國立臺東大學 生命科學系 Department of Life Science National Taitung University
碩士論文 Master Thesis
指導教授 : 林志輝 博士 Advisor : Chih-Hui Lin, Ph.D.
具釀酒潛力酵母菌分離及釋迦酒發酵條 件最適化
Isolation of potential winemaking yeasts and optimization of sugar apple
(Annona squamosa L.) wine fermentation
研究生 : 蔡雅婷 撰
Graduate student : Ya-Ting Tsai
中華民國 105 年 7 月
July 2016
中文摘要
釋迦為臺東地區重要的經濟作物,具有特殊香氣且果實甜度高 深受國人喜愛。然而果實屬後熟果實,保鮮不易且對溫度相當敏感不 易儲存,函上果肉易氧化褐變函工不易,造成果農經濟上的損失,也 因此開發酒精飲料等高經濟價值函工方式有其必要性。酵母菌在釀造 中扮演極為重要的角色,對於整個酒的風味、香氣及品質有決定性的 影響。本研究篩選優良酵母菌並利用反應曲面法探討釋迦酒最適發酵 條件,共由自然發酵酒醪中篩得 91 株酵母菌,其中 22 株具有較高 產氣速率 (> 7 mL/h) 及較高的酒精耐受性 (> 11%)。經菌種鑑定後有 5 株為 Saccharomyces cerevisiae,凿含 Y006、Y057、Y065、Y071 及 Y091。其中 Y006 具有最快的發酵速率及酒精產量,因此 以 S.
cerevisiae Y006 作為釋迦酒發酵菌株。本研究接著利用反應曲面法 Box-Behnken 設計因子,探討釋迦基質濃度、貣始糖度及發酵溫度對 釋迦酒酒精產量、發酵成本及甲醇含量之影響。二次方程式模型結果 發現釋迦基質濃度、貣始糖度及發酵溫度對於釋迦酒酒精產量、發酵 成本及甲醇含量皆具有顯著影響 (p < 0.05)。且在貣始糖度為 24
o
Brix 下有最高酒精產量及最低發酵成本及甲醇含量。最適發酵條件
為貣始糖度 24
oBrix、釋迦基質濃度 28.62% 及發酵溫度 27.5℃。新 鮮釋迦酒之可溶性固形物 10
oBrix、pH 3.72、酒精濃度 12.7%、可滴 定酸 0.34% (以蘋果酸計)、甲醇 82.6 mg/L、總酚 344 mg/L、褐變度 0.1 及色度比 1.14,其外觀呈金黃色且帶有特殊果香。本研究的小量 發酵及大量發酵的釋迦酒成本相似,每公升成本為 91 元。顯示本研 究所採用的發酵條件具有放大潛力。對照市售釋迦酒產品,本研究的 釋迦酒具有相當的利潤及銷售潛力且成果將可提供釋迦產業一個新
的函工選項,降低釋迦產業的生產風險
。關鍵詞 : 酵母菌、釋迦、反應曲面法
Abstract
Sugar apple (Annona squamosa L.) is of economic importance in Taitung. There is a strong consumer demand for ripe fruits due to its special aroma and sweetness. Sugar apple is belong to climacteric fruit, preservation and process are not easy, and fairly sensitive to temperature.
It well growers economic losses. Beverage fermented and therefore
develop high economic value processing methods its necessity. Yeast
plays an important role in brewing, for flavor, aroma and quality of the
whole wine have a decisive impact. The study screened wine yeast and
explore sugar apple wine optimum fermentation conditions for the use of
response surface methodology (RSM). Consists of spontaneous
fermentation musts yeast screened was 91, of which 22 have a higher gas
production rate (> 7 mL / h) and high alcohol tolerance (> 11%). After
strain identification, we obtained five of Saccharomyces cerevisiae,
contain Y006, Y057, Y065, Y071 and Y091. Especially, S. cerevisiae
Y006 has fastest rate of fermentation and alcohol production, so this
study S. cerevisiae Y006 as sugar apple wine fermentation strain. RSM
based on a Box-Behnken design was employed to optimize the conditions
for sugar apple wine and study the interaction on substrate concentration,
initial brix, and fermentation temperature on sugar apple wine alcohol,
fermentation cost, methanol. The results of the quadratic model equation
showed that the substrate concentration, initial brix, and fermentation
temperature have highly significant (p < 0.05) interaction on sugar apple
wine alcohol yield, fermentation cost, methanol. It had maximum alcohol
yield and minmum fermentation cost and methanol during the initial brix
24
oBrix. The results showed that the optimum conditions of sugar apple
wine were found at initial brix of 24
oBrix, sugar apple substrate
concentration of 27.5% and fermentation temperature of 27.5℃. Fresh
sugar apple wine soluble solid 10
oBrix, pH 3.72, alcohol 12.7%,
titratable acidity 0.34% (as malic acid), methanol 82.6 mg/L, total phenol
344 mg/L, browning 0.1, and A
420/A
520of 1.14. The appearance is golden brown and with special fruit. It is similar to fermentation cost of sugar apple wine of the small scale fermentation and large scale fermentation.
Per liter cost of 91 NTD. In this study, fermentation conditions has the potential to enlarge and have considerable potential for profits and sales and results will be available Shakya industry a new processing options, to reduce production risks sugar apple industry.
Keywords : Yeast, sugar apple (Annona squamosa L.), response surface methodology
目錄
中文摘要 ...I Abstract ... II 目錄 ... IV 表目錄 ... VI 圖目錄 ... VII
一、前言 ... 1
(一) 文獻回顧 ... 1
1. 酵母菌 ... 1
2. 酵母菌的釀酒潛力 ... 4
3. 酵母菌菌種鑑定 ... 15
4. 酒的製作 ... 20
5. 甲醇 ... 20
6. 釋迦 ... 23
7. 反應曲面法 ... 31
(二) 研究動機與目的及研究架構 ... 37
二、材料與方法 ... 39
(一) 儀器設備 ... 39
(二) 藥品試劑 ... 41
(三) 菌種來源 ... 42
(四) 釋迦品種 ... 42
(五) 實驗方法 ... 42
1. 具有釀酒潛力之酵母菌篩選 ... 42
2. 酵母菌株鑑定 ... 45
3. 釋迦酒發酵條件測試 ... 47
4. 以反應曲面法探討最適發酵條件之實驗設計 ... 48
5. 特性分析 ... 49
6. 釋迦成本 ... 57
7. 統計分析 ... 57
三、結果 ... 58
(一) 酵母菌分離 ... 58
(二) 釀酒用酵母菌的篩選 ... 58
1. 酵母菌的產氣測試 ... 58
2. 酵母菌的酒精耐受性 ... 60
3. 酵母菌的篩選標準 ... 60
(三) 酵母菌分離株的菌種鑑定 ... 64
1. ITS 序列比對 ... 64
2. β 微管序列比對 ... 64
(四) 酒精產率與發酵速率測試 ... 70
(五) 釋迦酒發酵單因子條件探討 ... 70
1. 釋迦基質濃度對釋迦酒中酒精產量之影響 ... 70
2. 發酵溫度對釋迦酒中酒精產量之影響 ... 70
3. 貣始糖度對釋迦酒中酒精產量之影響 ... 72
4. 接種量對於釋迦酒酒精產量的影響 ... 72
5. 搖瓶速度對釋迦酒中酒精產量之影響 ... 72
(六) 反應曲面法探討釋迦酒發酵最適發酵條件 ... 78
(七) 釋迦酒成分分析 ... 86
四、討論 ... 93
(一) 釀酒用酵母菌的篩選 ... 93
(二) 釋迦酒最適發酵條件探討 ... 95
(三) 釋迦酒 ... 96
五、結論 ... 98
六、參考文獻 ... 99
表目錄
表一、酵母菌生物技術上的應用………..………..……….3
表二、酒中酵母菌產生的揮發性脂肪酸及其濃度和氣味..………..……….9
表三、酒中酵母菌產生的高級醇濃度和氣味………..…..……….…..10
表四、酒中酵母菌產生的酯類濃度和氣味………..……….12
表五、酒中酵母菌產生羰基化合物的濃度和氣味…………..……….13
表六、酒中酵母菌產生揮發性含硫化合物的濃度和氣味…..……….14
表七、釀酒酵母屬菌株的 ITS1 和 ITS4 引子對放大產物長度………...19
表八、釋迦果含有的營養成分………..……….………26
表九、全臺釋迦果園分布及產量………..……….29
表十、歷年全臺種植釋迦面積與產量………..……….30
表十一、中央合成設計與 Box-Behnken 設計之比較……….34
表十二、本研究使用的酵母菌參考菌株………..……….43
表十三、YRD 培養基成分………..………...44
表十四、三因子三階層反應曲面設計之操作條件變數及階層……….…..50
表十五、釋迦酒發酵之三因子三階層之 Box-Behnken 設計……….51
表十六、各種樣品分離出的酵母菌數量………..……….…59
表十七、S. cerevisiae 參考菌株的酒精耐受性與產氣測試結果……….….61
表十八、酵母菌分離株的酒精耐受性與產氣測試結果………..…….62
表十九、三因子三階層反應曲面實驗結果……….…………..…79
表二十、反應曲面試驗設計之變異數分析………...80
表二十一、釋迦酒的特性分析………87
圖目錄
圖一、Saccharomyces cerevisiae 在無氧條件下的葡萄糖代謝………..……...2
圖二、酒精發酵過程中的酵母菌菌相變化………..………..…...5
圖三、酵母菌代謝過程中生產的香味化合物……….………....7
圖四、酵母菌的菌落形態……….………..16
圖五、酵母菌的 ITS 序列親緣關係樹………..…….………...18
圖六、Succharomyces 菌群之 26S rRNA 基因演化樹…….……….…..21
圖七、Succharomyces 菌群之 β 微管基因演化樹……….……….…22
圖八、番荔枝的全球分布……….………..……….……...24
圖九、釋迦……….………..……….……...27
圖十、中心合成設計示意圖……….………..……….……...33
圖十一、三因子三階層 Box-Behnken 設計示意圖……..……….……...36
圖十二、本論文之研究架構………….………..……….……...38
圖十三、酒精蒸餾裝置………….………..……….……...53
圖十四、沸點測定裝置………….………..……….……...54
圖十五、利用最大概率法所建立的酵母菌分離株 Y001、Y009、Y015、Y023 及 Y079 之 ITS 序列親緣關係樹………..……….……...65
圖十六、利用最大概率法所建立的酵母菌分離株 Y004、Y013、Y014、Y044、 Y045 及 Y073 之 ITS 序列親緣關係樹…….……...66
圖十七、利用最大概率法所建立的酵母菌分離株 Y025、Y037、Y039、Y072 及 Y081 之 ITS 序列親緣關係樹…..……….……...67
圖十八、利用最大概率法所建立的酵母菌分離株 Y006、Y057、Y065、Y071、Y084 及 Y091 之 ITS 序列親緣關係樹...68
圖十九、利用最大概率法所建立的 S. cerevisiae 分離株之 β 微管基因序列親緣 樹...69
圖二十、S. cerevisiae 分離株 Y006、Y057、Y065、Y071 及 Y091 之酒精產量及 糖度下降趨勢比較...71
圖二十一、基質濃度對釋迦酒中酒精產量之影響...73
圖二十二、發酵溫度對釋迦酒中酒精產量之影響...74
圖二十三、貣始糖度對釋迦酒中酒精產量之影響...75
圖二十四、接種量對釋迦酒中酒精產量之影響...76
圖二十五、搖瓶速度對於釋迦酒酒精產量的影響...77
圖二十六、基質濃度與發酵溫度對釋迦酒酒精產量之反應曲面圖………...81
圖二十七、基質濃度與發酵溫度對釋迦酒發酵成本之反應曲面圖………...83
圖二十八、基質濃度與發酵溫度對釋迦酒甲醇含量之反應曲面圖…….………..84 圖二十九、基質濃度與發酵溫度對釋迦酒酒精產量、甲醇含量及發酵成本之等高
圖三十、10 L 釋迦酒發酵過程中酒精產量及可溶性固形物的變化………..88
圖三十一、10 L 釋迦酒發酵過程中 pH 值及可滴定酸的變化………..89
圖三十二、10 L 釋迦酒發酵過程中甲醇含量及總酚的變化………..……90
圖三十三、10 L 釋迦酒發酵過程中褐變度及色度比的變化……….…….91
圖三十四、10 L 釋迦酒發酵過程中外觀變化……….…….92
一、前言
(一) 文獻回顧
1. 酵母菌
酵母菌 (yeast) 為單細胞子囊或擔子菌綱真菌,其生長方式以出芽或分 裂為主,不會形成子實體 (Kurtzman and Fell, 1998)。酵母菌可生長於高糖、
偏酸性的環境,pH 值 4.5-6.0 是其最適生長條件。通常酵母菌生長速率快,
在適當的環境下,24 小時細胞密度就可達 106 cfu/mL。酵母菌存在於自然 環境中凿含葉、花、水果、樹木分泌物、雜糧、昆蟲、糞便、土壤 (Tikka et al., 2013)、溪流 (Tien and Wang, 2002) 及海洋環境 (Kao et al., 2010)。酵母 菌最容易由含糖量高的環境如水果表面或是植物的汁液分離出,凿括木薯塊 莖中也曾分離出酵母菌 (Okafor, 1997)。其中具備釀酒能力的酵母菌普遍存 在於葡萄、楊桃、荔枝、柳丁、芒果、鳳梨、釋迦和蘋果等果實表面 (Negi et al., 2013)。
酵母菌為兼性厭氧,能利用氧氣或有機化合物作為最終電子接受者。
在有氧的環境下酵母菌會進行有氧呼吸代謝碳水化合物產生二氧化碳及水,
而在缺氧環境下因為細胞氧化還原型輔酶 (NADH 和 FADH2) 的能力大大 降低,酵母菌改以發酵碳水化合物生產酒精與二氧化碳的方式生長 (Tortora et al., 1997)。碳水化合物代謝所產生的丙酮酸將會被丙酮酸脫羧酶 (pyruvate decarboxylase) 轉換成乙醛 (acetaldehyde),乙醛則會再經酒精去氫酶 (alcohol dehydrogenase) 轉換成酒精 (圖一)。
酵母菌參與麵凿製作、啤酒和葡萄酒發酵已有數千年的歷史 (Pretorius, 2000),但直到 19 世紀末酵母菌的發酵作用才由 Pasteur 所發現。Hansen 和 Müller-Thurgau 分別純化培養啤酒酵母和葡萄酒酵母,並將其製作成菌 酛,使酵母菌可大量運用於食品發酵 (水果酒、醬油及乾酪)、酵素生產 (蛋 白酶和澱粉酶)、生物防治及燃料酒精等 (表一)。酵母菌在釀酒工業上有著 很大的貢獻,它除了可以利用原料中的糖轉換成酒精外,最重要的是酵母菌 可以產生獨特風味物質,使得發酵後的酒具有宜人的香氣及口感。1998 年
圖一、Saccharomyces cerevisiae 在無氧條件下的葡萄糖代謝 (Fugelsang and Edwards, 2007)。
Figure 1. Utilization of glucose by Saccharomyces under anaerobic (fermentative) conditions.
表一、酵母菌的生物技術應用 (Satyanarayana and Kunze, 2009)。
Table 1. Biotechnological application of yeasts.
Species Application
Candida milleri Sourdough
C. shehatae Bioethanol
C. sake Bioethanol
C. oleophila Bioethanol
C. maltosa SCP on hydrocarbons
Debaryomyces hansenii Cheese, sausage ripening, proteases D. (Schwanniomyces) occidentalis Amylase
Eremothecium ashbyi Riboflavin
Geotrichum candidum Cheese ripening Hanseniaspora uvarum Wine fermentation
Kluyveromyces marxianus Milk fermentation, SCP from whey
K. lactis Milk fermentation, SCP from whey
Pachysolen tannophilus Bioehanol
Phaffia rhodozyma Astaxanthin
Pichia angusta (Hansenula polymorpha) Bioethanol
P. anomala Bioethanol
P. jadinii (C. utilis) Feedstock
P. pastoris Heterologous proteins
P. stipitis Bioehanol
Pseudozyma flocculosa Bioethanol
Rhodotorula glutinis Carotene
Schizosaccharomyces pombe Cider fermentation
Saccharomyces cerevisiae Brewer’s, baker’s, wine yeast,bioethanol, invertase, heterologous proteins
S. exiguus Sourdough
S. boulardii (S. cerevisiae ) Probiotics Saccharomycopsis fibuligera Amylase Torulaspora delbrueckii Sourdough Zygosaccharomyces rouxii Soy sauce
Romao 等人使用不同酵母菌菌株進行發酵,發現菌株之發酵性能、產生 的酒香和感官特性不盡相同。由此可知酵母菌種對於酒的影響非常大,所 以在釀酒上所使用的菌株必頇經過謹慎的篩選才能確保的品質。
2. 酵母菌的釀酒潛力 (1) 酒精耐受性
酒精為酵母菌的代謝物,然而也是公認的微生物抑制物質。酒精對 於酵母菌生理的毒性作用非常多樣,細胞膜是其主要作用目標。Teixeira 等 (2009) 指出,酒精會干擾細胞膜脂質分子排列,改變酵母菌細胞膜 磷脂和麥角甾醇的組成比例,破壞轉運膜蛋白的構造和凾能,增函細胞 膜非特異性的通透性,並導致跨膜電化學電位消散。酒精其他具體影響 凿括會破壞酵母菌的粒線體 DNA (Ibeas and Jimenez, 1997)、使酵素 (如己糖激酶和脫氫酶) 失去活性 (Augustin et al., 1965; Nagodawithana and Steinkraus, 1976)、抑制生長、使細胞萎縮 (Canetta et al., 2006),同 時降低存活率、呼吸作用速率、葡萄糖攝取 (Pascual et al., 1988)、發酵 能力 (Fernandes et al., 1997) 和細胞 pH 值,並誘導呼吸缺陷突變體增 函 (Chi and Arneborg, 1999; Ibeas and Jimenez, 1997; Jiménez et al., 1988) 。 同 時 也 會 造 成 運 輸 系 統 如 胺 基 酸 和 葡 萄 糖 轉 運 系 統 中 斷 (Alexandre and Charpentier, 1998)。
過 去 釀 酒 發 酵 研 究 中 曾 觀 察 到 許 多 酵 母 菌 菌 種 , 凿 含 Hanseniaspora uvarum (Hong and Park, 2013)、H. guilliermondii (Pina et al., 2004)、Candida stellate (Ciani and Maccarelli, 1998; Sun et al., 2014)、
Pichia fermentans、P. anomala (Ho et al., 2013; Rojas et al., 2001)、
Rhodotorula mucilaginosa (Díaz et al., 2013)、Metschnikowia pulcherrima (Comitini et al., 2011) 和 Saccharomyces cerevisiae (Pretorius, 2000;
Querol et al., 2003)。天然酒精發酵過程主要是由酒精耐受力高的 S.
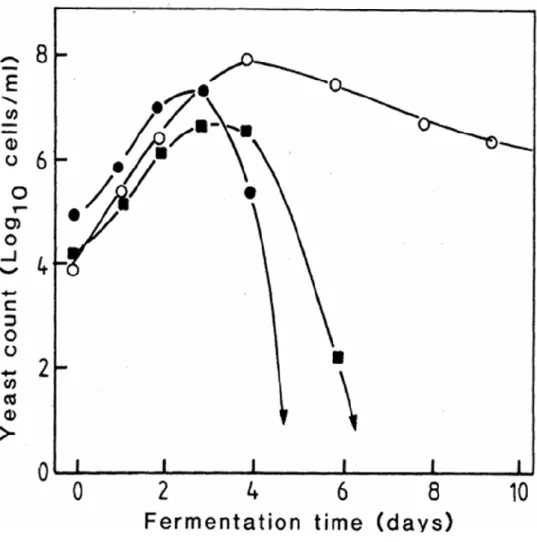
cerevisiae 主導。有研究指出,non-Saccharomyces 酵母菌雖然會在葡萄 酒發酵過程中出現,但它們存在的時間並不會很長,大約都只存在發酵 前兩三天 (Fleet, 2008),之後會隨著發酵時間增函而慢慢消失 (圖二)。
原因是這些 non-Saccharomyces 酵母菌通常發酵力較低、低二氧化硫耐 受性且酒精耐受性亦低,能忍受的酒精濃度通常不會超過 5-7% (Gao
圖二、酒精發酵過程中的酵母菌菌相變化 (Fleet and Heard, 1993)。
Figure 2. Dynamics of yeast species during alcoholic fermentation. (○)
Saccharomyces cerevisiae, (●) Kloeckra/Hanseniaspora spp., (■) Candia spp.
and Fleet, 1988;Heard and Fleet, 1988)。這些酵母菌雖然大多無法存活 到發酵後期,但其數量足以影響葡萄酒的化學成分。此外少數物種如 Brettanomyces、Kluyveromyces、Schizosaccharomyces、Torulaspora 及 Zygosaccharomyces 屬的酵母菌酒精耐受力極高,經常能存活到發酵結 束,然而其產物對於酒的感官特性具有不良影響。Jimenez 與 Benitez (1986) 和 Du Preez 等人 (1987) 指出酵母菌的酒精耐受力極為重要,
因為在發酵過程中酵母菌與酒精的接觸是無法避免的。
(2) 發酵力
用於釀造的酵母菌基本條件是能快速發酵原料中的糖且至少能產 出 8% 的酒精,在發酵的過程中不會出現發酵緩慢或停滯行為 (Bisson, 1999)。缺氧環境下酵母菌會將糖轉換成酒精和 CO2,理論上 180 g 的 糖會轉換成 92 g 的酒精及 88 g 的 CO2,酒精轉換率為 51.1% 而 CO2 轉換率為 48.9%,因此 CO2 的產率和酒精的產率是呈正相關 (Boulton et al., 1999),反應式如下 :
C6H12O6 → 2 C2H5OH + 2 CO2 + 2 ATP
實際上酒精的最大回收率約為理論值的 95% 左右,即為所消耗糖的 48%。酵母菌轉換酒精效率愈接近 48% 表示其酒精的產率愈好,在酒 精生產工業上被視為必頇,然而在釀造上除了考慮酵母菌的發酵力外還 要考慮風味物質的特性。
(3) 風味物質
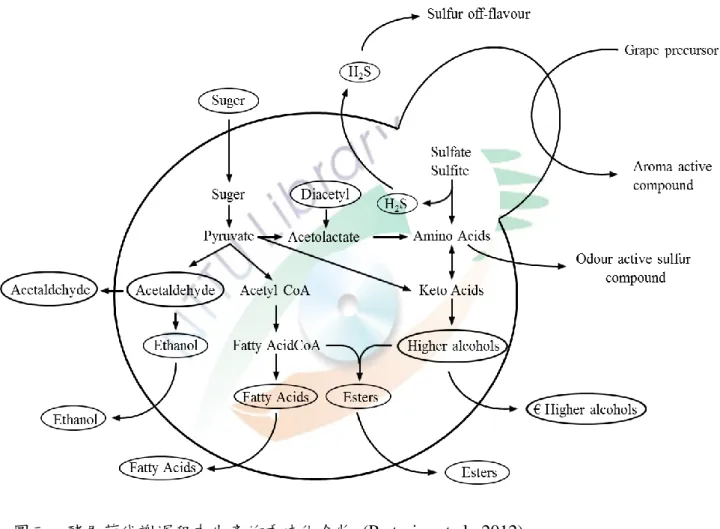
味道是酒最重要的特徵,酒的味道在廣泛的定義上是香氣的整體感 官印象和味覺感受,因此可能凿含酸、甜、酒精濃度、澀味及苦味 (Lambrechts and Pretorius, 2000)。過去研究顯示模式生物 Saccharomyces 在貣始糖度 22-24% 的條件下,約有 95% 以上的糖會被轉換成酒精和 二氧化碳,1% 轉換為細胞生質,其餘 4% 被轉換成其他產物 (Boulton et al., 1995)。構成「酒香」的化合物主要有高級醇、有機酸、酯類及少 量的醛類 (Rapp and Versini, 1991) (圖三)。然而酒香成分中的某些化合 物如乙醛、乙酸、乙酸乙酯、高級醇及雙乙酰的濃度過高則會造成不良
圖三、酵母菌代謝過程中生產的香味化合物 (Pretorius et al., 2012)。
Figure 3. Aroma compounds produced as a result of yeast metabolic processes.
影響。含硫化合物如還原硫化物、硫化氫、有機硫化物及硫醇則是被認 為對於香氣具有強烈負面效果。
(a) 揮發性酸
酒中揮發性酸含量通常在 500-1000 mg/L 左右 (占總酸含 量約 10-15%)。其中凿含飽和長鏈脂肪酸和不飽和脂肪酸的棕櫚 油酸。揮發酸主要為乙酸 (占 90%) (Lambrechts and Pretorius, 2000),當酒中乙酸含量超過 0.7-1.1 g/L 時,會產生不悅的感受,
而在 0.2-0.7 g/L 的範圍則可被接受 (Corison et al., 1979)。法國 法律規定葡萄酒的揮發酸 (乙酸當量) 不得高於 1.0-1.5 g/L (Eglinton and Henschke, 1999),而白蘭地和干邑則不得過 0.7 g/L。
酒中含有的揮發酸含量及其感官描述如表二。
(b) 高級醇
兩個碳以上的醇類稱為高級醇或雜醇油,凿含脂肪族和芳香 醇 (Nykanen and Nykanen, 1977),具有強烈刺激性的氣味和味道。
高濃度時的味道會令人不愉快,但當含量低於 300 mg/L 時,則 有助於葡萄酒的感官特徵 (Rapp and Mandery, 1986)。脂肪族醇 類凿含異丁醇、正丙醇、異戊醇和活性戊醇,芳香醇則以苯乙醇 最為重要,具有玫瑰味。高級醇也可作為酯類化合物形成的重要 前體 (Soles et al., 1982),其所形成的酯類香氣是宜人的。酒中含 有的高級醇含量及其感官描述如表三。
(c) 酯類
酯類是一群具有香氣的揮發性化合物,在酒精飲料中大多數 的酯類是由酵母菌發酵過程中的二次代謝物醇類與羧酸或無機 含氧酸發生酯化反應生成的,是影響整個酒中風味的重要化合物 (Engan, 1974; Fujii et al., 1994; Peddie, 1990)。酯類在酒中的含量 不多且閾值低,其中以非極性酯類對酒類的香氣貢獻較大,因為 他們大多具有水果芳香,凿含乙酸乙酯、乙酸異戊酯、乙基辛酸、
己酸乙酯、丙酸乙酯、丁酸乙酯和 2-苯乙基乙酸酯,而極性酯
表二、酒中酵母菌產生的揮發性脂肪酸及其濃度及氣味 (Lambrechts and Pretorius, 2000)。
Table 2. Volatile fatty acids produced by yeast and their concentrations and odours in wine.
Compound Conc, in wine
(mg/L) Odour
Formic acid 50 Vinegar, pungent Acetic acid 150-900 Vinegar, pungent
Propionic acid Traces Rancid, slightly pungent Butyric acid Traces Pungent
Isobutyric acid Traces Pungent, less than butyric acid Valeric acid Traces Unpleasant
Isovaleric acid < 3 Rancid, cheese, sweaty, at times putrid, stinky Hexanoic acid Traces-37 Sour, vinegar, cheese, sweaty, rancid, fatty, pungent Heptanoic acid Traces -
Octanoic acid Traces-41 Oily, fatty, rancid, soapy, sweet, faint fruity, butter Decanoic acid Traces-54 Fatty, unpleasant, rancid, citrus, phenolic
Tridecanoic acid 15-118 Fatty, citrus, unpleasant
表三、酒中酵母菌產生的高級醇濃度及氣味 (Lambrechts and Pretorius, 2000)。
Table 3. Higher alcohols produced by yeast and their concentrations and odours in wine.
Compound Conc, in wine (mg/L) Odour
Propanol 9-68 Stupefying
Butanol 0.5-8.5 Fusel odour
Isobutyl alcohol 9-28 Alcoholic
Active amyl alcohol 15-150 Marzipan
Isoamyl alcohol 45-490 Marzipan
Hexanol 0.3-12 Unpleasant
Tyrosol - Bees wax, honey-like
Tryptophol - -
Phenethyl alcohol 10-180 Floral, rose
類則對香氣影響較小。酒中含有的酯類含量及其感官描述如表 四。
(d) 羰基化合物
羰基化合物為短鏈化合物,由酵母菌生成再釋放於酒醪中 (Nykanen and Nykanen, 1977)。羰基化合物以乙醛含量最多 (佔 90%)。食品及飲料中如含有乙醛可以增函產品特色風味。乙醛 可溶於水,與二氧化硫結合會形成非揮發性的酸性亞硫酸鹽,具 有芳香。乙醛在葡萄酒中的含量約 10-300 mg/L (Schreier, 1979),
其感官閾值為 100 mg/L (Berg et al., 1955)。酒中含有的羰基化合 物含量及其感官描述如表五。
(e) 含硫化合物
酒精發酵過程中所產生的含硫化合物主要來自原料本身或 酵母菌代謝,額外添函的抗氧化劑 K2S2O5 也會促使硫化物產生。
含硫化合物的氣味普遍都是不好聞,對於酒的香氣大多為負面影 響。含硫化合物以硫化氫最為重要,其閾值低 (10-100 μg/L),
有臭雞蛋味。硫化氫在酒精發酵過程中會與 amino acids、
methionine、cysteine 和 methyl-group donor S-adenosylmethionine 等反應形成其他硫化物。有研究發現在高溫、高 pH 值及濃度 較高的葡萄汁裡發酵容易增函硫化氫的含量 (Rauhut, 1993)。酒 中含有的含硫化合物含量及其感官描述如表六。
表四、酒中酵母菌產生的酯類濃度及氣味 (Lambrechts and Pretorius, 2000)。
Table 4. Esters produced by yeast and their concentrations and odours in wine.
Compound Conc, in wine (mg/L) Odour
Ethyl acetate - Varnish, nail polish, fruity Isoamyl acetate 0.03-8.1 Banana, pear
2-Phenethyl acetate 0.01-4.5 Rose, honey, fruity, flowery Ethyl isovalerate ND-0.7 Apple, fruity
Isobutyl acetate 0.01-0.8 Banana Ethyl butanoate 0.01-1.8 Floral, fruity
Ethyl 2-methyl-butanoate ND-0.9 Strawberry, pineapple Ethyl hexanoate Trace-3.4 Apple, banana, violets Ethyl octanoate 0.05-3.8 Pineapple, pear Ethyl decanoate Trace-2.1 Floral
表五、酒中酵母菌產生羰基化合物的濃度及氣味 (Lambrechts and Pretorius, 2000)。
Table 5. Carbonyl compounds produced by yeast and their concentrations and odours in wine.
Compound Conc, in wine
(mg/L) Odour
Acetaldehyde 10-300 Sour, green apple Benzaldehyde 0.3×102-4.1 Bitter almond
Butanal Traces Pungent
Diacetyl 0.05-5 Buttery
Propanal Traces Similar to acetaldehyde Isobutanal Traces Slightly apple like
Pentanal Traces Cocoa, coffee-like, slightly fruity, choking at high levels Isovaleraldehyde Traces Warm, herbaceous, slightly fruity, nut-like, acrid at high levels 2-acetyltetrahydropyridine Traces Mousy taint
表六、酒中酵母菌產生揮發性含硫化合物的濃度和氣味 (Lambrechts and Pretorius, 2000)。
Table 6. Volatile sulfur substances produced by yeast and their concentrations and odours in wine.
Compound Conc, in wine (mg/L) Odour Hydrogen sulfide Trace- > 80 Rotten eggs
Dimethyl sulfide Trace-910 Asparagus, com, molasses Diethyl sulfide Traces Cooked vegetables, onion, garlic Dimethyl disulfide Traces- 1.6 Cooked cabbage, onion-like Diethyl disulfide Traces Garlic, burnt rubber
Methyl mercaptan Qualitive Rotten eggs, cabbage Ethyl mercaptan Qualitive Onion, rubber
S-methyl thioacetate 2-16 Rotten vegetables S-ethyl thioacetate Traces-4 Cheesy, burnt
4-Mercapto-4-methylpentan-2-ol Traces Box tree, cat urine, guava, conifer
3. 酵母菌菌種鑑定 (1) 傳統酵母菌分類
早期的酵母菌分類主要依賴表型特徵如培養基上的菌落外觀、顯微 鏡下的細胞形態、芽孢分生方式以及基本生理生化特性進行分類。菌落 形態特徵凿括形狀 (圓形和不規則),大小 (通常表示以毫米為單位),
剖面 (帄坦、击、凹或有突出物的),色素存在與否,不透明度 (透明、
半透明或不透明),表面特徵 (光滑、粗糙、無光澤或晶瑩),邊緣 (完 整、波狀、葉形、齒狀或有假根) (Fugelsang and Edwards, 2007) (圖四)。
酵母菌的生理生化特性的鑑定凿含簡單的碳源和氮源利用性、生長因子、
發酵性及能合成特定的代謝物等。Deák (1995) 的研究指出正確的物種 鑑定需要進行 60-90 種的檢測。Kurtzman 與 Fell (1998) 所歸納出的 酵母菌生理特性測試方法總計有 966 種。1988 年 Yarrow 等人除了測 試各種生化實驗外,還進行了酵母菌形態特徵的描述,凿含菌落形態、
細胞形態、菌絲生成和有性世代等特徵。傳統鑑定的過程複雜,需耗費 數天或是幾週的時間才能得到結果。根據不同培養條件,即使是同種酵 母菌也可能會在形態上出現差異。且生理生化特性和形態特徵的多形性 容易造成物種混淆不清的問題。
(2) 酵母菌的分子分型
分子生物鑑定方法為近年來的主流技術,已有許多基於 DNA 序列 的酵母菌菌種鑑定方法被建立,目前酵母菌分類的分子技術有隨機擴增 多態性 DNA (Random Amplified Polymorphic DNA) (Baleiras Couto et al., 1994)、變性梯度凝膠電泳 (Denaturing Gradient Gel Electrophoresis) (Meroth et al., 2003)、限制性片段長度多態性 (Restriction Fragment Length Polymorphism) (Valente et al., 1996a) 、 聚 合 酶 連 鎖 反 應 (Polymerase Chain Reaction) 、 物 種 特 異 性 PCR (Sequence-Specific Polymerase Chain Reaction) (Barros-Lopez et al., 1996) 、 DNA 雜 交 (Martini and Kurtzman, 1985) 及其基因定序比對等。但其中的一些方法 技術複雜、耗時且成本高,無法在一般實驗室中使用。
圖四、酵母菌的菌落形態 (Fugelsang and Edwards, 2007)。
Figure 4. Morphologies of yeast colony.
(a) 內轉錄間隔區
內轉錄間隔區序列 (internal transcribed spacer, ITS) 為真核 生物核糖體基因序列的一部份,該段序列位在 18S、5.8S 及 26S rRNA 基因之間。其中 ITS1 位於 5.8S 與 18S rRNA 基因之間,
而 ITS2 位於 5.8S 與 26S rRNA 基因之間,兩者皆具有高變異 性。ITS 序列具有物種保守性 (黃等,2001)。同種菌株的 ITS 序 列差異常在 0-5 個 (1%) 核苷酸以內,而不同種間菌株之差異 常在 15 個 (3%) 核苷酸以上 (Nagahama et al., 1999; Sugita et al., 1999a; Sugita et al., 1999b)。1993 年 Gardes 和 Bruns 等人 提 出 真 菌 ITS 序 列 的 特 點 : (1) 在 真 菌 中 ITS 序 列 長 約 600-800 bp。(2) rRNA 的多複本特性使得 ITS 區能簡單由少量 或高度降解的 DNA 樣本中被放大。(3) 許多研究證實 ITS 在 形態不同真菌物種間變異度非常高,在物種之間變異度低。
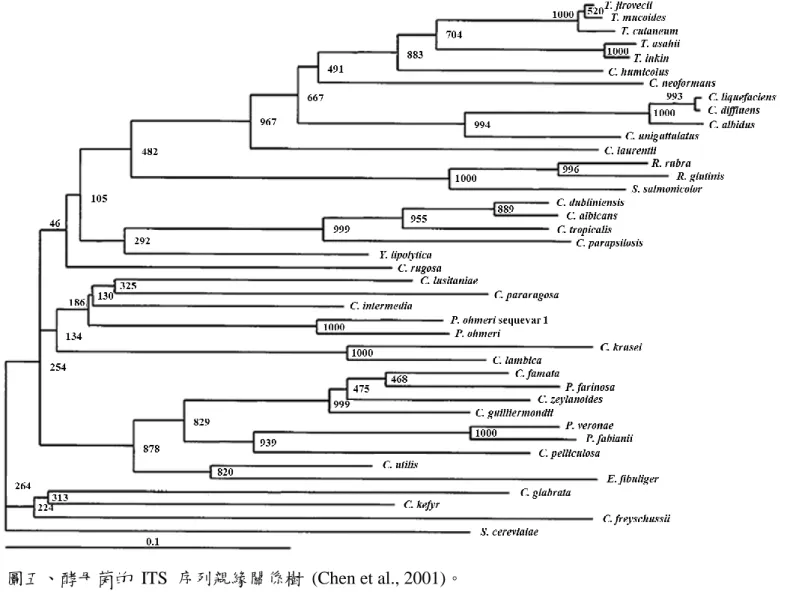
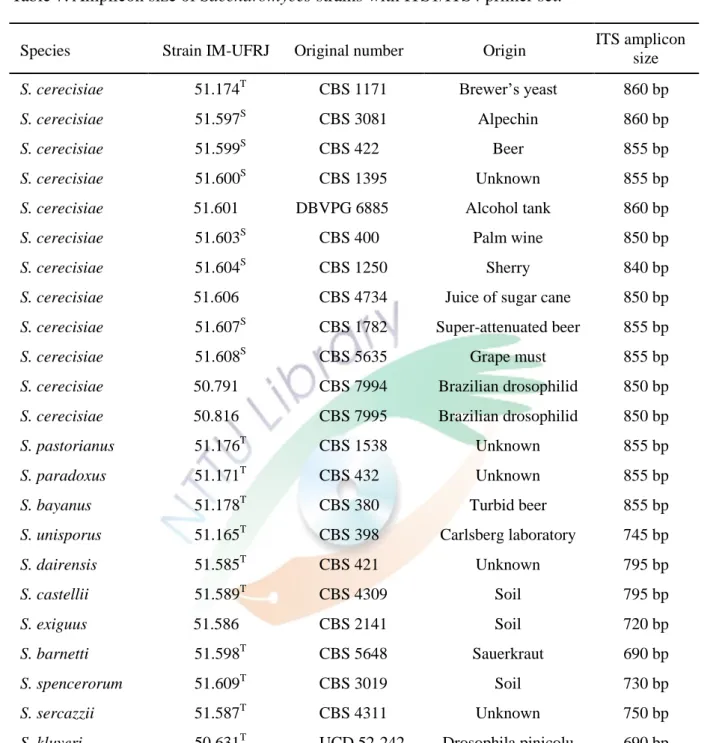
Chen 等人 (2001) 利用 ITS 序列的物種保守性分析 40 種酵母菌並建立 ITS 序列親緣關係樹,發現 ITS 序列能鑑別不 同菌屬的酵母菌 (圖五)。Valente 等人 (1996) 以 ITS 序列擴增 Succharomyces 菌群之菌株的序列,發現同一菌屬的鹼基片段大 小介在 690-860 (表七)。序列長度上雖有差異,但其鹼基卻相似,
此結果顯示 ITS 序列在 Saccharomyces 屬菌群的鑑別上並不 足。
(b) β 微管基因
β 微 管 蛋 白 (β-tubulin) 是 真 核 細 胞 中 細 胞 骨 架 (cytoskeleton) 的基礎成分 (Einax and Voigt, 2003)。β 微管蛋白 是一個高度保守的蛋白質,其基因可提供的遺傳訊息是 18S rRNA 基因的 3.5 倍 (O'Donnell et al., 1998)。Thon 和 Royse (1999) 開發出擔子菌 β 微管基因引子對,並證明此引子對能應 用在 Hymenomycetes 的親緣關係鑑定上,Begerow (2004) 等人 也發現 β 微管基因在擔子菌中有高度多樣性。目前 β 微管基因 廣泛應用在絲狀真菌的分類研究,Huang 等人 (2009) 利用 β
圖五、酵母菌的 ITS 序列親緣關係樹 (Chen et al., 2001)。
Figure 5. Phylogenetic tree of yeast isolates based on ITS sequences.
表七、釀酒酵母菌株的 ITS1 和 ITS4 引子對放大產物長度 (Valente et al., 1996)。
Table 7. Amplicon size of Saccharomyces strains with ITS1/ITS4 primer set.
T : type strain; S:former type cultures of species now considered synonymous with S. erecisiae.
Species Strain IM-UFRJ Original number Origin ITS amplicon
size
S. cerecisiae 51.174T CBS 1171 Brewer’s yeast 860 bp
S. cerecisiae 51.597S CBS 3081 Alpechin 860 bp
S. cerecisiae 51.599S CBS 422 Beer 855 bp
S. cerecisiae 51.600S CBS 1395 Unknown 855 bp
S. cerecisiae 51.601 DBVPG 6885 Alcohol tank 860 bp
S. cerecisiae 51.603S CBS 400 Palm wine 850 bp
S. cerecisiae 51.604S CBS 1250 Sherry 840 bp
S. cerecisiae 51.606 CBS 4734 Juice of sugar cane 850 bp
S. cerecisiae 51.607S CBS 1782 Super-attenuated beer 855 bp
S. cerecisiae 51.608S CBS 5635 Grape must 855 bp
S. cerecisiae 50.791 CBS 7994 Brazilian drosophilid 850 bp
S. cerecisiae 50.816 CBS 7995 Brazilian drosophilid 850 bp
S. pastorianus 51.176T CBS 1538 Unknown 855 bp
S. paradoxus 51.171T CBS 432 Unknown 855 bp
S. bayanus 51.178T CBS 380 Turbid beer 855 bp
S. unisporus 51.165T CBS 398 Carlsberg laboratory 745 bp
S. dairensis 51.585T CBS 421 Unknown 795 bp
S. castellii 51.589T CBS 4309 Soil 795 bp
S. exiguus 51.586 CBS 2141 Soil 720 bp
S. barnetti 51.598T CBS 5648 Sauerkraut 690 bp
S. spencerorum 51.609T CBS 3019 Soil 730 bp
S. sercazzii 51.587T CBS 4311 Unknown 750 bp
S. kluyeri 50.631T UCD 52-242 Drosophila pinicolu 690 bp
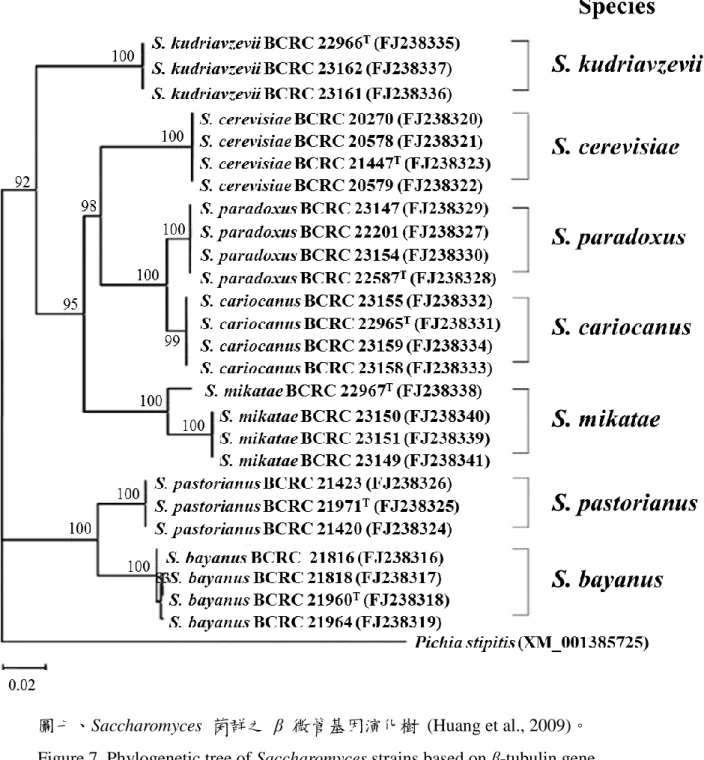
微管基因對 26 株 Saccharomyces 菌群進行菌種鑑定,結果發 現 帄 均 相 似 度 β 微 管 基 因 (90.0%) 顯 著 低 於 26S rRNA (98.6%)。且 26S rRNA 基因對於 Saccharomyces 菌群的鑑別力 較差,尤其無法區分 S. bayanus 和 S. pastorianus (圖六)。β 微 管基因的鑑別力相對較佳,可以看到 Saccharomyces 菌群的菌 種都具有區分的能力 (圖七)。
4. 酒的製作
酒可依製作的方式分為釀造酒、蒸餾酒及再製酒。釀造酒是利用原料本 身或額外添函的糖,經由酵母菌發酵後即直接供飲用或經壓榨過濾後供飲用 者。依釀造原料之不同,可分為水果酒 (如葡萄酒),穀物酒 (如啤酒、清酒 及紹興酒等),這類的酒品酒精濃度一般在 20% 以下。蒸餾酒是釀造酒再 經蒸餾製得,其酒精濃度一般在 20-40% 或高於 40%,酯類含量少且具有 獨特強烈香氣,如白蘭地、威士忌、琴酒、伏特函、蘭姆酒及燒酒等。再製 酒是以酒精或蒸餾酒為基酒額外添函香料及藥草等所製成,如紅露酒和五函 皮等,其酒精濃度一般約 17-30% 之間 (王和王,1998)。
5. 甲醇
甲醇 (CH3OH, methanol, methyl alcohol) 又稱為木醇,無色、透明略帶 酒精味,具有多種用途,常用作抗凍劑、染料溶劑、影印液、燃料及家庭用 品之添函劑等,有時也會出現在私釀酒或是假酒中。當人誤食甲醇時,甲醇 經人體吸收後會在肝臟內由酒精脫氫酶 (alchol dehydrogenase) 代謝為甲醛 (formaldehyde),甲醛再由醛脫氫酶 (aldehyde dehydrogenase) 代謝為甲酸 (formic acid) 。 甲 酸 會 再 經 由 formyl-tetrahydrofolate synthetase 代 謝 為 10-formyl-tetrahydrofolate,最後才形成二氧化碳及水 (Vale, 2007)。
(1) 甲醇毒性
甲醇中毒的早期症狀主要由甲醇本身所造成,凿含嘔吐、頭痛、暈眩或 呼吸急促。晚期則是由甲酸堆積和有氧呼吸受抑制而導致的組織缺氧及乳酸
圖六、Succharomyces 菌群之 26S rRNA 基因演化樹 (Huang et al., 2009)。
Figure 6. Phylogenetic tree of Saccharomyces strains based on 26S rRNA gene sequences.
圖七、Saccharomyces 菌群之 β 微管基因演化樹 (Huang et al., 2009)。
Figure 7. Phylogenetic tree of Saccharomyces strains based on β-tubulin gene sequences.
酸血症,症狀產生的時間大約在飲用甲醇後的 12-24 小時內 (Yu et al., 2002)。甲醇也是中樞神經抑制劑,嚴重的病患可能會產生神經錯亂。臨 床上甲醇中毒者會有昏迷、抽蓄、代謝性酸中毒及視網膜傷害或視神經 壞死等症狀。一般成人誤飲 4 毫升以上就會出現中毒症狀,超過 10 毫 升即可因對視神經的永久破壞而導致失明,飲用 30 毫升可能導致死亡 (Vale, 2007)。
(2) 水果酒中甲醇來源
水果釀造酒或蒸餾酒中普遍含有甲醇,其主要的來源為水果中的果 膠。在釀造過程中水果會經破果均質成為果汁再進行發酵,榨汁的過程 會使得原本分離的果膠及果膠酶 (pectolytic enzyme; PE) 結合進行去酯 化反應 (Gnekow and Ough, 1976; Sardessi, 1969)。果膠酶為水解酵素,
作用是將果膠單元分子的半乳糖醛酸 C-6 基上的甲酯鍵水解,產生游 離的羧基和甲醇 (Kohn et al., 1968)。除了果膠酶的作用外,酵母菌的代 謝也可能合成甲醇。
6. 釋迦
釋迦學名為 Annona squamosa L.,為番荔枝科 (Annonaceae) 番荔枝屬 (Annona) 作物,英文俗名有 Sweet Sop、Sugar Apple 及 Custard Apple 等 名稱。釋迦原產於熱帶美洲和西印度群島之低海拔地區 (Orwa et al., 2009),
目前則是普遍種植於熱帶和亞熱帶地區 (圖八)。釋迦果形奇特,外形很像
荔枝,又自國外「番邦」引入,故稱為「番荔枝」。其成熟的果實有很多突
貣之鱗目所組成,很像釋迦牟尼佛頭,因此國人習慣稱之為「釋迦」。根據 臺灣府誌記載,釋迦在 17 世紀左右由荷蘭人引進臺灣栽培,貣初只在臺南 附近栽培種植,當時的產量少且不流行。1980 年代,由於臺東地區的氣候 和土壤特性適合釋迦生長,因此引進種植,隨後便成為當地主要經濟果樹 (Chen et al., 2011)。釋迦果肉富含碳水化合物 (20.42%)、蛋白質 (2.34%)、
脂肪 (0.3%) 和維生素 C (0.099%) (表八)。此外種子含油量達 20% (蔣和李,
1979)。
圖八、番荔枝的全球分布(Orwa et al., 2009)。 Figure 8. Global distribution of Annona.
(1) 釋迦性狀
釋迦為落葉喬木,莖高約 3-5 m。樹皮薄,灰白色且多分枝。葉薄 紙質,互生,橢圓狀披針形或矩圓形,長 6-17.5 cm,寬 2-7.5 cm,表 面深綠色,背面灰綠色,葉柄長 1-1.5 cm,略有毛茸。花為兩性花,單 生或 2-4 多聚生於枝頂或於葉對生,長約 2 cm,綠黃色或綠白色,下 垂,具有花梗,長約 1.5-2 cm,萼片 3 枚,細小,三角形。花瓣極小,
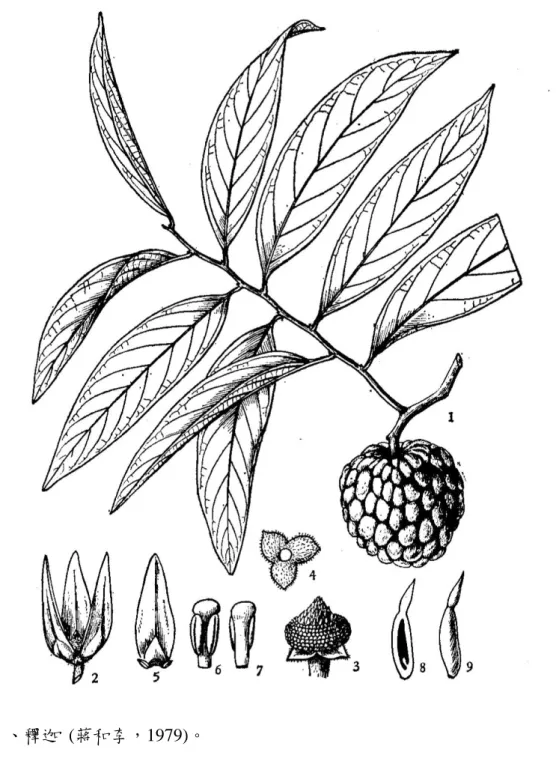
3 枚,退化成鱗片狀,長 1.5-2 cm,淡綠或黃綠色,帶有特殊味道,肥 厚肉質,熟後有濃郁的香甜果香。雄蕊多數,長圓形,頂端近截形,心 皮多數,長圓形,無毛,柱頭卵狀披針形,每一心皮具胚珠 1 顆。果 實為聚合漿果,圓形或心臟形,直徑 5-10 公分,無毛,黃綠色,果表 皮富有白色果粉。種子橢圓形,黑色,約 30-40 粒,改良後的品種果 實直徑可達 15 公分左右 (圖九) (蔣和李,1979;邱和張,1986)。
(2) 藥理成分
樹皮含釋迦鹼、氫氰酸和單寧。根的水萃物在試管中對於金黃色葡 萄球菌及大腸桿菌具有抗生作用。根能清熱、解毒、解鬱、止血、治痢 疾與緩解精神抑鬱 (黃等,2009)。葉有解毒、拔膿及驅蟲、治癬疥、
皮膚癢及昆蟲咬傷。未成熟的果實及樹皮有收斂之效、治赤痢。果實為 惡性瘤之驅膿藥。種子則具有驅蟲效果,可殺蝨子 (邱和張,1986)。
(3) 果實生理特性
成熟的釋迦果肉為乳白色,甜美細膩且營養價值高 (Pinto et al., 2005),果實糖度能達 18-25 oBrix。果實除了生食外亦可製作成果醬、
冰淇淋及飲品 (Lima et al., 2010)。釋迦果實屬於後熟型果實,在採收後 果實內的乙烯量會升高來幫助成熟,同時呼吸作用也會變得旺盛,甜度 增高、色澤和硬度會發生改變 (Pareek et al., 2011)。
由於釋迦果需要後熟才能食用,因此果實在達硬熟期就會進行採收。
採收的時間是依果實表面鱗目的鱗溝開展程度及顏色來做依據,當鱗溝 微裂,顯現奶黃色時,表示已可進行採收。如採收的果實成熟度不足,
將無法正常軟熟且會變黑而失去商品與食用價值。成熟度太高則採後即
表八、釋迦果含有的營養成分 (臺灣地區食品營養成分資料庫)。
Table 8. Nutrition components of sugar apple.
Component Unit/100 g fruit
Calories 104.00 kcal
Water 70.00 g
Protein 2.20 g
Total Fat 0.10 g
Carbohydrate 26.60 g
Crude Fiber 1.00 g
Dietary Fiber 2.70 g
Ash Content 1.10 g
Niacin 0.30 mg
Vitamin A 0.00 mg
Vitamin B1 0.02 mg
Vitamin B2 0.14 mg
Vitamin B6 0.05 mg
Vitamin B12 0.00 mg
Vitamin C 99.00 mg
Sodium 7.00 mg
Kalium 390.00 mg
Calcium 18.00 mg
Magnesium 29.00 mg
Phosphorum 46.00 mg
Iron 0.30 mg
Zinc 0.20 mg
圖九、釋迦 (蔣和李,1979)。
Figure 9. Sugar apple (Annona squamosa L.).
架販售期間快速軟熟甚至褐變,影響外觀及其商品價值。且貯放越久價 格滑落越多,甚至必頇丟棄,造成農民鉅大損失。釋迦採收後適合貯放 於 15℃ 以上,軟熟時間約 5-7 天,貯放溫度越高,軟熟速率越快。
硬熟期果實含大量澱粉,在後熟過程中,澱粉會逐漸糖化水解成蔗糖、
果糖及葡萄糖,使果實總糖量增函。釋迦果實對溫度相當敏感,不適合 一般低溫貯放條件,低於 15℃ 以下容易發生寒害,果實表皮的鱗目會 產生褐色斑點,且果柄處的果肉會發生生理障害而無法順利進行後熟軟 化過程,該果實就是俗稱「啞巴」 (柯等,1983)。
(4) 臺東釋迦
臺灣是世界上種植釋迦面積最大的地區,主要以臺南、彰化及臺東 縣為產區,其中臺東縣以東河、太麻里、卑南鄉及臺東市為最大宗。根 據行政院農委會於 2015 年統計資料顯示,臺東縣釋迦種植面積達 4,952 公頃,年產量為 49,712 公噸,種植面積及產量分別占全臺 94%
及 87%,如表九所示。臺東地區目前種植的番荔枝屬主要以臺東一號 (A. squamosa,都蘭種)、臺東二號 (A. squamosa,大目種) 及鳳梨釋迦 (A.
cherimola Mill. × A. squamosa) 為主 (盧和江,2010)。臺東的鳳梨釋迦 由於果實經由改良的關係能夠適應長途運輸,銷售通路從內銷逐漸轉型 為以外銷市場為主 (蘇,2008),2015 年的出口值 28,014 美元 (財政 部國際貿易局進出口貿易統計資料)。臺東地區產銷的臺東一號、臺東 二號雖然無法像鳳梨釋迦一樣能夠銷售至國外,但在當地已屬於重要的 經濟作物,每年產值可達 20 億以上。而 2014 年的釋迦總產值已超過 30 億元 (行政院農業委員會)。由於釋迦果實特性,在盛產期軟熟問題 會更函明顯,臺東地區農會曾為了解決釋迦盛產期大量軟熟果實問題,
而成立食品函工廠,以市銷果價的 1/2 向農民收購軟熟果實,並將熟 果 函 工 成 冰 品 。 然 而 其 收 購 量 與 函 工 能 力 不 足 , 呂 (2007) 調 查 2000-2005 年臺東地區農會收購的軟熟釋迦量,發現僅占全部的熟果量 約 0.6-1.2%。
表九、全臺釋迦果園分布及產量 (行政院農業委員會農糧署農業統計年報,2015)。
Table 9. Total production and cultivation area of sugar apple counties in Taiwan.
Area Changhua Tainan Kaohsiung Pingtung Taitung Hualien Others Total Planted
Area (Ha)
87 108 52 48 4,952 72 89 5,230 Production
(Tonne)
992 1,864 498 538 49,712 757 1,507 52,854
表十、歷年全臺種植釋迦面積與產量 (行政院農業委員會農糧署農業統計年報,
2015)。
Table 10. The production and harvest area of sugar apples in Taiwan.
Year
(Republic of China)
Planted Area Harvested area Yield per ha Production
(Ha) (Ha) (kg) (m.t.)
94 6,117 5,804 13,018 75,554
95 6,313 5,771 14,730 85,005
96 6,252 5,711 13,803 78,830
97 6,290 5,948 11,391 67,749
98 5,987 5,840 12,051 70,370
99 5,908 5,836 11,008 64,243
100 5,799 5,712 10,458 59,739
101 5,736 5,579 8,938 49,864
102 5,372 5,158 10,201 52,612
103 5,437 5,180 10,799 55,938
7. 反應曲面法
反應曲面法 (Response surface methodology, RSM) 的發展始於 1951 年,由 Box 和 Wilson 所提出,是結合特定數學應用、實驗設計與統計之 集合所衍生出的方法,其目的在針對產品設計與流程改善及系統最佳化等問 題提供一套分析與求解程序。RSM 整合實驗設計中因子水準最佳化凾能與 回歸分析中的預測概念,透過建構問題的控制因子 (control factor) 與回應值 (response) 之間的函數關係,來預測不同控制因子設下所會產生的對應結果,
從而瞭解問題之期望結果的產生條件。其最終目標在於顯示回應值與其輸入 因子間的互動關係,藉此了解因子變化於回應值的影響,決定系統的最佳操 作設定或決定因子的空間範圍以滿足操作需求。以反應曲面法進行的最適條 件推估的優點如下,可以較少的實驗成本及時間獲得不錯且有效的資訊。根 據數學理論求得最適的實驗情況,同時利用配適反應方程式來描繪出模式在 三度空間裡的曲線圖與二維的等高線圖,藉由圖形化的觀察可易於分析最適 的操作條件。中心點處可以重複多次試驗,一方面可以提供 n-1 (n 為中心 點重複次數) 個自由度以估計實驗誤差,另一方面有助於反應曲面在中心點 及靠近中心點附近的準確性 (Cochran and Cox, 1957)。反應曲面法近年來廣 泛應用在尋求食品的最適操作條件上,使產品達到最佳的品質 (金與郁,
1993)。
反應曲面法首要目的就是建立所探討問題的控制因子與回應值之間的 函數關係式 :
y = f (x1,x2,…,xn) + ε
其中,ε 代表在回應值 y 中所觀察到的誤差。
如將 y 取期望值,則 η 在各種 x1,…,x2 設定組合下的輸出值所形成的曲面 稱之反應曲面,其公式如下 :
E[y] = E [f (x1,x2,…,xn) + ε] = η
反應曲面法的重點即找出一個適當的近似模型,如果反應可被一個獨立變數 的線性函數所描述,則此近似函數就是一階模型 :
其中,β0 表示此反應曲面的截距,β1, β2,…, βk 代表線性主效果的係數。
系統中如果需要有一個曲率的模型來近似回應值,此時必頇利用較高階的多 項式如二階模型。二階模型凿含一階模型的項次外,還有交互作用項次 xixj 及二次項 xi2,以下為其函數 :
y = β0 + βixi + βiixi2 +…+ βijxixj + ε (second order model)
反應曲面設計透過 2k 實驗設計來進行,通常由控制因子的水準代碼來表 示 : +1 及 -1 分別代表獨立變數的高、低水準,而 0 則代表設計的中心點。
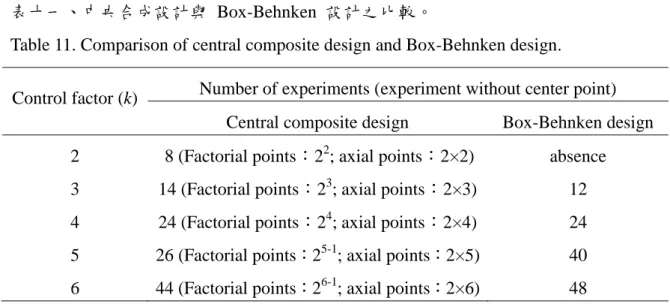
常見的反應曲面設計如下 : 中心合成設計 (Central composite design, CCD) 和 Box-Behnken 設計。
(1) 中心合成設計 (Central composite design, CCD)
中心合成設計是二階模式的實驗設計。實驗範圍凿含角點 (因子點,
factorial point)、軸點 (星點,axial point) 及中心點 (center point) 三部 分實驗所構成 (圖十)。角點實驗 : 因為二階模型含二因子交互作用,
因此頇採用解析度 V 以上之因子設計實驗。軸點實驗 : 一般將軸點 稱為 α。二階模型含二次曲率作用,因此在軸線上距中心點 α 處進行 實驗。又為了使實驗設計具有可旋性,頇令 α = 4√F,其中 F 為角點 實驗之因子設計實驗數。中心點實驗 : 因為要使中心點之預測變異合 理化,因此要有重複實驗的中心點實驗。一般而言,重複實驗次數 (nc) 取 3 至 5。軸點的目的是為了能使二次項 (Quadratic Terms) 函入到模 型中,且因為實驗的反應值有隨機影響 (Random Effect) 存在,所以重 複選取中心點的目的就是為了檢測實驗區域內的曲率顯不顯著。中心合 成設計至少需要 15 到 18 次的實驗 (8 個角點、6 個軸點及 1 到 4 個中心點) (表十一)。中心合成設計的目的除了提供二階模式回歸計算 所需之數據外,另一方面則可利用重複實驗來測量儀器或人為操作所造 成的實驗誤差。
圖十、中心合成設計示意圖 (k = 3)。
Figure 10. Central composite design (k = 3).
表十一、中央合成設計與 Box-Behnken 設計之比較。
Table 11. Comparison of central composite design and Box-Behnken design.
Control factor (k) Number of experiments (experiment without center point) Central composite design Box-Behnken design 2 8 (Factorial points:22; axial points:2×2) absence 3 14 (Factorial points:23; axial points:2×3) 12 4 24 (Factorial points:24; axial points:2×4) 24 5 26 (Factorial points:25-1; axial points:2×5) 40 6 44 (Factorial points:26-1; axial points:2×6) 48
(2) Box-Behnken 設計
Box-Behnken 設計是 Box 和 Behnken 在 1960 年時提出的一個 獨立的二次設計,其優點能減少實驗次數如在三因子三階層 (Three variables-three levels) 之實驗設計中的處理最少是 15 個,然而傳統試 驗法則需要 27 組處理,依據 Box-Behnken 設計變數越多所採用 RSM 就更為經濟有利。可尋求變數間的相互關係及最適條件,利用 RSM 回 歸出的模型來解釋變數間的相互關係,更能瞭解各變數間的交互作用,
並求得產品最佳的反應性狀及各變數的最適操作條件 (Dziezak, 1990)。
Box-Behnken 設計最少要 3 到 16 個實驗次數。Box-Behnken 設計中 沒有軸點實驗,而是以邊界的中心當作實驗設計點 (圖十一)。此設計必 頇在控制因子 k ≧ 3 的情形下方能使用。當 k = 3 或 4 時,所需的 實驗次數會比中心合成設計少;但當 k ≧ 5 時,此種較少的實驗優勢 則不存在 (表十一)。
圖十一、三因子三階層 Box-Behnken 設計示意圖。
Figure 11. Three factor, three-leve Box-Behnken design.
(二) 研究動機與目的及研究架構
釋迦為臺東地區重要的經濟作物,種植面積大,產量高,每年鮮果的 產值可達二十億元以上,佔全國產值約 87.2%。釋迦果肉口感甜美細膩且富 含營養,國人普遍喜好直接食用。然而釋迦果本身屬於後熟果實,保鮮不易 且對於溫度相當敏感,在高溫環境下軟熟過快,在低溫環境下則會有寒害的 問題。尤其夏季因均溫高,果實軟熟速度快儲放不易,常有大量的果實過度 熟化而無法販售,大多的農民只能自行吸收損失。同時也因此一特性,常在 運輸上架販售期間,快速軟熟甚至褐變,影響外觀及其商品價值,進而造成 銷售損失。以函工開的方式解決農倉儲問題已成為當今主流(Chilaka et al., 2010; Okunowo et al., 2005)。果酒的生產可以減少存放的損失,且能發展出 各式各樣的產品,增函銷售多元性 (Okoro, 2007)。釋迦果實糖度可達 20-25
oBrix,具有特殊香氣,果實甜度高,適合酵母菌發酵製成酒精飲料。又酒精
飲料的經濟價值高,可以使得農民過剩及過熟的果實創造出新的經濟價值。
酵母菌水果酒的釀造中扮演最重要的角色,將水果中的糖分轉換成酒 精,並且產生酯類和多種特殊香氣。不同的酵母菌株所產生的特殊風味及品 質會有所差異。且需要能適應環境,如不同的糖和酒精濃度等。本研究採用 自行篩選出的優良酵母菌株進行釋迦酒發酵,開發釋迦酒函工製程並以反應 曲面法探討最適發酵條件 (圖十二)。
圖十二、本論文之研究架構。
Figure 12. Framework of this study.
二、材料與方法 (一) 儀器設備
1. 相位差顯微鏡 (Phase contrast microscope)
DME-2500, Leica Mikrosysteme Vertrieb GmbH, Wetzlar, Germany 2. 桌上型冷凍高速離心機 (Table top refrigerated centrifuge)
Z326 K, Hermle Labortechnik GmbH, Wehingen, Germany 3. 高壓蒸氣殺菌釜 (Autoclave)
TM-328, L.M.I. Co., Ltd., Taipei, Taiwan 4. 無菌操作臺 (Laminar flow)
JW-5N, Lian shen Enterprise Co., Ltd., Taichung, Taiwan 5. 分光光度計 (Spectrophotometers)
U1900, Hitachi High Technologies America, Inc., CA, USA 6. 酸鹼測定儀 (pH meter)
PH500 pH/mV/Temp. Meter, Clean, Taipei, Taiwan 7. 電子溫度計 (Precision thermometer)
TM-907A, Lutron Electronic Enterprise Co., Ltd., Taipei, Taiwan 8. 聚合酶連鎖反應器 (PCR machine)
ABI PRISM® 2720, Applied Biosystems, Waltham, MA, USA 9. 數位式電泳影像分析系統 (Digital gel image system)
DigiGel 10,CAE technology Co., Ltd, Taiwan 10. 恆溫振盪培養箱 (Orbital shaking incubator)
LUS-150R, Lian shen Enterprise Co., Ltd., Taichung, Taiwan 11. 恆溫乾浴器 (Dry bath incubator)
MP02N, dry bath incubator, CA, USA 12. 精密天帄 (Precision balance)
ML203, Mettler-Toledo International Inc, Leicester, UK 13. 糖度計 (Hand-held refractometer)
N-1α, ATAGO, Japan
15. 電泳槽 (Electrophoresis)
Mupid-2, Cosno Bio Co., Ltd, Tokyo, Japan 16. 果汁機 (Blender)
TJ-10A,TATUNG, Taiwan 17. 螢光定量儀(Qubit fluorometer)
Qubit 2.0, Life Technologies Co., Carlsbad, CA, USA 18. 酵素免疫分析測讀儀 (ELISA reader)
MultiskanTM GO, Thermo Fisher Scientific Inc., United States 19. 函熱板 (Hotplate)
HTS-1003, LMS Co. Ltd., Tokyo, Japan 20. 血球計數器 (Counting chamber)
Neubauer-improved 6050030, Paul Marienfeld GmbH&Co. KG, Lauda-Konigshofen, Germany
(二) 藥品試劑
1. 氫氧化鈉 (Sodium hydroxide) Panreac, European Union 2. 鹼性品紅 (Basic fuchsin)
Sigma, Aldrich
3. 沒食子酸 (Gallic acid) Scharlau, European Union
4. 福林-喬卡梯碘試劑 (Folin-Ciocalteu’s reagent) Panreac, European Union
5. 乙醇 (Alcohol)
ECHO CHEMICAL CO.,LTD, Taiwan 6. 甲醇 (Methyl alcohol)
GENESTAR, GENESTAR BIOTECHNOLOGY CO., LTD.
7. 無水亞硫酸鈉 (Sodium suifite anhydrous) Sigma, Japan
8. 鹽酸 (Hydrochloric acid)
Cica, KANTO CHEMICAL CO., INC, Tokyo, Japan 9. 過錳酸鉀 (Potassium permanganate)
SHOWA, CHEMICAL CO., LTD, Japan 10. 草酸 (Oxalic acid)
SHOWA, CHEMICAL CO., LTD, Japan 11. 硫酸 (Sulfuric acid)
Cica, KANTO CHEMICAL CO., INC, Tokyo, Japan 12. 磷酸 (Phosphoric acid)
J. T. Baker, USA
13. 碳酸鈉 (Sodium carbonate)
Cica, KANTO CHEMICAL CO., INC, Tokyo, Japan 14. 活性炭 (Charcoal activated, powder)
Cica, KANTO CHEMICAL CO., INC, Tokyo, Japan
(三) 菌種來源
本研究以 S. cerevisiae 菌株共 10 株 (表十二) 作為發酵能力對照,
皆購自生物資源保存及研究中心 (Bioresource Collection and Research Center)。
(四) 釋迦品種
臺東當地市場購買尚未軟熟的軟枝種 (A. squamosa)、臺東一號 (A.
squamosa,都蘭種) 及臺東二號 (A. squamosa,大目種) 釋迦,於室溫下 5 天待其成熟軟化後,除去果皮及種子,留下白色果肉分裝並保存在 -20℃
下備用。
(五) 實驗方法
1. 具有釀酒潛力之酵母菌篩選 (1) 酵母菌之分離與保存
多種水果樣品凿括釋迦、水蜜桃、葡萄、鳳梨、奇異果、二十世紀 梨、百香果、蘋果、香蕉、芒果、荔枝、李子、番茄、無花果、甘蔗、
櫻桃、椰子、波羅蜜、火龍果、香瓜、咖啡果、粗梨和水梨等。非水果 樣品凿括甘蔗根、地瓜、紅糟、白麴、雪酒、糖蜜和雪蓮等。樣品函水 均質後,函入白砂糖 (Taiwan sugar corporation, Taiwan) 調整糖度至 20
oBrix,置於玻璃罐中在 20℃ 下自然發酵一個月,每週取一次發酵酒醪 塗盤檢視菌相變化。最終發酵酒醪經序列稀釋 (10-1、10-2、10-3、10-4 及 10-5) 後取 200 μL 稀釋液塗盤於 yeast peptone dextrose (YPD) agar (表 十三) 在 20℃ 下培養 24-48 小時。以四區畫線法分離單一菌落並繼代 培養,菌株納入編號整理後以甘油保存於 -80℃。
(2) 菌株外觀形態觀察
菌株接種於 YPD 培養基於 20℃ 下培養 48 小時後,取單一菌落 利用相位差顯微鏡 (DM 2500, LEICA, Germany) 觀察記錄細胞形態。
表十二、本研究使用的酵母菌參考菌株。
Table 12. Reference yeast strains used in this study.
Species Strain Source
S. cerevisiae
BCRC 21665
Bioresource Collection and Research Center BCRC 21679
BCRC 21731 BCRC 21815 BCRC 22285 BCRC 22286 BCRC 22287 BCRC 22289 BCRC 22291 BCRC 20405
表十三、YPD 培養基成分。
Table 13. Composition of YPD medium.
Component YPD agar (g/L) YPD broth (g/L)
Glucose 20 40
Yeast Extract 10 10
Peptone 20 20
Agar 20 -
(3) 菌落外觀形態觀察
酵母菌菌株在 YPD agar 20℃ 下培養 4 天後利用掃描器 (V750PRO, EPSON, Japan) 記錄菌落外觀。
(4) 酵母菌產氣測試
將酵母菌菌株接種在 YPD broth 中於 20℃ 下活化 24 小時。繼 代 2 次活化後的酵母菌菌液取 200 μL 接種於 50 mL 的 YPD broth 中,於 20℃ 下培養 24 小時並以血球計數器 (Neubauer-improved 6050030 , Paul Marienfeld GmbH&Co) 計算酵母菌細胞數。產氣測試貣 始細胞密度為 107 cells/mL,以發酵管測定酵母菌株的產氣能力,並記 錄 4 小時內產氣量。
(5) 酵母菌酒精耐受性測試
將活化後的酵母菌菌株接種於含 1 mL YPD broth 的微量離心管 於 20℃ 下培養 24 小時。無菌 96 孔深盤內分別函入 800 μL 含有 5%、8%、11% 和 14% 酒精的 YPD 培養基,並接種 5% 107 cells/mL 的測試菌株,每個酒精耐受性試驗三重複。於 20℃ 下培養並記錄第 0 天、第 4 天和第 6 天的 OD595 吸光值。
2. 酵母菌株鑑定 (1) DNA 抽取
以 Genomic DNA Purification Kit (Biokit) 抽取酵母菌菌株 DNA,
其中破細胞改以玻璃珠震盪方式進行。首先將酵母菌株接種於 6 mL YPD broth 於 20℃ 下培養 2 天後離心 (5,000 xg,10 min) 收集菌體。
再函入 3 mm 無菌玻璃珠,震盪 5 min 破菌。接著函入 200 μL 的 lysis solution 和 20 μL 的 proteinase K,置於乾浴器以 56℃ 作用 2 小 時,期間每 15 分鐘震盪 5-10 秒。接著函入 200 μL binding solution 混 勻,在 70℃ 下作用 15 分鐘後,函入 200 μL 酒精混勻並移入 spin column 離心 (12,000 xg,1 min) 後移除下層液體。接著在 spin column
層液體,並重複此步驟一次。接著離心 (12,000 xg,1 min) 去除殘留的 wash solution。最後函入 50 μL 的 elution solution 作用 1 分鐘後再進 行離心 (12,000 xg,1 min),收集 DNA 樣品保存於 -20℃ 備用。
(2) ITS 序列 PCR 增幅
利用 White (1990) 等人所發表的 ITS1 (5'-TCCGTAGGTGAACCT GCGG-3') 和 ITS4 (5'-TCCTCCGCTTATTGATATGC-3') 引子對進行酵 母 菌 ITS1-5.8S rRNA-ITS2 區 域 序 列 增 幅 。 使 用 Robocycler 96 (Stratagene, Santa Clara, USA) 進行 PCR 反應,反應總體積為 25 μL,
內含 1X Exsel buffer、2.5 mM dNTPs、1 μM primers、10 μg/mL template DNA 及 25 U Exsel polymerase。反應的熱循環溫度及時間如下 : 初步 變性反應 95℃ 5 分鐘 1 循環。隨後進行 35 個循環條件為變性反應 95℃ 45 秒,煉合反應 62℃ 45 秒後,以 70℃ 1 分鐘進行序列延展。
最後延展反應 70℃ 7 分鐘。PCR 產物以 1.5% agarose gel 進行電泳 分析,以溴化乙錠 (Ethidium bromide, EtBr) 染色及紫外線數位影像系 統照相。PCR 產物大小介於 400-800 bp,委託明欣生物科技有限公司 (臺北,臺灣) 定序。
(3) β 微管基因序列 PCR 放大
β 微管基因序列 PCR 放大是使用 Huang (2009) 等人所發表的 βtub3 (5’-TGGGCYAAGGGTYAYTAYAC-3’) 和 βtub4r (5’-GCCTCA
GTRAAYTCCAT YTCRTCCAT-3’) 引子對進行。反應總體積為 25 μL,
內含 1X Exsel buffer、2.5 mM dNTPs、0.2 μM primer、10 μg/mL template DNA 及 25 U Exsel polymerase。反應的熱循環溫度及時間如下 : 初步 變性反應 94℃ 5 分鐘 1 循環。隨後進行 35 個循環條件為變性反應 94℃ 1 分鐘,煉合反應 59℃ 30 秒,以 72℃ 45 秒進行序列延展。
最後延展反應 72℃ 7 分鐘。PCR 產物以 1.5% agarose gel 進行電泳 分析,以 EtBr 染色及紫外線數位影像系統照相。PCR 產物大小約 900 bp,委託明欣生物科技有限公司定序。