國立臺灣大學醫學院暨工學院醫學工程學研究所 碩士論文
Graduate Institute of Biomedical Engineering College of Medicine and Engineering
National Taiwan University Master Thesis
中草藥應用於關節軟骨組織工程之研究 The Effects of Chinese Herbal Medicine on Chondrocytes for Cartilage Tissue Engineering
林文央 Lin, Wen-Yang
指導教授:林峯輝 博士 Advisor: Lin Feng-Huei, Ph.D.
中華民國 96 年 7 月 July, 2007
ABSTRACT
The skeleton is composed of cartilage and bone which provide the functions such as support, protection, and movement in daily life. Articular cartilage tissue is composed of chondrocytes and extracellular matrix, where the chondrocytes only make up less than 10% of the total volume of cartilage. In healthy cartilage, the matrix is composed of collagens, especially type II collagen, proteoglycans, and noncollagenous proteins, and is filled with water because of the hydrophilic property of the framework.
Cartilage possesses limited ability to achieve spontaneous repair due to its dense extracellular matrix and lacking of blood vessels, lymphatics and innervation.
When lesions occur in articular cartilage, there is no bleeding and thus no mechanism for the replacement of lost or damaged tissue. Neighboring chondrocytes may respond by local proliferation; however, because they are sequestered in the dense matrix, they do not migrate into the damaged region to fill the void. If injury extends through the chondral layer to the subchondral bone and underlying vasculature, a repair response can hence occur, but the newly formed cartilage will be gradually populated with type I collagen and degenerate to a fibrocartilaginous scar tissue after 6–8 months.
Circumstances that impair chondrocytes function hence disrupt the balance of synthesis and catabolism and lead to the development of osteoarthritis. In the progression, osteoarthritis eventually impairs the function of a whole joint, including the cartilage, the subchondral bone, the synovium and the periarticular connective and muscular tissues. The current treatment options are fairly limited which include only symptomatic treatment of limited efficacy with analgesics, non-steroidal anti-inflammatory agents or intra-articular administration of steroids or hyaluronic acid, and if the joint destruction can not be halted, the ultimate measure is joint
replacement.
Tissue engineering has been a feasible way to regenerate cartilage in vitro, which combines cells, scaffolds, and signals to mimic the original environment of tissues in vitro. In addition, there are some Chinese herbal medicines have been used to treat the degeneration of the cartilage for thousands of years, but the precise mechanism of their potent chondrogenesis effects have not been fully elucidated.
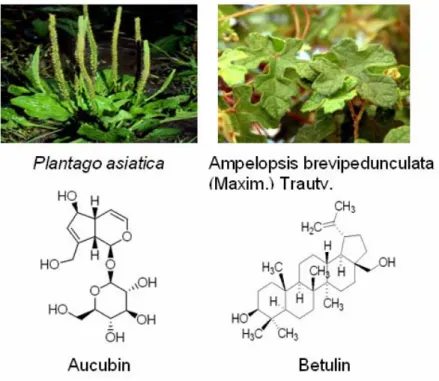
Therefore, in this study, we investigated the involved mechanism of two single chemical compounds: aucubin and betulin which were separately extracted from Plantago asiatic and Ampelopsis brevipedunculata (Maxim.) Trautv. by focusing on
their proliferation and antioxidant activity.
The experiment was divided into two parts. One was two-dimensional chondrocytes culture; the other was three-dimensional scaffold culture. After treating two-dimensional chondrocytes or three-dimensional seeded chondrocytes with optimal concentration of single chemical compounds, the proliferation and matrix productivity of them were evaluated by real-time reverse-transcriptase polymerase chain reaction, ELISA assays, and immuno-histochemical staining. The important role in scavenging free radicals by those extracted chemical compounds was detected by the chemiluminescence method.
The results showed that, in two-dimensional chondrocytes culture, both aucubin and betulin could effectively promote the mRNA expression of ECM and inhibit the mRNA expression related with ECM degradation at appropriate concentration, and the ability of O2•⎯ scavenging made aucubin and betulin as protectants of chondrocytes, which would stimulate chondrocyte proliferation and maintain the basic chondrocyte activities. In three-dimensional scaffold culture environment, betulin can significantly stimulate chondrocyte proliferation and maintain the basic chondrocyte activities until four-week cultivation, which suggested that in the future application
the in vitro three-dimensional cultivation of chondrocyte-scaffold hybrids should be maintained more than four weeks, and of course the addition of 0.32μg/ml of betulin into the cultured environment possessed positive effects toward chondrocytes and the whole cartilage-mimic tissue.
Keywords: Chinese herbal medicine, Cartilage, Tissue Engineering, Real-time RT-PCR, ROS, Aucubin, Betulin
中文摘要
人類的骨骼是由硬骨和軟骨構成的器官,具有支持、保護和運動的功能。軟 骨是由僅佔總體積百分之五的軟骨細胞和其所分泌的大量細胞外基質組成,在健 康的軟骨內,其細胞外基質主要為第二型膠原蛋白和多醣類,由於它們攜帶大量 的負電性,使水分子及正電離子易受吸引而聚集到軟骨中,因而使軟骨在受到壓 力時,能具有吸收震動力而達緩衝的效果。
然而受到破壞的軟骨其自我修復的能力很有限,除了因為軟骨中沒有血管或 神經的分布,使得受傷的部位沒有血流侵入而攜帶發炎因子,因而無法啟動修復 機制;另外在軟骨中的軟骨細胞數量本來就很有限,即使受傷部位鄰近的細胞可 以進行細胞分裂,但卻因數量過少且加上黏滯的細胞基質阻礙了細胞的遷移;而 當受傷侵入軟骨下方的硬骨時,受到破壞的硬骨內的血流便有機會侵入軟骨而啟 動修復,但此時新生成的軟骨組織,只具有分泌第一型膠原蛋白細胞外基質的能 力,而這種第一型膠原蛋白的機械特性和強度與原本第二型膠原蛋白相差甚大,
最終也將被逐漸降解而喪失軟骨的功能。
於是受到創傷而無法自我修復的軟骨會漸漸影響到整個關節,轉變為退化性 骨關節炎。目前臨床上的藥物療法皆著重在止痛而無法有根治之效,最常用的藥 物有普拿疼、第二型環氧脢抑制劑、非類固醇類抗發炎藥物、關節內注射透明質 酸、關節內注射類固醇,然而這些只治標不治本的療法,最終只能藉由置換人工 替代膝關節來取代壞損的關節。
利用組織工程在體外培養軟骨組織近年來已成為修補受損關節軟骨的途徑 之一,組織工程利用組織工程的三要素:細胞、生物材料骨架及訊號的刺激,模 擬組織在體內的狀態,以期在體外復育仿生組織。此研究從具有促進軟骨再生之
效的傳統中草藥複方中,萃取出主要來自於車前子及山葡萄兩種藥材中的兩種主 要有效單一化學成分,並以適當濃度添加到二維及三維支架的軟骨組織培養系統
中,而後利用同步定量聚合酵素鏈鎖反應、細胞總 DNA、細胞外基質葡萄胺聚
醣濃度、及免疫螢光染色確定細胞增殖情形;並利用化學發光技術配合適當的探 針,偵測軟骨細胞中自由基的產生量及其抗氧化的能力。
實驗結果發現,二維單層軟骨細胞的培養中,利用適當濃度的車前子或山葡 萄有效單一化學成分之添加,軟骨細胞能夠增加其細胞外基質相關的基因表現,
並能抑制與細胞外基質降解相關之蛋白質的基因表現;也由二維系統初步發現,
由山葡萄萃取之有效單一化學成分相較於由車前子萃取之有效單一化學成分,更 具活性氧之清除能力,因而使得軟骨細胞的整體活性在山葡萄萃取物之添加組更 為提升。在三維軟骨組織工程的培養中,經過兩週以上培養的軟骨細胞明顯地增 加細胞活性及複製能力,其細胞外基質相關的基因表現也明顯的上昇,而與細胞 外基質降解相關的基因被受到抑制;值得注意的是,經過四週以上培養的軟骨細 胞,其細胞整體活性相較於一週及兩週更為佳,或許建議在未來之臨床應用也宜 取經過四週培養之軟骨組織工程物為較佳。
關鍵詞:中草藥、軟骨、組織工程、定量聚合酵素鏈鎖反應、自由基、抗氧化
TABLE OF CONTENTS
ABSTRACT...i
中文摘要 ...iv
TABLE OF CONTENTS...vi
INDEX OF FIGURES ...ix
INDEX OF TABLES... xiii
LIST OF ABBREVIATIONS ...xiv
CHAPTER 1 INTRODUCTION ...1
1-1 Cartilage Biology...1
1-1.1 Definition and Composition of Cartilage ...1
1-1.2 Types of Cartilage...4
1-1.3 Homeostasis of Cartilage...5
1-2 Natural Limitations of Cartilage...8
1-2.1 Low Oxygen Tension on Cartilage Metabolism...8
1-2.2 Limited Cartilage Repair Response...9
1-3 Cartilage Pathology ...10
1-4 Current Strategies for Cartilage Treatment...13
1-4.1 Inadequate Pharmacological Options ...13
1-4.2 Novel Gene-based Approach for Cartilage Repair...15
1-5 Cartilage Tissue Engineering...17
1-5.1 Cell...18
1-5.2 Biomaterial Scaffold...19
1-5.3 Chondrogenesis Signals...20
CHAPTER 2 THEORETICAL BASIS ...21
2-1 Reactive Oxygen Species in Cartilage...21
2-1.1 Introduction of Reactive Oxygen Species ...21
2-1.2 Reactive Oxygen Species Formation in Cartilage...24
2-1.3 Antioxidant Systems in Cartilage ...25
2-1.4 Reactive Oxygen Species on Cartilage Matrix Degradation ...26
2-1.5 Reactive Oxygen Species on Cartilage Senescence ...27
2-1.6 Reactive Oxygen Species on Chondrocyte Death ...28
2-2 Chondrogenesis-related Chinese Herbal Medicine ...30
2-3 Purpose of this study...32
CHAPTER 3 MATERIALS AND METHODS ...34
3-1 Two-dimensional Cell-based Experiments ...34
3-1.1 Isolation and Culture of Chondrocytes...34
3-1.1.1 Specimen Isolation ...34
3-1.1.2 Matrix Digestion...34
3-1.1.3 Cell Culture...35
3-1.2 Effective Single Compound Treatment ...36
3-1.3 LDH Cytotoxicity Assay for Cytotoxicity...37
3-1.4 MTT Assay for Cell Viability ...38
3-1.5 Total DNA for Cell Proliferation Quantification ...39
3-1.6 DMMB Assay for Quantitative Measurement of Sulfated GAGs Content...40
3-1.7 Real-time Reverse-Transcriptase Polymerase Chain Reaction for mRNA Expression Quantification ...41
3-1.8 Chemiluminescence Method for Detecting Free Radicals Scavenging ...47
3-1.9 Histochemical Staining Evaluation ...49
3-1.9.1 Hematoxylin & Eosin staining ...50
3-1.9.2 Alcian blue Staining...51
3-1.10 Immunohistochemical Staining Evaluation...52
3-1.10.1 S100 Protein Immunohistochemical Staining ...52
3-1.10.2 Type II Collagen Immunohistochemical Staining ...53
3-2 Three-dimensional Tissue-based Experiments ...56
3-2.1 Isolation and Culture of Chondrocytes...56
3-2.1.1 Specimen Isolation ...56
3-2.1.2 Matrix Digestion...56
3-2.1.3 Cell Culture...57
3-2.2 Preparation of Modified Tri-copolymer Scaffolds ...58
3-2.3 Seeding and Culturing Porcine Chondrocytes in the Modified Tri-Copolymer Scaffolds ...59
3-2.4 Preparation of scaffold for scanning electron microscopy (SEM) ...60
3-2.5 WST-1 Assay for Cell Proliferation in Cell-Scaffold Hybrids ...60
3-2.6 Total DNA for Cell Proliferation Quantification ...61
3-2.7 DMMB Assay for Quantitative Measurement of Sulfated GAGs Content...62
3-2.8 Real-time Reverse-Transcriptase Polymerase Chain Reaction for mRNA Expression Quantification ...63
3-2.9 Statistical analysis...68
CHAPTER 4 RESULTS ...69
4-1 Two-dimensional Chondrocytes Culture ...69
4-1.1 LDH Cytotoxicity Assay ...69
4-1.2 MTT Cell Proliferation Assay ...71
4-1.3 Total DNA for Cell Proliferation Quantification ...73
4-1.4 DMMB Assay for Sulfated Glycosaminoglycans Content...75
4-1.5 Real-time Reverse-Transcriptase Polymerase Chain Reaction for mRNA Expression Quantification ...78
4-1.5.1 Gene Expression of Type I collagen...78
4-1.5.2 Gene Expression of Type II collagen...80
4-1.5.3 Gene Expression of Aggrecan ...82
4-1.5.4 Gene Expression of Decorin...84
4-1.5.5 Gene Expression of Sox9 ...86
4-1.5.6 Gene Expression of MT1-MMP...88
4-1.5.7 Gene Expression of MMP-2...90
4-1.5.8 Gene Expression of T1MP-1 ...92
4-1.5.9 Gene Expression of IL-1beta...94
4-1.5.10 Gene Expression of TGF-beta1 ...96
4-1.6 Chemiluminescence Method for Detecting Free Radicals Scavenging ...98
4-1.7 Histochemical Staining Evaluation ...101
4-1.8 Immunohistochemical Staining Evaluation...103
4-2 Three-dimensional Scaffold Culture...105
4-2.1 Scanning electron microscopy...105
4-2.2 WST-1 Assay for Cell Proliferation ...108
4-2.3 Total DNA for Cell Proliferation Quantification ...110
4-2.4 DMMB Assay for Sulfated Glycosaminoglycans Content...112
4-2.5 Real-time Reverse-Transcriptase Polymerase Chain Reaction for mRNA Expression Quantification ...115
4-2.5.1 mRNA Expression of Collagens...115
4-2.5.2 mRNA Expression of Proteoglycans ...118
4-2.5.3 mRNA Expression of ECM Regulators...120
4-2.5.4 mRNA Expression of Growth and Differentiation Factors ...123
4-2.5.5 mRNA Expression of Catabolic Cytokines ...126
CHAPTER 5 DISCUSSIONS ...127
5-1 Two-dimensional Chondrocytes Culture ...127
5-2 Three-dimensional Scaffold Culture...130
CHAPTER 6 CONCLUSIONS...132
REFERENCES...133
INDEX OF FIGURES
Figure 1 Cartilage covers each end of the bones of any joint. ...1
Figure 2 Cartilage is composed of chondrocytes and extracellular matrix...2
Figure 3 Diagram of proteoglycan aggregate...3
Figure 4 Degenerative processes in osteoarthritis cartilage... 11
Figure 5 Articular structures that are affected in osteoarthritis...12
Figure 6 Current osteoarthritis treatment options.. ...13
Figure 7 Targets for the development of disease- and symptom-modifying drugs for osteoarthritis...14
Figure 8 The central dogma of gene expressional molecular biology. ...15
Figure 9 The key concept of gene-based cartilage repair...16
Figure 10 Tissue engineering triad and its characteristic...17
Figure 11 Definition of the stem cells. ...18
Figure 12 Tissue engineering triad of tissue engineered cartilage. ...20
Figure 13 Generation of superoxide anion in the mitochondrial respiratory electron transport chain (ETC)...21
Figure 14 The biological functions of ROS constitute a paradox...23
Figure 15 Schematic representation of the often produced ROS in chondrocytes. 24 Figure 16 Schema of the antioxidant systems...25
Figure 17 A representative real time PCR amplification plot and some definiendum. ...41
Figure 18 Mechanisms of the SYBR® Green I Dye chemistry...42
Figure 19 Reactions leading to the chemiluminescence of luminol. ...47
Figure 20 The structures of hematoxylin and eosin ...50
Figure 21 The chemical structure of alcian blue. ...51
Figure 22 LDH cytotoxicity assay of aucubin-treated chondrocytes...69
Figure 23 LDH cytotoxicity assay of betulin-treated chondrocytes. ...70
Figure 24 MTT cell proliferation assay of aucubin-treated chondrocytes ...71
Figure 25 MTT cell proliferation assay of betulin-treated chondrocytes. ...72
Figure 26 Total DNA content of one-week chondrocytes treated with different concentrations of aucubin. ...73
Figure 27 Total DNA content of one-week chondrocytes treated with different concentrations of betulin...74
Figure 28 Linear standard curve of glycosaminoglycans standards ...75
Figure 29 Glycosaminoglycans content of one-week chondrocytes treated with different concentrations of aucubin...76
Figure 30 Glycosaminoglycans content of one-week chondrocytes treated with different concentrations of betulin. ...77 Figure 31 The mRNA expression of type I collagen of one-week chondrocytes under different concentrations of aucubin treatment. ...78 Figure 32 The mRNA expression of type I collagen of one-week chondrocytes under different concentration of betulin treatment ...79 Figure 33 The mRNA expression of type II collagen of one-week chondrocytes under different concentrations of aucubin treatment. ...80 Figure 34 The mRNA expression of type II collagen of one-week chondrocytes under different concentration of betulin treatment ...81 Figure 35 The mRNA expression of aggrecan of one-week chondrocytes under different concentration of aucubin treatment. ...82 Figure 36 The mRNA expression of aggrecan of one-week chondrocytes under different concentration of betulin treatment...83 Figure 37 The mRNA expression of decorin of one-week chondrocytes under different concentration of aucubin treatment ...84 Figure 38 The mRNA expression of decorin of one-week chondrocytes under different concentration of betulin treatment...85 Figure 39 The mRNA expression of Sox9 of one-week chondrocytes under
different concentration of aucubin treatment ...86 Figure 40 The mRNA expression of Sox9 of one-week chondrocytes under
different concentration of betulin treatment...87 Figure 41 The mRNA expression of MT1-MMP of one-week chondrocytes
under different concentration of aucubin treatment...88 Figure 42 The mRNA expression of MT1-MMP of one-week chondrocytes
under different concentration of betulin treatment ...89 Figure 43 The mRNA expression of MMP-2 of one-week chondrocytes under different concentration of aucubin treatment ...90 Figure 44 The mRNA expression of MMP-2 of one-week chondrocytes under different concentration of betulin treatment...91 Figure 45 The mRNA expression of TIMP-1 of one-week chondrocytes under different concentrations of aucubin treatment ...92 Figure 46 The mRNA expression of TIMP-1 of one-week chondrocytes under different concentration of betulin treatment...93 Figure 47 The mRNA expression of IL-1beta of one-week chondrocytes under different concentration of aucubin treatment. ...94 Figure 48 The mRNA expression of IL-1beta of one-week chondrocytes under different concentration of betulin treatment...95
Figure 49 The mRNA expression of TGF-beta1 of one-week chondrocytes under
different concentration of aucubin treatment ...96
Figure 50 The mRNA expression of TGF-beta1 of one-week chondrocytes under different concentration of betulin treatment...97
Figure 51 The free radicals scavenging effect of different concentrations of aucubin on lucigenin-dependent chemiluminescence of one-week chondrocytes. ...99
Figure 52 The free radicals scavenging effect of different concentrations of betulin on lucigenin-dependent chemiluminescence of one-week chondrocytes. ...100
Figure 53 H&E staining of one-week cultured chondrocytes (100×). ...101
Figure 54 Alcian Blue staining of one-week cultured chondrocytes (40×) ...102
Figure 55 S-100 protein antibody immunochemical stain of one-week cultured chondrocytes (200×) ...103
Figure 56 S-100 protein antibody immunochemical stain of one-week cultured chondrocytes (100×). ...104
Figure 57 type II collagen antibody immunochemical stain of one-week cultured chondrocytes (200×). ...104
Figure 58 An SEM photograph of gelatin/C6S/C4S/HA modified tricopolymer scaffold (400×)...105
Figure 59 SEM photographs of cell-scaffold hybrids which were taken immediately after cell seeding ...106
Figure 60 SEM photographs of cell-scaffold hybrids which were taken at the end of 1-week culture...106
Figure 61 SEM photographs of cell-scaffold hybrids which were taken at the end of 2-week culture...107
Figure 62 SEM photographs of cell-scaffold hybrids which were taken at the end of 4-week culture...107
Figure 63 WST-1 assay of three-dimensional scaffold culture ...109
Figure 64 Total DNA assay of three-dimensional scaffold culture... 111
Figure 65 Linear standard curve of glycosaminoglycans standards ... 112
Figure 66 The GAG contents of cell-scaffold hybrids control groups and betulin-treated cell-scaffold hybrids experimental groups after cultured for 1, 2, and 4 weeks ...114
Figure 67 The GAG contents of the scaffolds without cells seeded negative control group after cultured for 1, 2, and 4 weeks were shown...114
Figure 68 At the end of 1, 2, and 4-week, the values of –△Ct of relative type II collagen gene expression were obtained ...116
Figure 69 At the end of 1, 2, and 4-week, the values of –△Ct of relative
type I collagen gene expression were obtained...116 Figure 70 At the end of 1, 2, and 4-week, the values of –△Ct of relative
type X collagen gene expression were obtained...117 Figure 71 At the end of 1, 2, and 4-week, the values of –△Ct of relative
aggrecan gene expression were obtained...118 Figure 72 At the end of 1, 2, and 4-week, the values of –△Ct of relative
decorin gene expression were obtained ...119 Figure 73 At the end of 1, 2, and 4-week, the values of –△Ct of relative
TIMP-1 gene expression were obtained ...120 Figure 74 At the end of 1, 2, and 4-week, the values of –△Ct of relative
MMP-2 gene expression were obtained...121 Figure 75 At the end of 1, 2, and 4-week, the values of –△Ct of relative
MT1-MMP gene expression were obtained...122 Figure 76 At the end of 1, 2, and 4-week, the values of –△Ct of relative
TGF-β1 gene expression were obtained ...123 Figure 77 At the end of 1, 2, and 4-week, the values of –△Ct of relative
BMP-7 gene expression were obtained...124 Figure 78 At the end of 1, 2, and 4-week, the values of –△Ct of relative
IGF-1 gene expression were obtained...125 Figure 79 At the end of 1, 2, and 4-week, the values of –△Ct of relative
IL-1β gene expression were obtained ...126
INDEX OF TABLES
Table 1 Inhibition profile of TIMPs. ...7
Table 2 Some methods for the characterization of cell death...29
Table 3 Twelve single chemical compounds extracted from 7 chondrogenesis related Chinese herbal medicines...30
Table 4 Four chondrogenesis-related single chemical compounds extracted from Plantago asiatica and Ampelopsis brevipedunculata (Maxim.) Trautv. ....31
Table 5 Experiment Design ...33
Table 6 List of target genes for real-time RT-PCR in two-dimensional chondrocytes culture ...43
Table 7 Primer Sequences of two-dimensional chondrocytes culture...44
Table 8 List of target genes for real-time RT-PCR in three-dimensional scaffold culture...64
Table 9 Primer Sequences of three-dimensional scaffold culture. ...65
Table 10 Summary of two-dimensional chondrocytes culture results ...129
Table 11 Summary of three-dimensional scaffold culture results ...131
LIST OF ABBREVIATIONS ASCs Adult Stem Cells
AUC Area Under Curve
BMPs Bone Morphogenetic Proteins CAT Catalase
CL Chemiluminescences C4S Chondroitin-4-Sulfate C6S Chondroitin-6-Sulfate
cDNA Complementary DNA
COX2 Cyclooxygenase 2
DNA Deoxyribonucleic Acid
DAB 3, 3’-Diaminobenzidine Tetrahydrochloride DMMB 1,9-Dimethyl-Methylene Blue
MTT 3-[4,5-Dimethylthiazol-2-Yl]-2,5-Diphenyltetrazolium Bromide DMEM Dulbecco/Vogt Modified Eagle's Minimal Essential Medium ETC Electron Transport Chain
EGF Epidermal Growth Factor
EDC 1-Ethyl-3-(3-Dimethylaminopropyl) Carbodiimide EDTA Ethylenediamine Tetraacetic Acid
ECM Extracellular Matrix FGFs Fibroblast Growth Factors GI Gastrointestinal GPx Glutathione Peroxidase
GAPDH Glyceraldehyde 3-Phosphate Dehydrogenase GAGs Glycosaminoglycans
H&E Hematoxylin & Eosin
HCl Hydrogen Chloride
H2O2 Hydrogen Peroxide
•OH Hydroxyl Radical
OCl⎯ Hypochlorite Ion
iNOS Inducible NOS
IGF Insulin-like Growth Factor IL-1β Interleukin-1β
ICE Interleukin-converting Enzyme
LDH Lactate Dehydrogenase
MMP-2 Matrix Metalloproteinase-2 MMP-9 Matrix Metalloproteinase-9 MMPs Matrix Metalloproteinases
MT-MMPs Membrane-type Matrix Metalloproteinases MSCs Mesenchymal Stem Cells
NBF Neutral Buffered Formalin
NAD+ Nicotinamide Adenine Dinucleotide
NADP Nicotinamide Adenine Dinucleotide Phosphate NADPH Nicotinamide Dinucleotide Phosphate
•NO Nitric Oxide
NOS NO Synthase
NSAID Non-steroidal Anti-inflammatory Drug NF-κB Nuclear Factor-ΚB
O.D. Optical Density
OA Osteoarthritis PRDX Peroxiredoxins
PBS Phosphate-buffered Saline PMT Photo Multiplier Tube ROS Reactive Oxygen Species
PCR Real-time Polymerase Chain Reaction
RT-PCR Reverse Transcription Polymerase Chain Reaction RNase Ribonuclease
RNA Ribonucleic Acid
SEM Scanning Electron Microscopy
Sox-5 Sex-determining Region Y-Box-5 Sox-6 Sex-determining Region Y-Box-6 Sox-9 Sex-determining Region Y-Box-9 SMADs Small Mother Against Decapentaplegic sGAG Sulphate-glycosaminoglycans O2•⎯ Superoxide Anion
SOD Superoxide Dismutase
INT Tetrazolium Salt
CT Threshold Cycle
TIMP-1 Tissue Inhibitor Of Metalloproteinase-1 TIMPs Tissue Inhibitors of Metalloproteinases TGF-β Transforming Growth Factor-Β
TNF-α Tumor Necrosis Factor-Α COL1 Type I Collagen
COL2 Type II Collagen UV Ultraviolet
WST-1 Water Soluble Tetrazolium Salt
CHAPTER 1 INTRODUCTION
1-1 Cartilage Biology
1-1.1 Definition and Composition of Cartilage
Cartilage is an avascular, aneural and alymphatic connective tissue which belongs to the skeleton organ and covers the end of each bone of any joint (Figure 1) [1, 2]. This white-colored tissue has an elastic consistency to absorb mechanical shock and provides an extremely smooth bearing surface to facilitate daily motion between the bones [3], thereby preventing biomechanical damage caused by severe loading and protecting the underlying bones from mechanical stresses of normal joint use [4].
Figure 1 Cartilage covers each end of the bones of any joint [5].
In general, cartilage is composed of chondrocytes and extracellular matrix (ECM) [5]. The phenotypic cells, chondrocytes, are small cells with one oval-shaped nucleus and one to two nucleoli which only make up less than 10% of the total volume of cartilage and do not contribute to the mechanical properties of cartilage [6].
Chondrocytes attach themselves to the framework of ECM which they synthesize, but
do not form contacts with each other [7]. Because chondrocytes are derived from the same cell lineage as fibroblasts, chondrocytes are similar to fibroblasts [8].
The matrix is composed of a framework of macromolecules and is filled with water (Figure 2) [9]. The structural macromolecules are collagens, proteoglycans, and noncollagenous proteins which make up 30% to 40% of the wet weight of cartilage matrix [10]. Collagens form a tight collagen fiber network which contains highly hydrophilic gel of aggregated proteoglycans and other noncollagenous proteins in it and hence provide the tensile strength for cartilage [11]. And because of the hydrophilic property of this framework, the entrapped water and ions make up the remaining 60% to 70% of the wet weight of cartilage matrix [12]. In the cartilage tissue dry weight, collagens account for 50% to 60%, proteoglycans contribute 30% to 35%, and noncollagenous proteins are about 15% to 20% [13]. From the identification of electron microscopy, type II collagen consists 90% to 95% of total cartilage collagen, and the remaining percentage are lesser amounts of collagen types VI, IX, X and XI [14]. Type IX and type XI collagen have been reported to possess important roles in organizing and stabilizing the network of type II collagen [15].
Proteoglycans are composed of a protein core filament and one or more long glycosaminoglycan chains [16], which are unbranched polysaccharide chains of repeating disaccharides (Figure 3) [16, 17]. In each repeating disaccharide of glycosaminoglycan, one or more negative charges exist and hence attract positively charged ions or water molecules [18, 19]. Glycosaminoglycans that connect to proteoglycan core proteins include chondroitin sulfate, keratin sulfate, and dermatan sulfate [20]. When proteoglycans aggregate through the noncovalent interactions between hyaluronan filaments and link proteins, the resulting proteoglycan aggregate stabilizes these large macromolecules within the matrix (Figure 3) [21].
Figure 3 Diagram of proteoglycan aggregate.
The noncollagenous proteins in cartilage have not been extensively studied but seem to have some effects in organizing the matrix and maintaining the relationships between chondrocytes and the macromolecules of the matrix [11].
1-1.2 Types of Cartilage
There are three basic types of cartilage: articular cartilage, fibrous cartilage, and elastic cartilage [22]. Articular cartilage, also called hyaline cartilage, is the most common type among the three types of cartilage. It exists in articular surfaces of the joints, trachea tube rings, larynx, rib, costae, and the tip of the nose [23].
Perichondrium covers articular cartilage and provides nutrients to chondrocytes through the blood vessels in it [24]. Chondrocytes are entrapped in the lacunae structure [25]. The matrix of articular cartilage is smooth and basophilic, and the framework of macromolecules is mainly type II collagen [14].
Fibrous cartilage never occurs alone and exists in locations where high stress occurs such as the intervertebral disks, in front of the pelvic girdle between the pubic bones, and the glenoid cavity in the shoulder joint [26, 27]. It closely adheres to either articular cartilage or dense connective tissue. In fibrous cartilage, perichondrium is often absent. The chondrocytes are less in number and often widely separated, but are still enfolded in lacunae. The matrix of fibrous cartilage is mainly type I collagen [28, 29].
Elastic cartilage exists in the places where the maintenance of specific shapes is important, such as auricular, epiglottis, and the walls of the eustachian tube [30].
Chondrocytes in elastic cartilage are more tightly arranged than in articular cartilage.
And because the percentage of the matrix in elastic cartilage is higher than in articular cartilage, elastic cartilage can be easily deformed and spring back into the original shape. The major component of matrix macromolecules is elastin [31].
1-1.3 Homeostasis of Cartilage
Although there is only one type of cell in cartilage, like other tissues, cartilage still needs to keep its homeostasis [32]. Therefore, chondrocytes continually synthesize, combine and degrade ECM proteins to maintain the biochemical composition among cells, and in some manner are influenced and regulated by the mechanical and biochemical signs of ECM which they secrets [33]. This concept, in which chondrocytes maintain the balance and turnover between ECM and themselves, has been called dynamic reciprocity [34].
Under normal physiological condition, the proteins relates to ECM express continuously at constant level. Type II collagen is the most abundant protein in cartilage and constitute the basic framework of ECM. In addition to type II collagen, the fibril network of healthy cartilage also contains type IX and type XI collagen, and there are still small amounts of other types of collagen such as type III, VI, XII, and XIV collagen found in cartilage[35]. Type II and type VI collagens are demonstrated to possess the RGD sequences, which suggest the role in facilitating cell attachment [36]. However, when normal articular cartilage is somehow worn away, the wound healing process of cartilage prefers to deposit type I collagen rather than type II collagen in the newly formed ECM, which makes the articular cartilage become fibrous cartilage [37]. Contrary to type I and type II collagen which are synthesized by chondrocytes, type X collagen is synthesized by hypertrophic chondrocytes which play an important role in endochondral ossification [38]. Hypertrophic chondrocytes direct the mineralization of their surrounding ECM, lead adjacent perichondrial cells to become osteoblasts, and finally undergo apoptotic cell death [39].
The balance in the expression of matrix metalloproteinases (MMPs) and their inhibitors, tissue inhibitors of metalloproteinases (TIMPs), is another important factor for cartilage homeostasis [40]. Matrix metalloproteinases are a family of
zinc-dependent endopeptidases that possess the ability to degrade and remodel ECM [41]. There are 22 human MMPs and can be roughly divided into several groups:
collagenase such as MMP-1, MMP-8, and MMP-13, gelatinase such as MMP-2 and MMP-9, and membrane-type MMPs (MT-MMPs) such as MMP-14, 15, 16, 17, 24, and 25 [41, 42]. TIMPs inhibit MMPs in a noncovalent and reversible manner and can also positively regulate MMPs [41]. There are four types of TIMPs have been identified: TIMP-1, 2, 3, and 4, and each TIMP differs in its ability to inhibit or module different MMPs (Table 1) [41, 43]. TIMP-1 inhibits MMP-1 and MMP-3, while TIMP-2 and TIMP-3 inhibit MT1-MMP.
MMP-2, also known as gelatinase-A, is widely expressed in many noninflamed healthy connective tissues [44]. It is activated at the cell surface level to remodel the ECM and the basement membrane substrates such as laminin and fibronectin and hence modulate the migration and adhesion of cells. The activition of MMP-2 is tightly regulated by the expression of MT1-MMP (MMP-14) and TIMP-2 [45].
Table 1 Inhibition profile of TIMPs. (+) means the MMP is inhibited by the TIMP (Modified from the reference [41]).
Type of MMP TIMP-1 TIMP-2 TIMP-3 TIMP-4
MMP-1 + + + +
MMP-2 + + + +
MMP-3 + + + +
MMP-7 + + + +
MMP-8 + +
MMP-9 + + + +
MMP-10 + +
MMP-11 +
MMP-12 +
MMP-13 + + +
MMP-14 + +
MMP-15 + +
MMP-16 + +
MMP-17
MMP-19
MMP-20
1-2 Natural Limitations of Cartilage
1-2.1 Low Oxygen Tension on Cartilage Metabolism
Articular cartilage is a tissue without vascular invasion, so its nutrition and oxygen are mainly supplied by diffusion from the synovial fluid [46, 47]. Therefore, O2 tension in cartilage in vivo is only about 1% to 8% which is dramatically lower than in normal tissues where O2 tension is about 21% [48], and chondrocytes are hence adapted to working as anaerobic working cells [49]. The low O2 consumption in cartilage is because of the Crabtree effect, where the increased glucose concentrations inhibit the cellular respiration and limit the O2 consumption [50].
The natural low O2 tension in cartilage not only influences the homeostasis of ECM components but also stabilizes the phenotype of chondrocytes [51-56]. Under hypoxia environment, chondrocytes increase the synthesis of type II collagen and glycosaminoglycans and decrease the synthesis of type I collagen [51-54]. Moreover, MMP-9 mRNA is decreased by interleukin-1β (IL-1β), a catabolic cytokine, in low O2 tension environment [57].
1-2.2 Limited Cartilage Repair Response
Because the cartilage naturally lacks of the vascularity, it is in the very limited capacity of self-repair [4]. In most tissues, injury is associated with a rupture of blood vessels, an influx of cells and bioactive peptides from the blood, and the formation of a hematoma [58]. Undifferentiated mesenchymal progenitor cells migrate into the lesion, differentiate into the local cellular phenotype and synthesize repair tissue.
When lesions occur that are limited to the articular cartilage in adults, there is no bleeding and thus no mechanism for the replacement of lost or damaged tissue [59].
Neighboring chondrocytes may respond by local proliferation; however, because they are sequestered in the dense matrix, they do not migrate into the damaged region to fill the void. As such, focal lesions that are limited to the chondral layer usually remain for life. These can often expand with time and use, and lead to the more generalized cartilage loss associated with osteoarthritis (OA) [59, 60].
A repair response can occur when injury extends through the chondral layer to the subchondral bone and underlying vasculature. Local bleeding and fibrin clot formation causes an infiltration of bone marrow-derived cells which synthesize space-filling repair tissue. This may initially heal to resemble hyaline cartilage after 6–8 months, but in the longer term becomes increasingly populated with type I collagen, degenerating to a fibrocartilaginous scar tissue that does not have the appropriate physical and mechanical properties, and ultimately fails [61, 62].
1-3 Cartilage Pathology
Osteoarthritis (OA) is the most common disease of joints which affects millions of people all over the world [63]. It can occur in any joint but is most common in joints of the knee, shoulder, hand, spine, hip, and foot [4]. The development of OA starts in the cartilage degeneration, where the balance of anabolism and catabolism in cartilage is progressively disrupted by suppression in anabolic genes expression or increase in catabolic genes expression [4, 46, 64]. In this progression, OA eventually impairs the function of a whole joint, including the cartilage, the subchondral bone, the synovium and the periarticular connective and muscular tissues.
As we noted, cartilage continually maintains its homeostasis. Circumstances that impair chondrocytes function can disrupt the balance of synthesis and catabolism in favor of cartilage degradation, which over time can lead to OA [59].
The development of osteoarthritis begins at the degenerative processes in cartilage. Interleukin-converting enzyme (ICE) converts IL-1β to its active form and, therefore, activated IL-1β induces the expression of matrix proteases, which degrade the matrix components (Figure 4). The homeostasis in cartilage starts to be destroyed which contributes to the risk for cartilage degeneration by decreasing the ability of chondrocytes to maintain and repair the articular cartilage tissue [65]. The mitotic and synthetic activity of chondrocytes decline with advancing donor age [66]. In addition, human chondrocytes become less responsive to anabolic mechanical stimuli with ageing and exhibit an age-related decline in response to growth factors such as the anabolic cytokine insulin-like growth factor-I [66]. These findings provide evidence supporting the concept that chondrocytes senescence may be involved in the progression of cartilage degeneration.
Figure 4 Degenerative processes in osteoarthritis cartilage [4].
Metalloproteinases and ROS are the two main actors of matrix component degradation. They have been found to be overproduced in OA cartilage and synovium.
ROS may directly oxidize nucleic acids, transcriptional factors, membrane phospholipids, intracellular and extracellular components leading to impaired biological activity, cell death and matrix components breakdown [67-69]. From the different in vitro and animal studies, Y. Henrotin et al. concluded that in pathological circumstances, ROS contribute to cartilage degradation by directly degrading matrix components, by sustaining the activity of catabolic cytokines and by reducing cartilage repair capacities. Altogether, these observations support the concept of antioxidant therapy in rheumatic diseases and it might also decrease the progression of OA [70].
In severe osteoarthritis, the pathological changes result in radiological changes, such as loss of joint space, subchondral bone sclerosis and presence of osteophytes (bony spurs mostly located at the joint margins) (Figure 5) [4, 60]. These changes can result in joint symptoms such as pain, stiffness and loss of function. The symptoms
vary with time and between joint sites and individuals. The main risk factors for OA are age, obesity and any form of joint trauma. In some families, OA seems to be inherited [60].
Figure 5 Articular structures that are affected in osteoarthritis. a, Normal cartilage is without any fissures, no signs of synovial inflammation. b, Early focal degenerate lesion and ‘fibrillated’ cartilage, as well as remodelling of bone, is observed in osteoarthritis. This can lead to bony outgrowth and subchondral sclerosis [4].
1-4 Current Strategies for Cartilage Treatment
1-4.1 Inadequate Pharmacological Options
The specialized architecture and limited repair capacity of articular cartilage coupled with the high physical demands on this tissue make it exceedingly difficult to treat medically. The current treatment options as issued in the guidelines from the American College of Rheumatology are fairly limited (Figure 6) [71, 72]. In addition to non-pharmaceutical measures such as weight loss and physical exercise they include only symptomatic treatment of limited efficacy with analgesics, non-steroidal anti-inflammatory agents or intra-articular administration of steroids or hyaluronic acid. Because no drugs exist that prevent or halt osteoarthritis joint destruction, the ultimate measure is joint replacement.
Figure 6 Current osteoarthritis treatment options. Abbreviations: GI, gastrointestinal;
COX2, cyclooxygenase 2; NSAID, non-steroidal anti-inflammatory drug [4].
Chronic pain in osteoarthritis patients causes primarily on the activation of sensory neurons that innervate the affected joint. With the exception of cartilage, all joint tissues, including subchondral bone and synovium, are densely supplied by small-diameter nociceptive neurons (Figure 7) [4, 71]. Tissues, including subchondral bone and synovium, are densely supplied by small-diameter nociceptive neurons.
Potential targets for osteoarthritis is to interfere the nociception and possible pathways. Inhibiting the production of the inflammatory cytokine IL-1β or blocking its receptors or interrupting its subsequent intracellular signalling through nuclear factor-κB (NF-κB) and the blockade of bradykinin receptors are more recent approaches to developing symptom-modifying drugs with greater efficacy than non-steroidal anti-inflammatory drugs (NSAIDs) that inhibit the formation of the pain mediator prostaglandin E2 (Figure 7) [4].
Figure 7 Targets for the development of disease- and symptom-modifying drugs for osteoarthritis. Nociception and possible ways are targets to be interfered by pain relief
1-4.2 Novel Gene-based Approach for Cartilage Repair
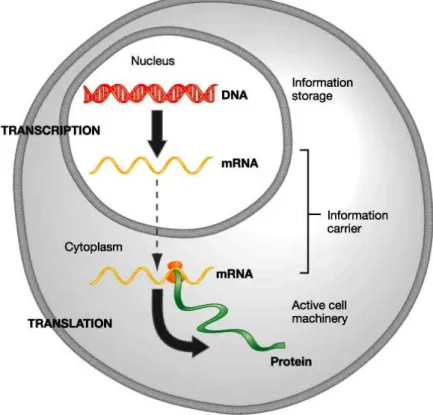
Existing pharmacologic, surgical and cell based treatments may offer temporary relief but are incapable of restoring damaged cartilage to its normal phenotype. Gene transfer provides the capability to achieve sustained, localized presentation of bioactive proteins or gene products to sites of tissue damage (Figure 8).
Figure 8 The central dogma of gene expressional molecular biology: Transcription of DNA to RNA and translation of RNA to protein.
A variety of cDNAs have been cloned which may be used to stimulate biological processes that improve cartilage healing. They are anabolic growth factors such as transforming growth factor (TGF-β superfamily), several of the bone morphogenetic proteins (BMPs), insulin-like growth factor (IGF), fibroblast growth factors (FGFs), and epidermal growth factor (EGF). Transcription factors such as Sox-9, L-Sox 5 and
Sox-6 that promote chondrogenesis or the maintenance of the chondrocyte phenotype present another class of biologics that may be useful. Signal transduction molecules, such as SMADs are also known to be important in chondrocyte differentiation.
Because these regulatory molecules function intracellularly and cannot be delivered to cells in soluble form, gene transfer is perhaps the only way to which they might be directly applied for medical use [73, 74].
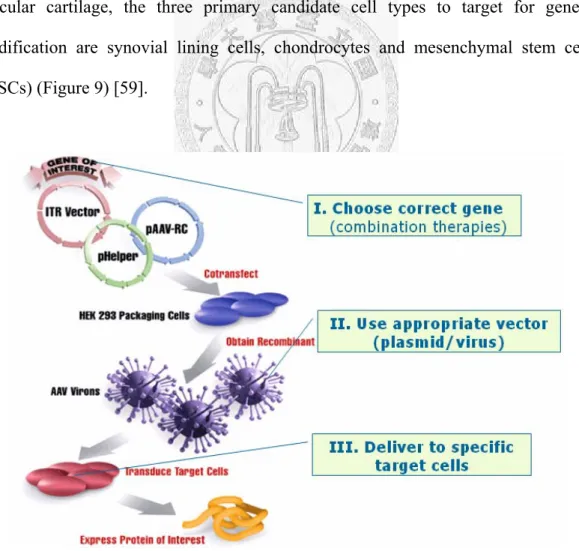
The challenge to gene-based treatment strategies is to devise methods that incorporate the correct gene or gene combination with the appropriate vector, delivered to specific target cells within the proper biological context to achieve a meaningful therapeutic response. Toward the treatment and repair of damaged articular cartilage, the three primary candidate cell types to target for genetic modification are synovial lining cells, chondrocytes and mesenchymal stem cells (MSCs) (Figure 9) [59].
Figure 9 The key concept of gene-based cartilage repair. It provides a way to convert
1-5 Cartilage Tissue Engineering
Compared with the former two strategies for cartilage repair, the cell-based cartilage tissue engineering seems a better way to repair the cartilage for several reasons. One of them is that strategies for engineering cartilage tissues are mainly focused on the restoration of pathologically altered structures, which were based on the transplantation of the so-called tissue engineering triad: cells in combination with supportive matrix and biomolecules [73, 75]. Since this approach comprises the interactive tissue engineering triad of responsive cells, a supportive matrix template, bioactive molecules promoting differentiation, it can mimic and further regenerate the tissue structure (Figure 10) [76]. Moreover, the cell source can origin from autologous cells or stem cells, and the cell number can be further augmented in vitro environment.
Furthermore, there are already lots of biodegradable materials can be used clinically.
The detailed descriptions are in following paragraphs.
Figure 10 Tissue engineering triad and its characteristic.
1-5.1 Cell
The cell is the first important component of the tissue engineering triad. Just as mentioned, cells can be obtained from autologous cells or stem cells. Although the cell used in cartilage tissue engineering triad can be directly obtained from patients, chondrocytes are difficult to isolate in humans, and they are also replicate in a slow rate and are prone to phenotypic dedifferentiation in vitro culture [77]. This can be further affected by donor age and health status [78]. In view of this, tissue engineering based on adult stem cells (ASCs) for tissue engineering cartilage is highly considered recently. Stem cells are highly replicative cells that have multilineage differentiation capacity (Figure 11). ASCs are less tumorogenic than their embryonic stem cells and are accessible from many tissues including bone, deciduous teeth, adipose tissue, umbilical cord blood, synovium, brain, blood vessels and blood [79-81]. To date, mesenchymal stem cells can not only derive from the bone marrow, but can also obtain from adipose tissue, muscle, skin and synovium. They are capable of self-renewal and can differentiate into several different phenotypes including bone, cartilage, adipocytes, and haematopoiesis supporting stroma [74, 82].
1-5.2 Biomaterial Scaffold
A critical requirement of cartilage tissue engineering is the design of specific biomaterials and scaffold structures. Biomaterial scaffolds control three-dimensional shape, guide tissue development and permit the convenient delivery of cells into patients [83]. For cartilage and intervertebral disc engineering, a suitable biomaterial should provide or support initial mechanical stability, even cell distribution and good tissue biocompatibility [84].
Increasing evidence suggests that three-dimensional cell cultures provide the advantage of anchorage-independent cell growth, maintaining the differentiated phenotype that allows the synthesis of cell-specific pericellular or intercellular matrix.
Furthermore, the macromolecular assembly of newly synthesized collagens and proteoglycans is critical for tissue engineering, as initially laden matrix serves as a template for subsequent matrix deposition and architecture. The polymers such as collagen, chondroitin sulfate, hyaluronate, fibrin, agarose, alginate and chitosan are often used in tissue engineering applications (Figure 12) [83]. Inclusion of chondroitin sulfate in scaffold may promote the secretion of proteoglycan and type II collagen [85], and using of hyaluronic acid can facilitates the integration of cells to the engineered cartilage [86].
Figure 12 Tissue engineering triad of tissue engineered cartilage.
1-5.3 Chondrogenesis Signals
A multi-factors network of metabolic pathways governs chondrogenesis (Figure 12) [60]. TGF-β, BMPs, and the sonic hedgehog gene (members of TGF-β superfamily) were found to enhance chondrogenesis. Insulin-like growth factor was shown to have a synergistic effect with a member of the TGF-β family in promoting progenitor cell chondrogenesis. FGF-2 also has a role in chondrogenesis, inducing MSC proliferation and promoting retention of multilineage differentiation capacity [60].
CHAPTER 2 THEORETICAL BASIS
2-1 Reactive Oxygen Species in Cartilage
2-1.1 Introduction of Reactive Oxygen Species
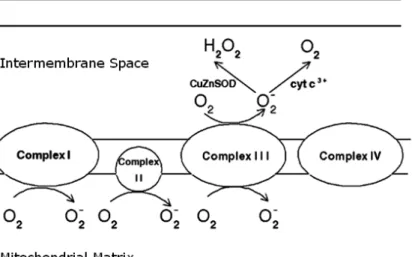
Reactive oxygen species (ROS) are a group of oxygen-containing and highly reactive species which are partially reduced from molecular oxygen, including molecules such as hydrogen peroxide (H2O2), radicals such as hydroxyl radical (•OH), ions such as hypochlorite ion (OCl⎯), and both an ion and a radical such as superoxide anion (O2•⎯) [46, 87]. Under normal physiological conditions, the formation of ROS is a natural consequence of cellular metabolism, where about 1% to 3% of oxygen uptook by the body is reduced into ROS [88, 89]. The major source of ROS exists in the mitochondrial aerobic respiratory electron transport chain (ETC). Among four multi-protein complexes of ETC, Complex I, II, and III were believed to produced superoxide anion byproduct by leaking electrons to oxygen (Figure 13) [90, 91].
Figure 13 Generation of superoxide anion in the mitochondrial respiratory electron transport chain (ETC). (Modified from reference [91])
After reducing oxygen by one electron, the produced superoxide anion (O2•⎯) can serve as the precursor of most other ROS. For example, superoxide anion can undergo dismutation to become hydrogen peroxide (H2O2), which can be further partially reduced to one of the strongest oxidants, hydroxyl radical (•OH) [92].
Furthermore, superoxide anion may also react with other radicals to produce new oxidants, such as peroxynitrite, an oxidant produced by the reaction of superoxide anion and nitric oxide (•NO) [93]. In addition to generating ROS by mitochondrial respiratory system, microsomal cytochrome P450 enzymes, flavoprotein oxidases, and peroxisomal enzymes involved in fatty acid metabolism also serve as the intracellular sources of ROS [88, 94].
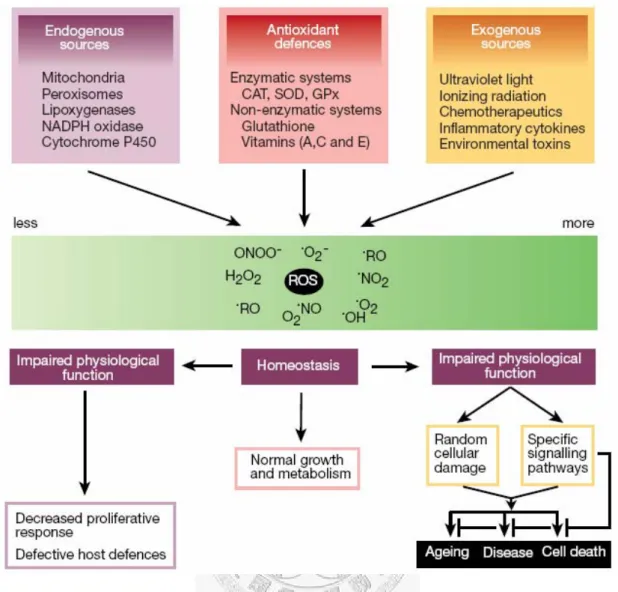
According to current research, the biological functions of ROS constitute a paradox. They are not only involved in signal transduction and homeostasis of tissue turnover but also involved in cell death and cellular degradation (Figure 14) [67, 95].
Under normal physiological conditions, the toxic effects of ROS are prevented by various cellular anti-oxidant systems, which mean that the production of ROS and the scavenging of ROS by antioxidants are in balance. Therefore, the resulting low concentration of ROS within cells act as intracellular second messenger molecules which regulate the expression of a number of genes, such as ECM components, MMPs, and cytokines [96, 97]. However, in several pathological circumstances, the ROS are somehow over-produced and the antioxidants defenses become insufficient, which lead to destroy the balance between intracellular and extracellular redox state [67, 98]. The resulting so-called “oxidative stress” is an abnormal catabolic state which can induce structural or functional changes in cells and tissues by oxidizing polyunsaturated fatty acids to increase membrane fluidity and permeability, oxidizing guanine to 8-hydroxyguanine to cause DNA damage, and oxidizing amino acids such
Figure 14 The biological functions of ROS constitute a paradox.
2-1.2 Reactive Oxygen Species Formation in Cartilage
The often produced ROS in chondrocytes are superoxide anion (O2•⎯) and nitric oxide (•NO), which can further generate derivative radicals such as ONOO⎯ and H2O2
(Figure 15) [102, 103]. Although O2•⎯ in vivo can be enzymatically or nonenzymatically produced, in cartilage, O2•⎯ is produced by the nicotinamide dinucleotide phosphate (NADPH) oxidase, which is a complex enzyme system consisting two membrane bound peptides: one is a two-peptide formed flavocytochrome, the other is a regulatory peptide called (Rap1A) [104].
Similarly, •NO in cartilage is also produced by an enzyme system, called NO synthase (NOS) [105]. Among three isoforms of NOS, chondrocytes express endothelial NOS (eNOS) and inducible NOS (iNOS) to generate nitric oxide [106]. On the other hand, in an in vitro environment, it was reported that O2•⎯ may be generated by tumor necrosis factor-α (TNF-α) stimulation or by cyclic stretch [107], and •NO generation may be stimulated by IL-1β, TNF-α, shear stress, or mechanical compression.
Figure 15 Schematic representation of the often produced ROS in chondrocytes.
(Modified from references [46] and [67] )
2-1.3 Antioxidant Systems in Cartilage
In order to prevent the toxic effects of ROS, chondrocytes possess various anti-oxidant defensive systems to scavenge the over-produced ROS, which include superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and peroxiredoxins (PRDX) (Figure 16) [108-111]. SODs are metalloproteins which possess metals in their reactive centers [112]. In chondrocytes, cytosolic Cu/Zn SOD and mitochondrial Mn SOD are constitutively expressed to catalyze two superoxide anions into one hydrogen peroxide and one molecular oxygen [113]. Because O2•⎯ is the most frequent and abundant ROS produced by normal cellular metabolism, the breakdown of O2•⎯ becomes the first defense mechanism against ROS [112]. However, the resulting hydrogen peroxide is also another kind of ROS and its accumulation is regarded as an oxidative stress as well. Therefore, catalase and glutathione peroxidase coexist and are functioned as scavengers of H2O2 [113, 114]. Besides this, peroxiredoxins, a newly found peroxidase family which possesses six isoforms in mammals, expresses type V isoform and eliminates H2O2 in cartilage [111].
Figure 16 Schema of the antioxidant systems. (Modified from reference [91, 115])
2-1.4 Reactive Oxygen Species on Cartilage Matrix Degradation
In pathological conditions, such as OA, reactive oxygen species have been described as an important factor on cartilage matrix degradation because they can cause cellular or organic changes by either directly oxidizing the amino acids of collagen and proteoglycan molecules or by modifying the expression of anabolic and catabolic related enzymes [46, 99-101, 116]. From several in vitro experiments, it have been reported that ROS directly attacked ECM components. For example, when treating the incubated type I collagen with O2•⎯, collagen was degraded and the ability of fibrils formation was lost [117, 118]. Moreover, in the •OH and oxygen coexisting environment, collagen was degraded into small peptides where the cleavages often happened on proline or 4-hydroxyproline residues, and the increase of glutamic acid and aspartic acid in the peptides were also observed [119]. Furthermore, HOCl was discovered to possess the ability to cleave hyaluronic acid and hence decrease the viscosity of synovial fluid [120]. On the other hand, some data also showed that ROS can change the response of chondrocytes to ECM-related anabolic enzymes or upregulate the expression of ECM-related catabolic enzymes. For instance, the concurrent generation of O2•⎯ and •NO could decrease the sensitivity of chondrocytes to IGF-1 and hence inhibit the synthesis of proteoglycan [121, 122]. Moreover, •NO could inhibit the sulfation of newly synthesized proteoglycan under the induction of IL-1 [123], and •NO also played an important role in activation of metalloprotease enzymes in articular cartilage [124]. Besides this, HOCl could directly activate the proenzymes of metalloprotease and inactivate the tissue inhibitors of metalloprotease (TIMPs) [125, 126].
2-1.5 Reactive Oxygen Species on Cartilage Senescence
Oxidative stress is an important factor of the aging process in cartilage. Because adult articular cartilage is considered as a post-mitotic tissue with the characteristic of limited ability to achieve tissue turnover [127], its aging process is not attributed to intrinsic replicative senescence accompanying with telomere shortening but is related to oxidative stress-induced extrinsic senescence [66, 128]. Several reports showed that the ability of chondrocytes to detoxify ROS and turnover the damaged macromolecules is dramatically inefficient over time [112]. Therefore, oxidative damage products such as peroxidated lipids [129], nitrotyrosine [130], and mutagenic 8-oxoguanine [131] gradually accumulate in chondrocytes and disturb normal cellular functions until the chondrocytes can not endure these structural changes. Notably, the production of mutagenic 8-oxoguanine has been reported to result from O2•⎯ attack which directly cause telomere erosion by injuring the guanine repeats in telomere DNA [132]. Moreover, it also has been reported that cell aging process correlates with the degeneration of mitochondria which may lead further leakage of electron transport chain and hence increase the production of ROS [133]. The accumulated ROS in mitochondria then attack mitochondrial 16-kilobase circular DNA, and the resulting mtDNA mutations contribute to the feature of accelerated aging [134].
2-1.6 Reactive Oxygen Species on Chondrocyte Death
In adult tissues, cell death plays an important role in maintenance of the balance of cell proliferation and the constancy of cell numbers [135]. Apoptosis is an active process of programmed cell death which is characterized by a series of morphological changes including DNA fragmentation, chromatin condensation, nucleus break-up, and cell fragmentation, while necrosis is the accidental death of cells that resulting from a pathologic injury [136].
It has been noted that aging [110], mechanical damage [137], and pathology such as osteoarthritis [138] often result in increased chondrocyte death in cartilage, and because of adult cartilage has limited ability for self-repair, the damaged cells may further accelerate the progression of the lesion [139]. From Blanco et al.’s research, it has been revealed that •NO is a primary inducer of chondrocyte apoptosis and is down-regulated by caspase-3 and tyrosine kinase activation [140]. However, from recent research, it has revealed that the toxicity of •NO needs to be further modulated by other ROS. For example, Del Carlo et al.’s research showed that the chondrocyte cell death mediated by •NO requires the generation of O2•⎯, which suggests that ONOO⎯ plays an important role in the process [141]. Moreover, in Kurz et al.’s finding, they found that both O2•⎯ and H2O2 play important roles in mechanically induced cell death [142]. From these examples, it might be suggested that O2•⎯ also plays an important role as •NO in chondrocyte cell death [46].
Several methods have been developed to detect cell death. However, it is often not sufficient to distinguish between apoptosis and necrosis by only applying a single assay. Therefore, combination of two to three methods is necessary to measure the morphological changes as well as the intracellular cell death parameters. Table 1 summarizes some methods for the characterization of cell death [136].
Table 2 Some methods for the characterization of cell death [136].
Cell Death Parameter Method Specific
Detection of Apoptosis Loss of cell attachment Staining of cells with crystal violet or
fluorescent DNA-binding dyes
-
Cellular ultrastructure Electron Microscopy + Externalization of
phosphatidyl serine
Annexin V binding -
Release of cytosolic compounds
51Cr release, 3H-labeled proteins, enzymatic activities in culture supernatants
-
Uptake of dyes Vital dyes (counting of cells) or fluorescent dyes for FACS
-
DNA laddering Agarose gel electrophoresis +
In situ DNA cleavage TUNEL -
Nuclear condensation and fragmentation
DAPI +
DNA degradation DNA content in sub G1 cells (FACS), agarose gel electrophoresis, DNA fragmentation ELISA
-
Internucleosomal DNA fragmentation
DNA fragmentation ELISA, agarose gel electrophoresis
+
Oxidative phosphorylation
MTT, Alomar Blue -
Mitochondrial
membrane polarization
Aggregation, uptake, sequestration of fluorescent dyes
-
Caspase activity Conversion of fluorogenic substrates + Caspase processing Western blot, immunhistochemistry + Cleavage of caspase
substrates
Western blot, immunhistochemistry +
2-2 Chondrogenesis-related Chinese Herbal Medicine
Circumstances that impair chondrocyte function can disrupt the balance of synthesis and catabolism in favor of cartilage degradation, which over time can lead to OA. The changes can result in joint symptoms such as pain, stiffness and loss of function. The symptoms vary with time and between joint sites and individuals. The main risk factors for OA are age, obesity and any form of joint trauma. In some families, OA seems to be inherited [143].
In Chinese herbal medicines, a number of herbs are used for treating joint symptoms in a long time treatment, and more and more researches have shown that the effective chemicals extracted from them possess various biological activities [144, 145]. From traditional Chinese herbal medicine ancient literatures and records, we preliminarily compared the prescriptions which have been used to treat the degeneration of the cartilage and targeted on 12 single chemical compounds which are extracted from 7 chondrogenesis-related Chinese herbal medicines (Table 1).
Table 3 Twelve single chemical compounds extracted from 7 chondrogenesis-related Chinese herbal medicines.
Then, by furthering systematic computer-aided search of the Medline database for the period 1990 through June 2006 and considering the commercializing achievability, we investigated the involved mechanism of 2 major single chemical compounds extracted from Plantago asiatica and Ampelopsis brevipedunculata (Maxim.) Trautv. by focusing on their antioxidant activity on cartilage tissue engineering. (Table 2)
Table 4 Four chondrogenesis-related single chemical compounds extracted from Plantago asiatica and Ampelopsis brevipedunculata (Maxim.) Trautv.
2-3 Purpose of this study
The objective of this study was to modify the current cartilage tissue engineering triad and find a better combination for regeneration the articular cartilage in vitro environment. We searched for the new signals which are more stable in
structure and cheaper in price than the traditionally used signals such as TGF-β1 and IGF, and suggested that because of the ROS were scavenged in cultured chondrocytes, the life span of cartilage tissue hybrids would be longer. Also, we modified the components of original gelatin-C6S-HA tricopolymer to the one which is more mimic the really ECM in organisms. We hope that the newly designed cartilage tissue engineering triad will improve the antioxidant and proliferation activity of chondrocytes.
In the beginning, we had to find the optimal concentration of these single chemical compounds when they were added in culture medium, and the experiment is divided into two parts: one part is 2D cell-based experiment, and the other part is 3D tissue engineering. In the 2D cell-based experiment, the important role in scavenging free radicals by these extracted chemical compounds was detected by the chemiluminescence method, and the proliferation and matrix productivity of chondrocytes were evaluated by real-time reverse-transcriptase polymerase chain reaction, ELISA assays, and immuno-histochemical staining. In the 3D tissue engineering experiment, also, the proliferation and matrix productivity were double evaluated by ELISA assays, real-time reverse-transcriptase polymerase chain reaction, and immuno-histochemical staining.
Table 5 Experiment Design.
CHAPTER 3 MATERIALS AND METHODS
3-1 Two-dimensional Cell-based Experiments
3-1.1 Isolation and Culture of Chondrocytes 3-1.1.1 Specimen Isolation
The articular cartilage used in the experiment was obtained aseptically from adult porcine knee joints which had no macroscopic signs of osteoarthritis. Within 4 h of slaughter, the integral part of adult porcine knee joint was cleaned by 75% ethanol (E7148, Sigma Co., St. Louis, MO, U.S.A.), iodine solution (35089, Sigma Co.) and phosphate-buffered saline (PBS, pH 7.2) in sequence. After trimming the undesired connective tissue with sterile scalpels, the cartilage tissue was sliced into 5mm×5mm×0.5mm pieces, and the action of slicing was avoided cutting deep into the subchondral bone [146]. Then the thin slices were washed three times in sterile PBS and were further sterilized by soaking in 5-fold antibiotics-added (50 units/mL penicillin, 50 μg/mL streptomycin, 100 μg/mL neomycin, P4083, Sigma Co.) PBS for 15 minutes.
3-1.1.2 Matrix Digestion
Enzymatic digestion of articular cartilage has been a feasible and useful technique for isolating chondrocytes based on degrading the extracellular matrix between cells. [147, 148]. In this experiment, the thin slices of cartilage were subjected to 0.2% collagenase (C0130, Sigma Co.) digestion in order to degrade the native collagen between animal cells and get the sufficient amount of cells after digestion [149-151]. The collagenase was dissolved in DMEM (31600-026, Gibco
Invitrogen Co., Burlington, Ontario, Canada) which was supplemented with 3.7 mg/ml sodium bicarbonate (S5761, Sigma Co.), 50 μg/ml ascorbic acid (A5960, Sigma Co.) [152], 50 units/50 μg/100 μg/mL penicillin/ streptomycin/ neomycin (P4083, Sigma Co.), and 10% heat-inactivated fetal bovine serum (100-106 Gemini Bio-Products, Woodland, CA) and filter-sterilized by passing through a 0.22 μm filter (SCGPT05RE, Millipore Co., Bedford, MA, U.S.A.). After incubating the sliced cartilage in a humidified incubator at 37°C, 5% carbon dioxide for 16 hours, the resulting chondrocytes/collagenase solution suspension was collected and centrifuged at 1500 rpm for 5 min (KUBOTA 5101, Canada). The supernatant was discarded and the pelleted cells were gently washed once in PBS to neutralize remaining collagenase.
Cells were centrifuged at 1500 rpm for 5 min again and resuspended in 10 ml of the same medium mentioned above. Cell number and viability was assessed by hemocytometer and trypan blue dye (T8154, Sigma Co.) exclusion.
3-1.1.3 Cell Culture
The chondrocytes were plated out to sterile 10cm tissue culture petri dishes (Cellstar, Greiner Bio-One, Longwood, Florida, United States) at 1×106 cells per dish as monolayer culture in the same DMEM mentioned above and maintained in a humidified incubator at 37°C, with 5% carbon dioxide [153]. Usually, the healthy chondrocytes would begin to adhere within the first two days, so in the 3rd day the culture medium was refreshed to discard the dead cells [154]. Moreover, often in the 7th day the cultured chondrocytes reached confluence and were detached by 0.05%
trypsin with EDTA•4Na (15400-054, Invitrogen Co.) for passage. Because most of the in vitro cultured chondrocytes lost their phenotype after the third passage [155], the cells used in the experiment were all in the beginning of the second passage.
![Figure 1 Cartilage covers each end of the bones of any joint [5].](https://thumb-ap.123doks.com/thumbv2/9libinfo/9608098.633739/17.892.255.658.562.847/figure-cartilage-covers-end-bones-joint.webp)