J. TOXIC0L.-TOXIN REVIEWS, 16(3), 79-1 13 (1997)
PHOSPHOLIPASES A2 OF ASIAN SNAKE VENOMS
Inn-Ho Tsai
Institute of Biological Chemistry, Academia Sinica, Institute of Biochemical Sciences, National Taiwan University
P. 0. Box 23-106, Taipei, Taiwan
ABSTRACT
This review up-dated the structural and functional information of various phospholipase A2 (PLA2) isoforms purified from Asian snake venoms. A
phylogenic tree of group I PLA2s was constructed herein based on many
recently resolved amino acid sequences of the venom enzymes. It was found that PLA2s of Asian elapid venoms are structurally different from those of sea- snakeiAustralian elapid venoms, and are usually associated with cardiovascular effect, although exceptions such as 0-bungarotoxins do exist. Two types of
venom PLA2s appear to be present in the venom of Asiatic viperinae such as
Daboia and Echis, one has a N-terminal residue Asn and the other has the residue Ser. In the venom of Asiatic crotalinae, up to four subgroups of PLA2 isoforms are present and each of them is characterized by a distinct substitution at residue 6 (Glu, Asn or Arg) or residue 49 (Asp or Lys) in their sequences. The venom PLA2s in each of the subgroup show more or less functional similarity specific for the subgroup: the Glu6-PLA2s are usually antiplatelet,
79
80 TSAI
the Asn6-PLA2s are neurotoxic andlor myotoxic and many Arg6-PLA2s are anticoagulating, while the Lys49-PLA2s are myotoxic and edema-inducing. Mechanisms for the pharmacological actions of venom PLATS have been
discussed, including neurotoxicity, myotoxicity, antiplatelet activity,
anticoagulating activity, heparin-binding, protein-acylation and deacylation. Conclusions derived from many recent studies on pancreatic PLA2 by method of protein engineering render valuable information about the structure-activity relationship of the secretory PLA2 superfamily. Site-directed-mutagenesis methods coupled with relevant and dissecting functional assays are essential
for understanding the structure-activity relationship of snake venom PLA2s
with special function or toxicity.
1. INTRODUCTION
Asia is the habitat of various species of venomous snakes including Elapidae, Hydrophilidae and Viperidae. According to statistics, the rate of
snake bite and envenomation in Asia has been the highest among the world [ 11.
The characterization of snake venom components is important because a
suitable medical treatment depends on a better understanding of the site and
mode of action of the venom components. Phospholipases A2 (PLA2s, EC
3.1.1.4) are a group of enzymes that catalyze the Ca2'-dependent hydrolysis of
the 2-acyl ester bond in 3-sn-phospholipid. Secreted forms of the enzyme are abundant in the mammalian pancreas and in snake and bee venoms. Amino acid sequences of many PLA2s have been determined, with most being about
120 amino acids long and having 14 Cys residues forming seven disulfide
bonds. Overall these proteins are closely related (>45% identity), with key residues that are required for catalysis and structure to be conserved (for review, see references [2-81). PLA2s are classified into two groups based on their Cys
PHOSPHOLIPASES A, 81 snake families and in the mammalian pancreas, and group I1 are from the viperid snake venoms and mammalian nonpancreatic sources. Interestingly, PLAz isoforms of diverse physiological functions exist in the same venom source and they usually synergize with other venom components to display special pharmacological effects. This mini-review would update the recent progress in purification, structural analysis, toxicology and reaction mechanism
of venom PLA2s from important species of Asia snakes. Since the 14 kDa
PLA2 family has been the subject of many comprehensive reviews [2, 51, I will focus on recent findings about molecular and mechanistic analysis related with the PLA2s from Asian snake venom, and examine roles of the PLAz isoforms in the context of toxinology of the whole venom and the phylogenic relationship between the PLAzs.
2. STRUCTURE OF PLAz ISOFORMS FROM SNAKE VENOM
a. Elapidae and Hydrophiidae
Highly homologous PLA2s from Asian elapid venoms were purified and characterized. More than 15 PLA2 sequences from cobra and ringhal venoms
were known [35]. Recently, the cDNAs encoding three PLAz isoforms in Nuju
nuja spututix [9] and two PLA2 isoforms in Naju nuju atru [ 10, 111 were cloned and sequenced.
Two acidic PLA2s with PI of 3.8 and 3.9 [12] and one with PI of 5.2 [13]
were purified from King cobra (Ophiophugus hannuh) venom. The amino acid
sequence of the latter PLA2 was completed [13, 271 and found to be similar to
that of PLAz I11 of Bungurus fusciutus venom and those of cobra PLAzs.
It was found that as many as eight PLA2 isoforms exist in the venom of
southeast- Asia golden krait Bungurus fusciutus, including four catalytically
active PLA2s (111, Vb-1, X-I, X-2), three less active Pro30 PLA2s (Va, Vb-2
82 TSAI and VI together count for about 70% of the total mass of the crude venom. The complete amino acid sequences of these PLA2 were all solved 114, 151.
Highly neurotoxic venom of Bungarus multicinctus contains the unique
pre-synaptic neurotoxin, P-bungarotoxin, in addition to a-bungarotoxin, the post-synaptic neurotoxins whose homologous toxins are commonly present in all the Elapidae and Hydrophiidae venoms. There are more than 10 isoforms of P-bungarotoxin present in the venom and each is a covalent heterodimer of
PLA2 (A chain, 14 kDa) and a homolog of Kunitz-type protease inhibitor (B
chain, 7 kDa) [16]. Six variants of the A chain of P-bungarotoxin have been identified and at least four of their cDNA sequences were analyzed [17-20].
Therefore, PLAzs in the venom of 5ungavus species are much diversified
as compared with cobra venom PLA2s. Interestingly, the venom PLA2s of B. fasciatus are not presynaptically neurotoxic but cardiotoxic [2 11 and structurally very different from those of B. multicinctus. This is in accord with the finding that their venom components other than PLA2 are also different: for
example, procoagulating factor X activator was found in venoms of 5.
fasciatus [22] and king cobra [23] but not in that of B. multicinctus. The acidic PLA2 from king cobra venom was found to damage both heart and skeletal muscle[24] and also inhibit platelet aggregation [27]. Venom of sea-snakes usually contains strong post-synaptic neurotoxins and myotoxic PLA2s. The primary structures of PLA2s from hydrophiid snake venom were solved for the following species: Enhydrina schistosa [25], Laticauda colubrina [26], Laticauda laticaudata [28], Laticauda semifasciatus 1291 and Aipysurus laevis
[30]. Some of the myotoxic sea-snake PLA2s are also neurotoxic [25, 261.
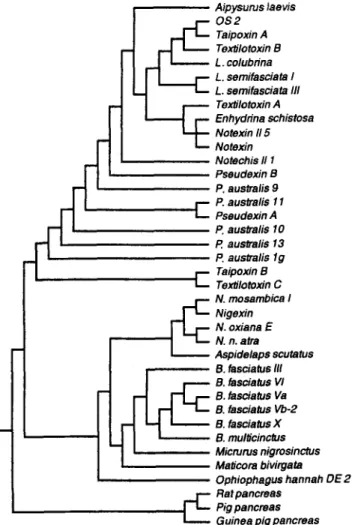
Results of phylogenic analysis of representative PLA2s from elapid and hydrophilid venoms (Fig. I ) suggest that sea-snake PLA2s are structurally more related with those of Australian snake venoms than with those from venoms of cobra, ringhal or krait, i.e. Asian and African Elapidae. Similar conclusions could also be drawn from the phylogenic tree contructed previously from
PHOSPHOLIPASES A, c Taipoxin B Texiilomxin C N. mosambica I Nigexin N. oxiana E N. n. atra 83
Fig. 1. Phylogenic tree of selected group I venom PLA2s based on the amino acid sequences. The sequences were extracted from the Swiss-Port data bank. The tree was constructed using program PAUP [156], and using pancreatic PLA2 as the outgroup.
another set of sequences [ 1021. P-Bungarotoxins are highly specialized and
phylogenically branch-off from other elapid PLA2s [ 1021.
Although the best known feature of envenoming by elapid and hydrophilid snakes is neurotoxicity, the cobra venom PLA2s are non-
84 TSAI
neurotoxic in clinical symptoms. Having a conserved structure with sequence identity of about 85%, they show cardiovascular effects either alone or in
synergy with the cytotoxin (cardiotoxin) in the same venom both in vivo and in
vitro [31-351. The acidic and neutral cobra PLAZs are more active than the basic cobra PLAzs in causing cardiac stimulation and transient hypotension [36]. The highly active cobra PLA2s also cause hyperkaliemia in the blood of victims due to membrane leakage and cell lysis [37]. It has been speculated that
the non-neurotoxic and monomeric PLAz in B. multicinctus venom may cause a
sharp fall in arterial blood pressure [37].
The three-dimensional structures of a single-chained PLA2 from Taiwan cobra venom [38] and of notexin, an Australian elapid PLA2 neurotoxin [39,40]
were solved by X-ray crystallography at 2
1.
The crystal structure ofp2-
bungarotoxin was also solved, showing a occluded substrate-binding surface and reduced hydrophobicity of the PLA2 subunit [41].
b. Subfamily Viperinae
The medically important viperinae snakes in Asia are Daboia russelli (Russell’s viper), Echis carinatus [42], and Cerastes. The primary structures of PLA2 isoforms from these venoms were not characterized until recently. Daboia russelli has been classified into at least four subspecies: Daboia russelli formosensis (Taiwan), Daboia russelli pulchella (Sri Lanka and southern India), Daboia russelli russelli (northern India and Pakistan), and Daboia russelli siarnensis (China and south-east Asia). The geographic variations of their bite-symptoms are remarkable and their antivenins usually show poor cross-neutralization. Venoms of four Russell’s viper subspecies were compared in terms of their HPLC profiles and partial amino acid sequences of their PLA2s. A potent, heterodimeric PLAz neurotoxin (designated as Russtoxin) was found in all the viper venoms analyzed except
PHOSPHOLIPASES A, 85
previously studied by Gowda et al. [44, 451 appear to be structurally the same
as those of D. r. pulchella (Sri Lanka), while the Russtoxins from D. r. russelli
(Pakistan) and D. r. siamensis (Thailand) resemble that from D. r. formosensis
[43]. Moreover, the published N-terminal amino acid sequences of the venom PLA2s of D. r. siamensis from Fujian (China) [46] and from Burma [47] are
also similar to that of D. r. formosensis. However, PLA2s of D. r. siamensis
from these three regions show some variations in their N-terminal sequences (Table 1). The structural and functional data of the venom PLA2s provide
evidence for the presence of two major types of Russell’s vipers. The species D.
r. forrnosensis, D. r siamensis (Fujian, Thailand, and Burma) and D. r. russelli (Pakistan) represent one type whose venom contains hypotensive and neurotoxic PLA2s having an Asn residue at the N-terminus [43, 481, while D. r. pulchella (southern India and Sri Lanka) represents the other type whose venom contains inyonecrotic PLA2s [49] with a N-terminal Ser residue. This finding is consistent with the reported antivenom differences between the Sri Lankan and the northern or western Indian subspecies [SO] and also in accord with the report that two distinct groups of Daboia russelli population was
found by means of multivariate morphometrics [S 11.
The complete sequences for heterodimeric neurotoxic PLA2 (F4-F7) from
the venom of D. r. formosensis were deduced from the cDNA sequences of
both subunits. They are 92% identical to the vipoxidinhibitor pair from the venom of Bulgarian Vipera ammodytes [48]. Structures of both subunits of the heterodimeric PLA2 toxin from Pseudocerastesfildi (false horned viper, Israel) were found to be over 90% identical to those of the F4-F7 and the vipoxin/ inhibitor pairs [52]. Moreover, similar heterodimer PLA2 toxins were found in the venoms of western-Asian species including Vipera aspis [53] and Vipera palaestinae [54]. The acidic subunit or charperon of the toxins plays the role of protecting the basic subunit against non-specific binding and thereby increases the probability of basic subunit reaching the neuromuscular junction or the
Asterisks denote numbering by each 10 residues. For PLA2 in each subgroups (separated by dashed lines), only the sequence of the first PLA2 and residues different from those of the first are shown. Abbreviations of PLA2s are the same as [43] with R stands for D. r. russelli, F for D. r. formosensis, S for D. r. siamensis (Thai), and P for D. r. pulchella (Sri Lanka), DRS (Fujian) for D. r. siamensis from Fujian, China [46]. DbTx for daboiatoxin from D. r. siamensis (Burma) [471.
PHOSPHOLIPASES A, 87
endothelium membrane causing neurotoxicity or hypotension (Tsai et al. to be published). Recently, synaptosomal binding of '251-labelled daboiatoxin from
Burmese Daboia r. siamensis was studied [55]. The toxin binding could not be
antagonized by the myotoxic PLA2s from south Indian D. russelli (i.e. D. r.
Pulchella) venom. However, it remains to be checked whether Burmese daboiatoxin present in the venom as a monomer or heterodimer.
Besides the heterodimeric toxic PLA~s, monomeric PLA2s of low lethal
potency were identified and sequenced for the venoms of Pakistan Cerastes
cerastes [56], west-Pakistan Eristocophis macmahoni [57], and those of D. r. siamensis and D. r. russelli (e.g. R1, Sl-1, S1-2 in Table 1) [43, 581. These homologous PLA2s appear to play a hypotensive or antiplatelet role in rats.
Venom of Echis carinatus sochureki (north India and Pakistan) contains a Ser49 PLA2 which was recently sequenced and characterized [59]. The substitution of Ca'l-binding Asp49 with a Ser49 in this enzyme does not abolish its enzymatic activity and this basic PLA2 could induce platelet aggregation. Previously, partial sequences of two PLAz from the venom of
Kenyan Echis pyramidum leakeyi (formerly E. c. leakeyi) were reported [60]
and one of the sequence is very similar to that of the E. c. sochureki Ser49
PLA2, while the other is similar to that of the Asp49 PLAz from Pakistan vipers
[56, 571. The Croatia/ Slovenija Vipera ammodytes venom also contain Ser49
myotoxic PLA2 (i.e. Ammodytin L) but without detectable enzyme activity [6 11.
A weakly basic PLA2 was purified from Indian E. carinatus venom but the
sequence was not reported [62].
c. Subfamily Crotalinae
The generic name of some of the Asiatic Agkistrodon species has been changed or remains controversial [42] e.g. the monotypic Calloselasma rhodostoma and Deinagkistrodon acutus have been renamed. Agkistrodon halys Pallas is designated as A . blomhof$i brevicaudus now. The green species
88 TSAI
of Trimeresurus have given rise to considerable confusion because of the great
similarity between different forms. T. gramineus has been mistakenly used to
name Trimeresurus stejnegeri (Taiwan) by toxinologists in reporting PLA2 and other components of this venom [42, 70, 711.
The venom PLA2 isoforms from the following species of Asian pit vipers
have been characterized and sequenced:
T.
flavoviridis [63-651, T. okinavensis[63],
T.
mucrosquamatus [66-691, T. stejnegeri (T. gramineus) [70, 711, A. b.brevicaudus [72-741, A . h. blomhoffii [75-771, Deinagkistrodon acutus [78-801. My classification of these PLA2 according to similarities in their structures and pharmacological properties resulted in four subgroups of the crotalid venom PLA2 (Fig. 2-5). Each subgroup has a distinct residue 6 or 49. Sequence identities within a subgroup are usually >70% while between the subgroups are
usually - 4 5 % . PLA2s in the Glu6 subgroup (Fig. 2) are inhibitors of platelet
aggregation [78, 811, those in the Arg6 subgroup (Fig. 3) are strong anticoagulant [82], direct-hemolytic [77] and bactericidal when associated with mammalian bactericidal/ permeability-increasing protein [75, 83-85]. PLA2s in the Asn6 subgroup( Fig. 4) are neurotoxic or/and myotoxic [67, 86-89], and
those in the Lys49 subgroup (Fig. 5 ) possess myotoxic [65, 71, 911, edema-
inducing [68] and membrane depolarizing activities [65, 921. The enzymatic
activities of Lys-49 PLA2s are usually hardly detectable or lower than those of
the Asp-49 enzymes [89, 931; however, the hydrolytic activity of the Lys49 PLA2s of T. flavoviridis [94] and of other venoms [91] could be demonstrated.
Three dimensional structures of crotalid PLA2s have been investigated by
X-ray diffraction on the crystals derived from the acidic dimeric PLA2 of T.
flavoviridis [95], the acidic monomeric PLA2 of A. h. blomhoffi venom [96],
the acidic PLA2 of A. b. brevicaudus venom [97], and the Lys49 PLA2 from A.
p . piscivorus venom [98] whose sequence is highly similar to those of the
Lys49 PLA2s of other Asian pit-vipers (Fig. 5). X-ray crystallographic
1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. Abb acidic Ahb 11 DAV acidic App-dimer C. adamanteus TMV-PL I TFV-PL 1, TFV-PL Ib T. okinavensis TGV-PL I TGV-PL I1 % identity 100 81 85 80 76 75 73 73 73 79 73 Fig. 2. Comparison of amino acid sequences of Crotalinae phospholipases A2 with Glu 6 substitution. Single-letter codes of amino acids were used. The numbering system follows that of Renetseder et al. [go]. Residue identical to that in the top line is denoted with a dot; gaps are marked with hyphens. Abbreviations and references: Abb, Ahb, App, DAV are Agkistrodon blomhoffii brevicaudus, Agkistrodon halys blomhoffi, Agkistrodon piscivorus piscivorus and Deinagkistrodon acutus [79]. respectively; TMV, TFV, TGV are venom of Trimeresurus flavoviridis [641, Trimeresurus mucrosquamatus [661, and Trimeresurus gramineus or stejnegeri [701, respectively.
W 0 1 10 20 30 40 50 60
+
+
+
+
++
+
1. TFV PL-X HLLQFRKMIKKNTG-KEPIVSYAF'YGCYCGKGGRGKPKDATDRCCFVHDCCYEKVT---GC--- 3. Abb-basic ... 4. Ahb-basic ... 2. TFV PL-X'...
... 5. TSV PL I1 ....L...N-....L..SK...MA...V...G...---S.--- 6. TMV PL I11 N.W ...E...m. .-...LTT.L..A....E....T...G.L.---A.--- 7. Rabbit ascite fluid...
D...-.RYT..-..ATT..GA-..H..V... 'X' (xxxxxxxxxxxxxxxxxxrowDDt 70 80 90 100 110 12 0 130 % identity+
++
+ + ++
1. --DPKWSWTYSLENGDIVCGG-DPYCTKVKCECDKAAAICFRFPDIFCT-DPTEGC 100 2. --....D...S...-.NP...EV...V...-...-... 94 3. --....DD....WK..T...-.DP.K.EV...K-....y.. .L.S-SKS.K. 84 4. --K.. .DD....WK...-.DP.K.QI....R...K-...AY...L..-SKS.K. 84 5. --ST. .DS,S. .W. 79 6. --s. 76 7. --x).FLS.KF.MK(- ) . . ' . . .A'. (B)K.QFY.ANR.S-GRPPS. Fig. 3. Comparison of amino acid sequences of Crotalinae PLA2 with an Arg 6. Single- letter codes of amino acids were used. The numbering system follows that of Renetseder et al. [go]. Residue identical to that in the top line was denoted with a dot, gaps are marked with hyphens, "x" denotes unknown residue. Abbreviations are as those in Fig. 2. References for sequences: 1 and 2 [63, 641, 3-5 [74-761, 6 [69] and 7 [851.ti
e
1. 2. 3. 4. 5. 6. 7. 8. 1. 2. 3. 4. 5. 6. 7. % identity 100 84 75 72 67 74 73 Fig. 4. Comparison of amino acid sequences of Crotalinae phospholipases A2 with a neutral residue 6 (or Asn 6). Single-letter codes of amino acids were used. The numbering system follows that of Renetseder et al. [go]. Residue identical to that in the top line is denoted with a dot: gaps are marked with hyphens. References for the sequences are 1-5 [79, 861, 6, 7 [881 and 8 [891.
W N 1. D. 2. T. 3. T. 4. T. 5. B. 6. B. 7. A. 8. A. acutus K49 mucrosquamatus K49 flavoviridis BPI gramineus PLA-V asper-I1 jararacussu-1 p. piscivorus K49 c. laticinctus MTX 1. 2. 3. 4. 5. 6. 7. 8. 60 70 80 + + + 90 100 110 120 130 identity + + + + + 100 100 81 18 78 78 74 77 Fig. 5. Comparison of amino acid sequences of Crotalinae PLA2 with Lys 49 substitution. Single-letter codes of amino acids were used. The numbering system follows that of Renetseder et al. [go]. Residue identical to that in the top line is denoted with a dot; gaps are marked with hyphens. References for the sequences are [71, 80, 911.
PHOSPHOLIPASES A, 93
from A . p. piscivorus venom were investigated and the crucial roles of its
residues 7 and 10 in interfacial binding were shown [99]. These
crystallographic studies confirmed the conservation of core area conformation of the PLAz molecules and variations in surface residues and loops. Notebly, results from recent NMR studies [loo, 1011 on the pancreatic PLA2 have demonstrated that the N-terminal residues 1-3 of the secretory 14 kDa PLA2 in solution are not ordered and the enzyme is activated upon binding to the aggregated substrate. The molecular dynamics were not revealed by previous crystallographic data.
3. EVOLUTION OF VENOM PLA2
Study on the evolution and the phylogeny of venom PLA2 at the molecular
level started several years ago [102]. The group I and group I1 PLA2s are
known to have evolved separately. Five cDNA and six genes encoding the
venom gland PLAzs of T. flavoviridis were sequenced [64], so were the PLA2
genes from T. grumineus [ 1031, those of Mojave toxin (C. s. scutulutus) [104],
and those from Vipera a. ummodytes [105]. The data confirmed a positive
Darwinian evolution of these genes. Interestingly, four and five exons were found in Crotalinae and Viperinae PLA2 genes, respectively. The first intron was retained in the mRNA of Crotalinae PLA2 in contrast to that of Viperinae PLA2, possibly due to change in secondary structure of the first exon of Croralinae PLA2 gene [105]. The protein-coding regions are much more diversified than the 5’ and 3 ’ untranslated regions (UTRs) and the introns except for the signal peptide domain. The numbers of nucleotide substitutions per site for the UTRs and the introns were approximately one-quarter of the numbers of nucleotide substitutions per synonymous site for the protein-coding regions. However, the UTRs and the introns of venom PLA2 genes have evolved at similar rates to those of non-venomous genes. Apparently, gene
94 TSAI
duplication and accelerated evolution in the protein coding regions is universal in PLA2 genes of Crotalinae and Viperinae venom. The venom genes have been evolving under adaptive pressure to acquire new physiological activities [64, 88, 103, 1061.
It has been shown that possibly four group I1 PLA2 genes which map to
the same chromosome are present in mammalian genome [107], and it is likely
that snakes also have several group I1 PLA2 genes expressed in non-venomous
tissues, and venom PLA2 isoforms may be derived from more than one non- venomous PLA2 gene.
Examination of nucleotide sequences for five group I PLA2 cDNAs from
four genera of elapids [I081 also revealed high similarity of the non-coding region and more variability in the coding regions; but the conserved sequences from the elapids have no nucleotide sequence similarity to corresponding regions in viperid PLA2 genes. Furthermore, when nucleotide sequences for cDNA clones of two metalloproteases from Agkistrodon contortrix laticinctus
[ 1091 and those from Echis pyramidum Zeakeyi [ 1 101 were compared, there
appears to be high conservation of noncoding DNA sequences for the
metalloprotease genes. Thus, most of the venom protein genes probably
undergo a coordinated and fast evolution to ensure the production of an effective venom to meet the requirment of snakes for food and defense.
4. BINDING PROTEIN AND ACTION MECHANISM
It has been difficult to define the pharmacological effect of a venom PLA2
in vivo and in vitro. Endeavours in mechanistic studies of venom PLA2s have been fruitful but many questions remain to be answered.
a. M-type receptor
This 180 kDa membrane protein, identified to be present in mammlian
PHOSPHOLIPASES A, 95
and the group I1 inflammatory-type PLA2 [ l 1 1-1 131. The receptor gene has
been cloned and expressed [l 1 1, 1141 and was found to have endocytic
property [ 1 151. Significance of the receptor in the regulation of PLAl action
remains to be elucidated.
b. Neurotoxic PLA2s and binding proteins
High affinity and specific receptor for presynaptically toxic
p-
bungarotoxin has been identified to be a voltage dependent K'-channel [ 1 16-
1 181. The binding proteins (or its subunit) on synaptic membrane for crotoxin and taipoxin have been found to be the norepinephrine transporter [119]. The
receptors for ammodytoxin A and C on the bovine brain were identified, too.
However, ammodytoxins could not inhibit all the high affinity binding of crotoxin, suggesting that receptors for crotoxin and ammodytoxin are not identical [ 120, 12 I]. The hypotensive PLA2s from Indian Duboiu r. russelli venom bind receptors rather differently from the receptors mentioned above [ 122- 1241. Recently, synaptic binding protein of daboiatoxin, the myotoxic and
neurotoxic PLAz from Burmese Duboiu russelli venom, has been identified to
be a 100 kDa protein with two subunits of 25 kDa and 75 kDa [55]. There was
no competition by ammodytoxin or crotoxin for the receptor. It remains to be
clarified whether the heterodimeric PLA2 toxins from various D. russelli
venoms [43,52] have similar binding sites as daboiatoxin or not. The diversified binding sites for presynaptic PLA2 toxins have been reviewed recently by Tzeng [125].
We and others have shown that the N-terminal region and regions 76-81 and 119-125 of PLA2 neutrotoxins are involved in their receptor binding or neurotoxicity [48, 67, 126, 1271. The cellular effects of the PLA toxins on the nerve-terminal have been reviewed in a recent paper [ 1281. The detailed mechanism after receptor binding and the role of phospholipid hydrolysis in the blockade of neurotransmitter release by the toxins are not clear.
96 TSAI
c. Anticoagulating activity
Basic motif with four or five ArgLys in strongly anticoagulant PLA2
from Nuja nigricollis venom [82] is involved in its inhibition of prothrombin
complex [ 1291. The anticoagulating human group I1 PLA2 inhibited
coagulation factor Xa or its interaction with factor Va also by a basic region at
residuses 5 1-62 [ 1301. However, specific sites in other anticoagulating PLA2s from viperid venoms, which are involved in a possibly similar mechanism, remains to be identified.
d. Antiplatelet activity
Venom PLA2 interferes with hemostasis by either anticoagulant or
antiplatelet effects. The inhibition of platelet aggregation by PLA2s may be mediated by the generation of lysophospholipid [131, 1321 and by resultant
change in cytoskeleton and hence the loss of release reaction of platelets [27].
However, the specificity for platelet membrane appears to be a prerequisite of the antiplatelet activity of venom PLA2. Recently, a structural feature responsible for this activity was proposed based on the 3-dimensional structure
of the antiplatelet PLA2 from A . b. brevicaudus (ie. A . h. Pallas) venom. A
unique aromatic patch (residues 20, 21, 113 and 119) surrounded by two acidic
residues (Glu 6 and Asp 1 15) on one face of the PLA2 molecule was postulated
to be implied in the recognition of platelet membrane [97].
e. Myonecrotic action and myotoxicity
In accord with their phylogeny relation (Fig. 1) hydrophiid and Australian elapid venom PLA2s cause systemic myotoxicity and myoglobinuria in
common while viperid basic PLA2s with or without neurotoxicity and the
Lys49 PLA2s produce local myonecrosis [24]. The damaged muscle showed
dilatation of sarcoplasmic reticulum, vascuolation, then disruption and
hypercontraction of the fibres, and inflammatory reaction. Influx of Ca2' into
PHOSPHOLIPASES A, 91
membrane damaging by PLA2s [24, 1331. PLA2 myotoxins showed differential
specificity for different types of skeletal muscle [134]. The Lys49 PLA2s are cytotoxic, edema-inducing and disrupt membrane by poorly understood Ca2+- independent mechanism. Their edema-inducing activity involves degranuation
of mast cell and PMN [68].
f. Binding to sulfated proteoglycans
A cluster of three or more basic residues is a potential motif in protein for
its binding to heparin or other sulfated proteoglycans [135]. This motif is present in some Lys49 PLA2 sequences and possibly also exists in the folded conformation of some Asp49 PLA2s. It was well documented that some venom
PLA2s bind heparin [136-1381, and so does the human group
I1
PLA2 [138,1391. The binding not only serves for anchoring or concentrating the PLA2 to specific cells but also modulates its enzyme activity.
g. Protein deacylation and autocatalytic acylation of PLA2
It has been shown that PLA2 may hydrolyze thioester bond of long chain acyl-CoA and show protein-deacylase activity for acyl-carrier protein [ 1401. It is likely that venom PLA2 may hydrolyse some acyl-protein anchored on cell membranes although no such case has been reported. A PLA2 may undergo autocatalyzed acylation of specific Lys residues to certain extend in the presence of phospholipid substrate [ 1411. On the other hand, Lys 49 and Ser 49 PLA2s are able to undergo acylation spontaneously with free fatty acids [ 1421. The acylated PLA2s usually became more hydrophobic and tightly bound to membrane or catalytically more active.
5. CONCLUSION AND PERSPECTIVES
Due to their abundance in snake venoms, thermal and acid stability, and relative ease of purification, PLA2s are among the best studied enzymes.
98 TSAl
Advances in technology including HPLC, mass spectrometry, polymerase chain reaction and nucleotide sequencing have facilitated the determination of amino acid sequences of more venom PLAzs. Secondary and tertiary structures of PLA2s were elucidated by X-ray crystallography, NMR and other methods [3, 1431. Although previous works, mainly chemical modification studies [ 1441, have accumulated rich data, the structure-activity relationship of venom PLAl remain to be reinvestigated for conclusion by incorporating new methods such as in vitro mutagenesis.
The enzymatic activity of PLA2 found in vitro may not well reflect that in
vivo; and bilayer, monomeric substractes need to be tested in addition to micellar substrates with different electrostatic properties or phospholipid substrates with different head-groups. For functional studies, relevant and diversified assays are ususlly needed to study the pharmacological effects of a venom PLA2. It may be essential to dissect the PLAZ action into steps including receptor targeting and binding, interfacial binding and activation, active site catalysis, kinetic specificity, etc. Independent analysis of each step is necessary to make a conclusion about the structure-activity relationship of the PLAz action. Besides, species specificity [23] and tissue specificity [ 1341 in toxin assays are important considerations. These are challenges for the researchers of venom PLA2 toxins. It would not be surprising to find that conformational change occurs upon binding of a venom PLA2 to its binding protein or specific target, since conformational dynamics of group 1 PLAzs at the N-terminal and 53-72 regions upon interfacial binding [98, 1451 or ligand binding [39] have been well documented.
In the past few years, site-directed-mutagenesis studies of pancreatic
PLA2s have shed considerable light on the role of its active-site residues [93, 146, 1471, the N-terminal [145, 1481 and the C-terminal regions [149], the disulfide bonds [150] and its interficial residues [97, 145, 148, 151, 1531. The results are important and fundamental for understanding the structure-activity
PHOSPHOLIPASES A2 99
relationship of the 14 kDa PLAz family. However, venom PLAzs have evolved from nontoxic enzymes to those with special toxicity or tissue specificity. More protein-engineering studies on their functional domains are apparently required.
Interfacial-binding mutants of a cationic PLAz from the venom of A . p .
piscivorus have recently been studied and the results showed that the critical
charged groups involved in interfacial adsorption for this groups
I1
PLAZs isvery different from those for pancreatic PLAz [99, 1541. It is likely that interfacial residues of each subgroup of venom PLAZs are characteristic. The interfacial binding surface of the bee venom PLAz was shown to be part of its
neuronal receptor-recognition domain by site-directed-mutagenesis [ 1551.
There is clearly much scope for future research on the structure-activity relationship of venom PLA2 isoforms with such complexicity and functional diversity.
Acknowledgement: I thank the National Science Council of Taiwan and
Academia Sinica for grant supports, and Dr. T. Y. Chiang at Department of
Biology, Cheng-Kung University for preparing phylogenic tree of PLAZ. The
secretarial helps of Dr. Yuh-Ling Chen and Miss
S.
H. Rao are greatlyappreciated.
REFERENCES
1. Warrell, D. A. Clinical features of envenoming from snake bites. In:
Envenoming and their treatment, edited by C . Bon and M. Goyffon,
Foundation Marcel Merieux, France, p. 63, 1996.
2. Rosenberg, P. Phospholipases. In: Handbook of Toxinology, edited by W. T.
Shier and D. Mebs, Marcel Dekker, Inc. New York, p. 67, 1990.
3 . Scott, D. and Sigler, P. B. Structure and catalytic mechanism of secretory
too TSAI
4. Danse, J. M., Gasparini, S and Menez, A. Molecular biology of snake venom
phospholipase A2. In: Venom Phospholipase A2 Enzymes: Structure,
Function and Mechanism, edited by R. M. Kini, John Wiley & Sons, UK, p.
29. 1997.
5 . Davidson, F. F. and Dennis, E. A. Structure, function, and mode of action of
snake venom and other phospholipase A2 In: Handbook of Natural Toxins,
edited by A. T. Tu, Marcel Dekker, Inc., New York, Vol. 5, p. 107, 1991.
6 . Hawgood, B. and Bon, C. Snake venom presynaptic toxins. In: Handbook of
Natural Toxins, edited by A. T. Tu, Marcel Dekker, Inc., New York, Vol. 5,
p. 3, 1991.
7. Harris, J. B. Phospholipases in snake venom and their effects on never and muscle. In: Snake Toxins, edited by A. L. Harvey Pergamon Press, Inc. New York, p. 91, 1991.
8. Kudo, I., Murakami, M., Hara, S. and Inoue, K. Mammalian non-pancreatic
phospholipase A2. Biochim. Biophys. Acta 117: 21 7, 1993.
9. Armugam, A., Earnest, L., Chung, M. C. M., Gopalakrishnakone, P., Tan, C. H., Tan, N. H. and Jeyaseelan, K. Cloning and characterization of cDNAs encoding three isoforms of phospholipase A2 in Malayan spitting cobra
(Naja naja sputatrix) venom. Toxicon 35: 27, 1997.
10. Pan, F. M., Yeh, M. S., Chang, W. C., Hung, C. C. and Chiou, S. H.
Sequence analysis and expression of phospholipase A2 from Taiwan cobra.
Biochem. biophys. Res. Commun. 91: 95, 1994.
1 1 . Pan, F. M., Chang, W. C. and Chiou, S. H. cDNA and protein sequences
coding for the precursor of phospholipase A2 from Taiwan cobra Naja naja
atra. Biochem. mol. Biol. Int. 33: 187, 1994.
13. Tan, N. H. and Saifuddin, M. N. Purification and characterization of two
acidic phospholipase A2 enzymes from king cobra (Ophiophagus hannah)
snake venom. Int. .IBiochem. 22: 481, 1990.
13. Chiou, J. Y., Chang, L. S., Chen, L. N. and Chang, C. C. Purification and
characterization of a novel phospholipase A2 from king cobra (Ophiophagus
hannah) venom. J. Protein Chem. 14: 45 1, 1995.
14. Liu, C. S., Kuo, P. Y, Chen, J. M., Chen, S. W., Chang, C. H., Tseng, C. C.,
Tzeng, M. C. and Lo, T. B. Primary structure of an inactive mutant of
phospholipase A2 in the venom of Bungarus fasciatus (Banded Krait).
J.
PHOSPHOLIPASES A2 101
15. Liu, C. S., Chen. J. M., Chang, C. H., Chen, S. W., Tsai, I. H., Lu, H. S. and
Lo, T. B. Revised amino acid sequences of the three major phospholipase A2
from Bungarus fasciatus (banded krait) venom. Toxicov 28: 1457, 1990.
16. Kondo, K., Toda, H., Narita, K. and Lee, C. Y. Amino acid sequence of three
(3-bungarotoxins ((33-, p4-, and p5-bungarotoxins) from Bungarus
multicinctus venom. Amino acid substitutions in the A chains. J . Biochem.
91:1519, 1982.
17. Chu, C. C., Li, S. H. and Chen, Y. H. Resolution of isotoxins in the (3-
bungarotoxin family. J . Chromatogr. A 694: 492, 1995.
18. Danse, J. M., Toussaint, J. L. and Kempf, J. Nucleotide sequence encoding
(3-bungarotoxin A2-chain from the venom of Bungarus multicinctus. Nucleic
Acids Res. 18: 4609, 1990.
19. Danse, J. M., Gamier, J. M. and Kempf, J. cDNA deduced amino acid
sequence of a new phospholipase from the venom of Bungarus multicinctus.
Nucleic Acids Res. 18: 46 10, 1990.
20. Chang, L. S., Wu, P. F and Chang, C. C. cDNA sequence analysis and
expression of the A chain of (3-bungarotoxin from Bungarus multicinctus
(Taiwan banded krait). Biochem. Biophys. Res. Commun. 221: 328,1996.
21. Lin-Shiau, S. Y., Huang, M. C. and Lee, C. Y. A study of cardiotoxic
principles from the venom of Bungarus fasciatus (Schneider). Toxicon 13:
189, 1975.
22. Zhang, Y., Xiong, Y. L. and Bon, C. An activator of blood coagulation
factor X from the venom of Bungarus fasciatus. Toxicon 33: 1277, 1995.
23. Lee, W. H., Zhang, Y., Wang, W. Y., Xiong, Y. L. and Gao, R. Isolation and
properties of a blood coagulation factor X activator from the venom of king
cobra (Ophiophagus hannah). Toxicon 33: 1263, 1995.
24. Huang, M. Z. and Gopalakrishnakone, P. Pathological changes induced by
an acidic phospholipase A2 from Ophiophagus hannah venom on heart and
skeletal muscle of mice after systemic injection. Toxicon 34: 201, 1996.
25. Lind, P. and Eaker, D. Amino acid sequence of a lethal myotoxic
phospholipase A2 from the venom of the common sea snake (Enhydrina
schistosa). Toxicon 19: 11, 1981.
26. Takasaki, C., Kimurs, S., Kokubun, Y. and Tamiya, N. Isolation, properties and amino acid sequences of a phospholipase A2 and its homologue without
activity from the venom of a sea snake, Laticauda colubrina, from the
102 TSAI
27. Huang,
M. Z.,
Gopalakrishnakone, P., Chung, M. C.M.
and Kini, R. M.Complete amino acid sequence of an acidic, cardiotoxic phospholipase A2 from the venom of Ophiophagus hannah (king cobra): a novel cobra venom
enzyme with “pancreatic loop”. Arch. Biochem. Biophys. 338: 150, 1997.
28. Guignery-Frelat, G., Ducancel, F., Menez, A. and Boulain, J. C. Sequence of
a cDNA encoding a snake venom phospholipase A2. Nucleic Acids Res. 15:
5892,1987.
29. Takasaki, C., Kuramochi, H., Shimazu, T. and Tamiya, N. Correction of
amino acid sequence of phospholipase A2 I from the venom of Laticauda
semfasciata (erabu sea snake). Toxicon 26: 474, 1988.
30. Ducancel, F. Guignery-Frelat, G. Bouchier, C. Menez, A. and Boulain, J. C.
Sequence analysis of a cDNA encoding a PLAl from the sea-snake
Aipysurus laevis. Nucleic Acids Res. 16: 9048, 1988.
31. Teng, C. M., Kuo, Y. P., Lee, L.
G.
and Ouyang,C.
Characterization of theanticoagulants from Taiwan cobra (Naja n. atra) snake venom. Toxicon 25:
201, 1987.
32. Fletcher, J. E. and Jiang, M. S. Possible mechanism of action of cobra
snake venom cardiotoxins and bee venom mellitin. Toxicon 31: 669, 1993.
33. Chen, Y. H., Lai, M. Z. and Kao, L.
S.
Destruction of liposome vesicles byTaiwan cobra cardiotoxin. Biochem. Int. 3: 385, 1981.
34. Bougis, P.
E.,
Marchot, P. and Rochat, H. In vivo synergy of cardiotoxin andphospholipase A2 from the elapid snake Naja mossambica mossambica.
Toxicon 25: 427, 1987.
35. Davidson F.
F.
and Dennis,E.
A. Amino acid sequence and circulardichroism of Indian cobra (Naja naja naja) venom acidic phospholipase A2.
Biochim. Biophy. Acta 1037: 7, 1990.
36. Ho, C. L. and Lee, C. Y. Cardiovascular effects of phospholipase Az purified
from various snake venoms. Proc. Natl. Sci. Counc. B, ROC 5: 18 1, 198 1.
37. Lee, C. Y. and Lee, S. Y. Cardiovascular effect of snake venoms. In: Snake
venoms. Handbook of experimental pharmacoiogy, edited by C .
Y.
Lee,Springer-Verlag, Berlin, 52: 546, 1979.
38. White, S. P., Scott, D. L., Otwinowski, Z., Gelb, M. H. and Sigler, P. B.
Crystal structure of cobra-venom phospholipase A2 in a complex with a
PHOSPHOLIPASES A, 103
39. Westerlund, B., Saarinen, M., Person, B., Ramaswamy, S., Eaker, D. and
Eklund, H. Crystallographic investigation of the dependence of calcium and
phosphate ions for notexin. FEBS Lett 403: 5 1, 1997.
40. Westerlund, B., Nordlund, P., Uhlin, U., Eaker, D. and Eklund, H. The 3-D
structure of notexin, a presynaptic neurotoxic phospholipase A2 at 2.0 A
resolution. FEBS Lett 301: 159, 1992.
41. Kwong, P. D., McDonald, N. Q., Sigler, P. B. and Hendrickson, W. A.
Structure of Pz-bungarotoxin: potassium channel binding by Kunitz modules
and targeted phospholipase action. Structure 3: 1109, 1995.
42. Wuster, W., Golay, P. and Warrell. D. A. Synopsis of recent developments in
venomous snake systematics. Toxicon 35: 3 19, 1997.
43. Tsai, I. H., Lu, P. L. and Su, J. C. Two types of Russell’s viper revealed by
variation in phospholipase A2 from venom of the subspecies. Toxicon 34: 99,
1996.
44. Kasturi, S. and Gowda, T. V. Purification and characterization of a major
phospholipase A2 from Russell’s viper (Vipera russelli) venom. Toxicon 27:
229, 1989.
45. Gowda, V. T., Schmidt, J. and Middlebrook, J. L. Primary sequence
determination of the most basic myonecrotic phospholipase A2 from the
venom of Vipera russelli. Toxicon 32: 665, 1994.
46. Li, Y . S., Liu K. F., Wang, Q. C . , Ran, Y. L. and Tu, G. C. Studies on a basic phospholipase A2 of Vipera russelli siamensis Smith venom from Fujian. Acta Biochim. Biophy. Sinica (PRC) 17: 607, 1985.
47. Maung-Maung-Thwin, Gopalakrishnakone, P., Yuen, R. and Tan, C. H. A
major lethal factor of the venom of Burmese Russell’s viper (Daboia russelli siamensis): isolation, N-terminal sequencing and biological activities of
daboiatoxin. Toxicon 33: 63, 1995.
48. Wang, Y . M., Lu, P. J., Ho, C. L. and Tsai. I. H. Characterization and
molecular cloning of neurotoxic phospholipase A2 from Taiwan viper (Vipera russelli formosensis). Eur. J. Biochem. 209: 635, 1992.
49. Gowda, T. V. and Middlebrook, J. L. effects of myonecrotic snake venom
phospholipase A2 toxins on cultured muscle cells. Toxicon 31: 1267, 1993.
50. Philips, R. E., Theakston, R. D. G., Warrell, D. A., Galigedara, Y.,
Abeysekera, D. T. D. J., Dissanayaka, P., Hutton, R. A. and Aloysius, D. J.
104 TSAI
(Vipera russelli pulchella) in Sri Lanka: failure of Indian (Haffiine)
antivenom. Q. J. Med. 68: 691, 1988.
51. Wuster, W., Otsuka, S., Malhotra, A. and Thorpe, R. S. Population
systematics of Russell’s viper: a multivariate study. Biol. J. Linn. SOC. 47:
97, 1992.
52. Francis, B., Bdolah, A. and Kaiser, I. I. Amino acid sequences of a
heterodimeric neurotoxin from the venom of the false homed viper (Pseudocerastesfieldi). Toxicon 33: 863, 1995.
53. Komori, Y., Nikai, T. and Sugihara, H. Comparative study of three
phospholipases A2 from the venom of Vipera aspis. Comp. Biochem. Physiol.
97B: 507, 1990.
54. Simon, T., Bdolah, A. and Kochva, E. The two component toxin of Vipera
palaestinae: contribution of phospholipase A2 to its activity. Toxicon 18: 249,1980.
55. Maung-Maung-Thwin, Gopalakrishnakone, P., Yuen, R. and Tan C. H.
Synaptosomal binding of 1251-lablled daboiatoxin, a new PLA2 neurotoxin
from the venom of Daboia russelli siamensis. Toxicon 34: 183, 1996.
56. Siddiqi, A. R., Shafqat, J., Zaidi, Z. H. and Jornvall, H. Characterization of
phospholipase A2 from the venom of homed viper (Cerastes cerastes). FEBS
Lett. 278: 14, 1991.
57. Siddiqi, A. R., Zaidi, Z. H. and Jomvall, H. Purification and characterization
of two highly different group
I1
phospholipase A2 isozymes from a singleviperid (Eristocophis macmahoni). Eur. J. Biochem. 201: 675, 1991.
58. Tsai, I. H., Lu, P. J. and Su, Y . C. Russtoxin: a new family of two-
component phospholipase A2 toxins from Russell’s vipers. In: Peptides;
Chemistry and Biology, Proc. 13th Amer. Peptide Symp., edited by R. S. Hodges and J. A. Smith. Escom Sci., The Netherland, p. 465, 1994.
59. Polgar, J., Mabnenat, E. M., Peitsch, M. C., Well, T. N. C. and Clemetson, J. C. Asp-49 is not an absolute prerequisite for the enzymic activity of low-
M, phospholipase Az: purification, characterization and computer modelling
of an enzymically active Ser-49 phospholipase A2, ecarpholin S, from the
venom of Echis carinatus sochureki (saw-scaled viper). Biochem. J. 319:
961,1996.
60. Desmono, H. P., Crampton, J. M. and Theakston, R. D. G. Rapid isolation
and partial characterization of two phospholipases from Kenyan Echis
carinatus leakeyi (Leakeyi’s saw-scaled viper) venom. Toxicon 29: 536, 1991.
PHOSPHOLIPASES A2 105
6 1. Krizaj, I., Bieber, A., Ritonja, A. and Gubensek, F. The primary structure of
ammodytin L. a myotoxic phospholipase A2 homologue from Vipera ammodytes venom. Eur. J. Biochem. 202: 1165, 1991.
62. Kemparaju, K., Prasad, B. N. and Gowda, V. T. Purification of a basic
phospholipase A2 from Indian saw-scaled viper (Echis carinatus) venom:
characterization of antigenic, catalytic and pharmacological properties.
Toxicon 32: 1187, 1994.
63. Kini, R. M., Kawabata, S. -I. and Iwanaga, S. Comparison of amino terminal
region of three isoenzymes of phospholipase A2 (TFV PL-Ia, TFA PL-Ib, TFV PL-X) from Trimeresurus flavoviridis (Habu snake) venom and the complete amino acid sequence of the basic phospholipase, TFB PL-X, Toxicon 24: 1 1 17, 1986.
64. Nakashima, K., Ogawa, T., Oda, N., Hattori, M., Sakaki, Y., Kihara, H. and
Ohno, M. Accelerated evolution of Trimeresurus flavoviridis venom gland
phospholipase A2 isozymes. Proc. Natl. Acad. Sci. USA 90: 5964,1993.
65. Kihara, H., Uchikawa, R., Hattori, S. and Ohno, M. Myotoxicity and
physiological effects of three Trimeresurus flavoviridis phospholipase Az. Biochem. Int. 28: 895, 1992.
66. Wang, Y. M. and Tsai, I. H. Cloning and sequencing of an acidic
phospholipase A2 from Taiwan habu (T. mucrosquamatus). J. Chinese.
Biochem. SOC. (R.O.C.) 23: 53, 1994.
67. Tsai, I. H., Lu, P. J., Wang, Y. M., Ho, C. L. and Liaw, L. L. Molecular
cloning and characterization of a neurotoxic phospholipase A2 from the
venom of Taiwan habu (T. mucrosquamatus). Biochem.
J.
(London) 311:895, 1995.
68. Liu, C. S., Chen, J. M., Chang, C. H., Chen S. W., Teng, C. M. and Tsai, I. H.
The amino acid sequence and properties of an edema-inducing Lys-49
phospholipase A2 homolog from the venom of Trimeresurus
mucrosquamatus. Biochim. Biophys. Acta 1077: 362, 1991.
69. Tsai, I. H., Lu, P. J., Wang, Y. M., Ho, C. L. andHwang, L. L. Molecular
cloning and characterization of snake venom phospholipases A2 from Taiwan vipers: Trimeresurus mucrosquamatus and Vipera russelli formosensis. In: Recent Advances in Molecular and Biochemical Research on Proteins, Proc. IUBMB Symposium on Protein Structure and Function,
edited by Y. H. Wei, C. S. Chen and J. C. &World Scientific, Singapore, p.
129, 1993.
70. Fukagawa, T., Nose, T., Shimohigashi, Y., Ogawa, T., Oda, N., Nakashima,
106 TSAI
amino acid-substituted phospholipase A2 isozymes from Trimeresurus gramineus (green habu snake) venom. Toxicon 31: 957, 1993.
71. Nakai, M., Fukagawa, T., Ogawa, T., Shimohigashi, Y., Hattori, S., Chang,
C. C. and Ohno, M. Purification and primary strucuture of a myotoxic
Lysine-49 phospholipase A2 with low lipolytic activity from Trimeresurus gramineus venom. Toxicon 33: 1469, 1995.
72. Kondo, K., Zhang, J., Xu, K. and Kagamiyama, H. Amino acid sequence of a
presynaptic neurotoxin, agkistrodotoxin, from the venom of Agkistrodon halys Pallas. J. Biochem. 105: 196, 1989.
73. Chen, Y. C., Maraganore, J. M., Reardon, I. and Heinrikson, R. L.
Characterization of the structure and function of three phospholipase A2
from the venom of Agkistrodon halys Pallas. Toxicon 25: 401, 1987.
74. Pan, H., Ou-Yang, L. L., Yang, G. Z., Zhou, Y. C. and Wu, X. F. Cloning of
the BPLA2 gene from Agkistrodon halys Pallas. Acta Biochim. Biophys. Sinica (PRC) 28: 579, 1996.
75. Forst, S., Weiss, J., Blackburn, P., Frangione, B., Goni, F. and Elsbach, P.
Amino acid sequence of a basic Agkistrodon halys blomhoffii phospholipase A2. Possible role of NH2-terminal lysines in action on phospholipids of Escherichia coli. Biochemistry 25: 4309, 1986.
76. Lee, S. Y., Wang, W. Y. and Xiong, Y. L. Purification, partial sequencing
and characterization of five phospholipases A2 from Trimeresurus stejnegeri
venom. Toxicon 35: 495, 1997.
77. Shukla, S. D. and Hanahan, D. J. Differences in the pattern of attack of
acidic, neutral, and basic phospholipase A2 of Agkistrodon halys blomhofii on human erythrocyte membranes: Problems in interpretation of
phospholipid location. Arch. Biochem. Biophys. 209: 668, 198 1.
78. Chen, R. H. and Chen, Y. C. Isolation of an acidic phospholipase A2 from
the venom Agkistrodon acutus (five pace snake) and its effect on platelet aggregation. Toxicon 27: 675, 1989.
79. Wang, Y. M., Wang, J. H. and Tsai, I. H. Molecular cloning and deduced
primary structures of acidic and basic phospholipases A2 from the venom of
Deinagkistrodon acutus. Toxicon 34: 49 1, 1996.
80. Wang, Y. M., Wang, J. H. Pan, F. M. and Tsai, I. H. Lys-49 phospholipase
homologs from venoms of Deinagkistrodon acutus and Trimeresurus mucrosquamatus have an identical protein sequence. Toxicon 34: 49 1, 1996.
PHOSPHOLIPASES A, 107
81. Huang, T. F., Yeh, H. I. and Ouyang, C. Mechanism of action of the platelet
aggregation inhibitor purified from Agkistrondon halys (mamushi) snake
venom. Toxicon 22: 243, 1984.
82. Kini, R. M. and Evans, H. J. Structure-function relationships of
phospholipases, the anticoagulant region of phospholipase A2. J. Biol. Chem.
262: 14402, 1987.
83. Weiss, J., Wright, G. W., Bekkers, A. C. A. P. A., van den Bergh, C. J. and
Verheij, H. M. Conversion of pig pancreas phospholipase A2 by protein
engineering into enzyme active against Escherichia coli treated with the
bactericidal/permeability-increasing protein. J. Biol. Chem. 266: 4 162, 199 1.
84. Forst, S., Weiss, J. and Elsbach, P. Structural and hnctional properties of a
phospholipase A2 purified from inflammatory exudate. Biochemistry 25:
8381, 1986.
85. Mizushima, H., Kudo, I., Horigome, K., Murakami, M., Hayakawa, M., Kim,
D. K., Kondo, E., Tomita, M. and Inoue, K. Purification of rabbit platelet
secretory phospholipase A2 and its characteristics. J. Biochem. 105: 520,
1989.
86. Faure, G., Choumet, V., Bouchier, C., Camoin, L., Guillaume, J. L.,
Monegier, B., Vuilhorge, M. and Bon, C. The origin of the diversity of
crotoxin isoforms in the venom of Crotalus durissus terrificus. Eur. J.
Biochem. 223: 16 1,1994.
87. Kaiser, I. I., Gutierrez, J. M., Plummer, D., Arid, S. D. and Odell, G. V. The
amino acid sequence of a myotoxic phospholipase from the venom of Bothrops asper. Arch. Biochem. Biophys. 278: 3 19, 1990.
88. Moura-da-Silva, A. M., Paine, M. J. I., Diniz, M. R., Theakston, R. D. G. and
Crampton J. M. The molecular cloning of a phospholipase A2 from Bothrops
jararaca snake venom: evolution of venom group I1 phospholipases A2 may
imply gene duplications. J. Mol. Evol. 41: 174, 1995.
89. Ownby, C. L., Collerg, T. R. and White, S. P. Isolation, characterization and
myotoxin from the venom of the
crystallization of a phospholipase A2
prairie rattlesnake (Crotalus viridis viridis). Toxicon 35: 1 11, 1997.
90. Renetseder, R., Brunie, S., Dijkstra, B. W., Drenth, J. and Sigler, P. B. A
comparison of the crystal structures of phospholipase A2 from bovine
pancreas and Crotalus atrox venom. J. Biol. Chem. 260: 11627, 1985.
91. Selistre de Araujo, H. S., White, S. P. and Ownby, C. L. Sequence analysis
108 TSAI
Toxicon 34: 1237, 1996.
92. Pedersen, J. Z., Arcuri, B. F., Morero, R. D. and Rufini, S . Phospholipase-
like myotoxins induce rapid membrane leakage of non-hydrolyzable ether-
lipid liposomes. Biochem. Biophys. Acta 1190: 177, 1994.
93. Li,
Y.,
Yu, B. Z., Zhu,H.,
Jain, M. K. and Tsai, M. D. Phospholipase A2engineering: structural and functional roles of the highly conserved active site residue aspartate-49. Biochemistry 33: 12724, 1993.
94. Shimohigashi, Y., Tani, A., Matsumoto, H., Kishino, J., Arita,
H.
and Ohno,M. Lysine-49 phospholipase A2 from Trimeresurus flavoviridis venom are
membrane-acting enzyme. J. Biochem. 118: 1037, 1995.
95. Suzuki, A., Matsueda, E., Yamane, T., Ashida, T., Kihara, H. and Ohno, M.
Crystal structure analysis of phospholipase A2 from Trimeresurus
flavoviridis (habu snake) venom at 1.5 A resolution. J. Biochem. 117: 730, 1995.
96. Tomoo, K., Ohishi, H., Doi, M., Ishida, T., Inoue, M., Ikeda, K., Hata, Y.
and Semejima, Y. Structure of acidic phospholipase A2 form the venom of
Agkistrodon halys blomhofii at 2.8 A resolution. Biochem. Biophys. Res.
Commun. 184: 137,1992.
97. Wang, X. Q., Yang, J., Gui, L. L., Lin, Z. J., Chen, Y. C. and Zhou, Y. C.
Crystal structure of an acidic phospholipase A2 form the venom of
Agkistrodon halys Pallas at 2.0 A resolution. J. Mole. Biol. 5: 669, 1996.
98. Scott, D. L., Achari, A,, Vidal, J. C. and Sigler, P. B. Crystallographic and
biochemical studies of the (inactive) Lys-49 phospholipase A2 from the
venom of Agkistrodon piscivorus piscivorus. J. Biol. Chem. 267: 22645,
1992.
99. Han, S. K., Yoon, E. T., Scott, D. L., Sigler, P. B. and Cho, W. Structural aspects of interfacial adsorption, a crystallographic and site-driected mutagenesis study of the phospholipase A2 from the venom of Agkistrodon
piscivorus piscivorus. J. Biol. Chem. 272: 3573, 1997.
IOO.Kilby, P. M., Primrose, W. U. and Roberts, G. C. Changes in the structure of
bovine phospholipase A2 upon micelle binding. Biochemistry 35: 935, 1995.
IOl.Berg, B. V. D., Terrair, M., Boelens, R., Dijkman, R., de Hass, G. H.,
Kaptein, R. and Verheij, H. M. NMR structures of phospholipase A2 reveal
conformational changes during interfacial activation. Nature Struct. Biol. 2:
PHOSPHOLIPASES A2 109 102. Davison, F. F. and Dennis, E. A. Evolutionary relationships and implications for the regulation of phospholipase A2 from snake venom to human secreted
forms. J. Mol. Evol. 31: 228, 1990.
103. Ogawa, T., Nakashima, K. L., Nobuhisa, I., Deshimaru, M., Shimohigashi,
Y., Fukumaki, Y., Sakaki, Y., Hattori, S. and Ohno, M. Accelerated evolution of snake venom phospholipase A2 isozymes for acquisition of
diverse physiological functions. Toxicon 34: 1229, 1996.
104. John, T. R., Smith, L. A. and Kaiser, I. I. Genomic sequences encoding the acidic and basic subunits of Mojave toxin: unusually high sequence identity
of non-coding regions. Gene 139: 229, 1994.
105.Gubensek, F. and Kordis, D. Venom phospholipase A2 genes and their
molecular evolution. In: Venom Phospholipase A2 Enzymes: Structure,
Function and Mechanism, edited by R. M. Kini, John Wiley & Sons, UK, p. 73, 1997.
106. Ogawa, T., Kitajima, M., Nakashima, K. I., Sakaki, Y., Ohno, M. Molecular
evolution of group I1 phospholipase AZ. J. Mol. EvoL 41: 867, 1995.
107.Johnson, L. K., Frank, S., Vades, P., Pruzanski, W., Lusis, A. J. and
Seilhamer, J. J. Localization and evolution of two human phospholipase A2
genes and two related genetic elements. In: Phospholipase A2, edited by P. Y.
K. Wong and E. A. Dennis, Plenum Press, New York, p. 17, 1990.
108.Ducance1, F., Bouchier, C., Tamiya, T., Boulain, J. C. and Menez, A.
Cloning and expression a cDNAs encoding snake toxins In: Snake Toxins,
edited by A. L. Harvey, Pergamon Press, New York,
NY,
p. 385, 1991.109. Seliatre De Araujo, H. S. and Ownby, C. L. Molecular cloning and sequence
analysis of cDNAs for metalloproteinases from broad-banded copperhead,
Agkistrodon contortrix laticinctus. Arch Biochem. Biophys. 320: 14 1, 1995. 110. Paine, M. J. I., Moura-da-Silva, A. M., Theakston, R. D. G. and Crampton, J.
M. Cloning of metalloprotease genes in the carpet viper (Echis pyramidum
leakeyi), further members of the metallopretease/disintegrin gene family. Eur.
J. Biochem. 224: 483, 1994.
1 1 l.Ishizaki, J., Hanasaki, K., Higashino, K. I., Kishino, J., Kikuchi, N., Ohara,
0. and Arita, H. Molecular cloning of pancreatic group I phospholipase A2
receptor.
J.
Biol. Chem. 269: 5897, 1994.112.Ancian, P., Lambeau, G. and Lazdunski, M. Multifunctional activity of the extracellular domain of the M-type (180 m a ) membrane receptor for
110 TSAI
113.Lambeau, G., Ancian, P., Nicolas, J. P., Beiboer, S. H. W., Moinier, D.,
Verheij, H. and Lazdunski, M. Structural elements of secretory
phospholipase A2 involved in the binding to M-type receptors. J. Biol. Chem.
270: 5534, 1995.
114.Lambeau, G., Ancian, P., Barhanin, J. and Lazdunski, M. Cloning and
expression of a membrane receptor for secretory phospholipase A2. J. Biol.
Chem. 269: 1575,1994.
115.Zvartch, E., Lambeau, G. and Lazdunski, M. Endocytic properties of the M-
type 180 kDa receptor for secretory phospholipase A2. J. Biol. Chem. 271 :
250, 1996.
116.Rowan, E. G., Pemberton, K. E. and Harvey, A. L. On the blockade of
acetylcholine-release at mouse motor-never terminals by P-bungarotoxin and
crotoxin. Br. J. Pharmacol. 100: 301, 1990.
1 17. Schmidt, R. R. and Betz, H. Cross-linking of P-gungarotoxin to chick brain
membranes. Identification of subunits of a putative voltage-gated K’ channel.
Biochemistry 28: 8346, 1989.
118.Benishin, C. G. Potassium channel blockade by the B subunit of
p-
bungarotoxin. J. Pharmacol. Exp. Ther. 38: 164, 1990.
1 19. Tzeng, M. C., Yen, C. H., Hseu, M. J., Tseng, C. C., Tsai, M. D. and
Dupureur, C. M. Binding proteins on synaptic membranes for crotoxin and
taipoxin, two phospholipase Az with neurotoxicity. Toxicon 33: 45 1, 1995.
12O.Krizaj, I., Rowan, E. G. and Gubensek, F. Ammodytoxin A acceptor in
bovine brain synaptic membranes. Toxicon 33: 437, 1995.
12 1. Krizaj, I., Faure, G., Gubensek, F. and Bon, C. Re-examination of crotoxin-
membrane interactions. Toxicon 34: 1003, 1996.
122. Bevan, P. and Hiestand, P. b-RTX. A receptor-active protein from Russell’s
Viper (Vipera russelli russeli) venom. J. Biol. Chem. 258: 53 19, 1983.
123.Slater, N. T., Freedman, J. E. and Larson-Prior, L. J. Russell’s viper venom
proteins: molecular probes for neurotransmitter receptors: a review. Comp. Biochem. Physiol. 91C: 51, 1988.
124.Freedman, J. E. and Snyder, S. H. Vipoxin. A protein from Russell’s viper
venom with high affinity for biogenic amine receptors. J. Biol. Chem. 256:
172,1981.
125. Tzeng, M. C. Interaction of presynaptically toxic phospholipase A2 with
PHOSPHOLIPASES A, 111
126.Tsai, I. H., Liu, H. C. and Chang, T. Toxicity domain in presynaptically
toxic phospholipase A2 of snake venom. Biochim. Biophy. Acta 916: 94, 1987.
127.Tsai, I. H. and Tzeng, M. C. N-terminal region of snake venom neurotoxic
phospholipase A2 is involved in its binding to presynaptic receptors. In:
Peptides: Chemistry and Biology, Proc. 12th Amer. Peptide Symp. edited by J. A. Smith and J. E. Rivier, Escom Sci., The Netherlands, p. 460, 1992.
128.Fletcher, J. E. and Rosenberg, P.The cellular effects and mechanisms of
action of presynaptically acting phospholipase A2 toxins. In: Venom
Phospholipase A2 Enzymes: Structure, Function and Mechanism, edited by
R. M. Kini, John Wiley & Sons, UK, p. 413, 1997.
129.Evans, H. J. and Kini, R. M. The anticoagulant effects of snake venom
phospholipase A2. In: Venom Phospholipase A* Enzymes: Structure, Function and Mechanism, edited by R. M. Kini, John Wiley & Sons, UK, p. 353, 1997.
130.Mounier, C. Franken, P. A., Verheij, H. M. and Bon, C. The anticoagulant
effect of the human secretory phospholipase A2 on blood plasma and on a cell-free system is due to a phospholipid-independent mechanism of action
involving the inhibition of factor Va. Eur. J. Biochem. 237: 778, 1996.
13 1. Yuan, Y., Jackson, S. P., Newnham, H. H., Mitchell, C. A. and Salem, H. H.
An essential role for lysophosphatidylcholine in the inhibition of platelet
aggregation by secretory phospholipase Az. Blood 86: 4 166, 1995.
132. Kini, R. M. and Evans, H. J. Effects of phospholipase A2 enzymes on platelet
aggregation. In: Venom Phospholipase A2 Enzymes: Structure, Function and Mechanism, edited by R. M. Kini, John Wiley & Sons, UK, p. 369, 1997.
133.Mebs, D. and Ownby, C. L. Myotoxic components of snake venoms: their
biochemical and biological activities. Pharmac. Ther. 48: 223, 1990.
134.Mel0, P. A. and Ownby, C. L. Different sensitivity of fast and slow-twitch
muscle to some snake venoms and myotoxins. Toxicon 34: 653, 1996.
135. Berryman, D. E. and Bensadoun, A. Site-directed mutagenesis of a putative
heparin binding domain of avian lipoprotein lipase. J. Biol. Chem. 268: 3372,
1993.
136. Lomonte, B., Moreno, E., Tarkowski, A., Hanson, L. A. and Maccarana, M.
Neutralizing interaction between heparins and myotoxin 11, a Lysine 49
phospholipase A2 from Bothrops asper snake venom. J. Biol. Chem. 269: