyses of the OL and /3
subunit between fish and mammalian
GTH
are summarized
in Table 4. These compar-
ative data gave further indication that the CY
subunit is conserved while the p subunit is
diversified
during the molecular
evolution
of vertebrate GTH.
ACKNOWLEDGMENTS
We express our particular appreciation to Mr. C. S. Chang for his invaluable amino acid determination. This work was supported by the National Science Council, ROC (NSC 77-0203-BOOl-07).
REFERENCES
Burzawa-Gerard, E. (1974). Purifcation d’une hor- mone gonadotrope hypophysaire de poisson tele- osteen, la carpe (Cyprinus carpio L.). Biochimie 53.545-552.
Chang, C. S., and Liu, C. S. (1988). A picomole-level amino acid analysis by means of gas-phase hy- drolysis and DABS-CYHPLC method. J. Chin. Biochem. Sot. 17, 12-19.
Chang, Y. S., Huang, F. L., Chen, C. T., and Lo, T. B. (1988a). Isolation and properties of the pi- tuitary gonadotropin from silver carp (Hypoph- thalmichthys molitrir). Znt. J. Pept. Protein Res. 31, 150-156.
Chang, Y. S., Huang, C. J., Huang, F. L., and Lo, T. B. (198813). The primary structures of carp go- nadotropin subunits deduced from cDNA nucle- otide sequences. Znt. J. Pept. Protein Res. 32, 5X-564.
Cornell, J. S., and Pierce, J. G. (1974). Studies on the disulfide bonds of glycoprotein hormones. .Z. Biol. Chem. 249,4166-4174.
Donaldson, E. M., Yamazaki, F., Dye, H. M., and Philleo, W. W. (1972). Preparation of gonadotro- pin from salmon (Oncorhynchus tshawytscha) pi- tuitary glands. Gen. Comp. Endocrinol. 18, 469- 481.
Esch, F. S., Mason, A. J., Cooksey, K., Nercads, M., and Shimasaki, S. (1986). Cloning and DNA se- quence analysis of the cDNA for the precusor of the g chain of bovine follicle stimulating hormone. Proc. Natl. Acad. Sci. USA 83, 6618-6621. Farmer, S. W., and Papkoff, H. (1977). A teleost (Ti-
lapia mossambica) gonadotropin that resembles luteinizing hormone. Life Sci. 20, 1227-1232. Fiddes, J. C., and Talmadge, K. (1984). Structure, ex-
pression and evolution of the genes for the human glycoprotein hormones. Recent Prog. Horm. Res. 40,43-78.
Greenwoods, F. C., Hunter, W. M., and Glover, J. S. (1963). The preparation of ‘311-labelled human growth hormone of high specific radioactivity. Biochem. J. 89, 116123.
Grunstein, M., and Hogness, D. (1975). Colony hy- bridization: A method for the isolation of cloned DNAs that contain a specific gene. Proc. Nutl. Acad. Sci. USA 12, 3%1-3%5.
Gubler, U., and Hoffman, B. J. (1983). A simple and very efficient method for generating cDNA li- brary. Gene 25, 263-269.
Hayashi, R. (1974). Carboxypeptidase Y in sequence determination of peptide. In “Methods in Enzy- mology” (C. H. W. Hirs and S. N. Timo- sheff, Eds.), Vol. 47, pp. 8+93. Academic Press, New York.
Hewick, R. M., Hunkapillar, M. W., Hood, L. E., and Dreyer, W. J. (1981). A gas-liquid solid phase peptide and protein sequenator. J. Biol. Chem. 256,799&7997.
Hunag, F. L., and Chang, Y. S. (1980). The gonado- tropic stimulation of androgen production on carp
testis in vitro. Proc. Natl. Sci. Count. Repub. China 4, 392400.
Huang, F. L., Huang, C. J., Lin, S. H., Lo, T. B., and Papkoff, H. (1981). Isolation and character- ization of gonadotropin isohormones from the pi- tuitary gland of pike eel (Muraenesox cinereus). Int. J. Pept. Protein Res. 18, 69-78.
Huang, F. L., Chang, Y. S., Chang, G. D., and Lo, T. B. (1982). Properties of subunits of pike eel gonadotropin. Proc. Natl. Sci. Count. Repub. China 6, 3&36.
Idler, D. R., and Ng, T. B. (1983). Teleost gonadotro- pins: Isolation, biochemistry and function. In “Fish Physiology” (W. S. Hoar and D. J. Ran- dell, Eds.), Vol. 9A, pp. 187-221. Academic Press, New York.
Itoh, H., Suzuki, K., and Kawauchi, H. (1988). The complete amino acid sequences of B-subunits of two distinct chum salmon GTHs. Gen. Camp. En- docrinol. 71,438-451.
Kidd, V. J., Wallace, R. B., Hakura, K., and Woo, S. L. C. (1983). a,-Antitrypsin deficiency detec- tion by direct analysis of the mutation in the gene. Nature (London) 304, 230-234.
Knecht, R., and Chang, J. Y. (1986). Liquid chro- matographic determination of amino acids after gas-phase hydrolysis and derivatization with (dimethylamino)azobenzenesulfonyl chloride. Anal. Chem. 58, 2375-2379.
Laemmli, U. K. (1970). Cleavage of structural protein during the assembly of the head of bacteriophage T+ Nature (London) 227, 68-5.
Liu, W. K., Nahm, S. H., Sweeney, C. M., Lamkin, W. M., Baker, H. N., and Ward, D. N. (1972). The primary structure of ovine luteinizing hor- mone. I. The amino acid sequence of the reduced and S-aminoethylated S-subunit (LH-a). J. Biol. Chem. 247, 43516364.
Lowry, 0. H., Rosebrough, N. J., Far, A. L., and Rondall, R. J. (1951). Protein measurement with the phenol reagent. J. Biol. Chem. 193, 265-275. Maurer, R. A. (1985). Analysis of several bovine lutropin B subunit cDNAs reveals heterogeneity in nucleotide sequence. J. Biol. Chem. 260,4684- 4687.
Maurer, R. A., Croyle, M. L., and Donelson, J. E. (1984). The sequence of a cloned cDNA for the B subunit of bovine thyrotropin predicts a protein containing both NH,- and COOH-terminal exten- sions. J. Biol. Chem. 259, 502&5027.
Pierce, J. G., Faith, M. R., and Donaldson, E. M. (1976). Antibodies to reduced S-carboxymeth- ylated alpha subunit of bovine luteinizing hor- mone and their application to study of the puriii- cation of gonadotropin from salmon (Onchohyn- thus tshawytscha) pituitary glands. Gen. Camp. Endocrinol. 30, 47-60.
Pierce, J. G., and Parsons, T. F. (1981). Glycoprotein hormones: Structure and function. Annu. Rev. Biochem. 50,465-495.
Rondle, C. J. M., and Morgan, W. T. J. (1955). The determination of glucosamine and galactosamine. Biochem. J. 61, 586-589.
Roser, J. F., Chang, Y. S., Papkoff, H., and Li, C. H. (1984). Development and characterization of a ho- mologous radioimmunoassay for equine prolactin. Proc. Exp. Biol. Med. 475, 510-517.
Sanger, F., Nicklen, S., and Coulson, A. R. (1977). DNA sequencing with chain-terminating inhibi- tors. Proc. Natl. Acad. Sci. USA 14, 5463-5467. Suzuki, K., Kawauchi, H., and Nagahama, Y. (1988).
Isolation and characterization of subunits from two distinct salmon gonadotropins. Gen. Camp. Endocrinol. 71, 302-306.
Trinh, K. Y., Wong, N. C., Hew, C. L., and Crim, L. W. (1986). Molecular cloning and sequencing of salmon gonadotropin B subunit. Eur. J. Bio- them. 159, 619-624.
Vaitukaitis, J. L., Roffms, J. B., Nieschlag, E., and Ross, G. T. (1971). A method of producing spe- chic antisera with small dose of hnmunogen. J. Clin. Endocrinol. Metab. 33, 988-991.
Ullrich, A., Shine, J., Chirgwin, J., Pictet, R., Tis- cher, E., Rutter, W. J., and Goodman, H. M. (1977). Rat insulin genes: Construction of plas- mids containing the coding sequences. Science
l%, 1313-1319.
Warren, L. (1959). The thiobarbituric acid assay of sialic acid. J. Biol. Chem. 234, 1971-1975. Winzler, R. J. (1955). Determination of serum glyco-