國立臺灣大學醫學院微生物學研究所 碩士論文
Graduate Institute of Microbiology College of Medicine
National Taiwan University Master Thesis
小分子合成物對於會引起心內膜炎的鏈球菌的生物膜 形成之抑制作用
Inhibition of endocarditis-inducing streptococcal biofilm formation by synthetic small molecules
陳宜婷 Yi-Ting Chen
指導教授:賈景山 博士 Advisor: Jean-San Chia, Ph.D.
中華民國 101 年 7 月
July, 2012
i
口試委員會審定書
ii
誌謝
兩年的碩士生活,真得是一轉眼就過了呢!這段日子,雖然走得跌跌撞撞,
但很幸運地,身旁有許多人陪伴著我、鼓勵著我,並隨時給予我必要的協助,讓 我得以完成這本碩士論文。首先,真的很感謝這兩年來所有指導我論文的老師們。
謝謝賈老師在平日的教導,無論是 seminar 的準備或實驗設計方面,總是給我很 多想法,讓我的眼界因此拓展了許多。另外,非常謝謝忻老師提供我小分子合成 物作為篩選材料,並一再地幫我補充所需要的藥量,讓我得以完成論文後半部的 動物實驗。也很感謝符老師、鄧老師和楊老師在 committee 時提供我許多寶貴意 見,讓我在研究上有改正和進步的機會。
實驗室的學姐們及夥伴們更是我完成論文的推力之一。感謝惠婷學姐、秋月 學姐和筱菁學姐在這段期間的大力幫助,不論在儀器的操作上、實驗的設計或是 報告前的 rehearsal,總是提供我許多協助。另外感謝鴻偉學長、李泱和學儒,因 為你們,我才得以學會動物實驗及許多跟細菌相關的基本操作。還有,一起打拼 的三位夥伴:杜杜、佳儒、昱璇,這兩年來咱們同甘共苦的時光將是我難忘的回 憶之一。謝謝小幫手佩青,最後這幾個月的動物實驗真是辛苦妳了!也謝謝 R1509 的其他夥伴:鏡文、派派、英哲、阿翔、Lori、高醫師和高高,因為你們,
整個實驗室總是洋溢著歡樂的氣氛。除此之外,感謝大學的好友們(冰箱、雨蓉、
志鴻)、杏林管弦的好夥伴們(恭仰、淑涵等)及微生物所上的好友群(婉倫、漢堡、
喬巴等)的大力相挺,你們的鼓勵,讓我更有力量面對每一天的挑戰。
最後,非常謝謝家人們的全力支持。感謝父母體諒我沒辦法長時間的陪伴,
也謝謝妹妹容忍我有時沒來由的任性。因為你們的支持和鼓勵,讓我這兩年可以 無後顧之憂的完成碩士學位。
iii
中文摘要
感染性心內膜炎(infective endocarditis)是一種高致死率、高復發率的心血 管感染疾病,主要由一些口腔中的鏈球菌例如:轉糖鏈球菌(Streptococcus mutans) 所引發。這些會引起心內膜炎的致病株可在受傷的瓣膜上形成生物膜(biofilm),
並與血小板、纖維蛋白(fibrin)及發炎細胞堆疊成贅疣(vegetation)。目前治療的 潛在問題是這些在贅疣中形成生物膜的致病株具有高抗藥性。先前實驗室曾利用 體外篩選模式(in vitro)模擬體內產生贅疣的狀況,證實血小板對於轉糖鏈球菌生 成生物膜是重要的、以及所生成的生物膜對於抗生素具有更高的抗性,因此有必 要進一步尋找一些小分子合成物來輔助抗生素抑制生物膜形成。本篇研究藉由體 外生成生物膜的方法,並搭配結晶紫(crystal violet)的染色,篩選出 CYY-X022, 47, 48, 58, 60 五種小分子合成物能夠抑制細菌形成生物膜,而 CYY-X011, 12, 22, 47, 52, 55, 56 則可以干擾由細菌和血小板所形成的生物膜的生成。進一步測定細菌 生長曲線,發現 CYY-X022, 47, 48, 58, 60 能有效地抑制細菌生長。利用血小板凝 集計(aggregometer) 分析血小板活性以及測定細菌和血小板之間交互作用的狀 況,發現 CYY-X011, 12, 22, 47, 52, 55, 56 能夠藉由干擾細菌和血小板之間的接觸 來抑制血小板凝集,進而抑制由細菌和血小板所形成的生物膜。在活體 (in vivo) 實 驗 中 , 利 用 實 驗 室 先 前 所 建 立 的 感 染 性 心 內 膜 炎 大 鼠 模 式 (experimental streptococcal endocarditis rat model),將 CYY-X022, 47, 52, 55, 56 分別注入大鼠體 內,發現能夠降低贅疣中生物膜生成的狀況。除此之外,當抗生素搭配小分子一 同使用,利用共軛焦顯微鏡的觀察以及菌落的計數,發現抑制效果比給予單一抗 生素處理還要好。本篇研究結果篩選出具有抑制生物膜生成的小分子合成物,希 望對於未來在感染性心內膜炎的治療上能夠提供重要的資訊,並且能夠在其他臨 床的研究上也有更多的應用。
iv
Abstract
Infective endocarditis (IE) is a cardiovascular disease with high mortality rate and usually caused by oral streptococci (such as Streptococcus mutans) infection. The characteristic of IE is the formation of vegetations, fibrin-platelet clots with the embedded bacteria forming biofilm, which is refractory to routine antibiotic treatment.
Previously, our data reported that platelets play important roles in vegetation formation and could enhance the resistance of streptococcal biofilm to antibiotics.
Therefore, to search novel prophylactic agents that specifically target the platelet-associated biofilm will provide effective strategy for the successful control of IE. In this study, 76 different synthetic molecules have been screened in vitro by using biofilm formation assay and crystal violet staining. We found that CYY-X022, 47, 48, 58 and 60 could eliminate homotypic bacterial biofilm formation, and CYY-X011, 12, 22, 47, 52, 55 and 56 could interfere with platelet-associated biofilm formation.
Among these effective synthetic molecules, CYY-X022, 47, 48, 58 and 60 could inhibit the bacterial growth , and CYY-X011, 12, 22, 47, 52, 55 and 56, alternatively, could interfere with the binding of S. mutans and platelets, as well as inhibiting the streptococci-induced platelet aggregation according to the results of platelet aggregation test. Consistent with the in vitro data, the vegetation size was decreased by intravenous administration of CYY-X022, 47, 52, 55 and 56 in the experimental streptococcal endocarditis rat model. Moreover, antibiotics combined with the targeted small molecules in vitro could effectively inhibit the biofilm formation based on the results of confocal laser scanning microscope observation and the measurement of survival bacteria. Taken together, the data suggest that these synthetic small molecules could eliminate streptococcal biofilm formation by inhibiting the bacterial
v
growth or the binding of S. mutans and platelets. For clinical treatment on infective endocarditis, these targeted small molecules may have promising applications and offer another strategy.
vi
Table of Contents
口試委員會審定書 ... i
誌謝 ... ii
中文摘要 ... iii
Abstract ... iv
Chapter 1: Introduction ... 1
1.1 Streptococcus mutans ... 1
1.2 Infective endocarditis (IE) ... 2
1.3 The role of platelets in IE model ... 3
1.4 Prophylaxis treatment ... 5
1.5 Biofilm dispersal ... 6
1.5.1 Dispersal factors ... 7
1.5.2 D-amino acids ... 8
1.5.3 Small molecules ... 9
1.6 Specific aim ... 10
Chapter 2: Materials and Methods ... 12
2.1 Bacteria strains and culture ... 12
2.2 Preparation of human platelets ... 12
2.2.1 Platelet-rich plasma (PRP) ... 12
2.2.2 Platelet-poor plasma (PPP) ... 13
vii
2.2.3 Platelet suspension (PS) ... 13
2.3 Preparation of small molecules ... 13
2.4 Biofilm formation assay ... 14
2.5 Bacteriostatic analysis ... 15
2.6 Platelet aggregation test ... 16
2.7 Bacteria-platelet interacting assay ... 17
2.8 Experimental streptococcal endocarditis rat model ... 18
2.8.1 Experimental schedule design ... 18
2.8.2 Preparation for bacteria, small molecules and anesthetic 19 2.8.3 Cardiac catheterization ... 19
2.8.4 Quantification and observation ... 19
2.8 Confocal laser scanning microscopy (CLSM) analysis ... 20
Chapter 3: Results ... 21
3.1 Identification of streptococci-platelet biofilm specific inhibitors ... 21
3.2 The correlation between small molecules concentration and bacterial number in inhibition effect ... 22
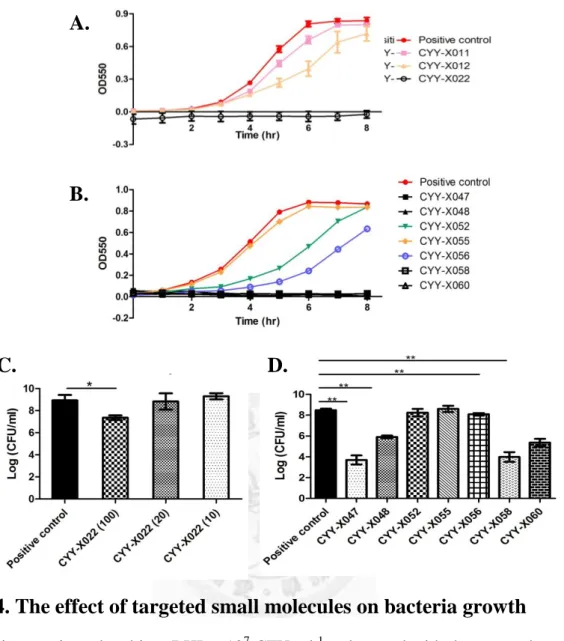
3.3 The effect of targeted small molecules on bacteria growth ... 23
3.4 Inhibition effect of targeted small molecules on bacteria-induced platelet aggregation ... 24
3.5 Inhibition effect of targeted small molecules on bacteria adherence to platelets ... 25 3.6 Inhibition effect of targeted small molecules on biofilm
viii
formation in rat endocarditis model ... 27
3.7 Synergistic effect with antibiotics ... 28
Chapter 4: Discussion ... 29
4.1 Summary ... 29
4.2 The inhibiting effect of the targeted small molecules on biofilm formation ... 31
4.3 Application of small molecules as biofilm dispersal factors ... 35
Chapter 5: References ... 37
Chapter. 6 Figures ... 43
Fig. 1. Identification of streptococci-platelet biofilm specific inhibitors by crystal violet staining. ... 44
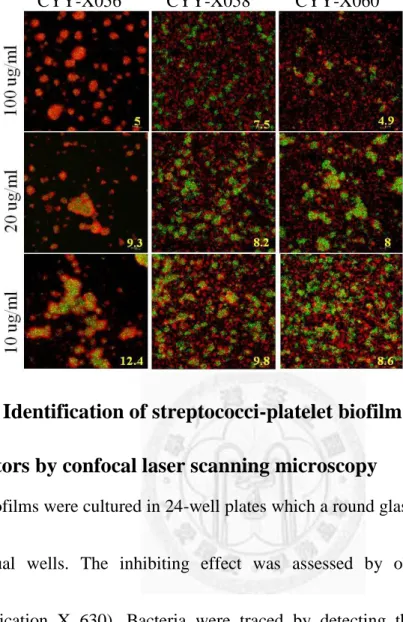
Fig. 2. Identification of streptococci-platelet biofilm specific inhibitors by confocal laser scanning microscopy... 46
Fig. 3. The correlation between small molecules concentration and bacterial number in inhibition effect ... 49
Fig. 4. The effect of targeted small molecules on bacteria growth .. 50
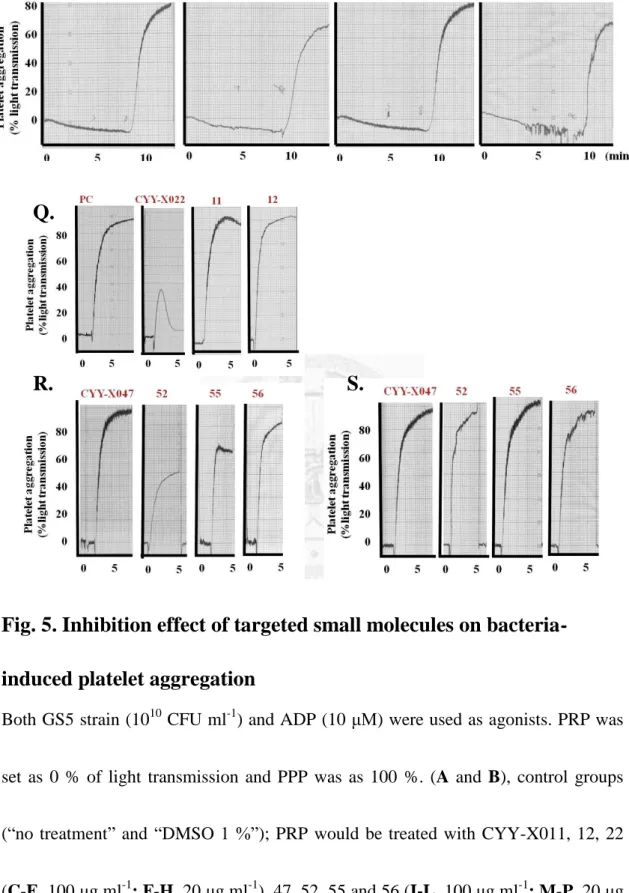
Fig. 5. Inhibition effect of targeted small molecules on bacteria-induced platelet aggregation ... 52
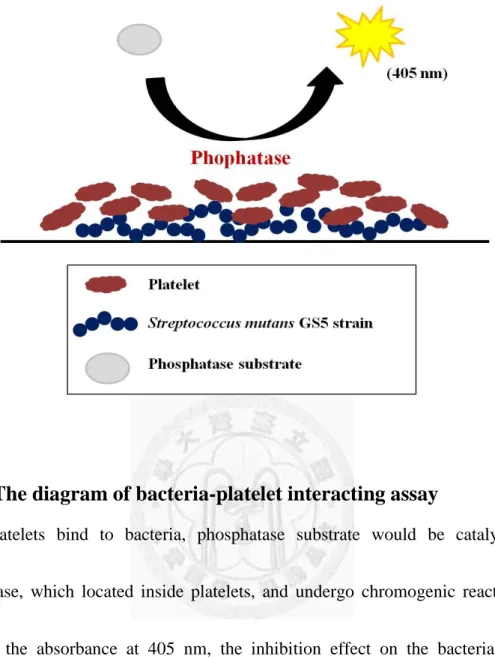
Fig. 6. The diagram of bacteria-platelet interacting assay ... 53
Fig. 7. Inhibition effect of targeted small molecules on bacteria adherence to platelets ... 55 Fig. 8. Inhibition effect of targeted small molecules on biofilm
ix
formation in rat endocarditis model ... 57 Fig. 9. Synergistic effect with antibiotics ... 59
Chapter. 7 Appendix ... 60
Appendix. 1. The pathogenesis of streptococcal-induced infective endocarditis. ... 60 Appendix. 2. The mechanism of S. aureus-mediated platelet
activation ... 61 Appendix. 3. The mechanisms of oral bacteria-induced platelet
activation ... 62 Appendix. 4. The dispersal factors ... 62
1
Chapter 1: Introduction
1.1 Streptococcus mutans
S. mutans was first identified by J Kilian Clarke in 19241and also completely
sequenced in 2002. It is a Gram-positive coccus-shaped organism, belonged into
viridans streptococci, and usually lives in facultatively anaerobic environment such as
the niche of teeth in oral cavity. They are also an important contributor to tooth decay.
By metabolizing sucrose to lactate, they create an acidic environment in mouth. As
time progresses, the acid makes the portions of the mineral content at the surface of
teeth dissolved and then causes dental caries2.
These oral streptococci must adapt to the adverse circumstances in oral cavity.
By equipping some specialized surface proteins, these bacterial could adhere onto the
teeth and aggregate as biofilm to form plaque. Antigen I/II and GTF, a
glucosyltransferase, the two proteins play an important role in adhesion and biofilm
formation by S. mutans. In the absence of sucrose, they use Antigen I/II, a cell surface
fibrillar protein, to anchor the surface of teeth at the initial adhesion3, 4. On the other
hand, GTF is responsible for synthesizing the glucan polymers which is related to
binding and biofilm formation in the present of sucrose5.
2
It has been reported that biofilms are involved in many microbial infection.
Biofilms are the structure that bacteria aggregate layer by layer, encased in an
extracellular matrix such like exopolysaccharides, proteins, extracellular DNA and
amyloid fibers, etc6-8. By forming biofilms, S. mutans could resist phagocytosis from
host immune cells and the threats from antibiotics. It makes difficulties in treatment9.
As many clinical reports, it is also known that biofilm formation by S. mutans is
strongly associated with dental plaque and infectious endocarditis2, 4, 5, 10
. Although a
lot of studies about the prevention or therapy have been published, the efficacy still
needs to be improved.
1.2 Infective endocarditis (IE)
Infective endocarditis (IE) is a form of cardiovascular disease. As a subacute
type that progresses more slowly than acute IE, it is usually caused by oral
commensal streptococci. In Taiwan, S. mutans is one of the prevalent infectious
agents. These oral microorganisms gain access to the vascular system through trauma
by surgery or other physics forces and cause the transient bacteremia11. If the
endocardial surface is abnormally roughened, it may offer a foothold for the
adherence. By the binding of fibrin-platelet clots and embedded bacteria layer by
3
layer, the vegetation with characterized by a firm architecture is made up10 (Appendix.
1). Bacteria in vegetation have higher resistance to antibiotics and could escape the
phagocytosis since that the dense nature of vegetation may restrict the migration of
phagocytes. Although IE is uncommon, it may have serious complications such like
stroke due to the embolism caused by dissection of vegetation12, 13. According to the
clinical reports, the frequency in recurrence and mortality rate are even up to 40% and
30% respectively. Unfortunately, there is still no appropriate way on prophylaxis and
diagnosis. .
1.3 The role of platelets in IE model
In IE model, the pathogenesis is complex. Briefly, the whole process could be
dissected into three parts: the oral bacteria accessing the circulation, escaping the
surveillance of immune cells and colonizing on valves to form vegetation10, 14. In our
previous study, we found that S. mutans would change the gene expression and
phenotype when exposure to human plasma at a low concentration (< 10%)15. They
can even bind to the components in plasma such as fibronectin to help them resist
phagocytosis14. Furthermore, the microbes can also interact with platelets through
their surface proteins or plasma components to induce platelet aggregation16.
4
When platelets (PLTs) are activated, they undergo aggregation and form
thrombus to plug the damaged endothelium. Bacteria take the advantage of that. By
interacting with specific surface receptors of PLTs to induce aggregation, they
promote the colonization at host tissue17 (Appendix. 3). In the case of staphylococci, S.
aureus could use fibrinogen to bridge between ClfA (clumping factor A) and
GPIIb/IIIa combined with immunoglobulin binding and induce the signal transduction
to stimulate platelets aggregation18 (Appendix. 2)16. In addition to fibrinogen, a fibronectin bridge may also take a part in S. aureus-induced platelet aggregation
(Appendix. 2). Similar observation could be made in streptococci. In the presence of
specific immunoglobulin to rhamnose-glucose polymers, S. mutans could trigger
PLTs activation by using these cell wall polysaccharides19. Moreover, in S. mutans, it
was also reported that protein antigen c (PAc) could both participate in extracellular
matrix binding and PLTs aggregation20. In S. gordonii, SspA/SspB belonged to
Antigen I/II family proteins could have the effect on PLTs adhesion and aggregation21.
Also, results obtained from our previous report suggest that activated platelets could
enhance biofilm formation of IE-inducing streptococci22. These PLTs-bacteria
biofilms are more highly resistant to antibiotics and refractory to routine antibiotics
treatment. Collectively, activated PLTs with some plasma components can offer
5
circulating bacteria adherence and enhance the pathogenesis.
1.4 Prophylaxis treatment
The history of prophylaxis treatment for streptococci infections could be back to
the 1950s23. At that time, penicillin was used in prophylaxis strategies for preventing
the diseases caused by streptococci such as rheumatic fever and infectious
endocarditis. People who may have a risk of bacteremia would be administered high
dosage of penicillin in oral or intramuscular manner to maintain the effective level of
drug in blood. In 2007, American Heart Association (AHA) provided the recent
guidelines for prophylaxis treatment of IE24. They recommended that: patients with
the high risk situation (i.e. those have predisposing cardiac condition such like
complex cyanotic congenital heart disease or equipping prosthetic valves or those
procedures may cause magnitude of bacteremia) would need the prophylaxis
treatment. With regard to antibiotics choice, amoxicillin (2g) is more often used in the
prophylaxis since that it is absorbed well in gastrointestinal tracks. If patients would
be allergic to β-lactams, they can take clindamycin (600mg), azithromycin (500mg)
or other cephalexin family drugs instead9, 25. Also, the guidelines recommended that
patients should be administered these antibiotics in an hour before the surgery or other
procedure at the high risk26.
6
Although many guidelines about IE prevention have been established, there are
some arguments about prophylaxis27. First, in consideration of ethic issue, the
effectiveness of prophylaxis treatment has only been shown in animal model, lack of
actual scientific evidence in human beings. Furthermore, it is known that the level of
bacteremia is one of essential factors, however, our daily activities, such like tooth
brushing, may offer a great entry for microbe to cause transient bacteremia. So, it
seems that the prophylaxis should not only focus on occasional procedure, but also
take personal background into consideration, for example, the general hygiene. What
is more, the factors associated with high risk of IE death should need more studies so
that it could be applied into prophylactic strategies in the future.
1.5 Biofilm dispersal
Biofilms are the structure whicht bacteria aggregate layer by layer, encased in an
extracellular matrix. These compact three-dimensional communities have the ability
to resist phagocytosis and antibiotics treatment. The processes of biofilm formation
can be divided into three steps: microbe attachment, population growth and biofilm
maturation. However, scientists found that mature biofilm would undergo dispersal
stage and then release planktonic cells which could migrate to new environment and
7
build up another communities, as a “biofilm life cycle”. Every step in biofilm life
cycle is highly regulated. Recently, there are many studies focusing on the final
dispersal stage, especially the mechanisms of regulation. A range of signal cues or
effects, from environment and even bacteria oneself, have been found to participate in
biofilm dispersal28 (Appendix. 4). In addition, there are also more and more small
molecules or products derived from nature for targeting bacterial biofilm. But their
effectiveness needs more demonstrations
1.5.1 Dispersal factors
Nutrients, one of the effectors from environment, could regulate the biofilm
dispersal, correlated with increase or decrease concentration. Some bacterial species,
like Pseudomonas aeruginosa, may trigger biofilm dispersal in response to carbon
limitation29; however, Acinetobacter sp. str. GJ12 would be packed when
encountering starvation30. Similarly, changes in temperature or the level of oxygen
and nitric oxide may also take part in the regulation of biofilm dispersal. For example,
in response to hypoxia and low concentration of nitric oxide, in P. aeruginosa,
biofilm dispersal would be triggered and sequencely release sessile cells31.
Quorum sensing, well known in regulation of bacteria biofilm formation, also
plays an important role in dispersal. AHLs (acyl-homoserine lactones) and AIP
8
(autoinducing peptide), secreted by Gram-negative and Gram positive bacteria
respectively, has been reported that could induce biofilm dispersal by activating the
signal transduction (i.e. cyclic di-GMP signaling) and producing the enzymes or
surfactants to degrade the hard biofilm structure. Previously, quorum sensing peptides
were applied in the inhibition against biofilms. They used CSP (competence
stimulating peptides, belonged to quorum sensing systems)32 hybridized with activate
antimicrobial peptides to serve as specifically targeted antimicrobial peptides
(STAMPs)33against S. mutans biofilm. Other dispersal factors such like
polysaccharide-degrading enzymes and rhamnolipids n P. aeruginosa could trigger
biofilm disassembly under the certain stimuli34, 35. Interestingly, bacteriophages are
also linked to biofilm dispersal. They may provide degrading enzymes by inducing
the related genes expression36.
1.5.2 D-amino acids
Recently, it is shown that D-amino acids also participate in biofilm disassembly.
D-amino acids, produced in bacteria stationary phase, can influences peptidoglycan
composition to adapt to changing environmental conditions37, 38. In Bacillus subtilis,
these amino acids incorporate into the anchored site of TasA fiber (TasA, subunits of
amyloid fibers) to cell wall and consequently govern the cell wall even triggering
9
biofilm disassembly. To further prove the importance of D-amino acids, Ilana
Kolodskin-Gal and her coworkers carried out liquid chromatography-mass
spectrometry followed by L-FDAA to identify the composition of cultured medium.
They found that D-tyrosine, D-leucine, and D-methionine were present and effective
in regulation of dispersal. Moreover, they also used racemases mutant strains, which
could not produce D-amino acids, and the bacteria failed to drive biofilm disassembly.
These phenomena could be observed in other species, such as P. aeruginosa and S.
aureus. It seems to be a general strategy in bacteria population for regulating the
biofilm disassembly. In addition to D-amino acids, norspermidine, another dispersal
factor, could interact with exopolysaccharides directly and together with D-amino
acids on biofilm-inhibiting effect39.
1.5.3 Small molecules
Biofilms are correlated with many diseases, such as whooping cough, cystic
fibrosis or endocarditis, etc. Therefore, more and more small molecules have been
developed for targeting biofilm-associated disease or for prophylaxis.
Chang Liu et al. screened a focus library of nitrogen-dense marine alkaloid
compounds, about 506 small molecules, and they found that eight of them had
inhibition against S. mutans biofilm formation3. The inhibiting effect is mainly on
10
biofilm-associated genes, such like pac (adherence-associated protein), gtfB
(glucosyltransferase), gbpB (glucan-binding protein) and comDE (quorum-sensing
associated protein). Genetic evidence showed that these molecules could interfere
with the genes expression and protein production level at 0.94 uM. Also, these small
molecules could have effect on planktonic cell growth at 2 uM.
In Gram-negative bacteria, curli and type I pili are important in mediating
biofilm formation. Lynette Cegelski and her groups discovered that dihydro thiazolo
ring-fused 2-pyridone is pilicides40, 41. These molecules structurally interfere with the
Escherichia coli pilus chaperon-subunits complex and consequently inhibit the fibers
assembly. Next, they modified some functional groups, such as the exchange of
cyclopropyl group, and generated another compound, which is not only pilicidal
ability but also curlicidal that can inhibit CsgA (curli subunit) polymerization42.
1.6 Specific aim
The characteristic of IE is the formation of vegetations which is refractory to
routine antibiotic treatment. Our data reported that platelets could promote vegetation
formation and enhance the resistance of streptococcal biofilm to antibiotics22.
Therefore, to search prophylactic agents that specifically target the platelet-associated
11
biofilm will provide effective strategy for the successful control of IE.
Previously, many studies made an effort toward inhibiting bacterial biofilm.
They used D-amino acids38, modified small molecules3, 42 or STAMPs33 to eliminate
homotypic biofilm formation. However, there is no report about the inhibition against
streptococci-platelet biofilm formation. Therefore, the objectives in my study are that:
1. To achieve the goal of searching the ideal candidates which could target
platelet-bacterial biofilm formation by screening a range of small molecules in
vitro (provided from Dr. Ling-Wei Hsin );
2. To investigate the potential inhibiting mechanisms of these targeted small
molecules;
3. To examine the effect of targeted small molecules in rat endocarditis model;
4. To treat bacteria with antibiotics addition to targeted small molecules and examine
the synergistic effect in vitro.
These results suggest that these synthetic small molecules may offer another strategy
for clinical treatment on infective endocarditis.
12
Chapter 2: Materials and Methods
2.1 Bacteria strains and culture
S. mutans GS5 strains were used in this study. Bacteria were grown and
maintained in brain-heart infusion (BHI) broth (BD, Bacto) and agar plate (BD,
Difco). Strains were cultured at 37 °C under an anaerobic atmosphere with 95% N2
and 5% CO2 for 16-18 hours. For confocal laser scanning microscopy observation,
GFPuv-tagged GS5 strains were generated by transformed with the GFPuv-contained
shuttle plasmid (pPDGFPuv) and selected by spectinomycin (500 μg ml-1).
2.2 Preparation of human platelets
Whole blood was collected from health donors in the laboratory by using venous
blood collection tubes containing sodium citrate 3.2 % (BD Vacutainer® citrate tube).
2.2.1 Platelet-rich plasma (PRP)
For preparation of platelet-rich plasma (PRP), whole blood was manipulated by
centrifugation at 1200 rpm for 12 min at 25°C and collected the upper layer. This PRP
would be used in biofilm formation and bacteria-platelet interacting assay to mimic
the condition in vivo.
13
2.2.2 Platelet-poor plasma (PPP)
The platelet-poor plasma (PPP), used as blank in platelet aggregation assay, was
collected by centrifugation at 3000 rpm for 10 min at 25°C. The upper layer would be
clearer than PRP.
2.2.3 Platelet suspension (PS)
The platelets suspension (PS) would be prepared for analyzing the direct
interaction between platelets and bacteria. Briefly, PRP was added with heparin (10 U
ml-1; B. Braun Melsungen AG) and Prostaglandin E1 (1 μM; Sigma-Aldrich) and
centrifuged at 3000 rpm for 6-8 min. Collect the pellet and resuspended gently with
Tyrode solution (11.2 mM glucose, 136.8 mM NaCl, 11.9 mM NaHCO3, 2.8mM KCl,
1.1 mM MgCl2, 0.33 mM NaH2PO4, 1.0 mM CaCl2, and 3.5 mg bovine serum
albumin per ml; pH 7.35~7.4). Heparin (6.4 U ml-1) and PGE1 (1 μM) were further
added to the suspension, and again, centrifuged at 2500 rpm for 5-7 min. After that,
the pellet was repeatedly resuspended with Tyrode solution. The concentration of PS
would be suggested at 3 x 108 ~5 x 108 platelets per ml.
2.3 Preparation of small molecules
A range of small molecules were kindly provided from Associate Professor
14
Ling-Wei Hsin (Graduate institute of Pharmaceutical Sciences, College of Medicine,
NTU). These 76 different small molecules were dissolved in dimethyl sulfoxide
(DMSO; Sigma-Aldrich) at 10 mg ml-1. The test concentrations in vitro were 100, 20,
10 μg ml-1 diluted with double-distilled water. For experiment in animal model, the
concentration was used at 6 mg kg-1.
2.4 Biofilm formation assay
To screen the targeted small molecules, the inhibiting effect would be test by
using bacterial biofilm formation assay in 96-well U-bottom polystyrene microtiter
plates (Greiner bio-one, NO. 650101) and condition was set in normal nutrient broth
(BHI) or PRP. S. mutans GS5 strain, cultured overnight, was centrifuged (3000rpm,
10 minutes) for removing the upper cultured medium. Pellet was washed by 1X
phosphate buffer saline (PBS) and suspended by sonication (40W) for 5 minutes.
After that, bacteria suspension was prepared as the concentration of 109 CFU ml-1
(optical density: A550 adjusted to 1.5) in 1X PBS and then seeded into wells as a ratio
of 1 to 100 in BHI supplemented 1 % glucose or 1 to 10 in PRP. Simultaneously,
small molecules with (or without) antibiotics (gentamicin 20 μg ml-1; penicillin 0.5 μg
ml-1) were added with. The incubation time was 18-20 hours at 37°C. After incubation,
15
the upper liquid was removed and washed twice with distilled water. The 96-well
plates were dried for 45 minutes and then stained with crystal violet (0.05%) for 5
minutes. After rinsed twice, the plates were distained with acetone-ethanol (ratio of 1
to 4) for 1 hour. By detecting the absorbance at 550 nm using a MicroELISA reader
(Dynatech Corp), the inhibition ability on biofilm formation would be semi-quantified.
Each experiment would be repeated three times in triplicate. For the confocal
microscopy observation, the biofilms would be cultured in 24-well plate which a
round glass coverslip was put in individual wells. After incubation, the coverslips
were washed by 1X PBS and fixed with 2 % paraformaldehyde.
2.5 Bacteriostatic analysis
To examine whether targeted small molecules could inhibit bacterial growth, it was necessary to quantify the bacterial population size and plot the values as a growth
curve. As mentioned before, bacteria suspension was prepared and inoculated into
BHI at 107 CFU ml-1. The inoculation was set as the first timepoint (“0 hour”), and
optical density was determined by measuring the absorbance at 550 nm. Repeatedly,
record the absorbance every hour until the 8th timepoint, and the 9th data was
measured at the 24th hour. All the records would be display as a curve by using
16
GraphPad Prism 5.
2.6 Platelet aggregation test
Platelets aggregation test could examine the ability of platelets activation. After
adding the agonist such as adenosine diphosphate (ADP) into PRP, platelets were
activated and sequencely underwent aggregation. Once platelets aggregated, the level
of light transmission would be increased. By measuring the difference of light
transmission level between pre- and post-stimulation, it could determine the ability of
platelets activation.
In this study, both S. mutans GS5 strain and ADP were used as agonists. The
concentration of bacterial treatment was 1010 CFU ml-1 and ADP was 10 μM.
Un-stimulated PRP was set as 0 % of light transmission and PPP was as 100 %. The
analysis was performed as following. PRP incubated with small molecules (100 and
20 μg ml-1) was pre-warmed for 3 minutes and stirred at 900 rpm in a test cuvette at
37 °C. After adding the agonist (bacteria or ADP), the level of platelet activation
would be monitored continually by the photocell in Lumi-Aggregometer (Payton
Scientific)43. If targeted small molecules have the inhibiting effect on
bacterial-inducing aggregation, light transmission level would not change for 25-30
17
minutes at least after addition of bacteria.
2.7 Bacteria-platelet interacting assay
For investigating whether targeted small molecules could interfere with the
interaction between bacteria and platelets, the experiment would be used as a
quantification assay. Bacterial cultured overnight were centrifuged at 3000 rpm for 10
minutes to remove the cultured medium, and the pellet was washed by 1X PBS. Again,
bacterial suspension was centrifuged and suspended by sonication (40 W) with
ELISA coating buffer instead. Next, bacteria were prepared as the concentration of
109 CFU ml-1 (optical density: A550 adjusted to 1.5) and then coated onto the 96-well
plates (Nunc MaxiSorp® flat-bottom 96 well plate) at 4 °C overnight. The upper
liquid part was removed and the plats were rinsed 1X PBST (1X PBS supplemented
with 1% Tween-20) once. Additionally, the wells were blocked by 1% bovine serum
albumin (BSA). The targeted small molecules were diluted with PRP
(indirect-binding assay) or PS (direct-binding assay), and then these samples were
added into wells which had coated with bacteria. The plates were incubated at 37°C
for 2 hours and sequencely washed by 1X PBST once again. Next, phosphatase
substrate (1 tablet per 5 ml buffer; Sigma-Aldrich) dissolved in specific buffer (0.1M
18
Na-acetate addition of 0.1% Triton-X100; pH 5.5) was added into wells, and the
plates were incubated at 37°C. If platelets bind to bacteria, phosphatase substrate
would interact with phosphatase, which located inside platelets, and undergo
chromogenic reaction (Fig. 6). By detecting the absorbance at 405 nm, the inhibition
effect on the bacteria-platelet interaction would be quantified.
2.8 Experimental streptococcal endocarditis rat model
All animal experiments in this study were approved by National Taiwan
University Institutional Animal Care and Use Committee.
2.8.1 Experimental schedule design
At the first day, rats were operated with cardiac catheterization. After 24 hour,
small molecules (I.V.) or Aspirin (I.P.) would be injected into these animals. 30
minutes later, these rats were treated with bacteria. For Aspirin (25mg kg-1;
Sigma-Aldrich treatment), there would be two more injections at 30 minutes and 3
hours after bacterial infection. To analyze the effect of targeted small molecules on
bacteria survive in circulation, blood was collected at three timepoints: 30 minutes, 3
hours and 24 hours after bacteria treatment. At the third day, these rats were sacrificed
and their hearts were removed (Fig. 8A).
19
2.8.2 Preparation for bacteria, small molecules and anesthetic
Preparation for bacteria (GFP-tagged or normal GS5 strain) and small
molecules were described as previously. The amount of bacterial infection was 109
CFU and the concentration of targeted small molecules was 6 mg kg-1. The anesthetic
(“Zoletil 50”, Virbac) were supplemented with muscle relaxant (“Rompun”, Bayer
HealthCare) and water (for injection only) as a ratio of 500:280:220 respectively.
2.8.3 Cardiac catheterization
8-week-old male rats (Wistar) were used as ideal endocarditis rat model. A
stainless steel (10 cm) embedded into a polyethylene tube (8 cm; I.D. 0.28mm and
O.D. 0.61mm; Becton Dickinson, PE10) as a catheter which was further bent as 1/4 of
circle by a tweezers. This catheter was then inserted into carotid artery through an
incision on the chest and moved on to the left ventricle along the vessel. The catheter
would tremble more and more intensely with heartbeat when it was closed to the
aortic valves. When it no longer went forward by the blood flow resistance, the
catheter remained right there and was fixed by sutures.
2.8.4 Quantification and observation
For quantification of the effect on bacteria survive in circulation, blood was drew
and lysed by 1% Triton-X100 (in 1X PBS). The cell lysate was plated on BHI agar
20
with optimized dilutions. To determine the density of bacteria colonized in
vegetations, the vegetations were removed and their masses were weighed. By
sonication in 1X PBS, the homogenous suspension was further plated on Mitis
Salivarius agar (MS agar; BD, Difco) added with 20% sucrose. Both BHI and MS
agars were incubated at 37 °C for 2 days. The biofilms inside vegetations were also
visualized by confocal microscopy and their thicknesses were measured by vertical
section quantification.
2.8 Confocal laser scanning microscopy (CLSM) analysis
By using confocal laser scanning microscopy (CLSM; Leica TCS SP5), the
detailed composition of biofilm formation in wells and inside vegetation would be
observed. Biofilms cultured in wells were washed with 1X PBS and fixed with 2 %
paraformaldehyde. GFP-tagged bacteria were visualized by detecting emission spectra
of GFP. For platelets observation, on the other hand, samples which were incubated
with 1% Triton-X100 for 5 minutes and stained with rhodamine-conjugated phalloidin
(1 to 500 dilution; Invitrogen) were traced by detecting the emission wavelength at
565 nm. For the vegetation observation, it was performed as describes previously.
21
Chapter 3: Results
3.1 Identification of streptococci-platelet biofilm specific inhibitors
For searching the ideal candidates which target streptococci-platelet biofilm
formation, 76 different small molecules, provided from Associate Professor Ling-Wei
Hsin, were screened in vitro. By using crystal violet, biofilms were stained and the
formation activity was semi-quantified by detecting the absorbance at 550 nm.
Among these compounds, CYY-X022, 47, 48, 58, and 60 could obviously inhibit the
homotypic biofilm formation in BHI (Fig. 1A and 1C); as regards the
platelet-associated biofilm, CYY-X011, 12, 22, 47, 52, 55 and 56 had an inhibiting
effect on that (Fig. 1B and 1D). DMSO, the solvent for small molecules, would not
have inhibition against biofilm formation (Fig. 1E and 1F). CYY-X058 and 60,
having an effect on normal biofilm formation (Fig. 1C), could also have partial
inhibition against platelet-bacterial biofilm (Fig. 1D). The absorbance upon the
treatment of CYY-X022 or 52 at 100 μg ml-1 was much higher than others (Fig. 1B
and 1D) since these compounds may precipitate with plasma components and cause
the pseudo-positive results (Fig. 1F).
The inhibition effect of these targeted small molecules was assessed by CLSM.
Compared with the controls (“no treatment” and “DMSO”) (Fig. 2A), the bacteria
22
aggregates in the groups treated with the targeted molecules were much smaller and
randomly distributed (Fig. 2B-D). The architectures were not compact anymore,
either. As the concentration decreased, the inhibition effect was diminished, with the
bacterial cluster becoming thicker and thicker. The biofilm thicknesses upon 100 μg
ml-1 of treatment was less than 10 μm in most treated groups except CYY-X022 (Fig.
2B-D) which may cause the precipitation with platelets and plasma components as
aggregates. These data indicate that some of small molecules may have ability to
inhibit biofilm formation, and, most importantly, seven of them could even target
platelet-associated biofilms.
3.2 The correlation between small molecules concentration and bacterial number in inhibition effect
To further find out the effective dosage of the targeted molecules, we analyzed
the correlation between molecules concentration and bacterial number. The bacterial
inoculation was manipulated by serial 10-fold dilutions (from 108 to 104 CFU) and
targeted small molecules, CYY-X047, 52, 55, and 56, were examined at five different
dosage: 100, 20, 10, 5, 1 and 0 μg ml-1. In BHI, CYY-X047 (100 μg ml-1) had broad
inhibiting effect under all different amount of inoculation (Fig. 3A); CYY-X052 and
23
56 (100 μg ml-1) would eliminate biofilm formation only when bacterial number was
below 105 CFU (Fig. 3B and 3D); CYY-X055 would still have no inhibition against
normal biofilm formation upon all of different bacterial amount treatment even at 100 μg ml-1 (Fig. 3C). In PRP, when bacterial amount was below 107 CFU, these four
molecules could inhibit the platelet-associated biofilm formation at five different
dosage (Fig. 3E-3H); however, when bacterial number was up to 108 CFU , the only
effective concentration would be 100 μg ml-1 (Fig. 3E-3H). These results indicated
that these molecules could inhibit biofilm formation with bacterial number below 107
CFU, and the most effective concentration would be 100μg ml-1. It suggested a
correlation between the molecules dosage and bacterial number for effective
treatment.
3.3 The effect of targeted small molecules on bacteria growth
At the first screening experiments, it was found that some of targeted small
molecules at 100 μg ml-1 could eliminate normal biofilm formation in BHI condition.
Therefore, we hypothesized that these molecules may have ability to inhibit bacteria
growth. To examine this hypothesis, bacteria were treated with the targeted small
molecules at 100μg ml-1 and the population of survival bacteria was quantified by
24
analyzing the changes in turbidity every hour. CYY-X055 had no inhibition against
cell growth (Fig. 4B). CYY-X011, 12, 52, and 56 partially interfered with the bacteria
growth; after 8 hours incubation, however, the inhibition would be compensated (Fig.
4A and 4B). The other targeted small molecules, CYY-X022, 47, 48, 58 and 60 had
an obvious inhibiting effect on bacteria growth (Fig. 4B), and this inhibition could
maintain for 24 hours (data not shown). These results were further analyzed by colony
counting. Compared with positive control (DMSO 1%), there was a significant
difference in the groups treated with CYY-X022, 47, 48, 58 and 60 at 100 μg ml-1
(Fig. 4C and 4D). Interestingly, these five molecules also inhibited homotypic biofilm
formation in BHI. The results revealed that these five molecules could interfere with
biofilm formation due to their ability to inhibit bacteria growth.
3.4 Inhibition effect of targeted small molecules on bacteria-induced platelet aggregation
In addition to having ability to inhibit homotypic biofilm formation, some of
them, such as CYY-X011, 12, 52, 55 and 56, could prevent the streptococci-platelet
biofilm forming (Fig. 1B and 5D), and some may even have dual abilities, like
CYY-X022 and 47 (Fig. 1A-1D). According to our previous results, platelets played
25
an important role in PRP biofilm formation22; moreover, the aggregation by induced
by bacteria was also another essential factor. Therefore, we supposed that these seven
molecules may inhibit PRP biofilm by interfering with bacteria-induced aggregation.
To prove the hypothesis, PRP incubated with targeted small molecules (100 and 20 μg
ml-1) for 20-30 minutes was added with agonist (GS5 strain and ADP) and then
examined the aggregation ability. Compared with control groups (Fig. 5A and 5B),
most PRP treated with the targeted molecules at 100 μg ml-1 did not aggregate within
25 minutes (Fig. 5C-E and 5I-L); however, as the concentration decreased to 20 μg
ml-1, the inhibition would be compensated, except CYY-X022 (Fig. 5F-H and 5M-P).
Considering these compounds may block all the physiological functions of platelets,
ADP stimulation was as a control (Fig. 5Q-S). After added with ADP, it was found
that the platelets supplemented with these molecules could aggregate as usual while
the magnitude upon the treatment of CYY-X022 (100 μg ml-1) was partially weaker.
According to above data, it indicated that CYY-X011, 12, 22, 47, 52, 55 and 56 could
inhibit platelet-associated biofilm through interfering with bacteria-induced
aggregation.
3.5 Inhibition effect of targeted small molecules on bacteria
26
adherence to platelets
In previous studies, it had been pointed out that S. mutans could induce platelet aggregation through its specialized polysaccharides, proteins19, 20 or the components
in plasma. To further explore the underlying mechanism of the inhibition against
platelets aggregation, it was assessed by the bacteria-platelet interacting assay (Fig. 6).
Compared with control groups, most PRP treated with the targeted molecules at 100 μg ml-1could not bind to bacteria significantly except CYY-X052 and 56, which
represented a partial inhibiting effect (Fig. 7A and 7B).With the dosage of treated
molecules decreased, the inhibiting effect would diminish. These data exhibited that
in PRP which was abundant in plasma proteins these targeted small molecules could
interfere with the indirect binding of streptococci and platelets, and the inhibiting
effect was dose-dependent. To analyze the effect on direct binding, PS purified from
PRP was pretreated with these small molecules and added into the wells coated with
bacteria. As the results revealed, CYY-X012, 47 and 52 at 100 μg ml-1 could interfere
with the direct binding while others had a slightly inhibiting effect (Fig. 7C and 7D).
The above results indicated that these small molecules had ability to inhibit bacteria
adherence to platelets.
27
3.6 Inhibition effect of targeted small molecules on biofilm formation in rat endocarditis model
Next, the inhibiting effect of these targeted small molecules on biofilm formation
was verified by using the experimental streptococcal endocarditis rat model. Rats
were infected with bacteria after pretreated with the targeted molecules (CYY-X012,
47, 52, 55 and 56) or Aspirin (Fig. 8A). According to the CLSM images, it was found
that compared with the controls (“no treatment” and “DMSO 1 %”) the biofilm
thickness of the treated groups decreased obviously (less than 40 nm) and the
structure also lost the multi-layer characteristic in vertical section (Fig. 8B). The
density of colonized bacteria in the groups treated with the targeted molecules,
however, decreased slightly (Fig. 8D) and the biomass of vegetation had no
significantly difference, either (Fig. 8E). To further investigate the effect on bacterial
survival in bloodstream, the blood was collected at the 0.5th, 3rd and 24th hour. The
data revealed that these targeted small molecules had no obvious inhibition against
bacteria survival in circulation (Fig. 8D).Taken together, CYY-X012, 47, 52, 55 and
56 could interfere with the biofilm formation in injured valves but have limited
inhibition on bacteremia and the density of colonized bacteria inside vegetations.
28
3.7 Synergistic effect with antibiotics
An attempt was made to further analyze whether antibiotics combined with these
targeted molecules could have more effective inhibition against platelet-associated
biofilm formation. Bacteria treated with antibiotics (penicillin and gentamicin) and
the small molecules (CYY-X011, 12 and 22) were seeded into 24-well plates. In the
CLSM images, it was found that the biofilms architecture in the treated groups
became more loosed and small-aggregated (Fig. 9B) compared with the untreated and
antibiotic-treated-only groups (Fig. 9A). With the dosage of the small molecules
decreased to 20 μg ml-1, the inhibiting effect would be diminished. For the effect on
bacterial survival in PRP biofilm, the results showed that CYY-X022 (100 μg ml-1)
addition to penicillin and gentamicin respectively could significantly inhibit the
bacterial survival (Fig. 9C and 9D), and the inhibiting effect would sustain even the
treated dosage reduced to 20 μg ml-1 ( gentamicin treated group). These data revealed
that CYY-X022 could assist antibiotics with inhibiting both the streptococci-platelet
biofilm formation and the bacterial survival in PRP biofilm, suggesting the role of
synergistic effect.
29
Chapter 4: Discussion
4.1 Summary
IE, an infectious disease with a high mortality rate, is characterized by the
formation of vegetations, fibrin-platelet clots with the embedded bacterial biofilm.
This firm architecture exhibiting highly resistant to antibiotics treatment makes it
difficult on the clinical management. Additionally, the prophylactic treatment to
prevent IE is also controversial. Previously, our data reported that platelets could
promote the vegetation formation and enhance the resistance of streptococcal biofilm
to antibiotics. Therefore, searching novel prophylactic agents that specifically target
the platelet-associated biofilm will provide effective strategy for the successful
control of IE.
In this study, 76 synthetic small molecules were screened and examined. These
results are summarized as following:
1. To achieve the goal of searching the ideal candidates which could target
platelet-bacterial biofilm formation by screening a range of small molecules in
vitro (provided from Dr. Ling-Wei Hsin );
Among the 76 synthetic molecules, it was found that CYY-X022, 47, 48, 58 and
60 could eliminate homotypic bacterial biofilm formation, and CYY-X011, 12,
30
22, 47, 52, 55 and 56, alternatively, could interfere with platelet-associated
biofilm formation. And 100 μg ml-1 was suggested as an effective concentration.
2. To investigate the potential inhibiting mechanisms of these targeted small
molecules;
By interfering with the bacterial growth, CYY-X048, 58 and 60 were able to
inhibit biofilm formation. On the other hand, CYY-X011, 12, 52, 55 and 56 may
reduce the platelet-associated biofilm formation through interfering with the
binding of streptococci and platelets. CYY-X022 and CYY-X047 had dual
abilities to inhibit both the bacterial growth and the interaction between bacterial
and platelets.
3. To examine the effect of targeted small molecules in rat endocarditis model;
Consistent with the in vitro data, the vegetation size was decreased by
intravenous administration of CYY-X022, 47, 52, 55 and 56 in the experimental
streptococcal endocarditis rat model; however, the inhibiting effect on bacteremia
and the colonized bacteria inside vegetations was limited.
4. To treat bacteria with antibiotics addition to targeted small molecules and examine
the synergistic effect in vitro;
Antibiotics combined with CYY-X011, 12 and 22 respectively could effectively
31
target the platelet-associated biofilm, suggesting that the role of synergistic effect
in vitro.
Taken together, these results indicated that these targeted synthetic small molecules
could eliminate streptococcal biofilm formation by inhibiting the bacterial growth or
the binding of bacteria and platelets.
4.2 The inhibiting effect of the targeted small molecules on biofilm formation
In this study, we found that some of small molecules could inhibit the biofilm
formation through different mechanisms, such as interfering with bacterial growth or
the indirect (or direct) binding of bacteria and platelets. However, some side effects
need to be solved:
1. In the in vitro biofilm formation assay, CCY-X022 and CYY-X052 precipitated in
plasma as small aggregates surrounded by rhodamine-tagged platelets (Fig. 1F
and 2B). It indicated that the two molecules were slightly hydrophobic and may
precipitate with plasma components in PRP. The functional group of these
molecules will be modified additionally for more effective inhibition against PRP
biofilm formation.
32
2. In the animal model, the thickness of the biofilms inside vegetation was decreased
after prophylactic treatment of targeted small molecules. However, after
pretreated with CYY-X047, 52, 55 and 56 by intravenous injection, these rats
performed hematuria within 30 minutes. Also, CYY-X022 could interfere with the
magnitude of normal platelet aggregation slightly in response to ADP (Fig. 5Q).
The cytotoxicity-associated issue would need further investigation.
The effective concentration of these targeted molecules was 100μg ml-1
according to the in vitro and in vivo experiments (Fig. 1, 2 and 8). Based on the data
in this study, it was shown that when bacterial number below 107 CFU, these targeted
molecules, even at 1 μg ml-1, could perform effective inhibition against
streptococci-platelet biofilm formation (Fig. 3E-3H). However, in biofilm formation
assay and animal experiments, the number of the bacteria treatment was up to
108-1010 CFU, for the GFP tracing in CLSM observation. In regard to clinical cases,
the amount of bacterial infection would be less than 108 CFU per exposure. Therefore,
the effective concentration of the molecules could be adjusted to a lower dosage.
According to the result, it indicated that CYY-X048 could effectively inhibit the
bacterial growth. Nevertheless, it still could not eliminate the platelet-associated
biofilm formation. On the other hand, CYY-X022 and CYY-X047 had more effective
33
inhibition against biofilm formation owing to equipping with dual abilities to inhibit
both bacterial growth and the interaction between bacteria and platelets. Together, it
revealed that uni-target of inhibition was not enough to eliminate the firm architecture
of the complex biofilm.
Aspirin, an anti-platelet drug44, was reported to be applied in pre-treatment of IE
patients before the disease onset45. This strategy would be associated with a lower risk
of embolism. Consistent with our published data, we also found that Aspirin could
inhibit S. mutans-induced platelet aggregation and reduce the platelet-biofilm
formation in rat experimental model and in vitro assay22. Similar results in animal
model were observed in this study (Fig. 8B). Compared with Aspirin, the targeted
small molecules had comparable inhibiting effect on platelet-associated biofilm
formation (Fig. 8B) since that both of Aspirin and those targeted molecules have the
ability to eliminate the bacteria-induced platelet activation (Fig. 5).
Besides making an effort on searching effective prophylactic agents, we also
attempted to explore the potential mechanisms. We found that some of the molecules
(CYY-X048, 58 and 60) could target the bacterial growth, and some (CYY-X011, 12,
52, 55 and 56) may alternatively have inhibiting effect on the interaction between
bacteria and platelets. What’s more, some of them (CYY-X022 and 47) could have
34
dual abilities to target both bacterial growth and platelet aggregation. However, the
detail about the inhibiting mechanism remains unclear. There may be a range of
possible targets for eliminating biofilm formation. In genomic level, the molecules
could inhibit the biofilm-associated genes expression in planktonic cell, such as gtfB,
gbpB or comDE and then cause the inhibition against the bacterial biofilm formation;
or, in proteomic level, the production of adhesion-associated proteins, such like GTF
and Antigen I/II, may be blocked by these small molecules so that bacteria could not
adhere to platelets and build a firm biofilm anymore. Others like metabolic or
signalling-associated proteins, i.e. DAG (diacylglycerol), may be the potential targets.
DAG and DAG kinase, ubiquitous in proeukaryotic and eukaryotic cells, participate
in many essential signaling pathways, such like stress response and lipid recycling46-48,
etc. In proeukaryote, if these proteins have defect, there would be a severe impact on bacterial survival. In platelets, DAG plays an important role in activation. After
priming, DAGs are generated through Gq protein-PKC (protein kinase C) pathway
and sequencely induce the secondary aggregation49-51. Defect in DAG would fail to
secretion and aggregation in response to ADP and thrombin49. Therefore, we
hypothesis that : after cells uptake CYY-X022 and 47 through some membrane
transporters, these molecules may inhibit both bacterial growth and platelet
35
aggregation by targeting bacterial and cellular DAG. The additional studies may shed
light on the underlying mechanisms.
4.3 Application of small molecules as biofilm dispersal factors
In perspective of biological evolution, biofilm dispersal is beneficial for the
whole population maintenance. In response to the changes of nutrients level or the
competition, microcolonies in biofilm would be driven from mature to dispersal stage
through different signaling regulation, and these dispersal cells would move away
from the parent colonies to a new environment and form another communities. Taking
advantage of this characteristic, nowadays, scientists put it in use. They modified the
nature dispersal factors, trying to regulate biofilm life cycle instead of killing biofilm.
Previously, many studies made an effort toward the application of biofilm dispersal
factors. They used D-amino acids38, 39, which were produced in bacteria stationary
phase, to trigger biofilm disassembly. Other examples like synthesizing some
molecules, which could targeted some biofilm-associated genes3 or
adhesion-associated protein3, 42, to eliminate homotypic bacterial biofilm formation.
Based on the previous studies, bacteria-induced platelet aggregation was benefit for
bacterial survival16, 17. Moreover, according to our prior study, platelet could promote
36
the vegetation formation and enhance the resistance of streptococcal biofilm to
antibiotics in IE22. Therefore, the attempt of this study is to search some novel agents
that target the platelet-associated biofilm, especially the interaction between bacteria
and platelets. By interfering with the biofilm formation, bacteria, as planktonic-like
population, would become more sensitive to antibiotics. According to the data in this
study, we found that CYY-X022, which has multi-ring-fused structure with two short
chains, has better inhibition against platelet-associated biofilm. This may reveal that
the characteristic structure may play an important role in the inhibition and have
potential therapeutic value. However, for the clinical application, many aspects need
to be considered, such as solubility, cytotoxicity and immunogenic properties, etc.
Accordingly, the future studies are required to further probe into these important
issues.
37
Chapter 5: References
1. Clarke, J.K. On the bacterial factor in the aetiology of dental caries. Br J Exp Pathol 5, 141-147 (1924).
2. Hamada S, S.H. Biology, immunology, and cariogenicity of Streptococcus mutans. Microbiol Rev 44, 331-384 (1980).
3. Liu, C., Worthington, R.J., Melander, C. & Wu, H. A new small molecule specifically inhibits the cariogenic bacterium Streptococcus mutans in multispecies biofilms. Antimicrob Agents Chemother 55, 2679-87 (2011).
4. Mitchell, T.J. The pathogenesis of streptococcal infections: from tooth decay to meningitis. Nat Rev Microbiol 1, 219-30 (2003).
5. Kazuhiko Nakano, R.N., Takashi Ooshima. Streptococcus mutans and cardiovascular diseases. Japanese Dental Science Review 44, 29-37 (2008).
6. Flemming, H.C. & Wingender, J. The biofilm matrix. Nat Rev Microbiol 8, 623-33 (2010).
7. Hall-Stoodley, L., Costerton, J.W. & Stoodley, P. Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2, 95-108 (2004).
8. Abee, T., Kovacs, A.T., Kuipers, O.P. & van der Veen, S. Biofilm formation and dispersal in Gram-positive bacteria. Curr Opin Biotechnol 22, 172-9 (2011).
9. Que, Y.A. & Moreillon, P. Infective endocarditis. Nat Rev Cardiol 8, 322-36 (2011).
10. Moreillon, P. & Que, Y.A. Infective endocarditis. Lancet 363, 139-49 (2004).