~bur~,rof
Biomedical
Science
Original Paper
J Biomed Sci 2003;10:219-227 DOt: 10.1159/000068711 Received: Juty 3, 2002 Accepted: October 16, 2002Green Tea Constituent (-)-Epigallocatechin-
3-Gallate Inhibits Hep G2 Cell Proliferation and
Induces Apoptosis through p53-Dependent and
Fas-Mediated Pathways
P o - L i n K u o C h u n - C h i n g Lin
Graduate Institute of Natural Products, Kaohsiung Medical University, Kaohsiung, Taiwan, ROC
Key Words
EGCG, p53. p21/WAF1 • Fas/APO-1 • Fas ligand, apoptosis
Abstract
(-)-Epigallocatechin-3-gallate (EGCG) is a polyphenolic compound found in green tea. It has been reported to possess a wide range of pharmacological properties, and is one of the most promising chemopreventive agents for cancer. To provide a better understanding of the preventive effect of EGCG on liver cancer, we exam- ined EGCG for its effect on proliferation and cell cycle progression in a human liver cancer cell line, Hep G2. The results showed that EGCG inhibited the prolifera- tion of Hep G2 by inducing apoptosis and blocking cell cycle progression in the G1 phase. ELISA showed that EGCG significantly increased the expression of p53 and p21/WAF1 protein, and this contributed to cell cycle arrest. An enhancement in Fas/APO-1 and its two form ligands, membrane-bound Fas ligand (mFasL) and solu- ble Fas ligand (sFasL), as well as Bax protein, was responsible for the apoptotic effect induced by EGCG. Taken together, our study suggests that the induction of
p53 and the activity of the Fas/FasL apoptotic system play major roles in the antiproliferative activity of EGCG in Hep G2 cells.
Copyright © 2003 National Science Council, ROC and S. Karger AG, Basel
Introduction
Recently, natural herbs and pure compounds that are present in the human diet have attracted extensive atten- tion as cancer chemopreventive agents. Green tea
(Ca-
mellia sinensis)
has been demonstrated to display cancer chemopreventive effects in different systems due to its striking inhibition of diverse cellular events related to cancer development [5, 40]. These effects are associated with antioxidative properties, blocking angiogenesis, car- cinogenesis, metastasis and proliferation of tumor cells [1, 6, 11, 15, 39, 43]. The main active constituents of green tea are polyphenolic compounds, which consist of (-)-epigallocatechin-3-gallate (EGCG), (-)-epigallocate- chin (EGC), (-)-epicatechin-3-gallate (ECG) and (-)-epi- gallocatechin (EC) [2]. Among them, EGCG is the most abundant ingredient. Many studies have reported that EGCG possesses direct tumor growth inhibition ability inKA KG E K
Fax +41 61 306 t2 34 E-Mail [email protected] "~e.karger.com© 2003 National Science Council, ROC S. Karger AG, Basel
1021-7770/03/0102-0219519.50 Accessible online at:
www.karger.com/jbs
Dr. Chun-Cbing Lin
Graduate Institute of NaturaI Products, Kaohsiung Medical University No. 100, Shin-Chuan tst Road
Kaohsiung 807, Taiwan (ROC)
several types of tumor cells, including h u m a n liver cancer [8, 26, 31, 32, 37], which is one of most common and the most lethal malignant diseases in the world [25]. How- ever, the precise antitumorigenic mechanisms of EGCG in hepatoma cells remain largely unknown. In this study, we determined the antiproliferative activity of EGCG in the h u m a n liver cancer cell line, Hep G2, and examined its effect on cell cycle distribution and apoptosis in Hep G2 cells. Furthermore, to establish the anticancer effects of EGCG, we assayed the levels of p53, p21/WAF 1, Fas ligand and Fas/APO- 1 receptor, which are strongly associ- ated with the signal transduction pathway of apoptosis and affect the chemosensitivity of tumor cells to antican- cer agents.
Materials and M e t h o d s
Materials
Fetal calf serum (FCS), penicillin G, streptomycin and amphoter- icin B were obtained from Gibco BRL (Gaithersburg, Md., USA). EGCG, dimethyl sulfoxide (DMSO), Dulbecco's modified Eagle's medium (DMEM), ribonuclease (RNase), and propidium iodide (PI) were purchased from Sigma (St. Louis, Mo., USA). XTT and p53 pan ELISA kits were obtained from Roche Diagnostics GmbH (Germa- ny). Nucteosome ELISA, WAF1 ELISA and Fas/APO-t ELISA kits were purchased from Calbiochem (Cambridge, Mass., USA).
Preparation of EGCG
EGCG was dissolved in DMSO and stored at - 2 0 ° C . For all experiments, final concentrations of the tested compound were pre- pared by diluting the stock with DMEM (Sigma). Control cultures received the carrier solvent (0.1% DMSO).
Ceil Line and Culture
Hep G2 cells [American Type Culture Collection (ATCC) HB8065] were maintained in DMEM supplemented with 10% FCS, 100 units/ml of penicillin G, 100 gg/ml of streptomycin, and 0.25 gg/ml of amphotericin B in a CO2-air mixture.
Assay for Cell Proliferation Inhibition
Cell proliferation inhibition was assessed by XTT assay [sodium 3"-(1-(phenylamino-carbonyl)-3,4-tetrazolium)-bis(4-methoxy-6-ni- tro) benzene-sulfoic acid hydrate]. Briefly, the cell line was subcul- tured into a 96-well plate with 1 x t04 cells per well in 100 gl medium. After 24 h of incubation, the medium in the 96-well plate was discarded and replaced by 90 gl of DMEM, and 10 gl of EGCG was then added to each well to final concentrations of 10, 50, 100, and 200 g M each in triplicate. After the addition of the sample, the plates were incubated in a 37 ° C humidified incubator with 5% CO2 for 12, 24, 48, and 72 h. At the end of the incubation, the medium containing the various concentrations of EGCG was discarded. The cells were then washed once with PBS. Fifty microliters of XTT test solution was prepared by mixing 5 ml of XTT-labeling reagent, and 100 ~tl of electron-coupling reagent was then added to each well. After 6 h of incubation at 37 °C in a 5 % CO2 incubator, the absorbance of test samples was measured with an ELISA reader (Multiskan EX,
Labsystems) at a test wavelength of 492 nm and a reference wave- length of 690 nm [12, 27].
Assay for Cell Cycle Distribution
Hep G2 cells were cultured in triplicate in 6-well plates at a con- centration of 5 x 105/well. After 24 h of incubation, cells were treated with 50, 100, and 200 g M EGCG were analyzed in paralM with cells grown in the absence of green polyphenol compounds to determine the effects on cell cycle distribution. Cells were collected and fixed with 70% ethanol. Cell pellets were suspended in 2 gl of 10 gg/ml RNase containing 0.5% Triton (J.T. Baker, Inc.) plus the same volume of 20 gg/ml PI (Sigma), followed by incubation in the dark at room temperature for 30 rain. Celt suspensions were filtered through a 60-gin mesh filter (Spectrum Medical Industries, Calif., USA). Data acquisition and analysis were performed on an EPICS flow cytometer (Coulter Electronics), and data from 10,000 cells were collected for each data file. Cell cycle analysis was performed with Multicycle software (Phoenix Flow Systems, San Diego, Calif., USA) [30].
Assay for Apoptosis by Nucleosome ELISA
For the quantification of apoptotic cells, we used the Nucleosome ELISA kit, operated according to the manufacturer's protocol. Brief- ly, Hep G2 cells were treated with 50, 100, and 200 g M o f E G C G for 6, 12, 24, and 48 h. Samples of the cell lysate and the biotinylated detector antibody were placed (triplicated) in the 96-well plate that were coated DNA binding protein, with final cell counts of 1 x 106 per well. The assayed mono- and oligonucleosomes were captured on plates that were pre-coated with DNA binding proteins. Antihistones 3 (H3) biotin-labeled antibody was then bound to the histone compo- nent of captured nucleosomes and was detected following incubation with streptavidin-linked horseradish peroxidase (SA-HRP) conju- gate. HRP catalyzed the conversion of colorless tetramethylbenzid- ine to blue. The addition of stop solution changed the color to yellow, the intensity of which was proportional to the number of nucleo- somes in the sample. Absorbance was measured with an ELISA read- er at a test wavelength of 450 nm. By comparing the absorbance obtained from a sample containing an unknown amount of nucleo- somes with that obtained from the standards, one can assign a nucleosome unit value to the unknown sample. In addition, one can assign an apoptotic index (ratio) to an unknown sample when the absorbance of the treated (apoptotic) sample is divided by that of the untreated (control) sample [18, 38].
ELISA Assay of p53, p21/WAF1, Fas Ligand and Fas/APO-1 Receptor
For the detection of p53, p21/WAF1, Fas ligand and Fas/APO-1 receptor, we used the p53 pan ELISA, WAF1 ELISA, Fas Ligand and Fas/APO-t ELISA kits. Briefly, Hep G2 cells were treated with 50, 100, and 200 gM of EGCG for 6, 12, 24, and 48 h. The samples of cell lysate and the biotinylated specific detector antibody were placed (triplicated) in 96-well microtiter plates coated with monoclonal anti- bodies with final cells counts of 1 x 106 per well, and were incubated for 1 h (Fas/APO-1), 2 h (p53 or p21/WAF1) or 3 h (FasL) at room temperature. Particularly, the soluble Fas ligand in the cell cnlture supernatant also needed to be determined by using an enzyme-linked immunosorbent assay (Fas Ligand ELISA). After removing the unbound material by washing with PBS, horseradish-peroxidase- conjugated streptavidin was added to bind to the antibodies. Horse- radish peroxidase catalyzed the conversion of chromogenic substrate
(tetramethylbenzidine) to a colored solution with a color intensity proportional to the amount of protein (p53, p21/WAF1, Fas L or Fas/APO-1) present in the sample. The absorbance of each well was measured at 450 nm and concentrations of p53, p21/WAF1, FasL, and Fas/APO-1 were determined by interpolating from standard curves obtained with known concentrations of protein [7, 22, 35,
36].
Assay for Western Blotting
Cells treated with 50, 100, and 200 p5~/EGCG for 24 h were lysed and the protein concentration was determined by using a Bio-Rad Protein Assay (Bio-Rad Laboratories, Richmond, Calif, USA). For Western blotting, 50-100 gg of total ceil lysates were subjected to SDS-PAGE. The protein was transferred to PVDF membranes using transfer buffer (50 m M Tris, 190 m M glycin, and 10% methanol) at 100 V for 2 h. The membranes were incubated with blocking buffer (50 m M Tris, 200 m M NaC1, 0.2% Tween 20, and 3% BSA) over- night at 4 ° C. After washing three times with washing buffer (block- ing buffer without 3% BSA) for 10 rain each, the blot was incubated with Bcl-2 or Bax antibody for 2-15 h, followed by horseradish per- oxidase-labeled secondary antibody for 1 h. The membranes were washed again, and detection was performed using the enhanced chemiluminescence Western blotting detection system (Amersham, USA) [301.
Results
Effect of EGCG on Hep G2 Celt Proliferation
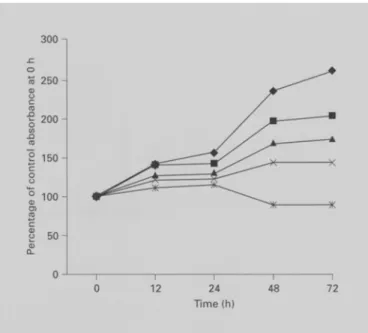
We first tested the antiproliferative effect of EGCG in a liver cancer cell line, Hep G2. As shown in figure 1, the growth-inhibitory effect of EGCG was observed in a dose- dependent manner and with an ICs0 vaiue of 147.26 + 5.3 gM.
EGCG-Induced Cell Cycle Arrest and Apoptosis in Hep G2 Cells
The effect of EGCG on cell cycle progression of Hep G2 was determined by flow cytometric analysis 24 h after its addition. As shown in figure 2, the results indicated that, compared with the control, EGCG increased the population in the G1 phase. At the dose of 100 gM EGCG, the percentage of cells in the G 1 phase increased from 33.8% to 57.2%, and those in the G2 phase de- creased from 34.3 to 11.9%. These effects were enhanced when Hep G2 cells were treated with 200 gM EGCG. Our data suggest that EGCG arrests the cell cycle of Hep G2 cells at the G1 phase.
Apoptosis is an active process of cell death. It is charac- terized by nuclear condensation, DNA fragmentation and blebbing of the plasma membrane. Activation of endonu- cleases during the process of apoptosis leads to the frag- mentation ofchromatin into about 180-bp oligonucleoso- real fragments, with subsequent releases of nucleosomes
300 - o "~ 2 5 0 -
~
2 0 0 - "~ 1 5 0 - g ~5 100 g 0 J f u __ I 0 12 24 4 8 7 2 Time (h)Fig. 1. Growth inhibition of Hep G2 cells by EGCG. Adherent cells that proliferated in 96-well plates (104 cells/well) were incubated with different concentrations of EGCG for various time intervals. Cell proliferation was determined by XTT assay. Results are expressed as percent of cell proliferation of control at 0 h. The data shown are the mean from three independent experiments each with triplicate wells. Standard deviations are less than 10 %.
into the cytoplasm. By contrast, necrotic cell death often occurs with extensive tissue damage, resulting in an intense inflammatory response. The main characteristic of necrosis is the swelling oforganelles and cells. Later, the cell loses membrane integrity and releases randomly di- gested DNA fragments. Thus, the quantification of nu- cleosomes in the cytoplasm by a nucleosome ELISA assay can precisely measure apoptotic response [28].
Figure 3 shows the time course of DNA fragmentation in continuous treatment with 50, 100, and 200 gM EGCG. DNA fragmentation of Hep G2 was noted at 6 h and maximized at 48 h after addition of EGCG. In con- trast to the controls, when cells were treated with higher concentrations of EGCG, the number of cells undergoing apoptosis was further enhanced from about 5-fold at
100 g M E G C G to 9-fold at 200 gMat 48 h.
EGCG Increases the Expression of p53 and p21/WAF1 Proteins in Hep G2 Cells
To determine whether tumor suppression factor p53 is involved in the EGCG-mediated antiproliferation of Hep G2 cells, three concentrations of EGCG (50, 100, and 200 gM) were added to Hep G2 cells, then the levels of
EGCG Inhibits Hep G2 Cell Proliferation and Induces Apoptosis
Control
+Aggregates
Multicycle suggestions (a guideline only): No abnormal DNA content is observed The diploid %S = 31.9, %G2 = 34.3 ~¢ The S phase confidence is good
0 64 128 192 256 320 384 448 512
DNA content EGCG/50
+Aggregates
Multicycle suggestions (a guideline only): No abnormal DNA content is observed The diploid %S = 30,6, %G2 = 19,5 The S phase confidence is good
0 64 128 192 256 320 384 448 512
DNA content EGCG/100
4oo j +Aggregates
35o Multlcycle suggestions (a guideline only): 320 No abnormal DNA content is observed
The diploid %S = 30,9 %G2 = 11.9, 280 ~The S phase confidence is good
240 o~ ~ 200 120
°oj
'oi
0 64 128 192 256 320 384 448 512 DNA content 270 240 210 ¢o 180 120 SO 4 60 4 30 4 EGCG/200 +AggregatesMulticycle suggestions (a guideline only): No abnormal DNA content is observed The diploid %S = 26.7, %G2 = 5,4 ;~The S phase confidence is good
0 64 ~28 192 256 320 384 448 512
F i g . 3 . Induction of apoptosis in Hep G2 by EGCG. Hep G2 cells were cultured with 50, 100, and 200 pM EGCG for 6, 12, 24, and 48 h. Cells were harvested and lysed with lysis buffer. Cell lysates that contained cytoplasmic oligonucleosomes of apoptotic cells were ana- lyzed in the Nucleosome ELISA. The data shown are the mean + SD of three independent experiments each with triplicate wells. The asterisk indicates a significant difference between control and EGCG-treated cells, * p < 0.05.
Fig. 4. Effects of EGCG on protein expression of p53 (a) and p21/ WAF1 (b) in Hep G2 cells. Hepatoma cells were treated with 50, 100, and 200 gM EGCG. Lysates were prepared from these cells and p53 and p21/WAFt levels were determined by p53 pan ELISA and WAF 1 ELISA kits, respectively. The detailed protocol is described in 'Materials and Methods'. The data shown are the mean -+ SD of three independent experiments each with triplicate wells. The aster- isk indicates a significant difference between control and EGCG- treated cells, * p < 0.05. 800 700 600 ,-$ 500 o ..~ 400 _~ 300 CrJ D. 200 100 0 F-n0 10 b e- _~ 5 > 4 so 1oo 12 2o0 24 "time (h) 48 6 12 24 48 Time (h)
p53 protein were assayed b y ELISA. M a r k e d induction o f p53 p r o t e i n was o b s e r v e d in a d o s e - d e p e n d e n t m a n n e r (fig. 4a). T h e upregulation o f p53 b y E G C G started to increase 6 h after t r e a t m e n t with E G C G , a n d r e a c h e d m a x i m u m expression at 12 h.
Fig. 2. Inhibition of cell cycle progress in Hep G2 cells by treating with EGCG. Cell cycle analysis of Hep G2 cells following treatment with 50, 100, and 200 gM EGCG for 24 h. Cells were fixed and stained with propidium iodide. Cell cycle distribution was analyzed by flow cytometry.
p 2 1 / W A F I protein is first characterized as a down- s t r e a m target o f p53, a n d is t h o u g h t to be responsible for G1 cell cycle arrest [21]. Thus, we also assessed the p 2 t / WAF1 expression o f p53-expressing H e p G 2 cells using a WAF1 E L I S A assay. Figure 4b shows that an increase in p 2 1 / W A F 1 protein was a p p a r e n t at 6 h a n d reached m a x i - m u m induction at 24 h in E G C G - t r e a t e d H e p G2 cells. M o r e o v e r , p 2 1 / W A F 1 was induced in a d o s e - d e p e n d e n t m a n n e r . Based o n these data, we suggest that E G C G - m e d i a t e d cell cycle arrest operates t h r o u g h the induction o f p 2 1 / W A F 1 protein in a p 5 3 - d e p e n d e n t event in H e p G 2 cells.
EGCG Inhibits Hep G2 Cell Proliferation and Induces Apoptosis
D 0 16-~a 14- -~ 12 o= a v 6 u_ 4 5o 100 12 Time (h) 200 24 48 C~ E l O ~ b 9 8 7 6 5 4 3 2 1 0 10 ¢ 9 8 7 6 5 4 3 2 1 0 6 12 24- 48 ]]me (h) 6 12 24 48 .time (h)
Fas/FasL System Is an Essential Pathway o f EGCG-Mediated Apoptosis
Recent studies have demonstrated that the Fas/FasL system is an important mediator of the apoptotic process, particularly in chemotherapy-induced cancer cell apopto- sis [16, 24]. The apoptotic effect of a chemotherapy agent acts by upregutating the expression of either the Fas/ APO-1 receptor or Fas ligand (FasL) [23]. Therefore, we wondered whether the levels of Fas/APO-1 and FasL in Hep G2 cells were modulated by EGCG. First, the level of Fas/APO-1 expression was measured after the treatment of Hep G2 cells with 50, 100, and 200 g M o f E G C G for 6, 12, 24, and 48 h. As shown in figure 5a, the expression of Fas/APO-1 increased following EGCG treatment. This change occurred at 6 h and reached its peak at 24 h.
Recent reports have indicated that both soluble Fas ligand (sFasL) and membrane-bound Fas ligand (mFasL) might be involved in the activation of the Fas/FasL sys- tem [29, 34]. Thus, we also assayed the alteration of sFasL and mFasL in Hep G2 cells with treatments of 50, 100, and 200 gMEGCG. At the mFasL level, ligand accumula- tion appeared 6 h after EGCG treatment and progressive- ly increased up to 24 h (fig. 5b). A similar result was observed for sFasL. Six hours after the addition of EGCG, sFasL was significantly induced and remained at a high level for 12-24 h (fig. 5c). However, the increase in mFasL with EGCG was greater than the increase in sFasL at 24 h (30-fold vs. 9-fold in Hep G2 cells treated with 200 gM EGCG ). Interestingly, the induction of Fas/ APO-1, mFasL as well as sFasL showed slight differences between cells treated with 50 and 100 g M EGCG, but the differences greatly increased when Hep G2 cells were treated with 200 g M EGCG.
EGCG Treatment o f Hep G2 Cells' Results in an Increase in Proapoptotic Protein Bax
Bcl-2 family proteins are important regulators in the apoptotis signaling pathway. These proteins, including Bax and Bcl-2, are regulated by p53 [21]. Here, we exam-
F i g . 5. The Fas/FasL system was involved in EGCG-mediated apo- topsis. Hepatoma cells were incubated with 50, 100, and 200 g M EGCG for the time indicated, a Amount of Fas/APO-1 receptor in Hep G2 cells, b Amount of mFasL in Hep G2 cells, c Amount of sFasL in Hep G2 cells. The data shown are the mean -+ SD of three independent experiments each with triplicate wells. The asterisk indicates a significant difference between control and EGCG-treated cells, * p < 0.05.
ined the effect of EGCG on Bax and Bcl-2 protein expres- sion (fig. 6). After 24 h of exposure to 50, 100, and 200 p M of EGCG, the amount of Bax protein increased in a dose- dependent manner. In contrast, we were unable to detect Bcl-2 protein expression in Hep G2 cells in the absence or presence of EGCG. This result was similar to that of a previous report which indicated that Hep G2 cells did not express Bcl-2 [33].
Discussion
In recent years, natural antioxidants that are present in food and beverages consumed by humans have earned extensive attention because of their cancer chemopreven- tive effects. EGCG, a potyphenolic compound present in green tea, is a potent antioxidant with tumor-preventive effects [1, 5, 40]. Many studies have demonstrated that EGCG has an important antiproliferative effect against certain cancers, including lung, bronchial, colon, prostate, breast, liver and leukemia cancer cells [2, 11, 15, 32, 39, 43]. The mechanism of action of EGCG on tumor cell growth inhibition is related to the induction of apoptosis and the blockade of cell cycle progression. EGCG inhibit- ed the activities of several key G1 regulation proteins by arresting cells in the G1 phase [19]. EGCG can also induce apoptosis by several molecular mechanisms, in- cluding inhibition of growth-related signal transduction pathways that lead to the activation of an important tran- scription factor activator protein (AP-1) [10, 20, 42], trig- gering apoptosis by directly binding to Fas receptor [13], and blocking the activity of topoisomerase I [3]. More- over, a recent study showed that the synergistic cancer- preventive effects occurred when EGCG was combined with EGC, sulindac or tamoxifen [32]. In contrast to its antioxidant effect, EGCG may also act as a prooxidant through H202 production to induce apoptosis in human lung cancer H661 cells [41, 42]. In the study by Uesato et al. [37], EGCG exhibited cell growth inhibition in hepato- ma cell lines, but the precise mechanism against liver can- cer was not reported. In our study, we demonstrated that ECGC restores Hep G2 cell apoptosis sensitivity and arrests cells in the G1 phase. Moreover, this effect was found to be mediated through a p53-dependent- and Fas/ FasL-system-mediated ceil death pathway.
Normal p53 function has been found to play a crucial rote in the induction of apoptosis and triggering of cell cycle checkpoints in human and murine cells following DNA damage [21 ]. This notion has further been support- ed by findings that p53 is the most commonly mutated
0 50 100 200 pM
Fig. 6. Effect of EGCG on protein expression of Bax. As detailed in 'Materials and Methods', the cells were treated with 50, 100, and 200 p M of EGCG. The cells were harvested after 24 h. Total cell lysates were separated, and 50 gg of proteins were separated via SDS- PAGE followed by Western blot analysis and chemilmninescence detection.
tumor suppressor gene and a lacking p53 function is relat- ed to an enhanced risk of carcinogenesis [9]. Moreover, the chemosensitivity of cancer cells to chemotherapy agents is greatly influenced when the function of p53 is abrogated [4]. Our results indicated that p53 plays a key role in EGCG-mediated antiproliferation in Hep G2 cells. p53 cannot only arrest the cell cycle in the G1 phase, but also induce apoptosis. This hypothesis is supported by the following results: (a) Flow cytometric analysis indicated that EGCG could arrest Hep G2 cells in the G1 phase. The blockade of cell cycle progression was attributed to the amount of enhancement of p21/WAF 1 protein that is induced in a p53-dependent manner. A similar result was found in EGCG-treated breast carcinoma cells [19]. (b) Both proapoptotic downstream targets of p53, Fas/ APO-1 and Bax proteins were increased by EGCG. More- over, the expression levels of these proteins were greatly increased (at 24 h) after maximal accumulation of p53 protein (at 12 h) in Hep G2 cells.
The Fas/FasL system is a key signaling transduction pathway of apoptosis in cells and tissues. Binding of Fas ligand to Fas/APO-1 induces receptor oligomerization and formation of death-inducing signaling complex (DISC), followed by activation of a series caspase cas- cades resulting in apoptotic celt death [ 14]. FasL is a TNF- related type II membrane protein. Cleavage of mem- brane-bound Fas ligand (mFasL) by a metalloprotease- like enzyme results in the formation of soluble Fas ligand (sFasL) [t7]. Both mFasL and sFasL can bind to Fas/ APO- 1, and subsequently trigger the Fas/FasL system, but sFasL has been reported to be a weaker inducer of apopto- sis than mFasL [29]. Our study indicated that both Fas ligands, mFasL and sFasL, were increased in EGCG- treated hepatoma cells. Moreover, the level of Fas/APO-1
EGCG Inhibits Hep G2 Cell Proliferation and Induces Apoptosis
was simultaneously enhanced in FasL-upexpressing Hep G2 cells. Thus, we demonstrated that the Fas/FasL system participates in EGCG-mediated apoptosis.
In summary, our study demonstrates that EGCG can effectively inhibit Hep G2 cells proliferation by two dif- ferent cellular responses. Firstly, EGCG can block celt cycle progression and cause a G1 arrest via p21/WAF1 activation. Secondly, the apoptotic sensitivity of Hep G2 cells was restored by a p53- and Fas/FasL-dependent
pathway. Our study has clearly demonstrated that EGCG might be a promising chemopreventive agent for liver cancer, even when absorbed solely by means of diet.
Acknowledgements
This study was supported by a research grant from the National Science Council of Taiwan (NSC 90-2320-B-037-021).
References
1 Anderson RF, Fisher LJ, Hara Y, Harris T, Mak WB, Melton LD, Packer JE. Green tea catechins partially protect DNA from (.)OH radical-induced strand breaks and base damage through fast chemical repair of DNA radicals. Carcinogenesis 22:1189-1193;200t.
2 Balentine DA, Wiseman SA, Bouwens LC. The chemistry of tea flavonoids. Crit Rev Food Sci Nutr 37:693-704; 1997.
3 Berger SJ, GuNa S, Belfi CA, Gosky DM, Mukhtar H. Green tea constituent (-)-epigallo- catechin-3-gallate inhibits topoisomerase I ac- tivity in human colon carcinoma cells. Bio- chem Biophys Res Commun 288:101-105; 2001.
4 Brown JM, Wouters BG. Apoptosis, p53, and tumor cell sensitivity to anticancer agents. Can- cer Res 59:1391-1399; 1999.
5 Bushman JL. Green tea and cancer in humans: A review of the literature. Nutr Cancer 31:151 -
159;1998.
6 Cao Y, Cao R. Angiogenesis inhibited by drinking tea. Nature 398:381; 1999.
7 Castaneda F, Kinne RK. Apoptosis induced in HepG2 cells by short exposure to millimolar concentrations of ethanol involves the Fas- receptor pathway. J Cancer Res Cin Oncol 127: 418-424;2001.
8 Chen ZP, Schell JB, Ho CT, Chen KY. Green tea epigallocatechin gallate shows a pro- nounced growth inhibitory effect on cancerous cells but not on their normal counterparts. Can- cer Lett 129:173-179; 1998.
9 Donehower LA, Harvey M, Slagle BL, McAr- thur M J, Montgomery CA Jr, Butel JS, Bradley A. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tu- mours. Nature 356:215-221;1992.
10 Dong Z, Ma W, Huang C, Yang CS. Inlaibition of tumor promoter-induced activator protein 1 activation and cell transformation by tea 13o13,- phenols, (-)-epigallocatechin gallate, and thea- flavins. Cancer Res 57:4414-4419; 1997. 11 Garbisa S, Sartor L, Biggin S, Salvato B, Benelli
R, Albini A. Tumor getatinases and invasion inhibited by the green tea flavanol epigallocate- chin-3-galIate. Cancer 91:822-832;2001.
12 Goodwin CJ, Holt SJ, Downes S, Marshall NJ. Microculture tetrazolium assays: A comparison between two new tetrazolium salts, XTT and MTS. J Immunol Methods 179:95-103; 1995. 13 Hayakawa S, Saeki K, Sazuka M, Suzuki Y,
Shoji Y, Ohta T, Kaji K, Yuo A, Isemura M. Apoptosis induction by epigallocatechin gallate involves its binding to Fas. Biochem Biophys Res Commun 285:1102-1106;2001.
14 Hengartner MO. The biochemistry of apopto- sis. Nature 407:770-776;2000.
15 Hong J, Smith T J, Ho CT, August DA, Yang CS. Effects &purified green and black tea poly- phenols on cyclooxygenase- and lipoxygenase- dependent metabolism of arachidonic acid in human colon mucosa and colon tumor tissues. Biochem Pharmaco162:1175-1183;2001. 16 Jiang S, Song MJ, Shin EC, Lee MO, Kim SJ,
Park JH. Apoptosis in human hepatoma cell lines by chemotherapeutic drugs via Fas-de- pendent and Fas-independent pathways. Hepa- tology 29:101-110; 1999.
17 Kayagaki N, Kawasaki A, Ebata T, Ohmoto H, Ikeda S, Inoue S, Yoshino K, Okumura K, Ya- gita H. Metalloproteinase-mediated release of human Fas ligand. J Exp Med 182:1777-1783; 1995.
18 Kikuchi S, Hiraide H, Tamakuma S, Yamamo- to M. Expression of wild-type p53 tumor sup- pressor gene and its possible involvement in the apoptosis of thyroid tumors. Jpn J Surg 27: 226-233;1997.
19 Liang YC, Lin-Shiau SY, Chen CF, Lin JK. Inhibition of cyclin-dependent kinases 2 and 4 activities as well as induction ofCdk inhibitors p21 and p27 during growth arrest of human breast carcinoma cells by (-)-epigallocatechin- 3-gallate. J Cell Biochem 75:1 - 12; 1999. 20 Lin YL, Lin JK. (-)-Epigallocatechin-3-gallate
blocks the induction of nitric oxide synthase by down-regulating lipopolysaccharide-induced activity of transcription factor nuclear factor- kappa B. Mol Pharmaco152:465-472; 1997. 21 May P, May E. Twenty years of p53 research.
structural and functional aspects of the p53 protein. Oncogene 18:7621-7636; 1999. 22 Mediavilla MD, Cos S, Sanchez-Bareeto EJ.
Melatonin increases p53 and p21WAF1 ex- pression in MCF-7 human breast cancer cells in vitro. Life Sci 65:415-420;1999,
23 Muller M, Strand S, Hug H, Heinemann EM, Walczak H. Drug-induced apoptosis in hepato- ma cells is mediated by the CD95 (APO-1/Fas) receptor/ligand system and involves activation of wild-type p53. J Clin Invest 99:403-413; 1997.
24 Nagata S, Golstein P. The Fas death factor. Science 267:1449-1456; 1995.
25 Okuda K. HepatocelMar carcinoma. Recent progress. Hepatology 15:948-963; 1992. 26 Paschka AG, Butler R, Young CY. Induction
of apoptosis in prostate cancer cell lines by the green tea component, (-)-epigallocatechin-3- gallate. Cancer Lett 130:1-7; 1998.
27 Roehm NW, Rodgers GH, Hatfield SM, Glase- brook AL. An improved colorimetric assay for cell proliferation and viability utilizing the te- trazolium salt XTT. J Immunol Methods 142: 257-265;1991.
28 Salgame P, Varadhachary AS, Primiano LL, Fincke JE, Muller S, Monestier M. An ELISA for detection of apoptosis. Nucleic Acids Res 25:680-681; 1997.
29 Schneider P, Holler N, Bodmer JL, Halme M, Frei K, Fontana A, Tschopp J. Conversion of membrane-bound Fas(CD95) ligand to its solu- ble form is associated with downregulation of its proapoptotic activity and loss of liver toxici- ty. J Exp Med 187:1205-1213;1998.
30 Shirin H, Sordillo EM, Oh SH, Yamamoto H, Delohery T, Weinstein IB, Moss SF. Helico- bacter pylori inhibits the G1 to S transition in AGS ~stric epithelial cells. Cancer Res 59: 2277-2281;1999.
31 Smith DM, Dou QP. Green tea polyphenol epi- gallocatechin inhibits DNA replication and consequentIy induces leukemia cell apoptosis. Int J Mol Med 7:645-652;2001.
32 Suganuma M, Okabe S, Kai Y, Sueoka N, Suenka E, Fujiki H. Synergistic effects of (-)- epigallocatechin gallate with (-)-epicatechin, sulindac, or tamoxifen on cancer-preventive activity in the human tung cancer cell line PC- 9. Cancer Res 59:44-47; 1999.
33 Takehara T, Liu X, Fujimoto J, Friedman SL, Takahashi H. Expression and role of Bcl-xL in human hepatocellular carcinomas. Hepatology 34:55-61 ;2001.
34 Tanaka M, Suda T, Takahashi T, Nagata S. Expression of the functional solubIe form of human fas ligand in activated tymphocytes. EMBOJ 14:1129-1135;1995.
35 Tarkowski E, Rosengren L, Blomstrand C, Jensen C, Ekholm S, Tarkowski A. Intrathecal expression of proteins regulating apoptosis in acute stroke. Stroke 30:321-327; 1999. 36 Tendler Y, Weisinger G, Coleman R, Diamond
E, Lischinsky S, Kerner H, Rotter V, Zinder O. Tissue-specific p53 expression in the nervous system. Brain Res Mol Brain Res 72:40-46; 1999.
37 Uesato S, Kitagawa Y, Kamishimoto M, Ku- magai A, Hori H, Nagasawa H. Inhibition of green tea catechins against the growth of can- cerous human colon and hepatic epithelial cells. Cancer Lett 170:41-44;2001.
38 Wang L, MaW, Markovich R, Chen JW, Wang PH. Regulation of cardiomyocyte apoptotic signaling by insulin-like growth factor I. Circ Res 83:516-522;1998.
39 Yamane T, Takahashi T, Kuwata K, Oya K, Inagake M, Kitao Y, Suganuma M, Fujiki H. Inhibition of N-methyl-N'-nitro-N-nitroso- guanidine-induced carcinogenesis by (-)-epi- gallocatechin gallate in the rat glandular stom- ach. Cancer Res 55:2081-2084;1995.
40 Yang CS, Chung JY, Yang G, Chhabra SK, Lee MJ. Tea and tea polyphenols in cancer pre- vention. J Nutr 130:472S-478S;2000.
41 Yang GY, Liao J, Kim K, Yurkow EJ, Yang CS. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 19:611-616; 1998.
42 Yang GY, Liao J, Li C, Chung J, Yurkow EJ, Ho CT, Yang CS. Effect of black and green tea polyphenols on c-jun phosphorylation and H202 production in transformed and non- transformed human bronchial cell lines. Possi- ble mechanisms of ceil growth inhibition and apoptosis induction. Carcinogenesis 21:2035- 2O39;2000.
43 Zhang G, Miura Y, Yagasaki K. Induction of apoptosis and cell cycle arrest in cancer cells by in vivo metabolites of teas. Nutr Cancer 38: 265-273;2000.
E G C G Inhibits Hep G 2 Cell Proliferation a n d Induces Apoptosis