國立台灣大學生命科學院漁業科學研究所 博士論文
Institute of Fisheries Science College of Life Science National Taiwan University
Doctoral Dissertation
從耳石日成長輪及微化學探討西北太平洋區鱸鰻之生活史特性
LIFE HISTORY TRAITS OF THE GIANT MOTTLED EEL ANGUILLA MARMORATA IN THE NORTHWESTERN PACIFIC AS REVEALED FROM OTOLITH DAILY
GROWTH INCREMENT AND MICROCHEMISTRY
林德
Nico Jose Sarmiento Leander
指導教授:曾萬年 博士, 韓玉山 博士
Advisors: Wann-Nian Tzeng, Ph.D., Yu-San Han, Ph.D.
民國103年06月 June 2014
ii
iii
ACKNOWLEDGEMENT
This dissertation is the culmination of all the research works that I have done in the past years as a PhD student in the Fisheries Biology Laboratory, Institute of Fisheries Science, National Taiwan University and I would like to express my sincerest gratitude to the people who has been a part of this.
First and foremost, I am profoundly grateful to my doctoral supervisor, Prof. Wann-Nian Tzeng for accepting me to work in his lab and for his unwavering support and continuing guidance all these years. His profound knowledge and experience in fish biology and his untiring counsel inspired me to greater efforts in my research and encouraged my interest in academic work. Special thanks to the former students and staffs of Fisheries Biology Laboratory for their support and encouragement: Kang-ning Shen, Bruse Chang, Mei-Yu Chang, Arthur Hsu, Yu-Jia Lin, Yu-Ting Lin, Kuan-Yu Chen, Chih-Chen Cheng, Chun-I Chiang, Nian-Tzu Yeh, Dai-Ni Chi, Shih-Hung Yang, Yu-Hsiang Chen and Dai-Yu Lee for their invaluable help and encouragement. To Prof. Yu-San Han, my other doctoral supervisor, for his support and guidance and for providing me with a place in his lab to work during my remaining year in the Institute of Fisheries Science. Also, his students and staff in the Aquatic Molecular Biology lab were helpful and accommodating all throughout my stay in their lab. I am thankful to Dr.
Yoshiyuki Iizuka and his staffs at the Electron Probe Micro-analyses Lab, Institute of Earth Sciences, Academia Sinica for their assistance in the otolith microchemistry analyses.
Many thanks to the members of my PhD dissertation committee, Prof. Kwang-Tsao Shao (Academia Sinica), Prof. Kuo-Tien Lee (National Taiwan Ocean University), Prof. Chyng-Shyan Tzeng (National Tsing-Hua University), Prof. Chia-Hui Wang (National Taiwan Ocean
iv
University) and Prof. Hui-Yu Wang (National Taiwan University) for their valuable comments, insights and criticisms that were very helpful in my research.
Monthly fish collection for 4 years is a very big and daunting task so I am very grateful to Dr. Ruong-Tzung Chen, Dr. Ming-Fon Yeh, Prof. Chu-Fa Tsai and all the staff of Endemic Species Research Institute, Nantou, Taiwan for their assistance in fish sampling, loan of fish specimens from their extensive ichthyological collection and for putting up with my weird and sometimes difficult and complicated requests, etc.
A big thanks to my fellow students and friends here in Taiwan for the encouragement and the laughter: Bro. Tony, Ate Shelly, Kuya Toto, Ate Nanette, Ate Judy, Ate Fely, Cynthia, Doc.
Em and Doc. Anne. Special thanks to Brian, Rodolph, Faith Ate Juvia and Ate Malona for all the sleepless nights we spent together brainstorming about our researches and whatnot and for helping me with everything and for keeping me sane.
Lastly, I am eternally thankful to my family for the inspiration and for their patience, love and support all these years. I couldn’t have come this far without them.
v 摘要
為了瞭解西北太平洋熱帶性鱸鰻之加入動態和初期生活史以及菲律賓水域同種鰻的 洄游環境史。本研究調查2005-2009年台灣東部秀姑巒溪河口玻璃鰻的種類組成,結果發 現鱸鰻的玻璃鰻數量最多,占總捕獲量的98.4%;另外有少量的太平洋雙色鰻 (1.6%) 及日 本鰻(<1%)。熱帶性鱸鰻的玻璃鰻的主要出現季節為春夏天,但幾乎終年都可發現;而太 平洋雙色鰻的玻璃鰻的主要出現季節為秋天;溫帶性日本鰻的玻璃鰻的出現季節則是在冬 天。
透過東北亞地區不同河口所捕獲的鱸鰻和日本鰻玻璃鰻的耳石日周輪分析顯示,柳 葉鰻變態的日齡及早期成長率對於此兩種同域分布鰻魚的分離洄游及緯度分布扮演重要的 角色。成長快、變態早的鱸鰻柳葉鰻較早加入到菲律賓;而成長慢、延遲變態的日本鰻柳 葉鰻繼續向南(經由明答那峨海流)及向北(經由黑潮)飄送。另一方面,日本鰻柳葉鰻抵達 菲律賓水域時尚未到變態階段,所以也不會洄游到河口,而是繼續向北散佈。這可能就是 日本鰻很少在菲律賓發現而鱸鰻在菲律賓數量很多的原因。
電子微探儀(EPMA)分析顯示,採集於菲律賓呂宋島東部河川的鱸鰻耳石鍶鈣比數 據和之前所報導的日本及越南的鱸鰻以及日本鰻的洄游環境史非常不同。在耳石鰻線標記
之後到耳石邊緣的鍶鈣比都低於4 x 103,顯示菲律賓呂宋島東側的鱸鰻在鰻線階段進入
淡水溪流後一直到被捕獲的黃鰻階段為止都悽息在淡水環境中。此結果和台灣的鱸鰻一,
在黃鰻階段都只棲息在淡水中。而日本鰻在黃鰻階段的棲地利用則較為彈性,可在海水、
鹹淡水和淡水之間洄游。種間競爭、環境喜好性以及生產力可能是造成鱸鰻和日本鰻的棲 地喜好性不同的原因。本研究之發現可提供鰻魚資源保育之參考。
vi
關鍵字:耳石、日成長輪、鱸鰻、玻璃鰻、加入動態、洄游環境史
vii ABSTRACT
The present study investigated the recruitment dynamics and early life history of the tropical eel Anguilla marmorata in the northwestern Pacific as well as the migratory environmental history of the eel in the Philippine waters.
Analysis of the species composition of the recruiting glass eels in the estuary of the Hsiukuluan River, Eastern Taiwan from 2005-2009 revealed that A. marmorata was the most dominant eel species making up to 98.4% of the total catch while there were very few A. bicolor pacifica (1.6%) and A. japonica (<1%). Tropical eel species A. marmorata recruited mainly to the estuary during spring to summer but can be found year-round while A. bicolor pacifica recruited mainly during autumn. The temperate species, A. japonica, recruited mainly during winter.
Examinations of the otolith daily growth increments of A. marmorata and A. japonica glass eels collected from various rivers and estuaries in East Asia from 1992-2008 indicated that age at metamorphosis and early growth rate seem to play an important role in the segregative migration and latitudinal distribution of these two sympatric eel species in the northwestern Pacific. Faster-growing and earlier-metamorphosing leptocephali of A. marmorata recruited earlier in the Philippines while its slower-growing, delayed metamorphosing leptocephali dispersed southward (via the Mindanao Current) and northward (via the Kuroshio Current). On the other hand, the A. japonica leptocephali which arrive in the Philippine waters are apparently too young to metamorphose and migrate towards the estuaries so it will continue to drift northwards. This must be the reason why Japanese eels are seldom found in the Philippines while A. marmorata occurs in abundance.
The Sr:Ca profile in the otoliths of yellow-stage A. marmorata collected in the river of eastern Luzon, the Philippines in August 2008 revealed that it’s migratory environmental history
viii
is quite different from that previously reported from Japan and Vietnam and from A. japonica.
Electron probe microanalyzer showed that after the elver check, the Sr:Ca ratio until the otolith edge were less than 4 x 10-3, indicating that after recruitment, A. marmorata just stayed in freshwater until capture, which is similar to that of A. marmorata in Taiwan. On the contrary, A.
japonica has a more flexible migratory behavior in the yellow stage. It can migrate among seawater, brackish water and freshwater in the yellow eel stage. Interspecific competition, environmental factors and the productivity of the environment may play an important role in the habitat preference of A. marmorata throughout its species range. The findings of this study can provide the information for the eel conservation and management.
Keywords: Otolith, daily growth increment, giant mottled eel, glass eels, recruitment dynamics, migratory environmental history
ix
TABLE OF CONTENTS
Title page……….i
Approval sheet………...ii
Acknowledgement……….iii
Abstract………...v
Table of Contents………..ix
List of Tables………...xiii
List of Figures………...xv
List of Appendices………xxiii
CHAPTER 1: INTRODUCTION 1.1 Life history pattern of freshwater eel (Anguilla spp.)………1
1.2 Freshwater eel fishery and its problem………..4
1.3 Population decline of freshwater eel………..4
1.4 Biology and population structure of Anguilla marmorata………...10
1.5 Application of otolith daily growth increment and microchemistry to life history study……….14
1.6 Current research status of Anguilla marmorata………...17
1.7 Objectives of this study………19
CHAPTER 2: MATERIAL AND METHODS 2.1 Glass eel collection 2.1.1 Temporal sampling………...20
2.1.2 Spatial sampling………24
2.2 Juvenile eel collection………..27
2.3 Glass eel species identification and morphometric measurements………..29
x
2.4 DNA extraction, polymerase chain reaction (PCR) amplification and phylogenetic
analysis………..30
2.5 Determination of glass eel developmental stages………33
2.6 Morphometric measurements of adult eels………..37
2.7 Otolith extraction and preparation for daily growth increment and microchemistry analyses………...39
2.8 Life history parameters obtained from otolith daily growth increment analyses……40
CHAPTER 3: RESULTS 3.1 Species identification for the glass eel recruiting to Hsiukuluan River, Eastern Taiwan ………...44
3.2 Species composition of the glass eel recruited to Hsiukuluan River………...49
3.3 Stage composition of the glass eel recruited to Hsiukuluan River ………...52
3.4 Comparison of morphometric characters of glass eels among species………....55
3.5 Daily growth increments and growth checks in the otoliths of glass eel of Anguilla marmorata ………...62
3.6 Comparison of the developmental stage composition between Anguilla marmorata and A. japonica glass eels………....72
3.7 Differences in size and age at estuarine arrival between Anguilla marmorata and A. japonica glass eels………76
3.8 The timing of metamorphosis from leptocephalus to glass eel as indicated by otolith daily growth rates and Sr:Ca ratios………...79
3.9 Back-calculated hatching dates of Anguilla marmorata and A. japonica…………...85 3.10 Age at metamorphosis in relation to the growth rate and distance from the spawning
xi
grounds………..87 3.11 Migratory environmental history and habitat use of Anguilla marmorata juveniles in the Philippines as indicated by otolith Sr:Ca ratios………...95 CHAPTER 4: DISCUSSION
4.1 The differences in spatial recruitment patterns among species of glass eel in
Taiwan …………...102 4.2 Differences in seasonal occurrence between Anguilla marmorata and A. japonica in Taiwan………106 4.3 The metamorphosis timing differs between Anguilla marmorata and A. japonica and its ecological evolution significance………...107
4.4 Delayed metamorphosis as a means of long-distance dispersal of the eel…………110 4.5 The early growth rate affects dispersal range of leptocephali ………..112 4.6 Spatial and temporal population genetic structure of the giant mottled eel Anguilla marmorata in the northwestern Pacific………..113 4.7 Migration behavior and habitat use of Anguilla marmorata in eastern Luzon,
Philippines………...115 CHAPTER 5: CONCLUSION AND PERSPECTIVE
5.1 Species-specific geographic distribution………...118
5.2 Role of larval growth rate and metamorphosis timing in determining the geographical distribution of the anguillid eel ………..118
5.3 Migratory environmental history of Anguilla marmorata is different from that of A.
japonica ………..119 REFERENCES………...121
xii
APPENDICES………140
xiii
LIST OF TABLES
TABLE 1. Sampling information and sample sizes of the anguillid glass eel collected in the lower reach of Hsiukuluan River, eastern Taiwan……….23
TABLE 2. Sampling information and sample sizes of A. japonica and A. marmorata glass eel collection in various rivers and estuaries in East Asia. Values inside the parentheses indicate the number of individuals used for otolith analyses………26
TABLE 3. Development of pigmentation in anguillid eels (Tesch2003)……….36
TABLE 4. Species composition of Anguilla glass eels collected in the lower reach of Hsiukuluan River in Eastern Taiwan from 2005-2009……….50
TABLE 5. Pigmentation stages of the different Anguilla eel species collected from 2 stations in the lower reach of Hsiukuluan River……….53
TABLE 6. Comparison of morphometric characters among the 4 Anguilla species………59
TABLE 7. Sampling sites, sample sizes, sampling dates, total lengths, daily age of glass eels at the estuary (Tt), daily age at metamorphosis from leptocephalus to glass eel (Tm), and the time between the metamorphosis check and estuarine arrival (Tt-m) of Anguilla japonica and A.
marmorata……….73
xiv
TABLE 8. Pigmentation stages of glass eels of Anguilla japonica and A. marmorata collected from various rivers and estuaries in East Asia………...75
TABLE 9. Mean (± SD) increment widths of otoliths radii Rm, Rt, and Rt-m in of Anguilla japonica and A. marmorata glass eels.………..80
TABLE 10. Age and estimated hatch date of glass eels of Anguilla japonica and A. marmorata recruiting in East Asia………86
TABLE 11. Sampling information of the adult eels collected from a river system in Aurora province, eastern Luzon, Philippines on August 6, 2012. ………95
TABLE 12. Sampling information of the adult eels collected from an aquaculture farm in Sanya, southern Hainan province, China………...96
xv
LIST OF FIGURES
FIGURE 1. Life cycle and catadromous migration of anguillid eels……….3
FIGURE 2. Recruitment (3 year running averages of geomeans of indices as % of 1979-1994 means) for the European (Moses-Saunders Index – 7 years), American (adjusted to year of arrival) and Japanese glass eels from Taiwan………..5
FIGURE 3. Population decline in Anguilla japonica (a; Tsukamoto et al. 2009) and A.
marmorata (b; Prof. C.S. Tzeng, unpublished)………..8
FIGURE 4. Migration cycle of the northwestern Pacific population of Anguilla marmorata….12
FIGURE 5. Map showing the species range (thick black lines along the coasts) of Anguilla marmorata (a) and A. japonica (b) in East Asia and the geographic distribution of their larvae (data from Kuroki et al. 2006 and Tsukamoto 2009)………13
FIGURE 6. The orientation of the semicircular canals and otoliths within the inner ear of teleost fish (a) and the transverse section of the inner ear showing the positions of each otoliths (b) (image modified from Panfili et al. 2002)……….16
FIGURE 7. Map showing glass eel sampling stations (1 and 2) in the lower reach of the Hsiukuluan River in eastern Taiwan………..21
xvi
FIGURE 8. Fish way trap used to collect the migrating fish larvae in the lower reach of Hsiukuluan River (a) and the dimension of the tin trap (b)………...22
FIGURE 9. Map showing the geographic distributions of Anguilla japonica (thick gray lines on the coastlines) and A. marmorata (thick black lines on the coastlines) in East Asia and collection sites of samples analyzed in this study (∆, A. marmorata; Ο, A. japonica ; Cheng and Tzeng 1996)………..25
FIGURE 10. Sampling location of the juvenile eels in a river system in Aurora province in eastern Luzon, Philippines……….27
FIGURE 11. Electro-fishing gear (a) used to collect juvenile eels in a river system (b) in Aurora province, eastern Luzon, Philippines……….28
FIGURE 12. Schematic diagram of the methods used for anguillid glass eel species identification………..32
FIGURE 13. Diagram showing morphometric measurements of the glass eel………33
FIGURE 14a. Progressive sub-epidermal pigmentation in the anal region of the glass eel used to determine the different ontogenetic stages (after Strubberg, 1913; Bertin 1956; Tesch 2003)….34
xvii
FIGURE 14b. Developmental stages (VIA1 to VIB) in Japanese eels showing pigmentation process from glass eel to the fully pigmented elver stage at VIB. Arrows indicates the location of the unique pigmentation characteristic of each stage (adapted from Fukuda et al.
2013)………..35
FIGURE 15. Diagram showing morphometric measurements of the adult eel………38
FIGURE 16. Schematic diagram of the measurements of radii and counts of the daily growth increments in the otoliths of the elvers (modified from Cheng and Tzeng 1996; Wang and Tzeng 2000)………..43
FIGURE 17. Origins of the dorsal fin (black triangle) and anal fin (gray triangle) in longfin (a, A. japonica and b, A. marmorata, A. celebesensis and A. luzonensis) and shortfin (c, A. bicolor pacifica) glass eels………46
FIGURE 18. Caudal in and tail bud pigmentation patterns of anguillid glass eels………..47
FIGURE 19. Phylogenetic tree of the genus Anguilla inferred from cytochrome b sequences...48
FIGURE 20. Species composition of the recruiting glass eels collected in 2 stations at the lower reach of Hsiukuluan River, Eastern Taiwan from 2005-2009………...51
xviii
FIGURE 21. Developmental stage composition of recruiting glass eels according to sampling station……….54 FIGURE 22. Length frequency distribution of glass eels of Anguilla japonica (a), A. bicolor pacifica (b), and A. marmorata (c) caught in the lower reach of Hsiukuluan River of Eastern Taiwan and A. celebesensis (d) (Tzeng 1982)………56
FIGURE 23. Relationship between pre-dorsal fin length and total length in the glass eels of Anguilla japonica, A. bicolor pacifica, A. marmorata and A. celebesensis (from Tzeng 1982)………..60
FIGURE 24. Relationship between ano-dorsal fin length and total length in the glass eels of Anguilla japonica, A. bicolor pacifica, A. marmorata and A. celebesensis (from Tzeng 1982)...61
FIGURE 25. SEM photographs showing daily growth increments (DGIs) and growth checks in an otolith of an Anguilla marmorata elver………64
FIGURE 25a. Peripheral region of the Anguilla marmorata glass eel otolith showing the estuarine check (white arrow) and the otolith edge (black arrow)……….65
FIGURE 25b. SEM photograph showing the metamorphosis check (black arrow) in the Anguilla marmorata glass eel otolith………...66
xix
FIGURE 25c. SEM photograph showing the discontinuous (dark band, black arrow) and increment (light band, white arrow) zones in the Anguilla marmorata glass eel otolith………...67
FIGURE 25d. Core region of the of the Anguilla marmorata glass eel otolith showing the primordium (gray arrow), yolk-sac stage (red line), hatching check (white arrow) and the 1st feeding check (black arrow)………...68
FIGURE 26a. Core region of the A. marmorata glass eel otolith showing a distinct and well- defined primordium………...69
FIGURE 26b. Core region of the A. marmorata glass eel otolith showing an undefined primordium………70
FIGURE 26c. Core region of the A. marmorata glass eel otolith showing twin primordia………71
FIGURE 27. Developmental stage composition of recruiting Anguilla marmorata (a) and A.
japonica glass eels……….74
FIGURE 28. Length-frequency distribution of recruiting Anguilla marmorata glass eels from the Philippines, Taiwan, China and Japan……….77
xx
FIGURE 29. Length-frequency distribution of recruiting Anguilla japonica glass eels from the Taiwan, China and Japan………...78
FIGURE 30. Profiles of otolith growth increment width (blank circle) and Sr:Ca concentration ratios measured using electron probe microanalyzer (solid diamond) from the primordium to the edge in A. marmorata glass eels………81
FIGURE 31. Otolith Sr:Ca profiles of the A. marmorata glass eels collected from the Philippines……….82
FIGURE 32. Otolith Sr:Ca profiles of the A. marmorata glass eels collected from Taiwan…...83
FIGURE 33. Otolith Sr:Ca profiles of the A. marmorata glass eels collected from Japan …...84
FIGURE 34. Spatial changes in age at metamorphosis in A. marmorata (a) and A. japonica (b)...88
FIGURE 35. Spatial changes in age at recruitment in A. marmorata (a) and A. japonica (b)………...89
FIGURE 36. Spatial changes in length at recruitment in A. marmorata (a) and A. japonica (b)………...90
xxi
FIGURE 37. Spatial changes in time between metamorphosis and age at recruitment in A.
marmorata (a) and A. japonica (b)………91
FIGURE 38. Relationship between the age at metamorphosis (Tm) and growth rate (Gm) before metamorphosis in Anguilla marmorata……….92
FIGURE 39. Relationship between the age at metamorphosis (Tm) and growth rate (Gm) before metamorphosis in Anguilla japonica……….93
FIGURE 40. Relationship between the larval dispersal distance and age at metamorphosis (Tm) in Anguilla marmorata (open symbols) and A. japonica (solid symbols)……….94
FIGURE 41. An otolith sample of a wild Anguilla marmorata (553.86 mm TL) from Aurora province, Philippines………..97
FIGURE 42. An otolith sample of a cultured Anguilla marmorata (785 mm TL) from Hainan province, China………..98
FIGURE 43. Temporal changes in the otolith Sr:Ca ratios (measured from the core to the edge) of A. marmorata collected from the Philippines………..100
FIGURE 44. Temporal changes in the otolith Sr:Ca ratios (measured from the core to the edge) of A. marmorata collected from an aquaculture farm in Sanya, southern Hainan Province,
xxii
China………101
FIGURE 45. Distribution patterns and abundances of A. marmorata and other anguillid eels in Taiwan………..105
FIGURE 46. Anguillid eel metamorphosis from leptocephalus to glass eel and some of the accompanying physiological changes………..109
xxiii
LIST OF APPENDICES
APPENDIX 1. Fisheries Administrative Order 242 of the Philippines………..140
APPENDIX 2. Papers, abstracts and presentations related to this dissertation………..142
1
1. INTRODUCTION 1.1 Life history pattern of freshwater eel (Anguilla spp.)
Anguillid eels (Genus Anguilla) are catadromous fishes, spending the majority of their life cycle in freshwater or estuaries until sexual maturity and migrating to the open ocean to spawn and die (Fig. 1). These long migrations to spawn far offshore appeared to be unique among the >800 anguilliform species (Miller 2009). At the moment, only the spawning grounds of four freshwater eel species in both the Atlantic and the Pacific Oceans have been identified (Schmidt 1922; Tsukamoto 1992; Kuroki et al. 2009). The spawning areas of the other freshwater eel species around the world are still not well understood, thus research interest in eel migration is still focused on the actual spawning location for anguillid eels. Interestingly, these eels in both the Atlantic and the Pacific spawn in similar westward flowing currents at the southern edges of the subtropical gyres in both oceans so that their leptocephali can be passively transported to their respective recruitment areas where they feed and grow (Tsukamoto et al.
2002). At the end of their long transoceanic migration, the leptocephali metamorphose into glass eels and invade coastal and inland habitats. The eels undergo two major metamorphoses throughout their long lives: the first one is the metamorphosis from leptocephalus into glass eel during their migration from their offshore marine spawning ground to their continental freshwater growth habitat. Otolith microchemistry studies have revealed that the glass eels after reaching coastal waters may either migrate further inland and colonize freshwater habitats or stop their migration and settle in seawater or estuary (Tzeng et al. 2002; Arai et al. 2004; Daverat et al. 2006). The timing of the metamorphosis by a leptocephalus into glass eel and the transportation of the oceanic currents are considered to be the key determinant of the ultimate destination of the eel (Cheng and Tzeng 1996; Wang and Tzeng 2000; Tzeng 2003). The second metamorphosis on the other hand, is from yellow eel to silver eel in a process called silvering
2
and it occurs during their downstream migration from their continental freshwater habitat to their offshore spawning ground in the open ocean. During silvering, their body colouration changes, their swim bladder changes and their eyes enlarge, all of which would facilitate the new demands of deep-water swimming, predator avoidance and possibly visual mate location (Rousseau et al.
2002; Aoyama and Miller 2003). These series of metamorphoses in yellow eels in both freshwater and coastal marine habitats are accompanied by drastic morphological, physiological and behavioural changes to fit their habitat shift and activities associated with a long migration in the open ocean (Bruijs and Durif 2002). Recent study by Tsukamoto et al. (2011) reported that the morphological condition of spawning adults suggests that freshwater eels have the capacity for multiple spawning during a spawning season. However, their highly modified bodies and degenerated condition confirm that they have only one spawning season in their life and die after spawning a few times.
3 Fig. 1. Life cycle and catadromous migration of anguillid eels.
4 1.2 Freshwater eel fishery and its problem
Anguillid eels are an economically important fish species in many regions of the world particularly in Europe and East Asia where there is a long standing tradition of its consumption.
The Japanese consume around 100, 000 tons of eels every year, of which more than 99.5% are farmed eels (FAO 2012 Statistics). Intensive commercial culture of anguillid eels (particularly A.
japonica) is an important industry not only in Japan where it was first initiated but also in China, Taiwan and Korea. In fact in recent years, imports of farmed eels from these two countries have grown considerably that they now comprise around 70% of all eels consumed in Japan. The seeds needed for aquaculture production still rely on wild-caught glass eels but because the glass eel resources are in sharp decline globally, the eel farming industry is at a critical point. To ease the reliance for wild-caught eels for aquaculture production and to help protect the wild populations, researches on technology for producing artificial glass eels were initiated. But at the moment, problems such as poor egg quality, appropriate food for the larvae and high density rearing method for leptocephalus among other things, have hindered the completion of its artificial production.
1.3 Population decline of freshwater eel
The world’s eel resources have been in sharp decline since the second half of the last century (Fig. 2). In the Pacific Ocean, the annual catch of the Japanese eels is reported to be fluctuating with a continuously decreasing trend (Tatsukawa 2003). Its decline started in around 1970 and it now stand at about 10% of the levels seen in the 1960’s. This decreasing trend however is not only observed in/limited to the Japanese eel. In the Atlantic Ocean, European and American eel recruitment is down to less than 1% of its peak in some areas (Richkus and Whalen 2009; Dekker 2008). South Pacific eels are also reported to be in decline (Jellyman et al. 2000;
5
Fig. 2. Recruitment (3 year running averages of geomeans of indices as % of 1979-1994 means) for the European (Moses-Saunders Index – 7 years), American (adjusted to year of arrival) and Japanese glass eels from Taiwan.
6
Hoyle and Jellyman 2002). The reasons for the decline are unclear but are probably due to spawning stocks reduction (Clevestam et al. 2011), swim bladder parasites and virus infection (Haenen et al. 2002; Szekely et al. 2002; Kirk 2003; Sures and Knopf 2004; van Ginniken et al.
2005) , overfishing (Moriarty and Dekker 1997; Briand et al. 2003; Dekker 2003), growth habitat reduction (McCleave 2001), pollution (Amiard-Triquet et al. 1988; Robinet and Feunteun 2002;
van Ginneken et al. 2009), global climate change (Castonguay et al. 1994; Kimura et al. 2001;
Knights 2003; Friedland et al. 2007; Bonhommeau et al. 2008) and solar cycle (Tzeng et al.
2012). Previous researches also highlighted the environmental factors in the estuaries such as temperature, salinity, turbidity, pH, stream water odour and tidal cycle as well as moon phase and rainfall, may act alone or in combination to influence recruitment (Sloane 1984; Tzeng 1985;
Sorensen and Bianchini 1986; Tosi et al. 1990; Chen Lee et al. 1994).
In the northwestern Pacific, intense fishing pressure in adult A. japonica and A.
marmorata and the widespread harvesting of their glass eels for aquaculture caused their population to decline (Fig. 3). Also, constructions along the river systems like dams and other water impoundments impeded the downstream migration of the adult eels towards the sea and the upstream migration of elvers, further affecting the dwindling eel population. It is because of this that the Taiwanese government was prompted to impose fishing and aquaculture ban on A.
marmorata and listed it as an endangered species according to the Wildlife Conservation Act of Taiwan. More recently, the Philippine government, through its Bureau of Fisheries and Aquatic Resources (BFAR) issued Fisheries Administrative Order (FAO) No. 242 series of 2012 repealing FAO 159 series of 1986 that allows elver exportation and reinstate the ban on the exportation of live elvers caught in the Philippine waters in a bid to stop the rapid and rampant
7
exploitations of the anguillid eels in the country (Appendix I). According to BFAR regional office in northern
8
Fig. 3. Population decline in Anguilla japonica (a; Tsukamoto et al. 2009) and A. marmorata (b;
Prof. C.S. Tzeng unpublished ).
9
Philippines, approximately more than 1 million elvers were caught every day during the main fishing season (November to February every year) and from PhP 2,500 (US$ 59.19 or NT$
1,766.09) per kilo (approx. 5-6 thousand elvers) in 2012, the buying price suddenly went up to 17 thousand (US$ 402.31or NT$ 12,009.43) to 28 thousand (US$ 622.58 or US$ 19,783.14) PhP from January to March 2013. Because of the high demand and price for the elvers, indiscriminate and undocumented elver collections were reported in northern Philippines specifically near the river mouth of Cagayan River. The FAO carries a penalty of eight years imprisonment, confiscation of catch or a fine equivalent to double the export value of the same, and revocation of fishing and/or export permit. In recent years, A. marmorata is rapidly gaining popularity as an aquaculture species in East Asia due to the declining supply of A. japonica and the soaring prices of its glass eels. Anguilla marmorata glass eels are still in abundant supply and its price is much cheaper than those of A. japonica glass eels. Despite the ban the Philippines is still supplying increasingly large quantities of A. marmorata glass eels caught in major river systems in northern Luzon and in Mindanao, to numerous eel farms in East Asia and are now increasingly being used in many eel farms in China, Taiwan and Korea. Early this year, the International Union on the Conservation of Nature listed A. japonica as an endangered species due to a number of factors that includes overfishing and loss of habitat (IUCN, 2014). Because of this, it is expected that A. marmorata is poised to take-over A. japonica as the major eel species for aquaculture in East Asia. But to prevent A. marmorata from following the same fate as A.
japonica, understanding its life history will help in formulating regulatory measures for its conservation and sustainable use.
10
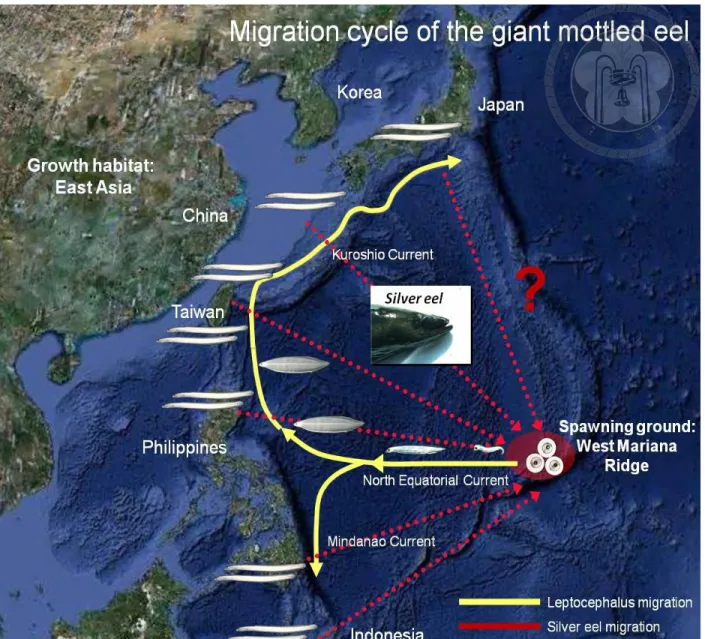
1.4 Biology and population structure of Anguilla marmorata
The tropical anguillid eel Anguilla marmorata is the most widespread in all of the 16 species and 3 subspecies of freshwater eels in the world (Ege 1939; Watanabe 2003). It is found from the southeast coast of Africa in the Indian Ocean, eastward through the islands like Madagascar and Reunion, northward through Indonesia up to southern Japan and Korea and through the tropical western Pacific, including many small islands in the South Pacific (Tesch 2003). More recently, the occurrence of A. marmorata outside of its known distribution range has been reported like in Caroline Islands (Myers and Donaldson 2003; Donovan et al. 2012), Palmyra Atoll (Handler and James 2006) and even farther east in the Galapagos (McCosker et al. 2003). It has been widely documented that this species can live in sympatry with the other tropical and temperate eel species. Recent genetic (mtDNA and microsatellite) and morphological (total number of vertebrae) studies suggested that A. marmorata has at least four different regional populations in the whole Indo-Pacific (Ishikawa 1998; Ishikawa et al. 2004; Minegishi et al. 2008; Watanabe et al. 2008; Watanabe et al. 2009a). Such type of regional divergence within the same species can be due to each population having different migration loops or migratory pathways brought about by barriers to migration in the Pacific and Indian Oceans. This causes each population to evolve to have unique or specific migration loops adapted to the geography and hydrology of each area.
Therefore, the shift in the migration loop could possibly a step towards speciation in freshwater eels. At the moment with its four to five different regional populations, only one spawning ground of A. marmorata has been identified and it is in the north equatorial current (NEC) region of the western North Pacific Ocean in about the same region as the spawning area of A. japonica (Kuroki et al. 2009; Tsukamoto et al. 2011). Both A. japonica and A. marmorata were proposed to be spawned in the waters west of the Mariana Islands in the Pacific Ocean (Fig. 4) (Kuroki et
11
al. 2009; Tsukamoto et al. 2011). Adult individuals migrate thousands of miles from their freshwater growth habitat in East Asia to this location under starving condition because they do not assimilate marine food sources during their spawning migration (Chow et al. 2010). After hatching, their marine larvae, the leptocephalus were transported westward by the NEC from their spawning ground to the continental shelf of the northwestern Pacific. Furthermore, A.
marmorata will drift both in the northward flowing Kuroshio Current and the southward flowing Mindanao Current (Fig. 5a) while A. japonica will only enter the northward flowing Kuroshio Current that will transport them to East Asia particularly in Taiwan, China, Japan and Korea (Fig.
5b). This indicates that the larvae of A. marmorata disperses and recruits to a wider area in the northwestern Pacific region than A. japonica (Tsukamoto 1992; Kimura et al. 1994; Miller et al.
2006; Kuroki et al. 2006). Why A. japonica larvae only enter the Kuroshio Current Region while A. marmorata, after being transported by the NEC, can enter both the northward Kuroshio Current and the southward Mindanao Current that carry them to their recruitment areas in northern Indonesia, the Philippines, Taiwan, eastern China, southern Japan and Korea, is still unclear (Zenimoto et al. 2009). Recently, Han et al. (2012) pointed out that recruitment temperature preference and oceanic current system control the distinct biogeography of A.
japonica and A. marmorata. However aside from these environmental controls, biological factors should also be taken into consideration in explaining this ecological discrepancy but the early life histories of these two species particularly during the oceanic phase are not yet fully understood especially that of A. marmorata. Upon reaching the continental waters, it was also found that their distribution in the estuaries was also geographically different. In addition, it was also observed that in the same river system, A. marmorata occupied the upper reaches while A.
japonica occupied the lower reaches (Shiao et al. 2003).
12
Fig 4. Migration cycle of the northwestern Pacific population of Anguilla marmorata.
13
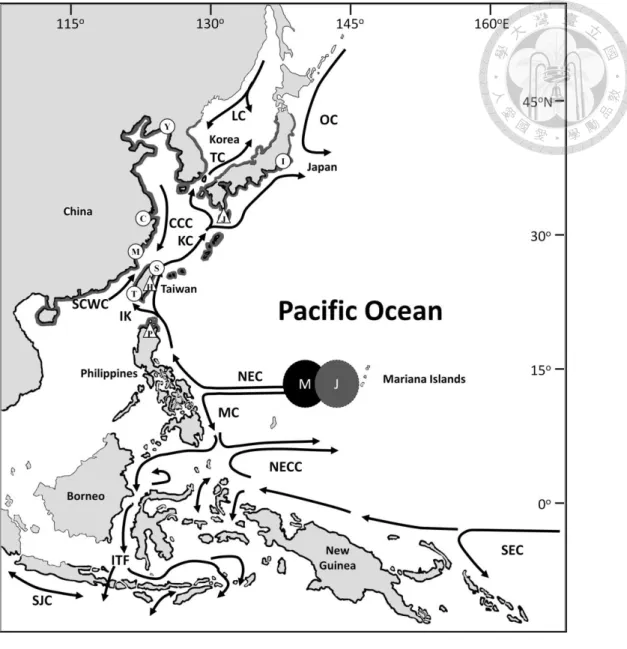
Figure 5. Map showing the species range (thick black lines along the coasts) of Anguilla marmorata (a) and A. japonica (b) in East Asia and the geographic distribution of their larvae (data from Liao et al. 1999, Kuroki et al. 2006 and Tsukamoto 2009).
Preleptocephali (≤ 7 mm TL) (), small leptocephali (≤ 10 mm TL) (♦), leptocephali (>10 mm TL) (●), metamorphosing leptocephali (▲) and oceanic glass eels (■). The detailed patterns of the general currents in the western North Pacific and central Indonesian Seas were also shown (adapted from Nitani 1972 and Lukas et al. 1991). NEC: North Equatorial Current, KC: Kuroshio Current, OC:
Oyashio Current, TS: Tsushima Current, CCC: China Coastal Current, SCSWC: South China Sea Warm Current, IK: Intruded Kuroshio, MC: Mindanao Current, NECC: North Equatorial Counter Current.
14
1.5 Application of otolith daily growth increment and microchemistry to life history study For many years, the life history events of anguillid eels have been the focus of a variety of researches. However such information about the early life histories of the anguillid eels have been slow to accumulate primarily because of the difficulty in collecting and keeping them alive due to their size and fragile body structure. Because of these constraints, otolith microstructure analysis has emerged as a useful and powerful technique in elucidating the early life history of the anguillid eels. Otoliths are small, paired calcified structures that aids in hearing and in the maintenance of equilibrium of all teleost fishes and are located in the fluid-filled chambers of the inner ear (Fig. 6). Fish otoliths are composed of calcium carbonate and an organic matrix and are both deposited in daily and annual increments as the fish grows, allowing the ages of the fishes to be determined on a daily and annual basis (Panella 1980). Recent chemical analytical techniques have enabled the identification of the life-history events in a variety of fish species by detecting major (>100 ppm), minor (1 ≤ ppm <100) and trace (<1 ppm) elements in the microstructures of their otoliths (Campana 1999; Arai 2002). This is because the depositions of the various elements in the growth increments of the otolith represent a permanent record of the environmental conditions experienced by the fish at a particular time (Campana et al. 2000). As a result of these advances, the growth increments in the otoliths have been widely used to study the different life history events in various eel species. It has been proven that the examination of the otolith microstructures and microchemistry is a useful tool in estimating various events in the life of the eel such as spawning and hatching dates, growth rate and life stage and habitat transitions.
Recent progress in otolith analytical techniques have revealed considerable details of the early life history of various anguillid eels species including information on age, growth patterns, timing of metamorphosis of leptocephali, larval duration, recruitment to coastal waters, timing of
15
inshore migration of glass eels and others (Arai et al. 1997, 1999a,b,c, 2001a,b; Wang and Tzeng 1998; Ishikawa et al. 2001; Marui et al. 2001; Kuroki et al. 2008; Shiao et al. 2002). It was found that the growth increments in the otoliths of the newly hatched larvae and elvers of both temperate and tropical eel species are deposited on a daily basis thus it is possible to determine their various early life history details such as hatching date, age at first feeding, duration of leptocephalus stage, age at metamorphosis, oceanic glass eel stage and age at recruitment (Tsukamoto 1989; Martin 1995; Arai et al. 2000a; Cieri and McCleave 2001; Sugeha et al. 2001, Kuroki et al. 2006; Budimawan and Lecomte-Finiger 2007; Reveillac et al. 2009; Leander et al.
2013). It is also possible to calculate the growth rate by measuring the otolith growth increments.
In addition, it was confirmed by various studies that there is a relationship between otolith microstructure and microchemistry (Otake et al. 1994; Arai et al. 1997; Arai et al. 2000b). A marked increase in otolith increment width coincides with a drop in otolith Sr: Ca ratios indicates the onset of metamorphosis, a common phenomenon in all anguillid eels (Otake et al. 1994; Arai et al. 1999a,b,c; Marui et al. 2001). Otake et al. (1994) suggested that this marked drop of otolith Sr: Ca ratios is related to the internal physiological changes that occurs during metamorphosis.
Furthermore, Arai et al. (1997) reported that metamorphosis was completed before the increment width reached the maximum peak. These changes in otolith microstructure and microchemistry allowed us to have an insight on the timing and duration of the metamorphic period of the different anguillid eel species.
16
Fig. 6. The orientation of the semicircular canals and otoliths within the inner ear of teleost fish (a) and the transverse section of the inner ear showing the positions of each otoliths (b) (image modified from Panfili et al. 2002).
17 1.6 Current research status of Anguilla marmorata
Several aspects of the life history and ecology of the elvers of the temperate eel A.
japonica in the estuaries around Taiwan has been well documented, including reports on metamorphosis and estuarine arrival (Tzeng 1990; Cheng and Tzeng 1996), timing of estuarine recruitment in relation to environmental conditions (Tzeng 1984a, 1985; Chen et al. 1994), fishing exploitation rates (Tzeng 1984b), effects of surface seawater temperature on the recruitment of elvers (Tzeng 2006), genetic composition of the recruiting glass eels (Chang et al.
2007) and the possible effects of environmental variations such as sunspot cycles and El Nino events on the glass eel catch (Han et al. 2009) and others. However, little is known concerning the tropical eel species on the eastern coast of Taiwan as previous studies were focused on the A.
japonica populations on the southern, western and northern coast of Taiwan where their elvers were regularly collected for eel aquaculture. Accordingly, as compared with the information on A. japonica, relatively little is known about the life history of A. marmorata. It also did not help that A. marmorata was declared as an endangered species in Taiwan. Because of this ban, little information has been known about its population in Taiwan because its fishing and aquaculture is illegal before April 2009.
Not only its life history during the continental growth phase but also its marine larval life stage from the spawning ground to the estuary is still not well understood for A. marmorata as compared with that of the temperate eel species (McCleave 1993; Cheng and Tzeng 1996; Wang and Tzeng 2000; Tzeng 2003; Edeline et al. 2009; Miller 2009). Although recent research efforts have resulted in a basic understanding of the ecology of the anguillid eels, early life history details such as metamorphosis from leptocephalus to glass eel stage as well as from glass eel to elvers stage remains to be understood. Similarly, in the Philippines, the first country the drifting
18
leptocephalus encountered as they migrate and colonize continental East Asia, information such as population structure, recruitment dynamics, species composition and abundance, etc. is still lacking. Such information is essential for the sustainable fishery and resource conservation of the tropical anguillid eel populations. It is because of this need for information regarding the life history of A. marmorata that this present study was conceived.
Also, to understand the decline in glass eel recruitment, it is necessary to study the early life history of anguillid eels. The life history of eels during the continental growth phase is well documented, but knowledge of the marine larval life stage from the spawning ground to the estuary is still fragmented (McCleave 1993, Cheng and Tzeng 1996, Wang and Tzeng 2000, Tzeng 2003, Edeline et al. 2009, Miller 2009). Early life-history information is very important because it is a key factor in understanding possible reasons for recruitment success or failure of anguillid eels and also for their artificial propagation.
19 1.7 Objectives of this study
The present study aims to examine the life history of the tropical eel A. marmorata in northwestern Pacific from marine larval phase up to continental growth phase by evaluating its recruitment pattern in the estuary and by analyzing its otolith microchemistry and microstructures. In addition, its life history and evolution in the northwestern Pacific were compared with that of A. japonica. Specifically this study aimed to:
Examine the early oceanic growth phase of A. marmorata using information such as the
timing of metamorphosis from leptocephalus to glass eel, inshore migration period of the glass eel, age and size at estuarine arrival and the growth rate of the leptocephalus and elver.
Determine the possible mechanism for the segregative migration and recruitment of A.
marmorata and A. japonica in the northwestern Pacific.
Assess the recruitment pattern of A. marmorata and other anguillid eel species in
Hsiukuluan River, Eastern Taiwan from 2005-2009 to clarify their seasonality and abundance.
Determine the migratory environmental history and habitat use of adult A. marmorata in eastern Luzon, Philippines.
20
2. MATERIALS AND METHODS 2.1 Glass eel collection
2.1.1 Temporal sampling
To determine the species composition, seasonal occurrence and recruitment pattern of anguillid eel in eastern Taiwan, glass eel were sampled monthly in 2005 and 2007-2009 at 2 stations in the lower reach of the Hsiukuluan River (Fig. 7). Hsiukuluan River is the largest river in eastern Taiwan with a length of 81 km and a drainage area of 1790 km2 (Shiao et al. 2003).
The 1st station was located in the river mouth, and a traditional triangle net was used for glass eel collection; the 2nd station was located 2 km upstream where a fish way trap was set up in an artificially dug water channel that was about 20 m long, 1.5 m wide, and 25 cm deep (Fig. 8).
After collection, specimens were immediately preserved in 75% ethanol. Environmental parameters such as dissolved oxygen (DO), salinity (conductivity), pH, water temperate, water velocity, and turbidity were also recorded at every sample collection.
21
Fig 7. Map showing glass eel sampling stations (1 and 2) in the lower reach of the Hsiukuluan River in eastern Taiwan. The bars indicate the Juisiu and Long Rainbow Bridges.
22
Fig. 8. Fish way trap used to collect the migrating fish larvae in the lower reach of Hsiukuluan River (a) and the dimension of the tin trap (b). Arrows represents the direction of the river flow. Figure provided by Dr. R.T. Chen.
23
Table 1. Sampling information and sample sizes of the anguillid glass eel collected in the lower reach of Hsiukuluan River, eastern Taiwan.
Stn. 1 Stn. 2 Total
2005 June 0 151 151
2007 July 7 19 26
Sept. 0 8 8
2008 Apr. 0 1 1
May 0 48 48
June 31 23 54
July 0 13 13
Aug. 56 0 56
Sept. 4 1 5
Oct. 36 0 36
Nov. 280 0 280
Dec. 62 0 62
2009 Jan. 1 0 1
Feb. 1 0 1
Mar. 42 2 44
Apr. 64 0 64
May 17 0 17
June 61 0 61
July 69 0 69
Aug. 4 0 4
Sept. 1 2 3
736 268 1004
Sample size Sampling period
Total
24 2.1.2 Spatial sampling
A total of 168 A. marmorata glass eels were collected from the estuaries of the Hsiukuluan River, eastern Taiwan (n = 86) on 20 May 2008, the Cagayan River, northern Philippines (n = 45) on 19 May 2008, and the Kurio River, southern Japan (n = 37) on 6 June 1996 (Fig. 9) to determine the possible effects of early life history traits on its latitudinal distribution in the northwestern Pacific together with A. japonica. All specimens were immediately preserved in 95% ethanol after measuring the total lengths (TLs) to the nearest millimeter (mm). On the other hand, A. japonica specimens examined from a previous study (Cheng and Tzeng 1996) were collected from estuaries of the Tungkang River, southern Taiwan (n = 60) on 30 December 1992 and 24 March 1993, the Shuangshi River, northern Taiwan (n = 60) on 20 December 1992 and 17 February 1993, the Mingchiang River, eastern China (n = 30) on 1 March 1993, the Chyantarng River, eastern China (n = 30) on 17 February 1993, the Yalu River, northern China (n = 30) on 3 May 1993, and the Ichinomiya River, eastern Japan (n = 30) on 10 January 1994 (Fig. 9).
25
Fig. 9. Map showing the geographic distributions of Anguilla japonica (thick gray lines on the coastlines) and A. marmorata (thick black lines on the coastlines) in East Asia and collection sites of samples analyzed in this study (∆, A. marmorata; Ο, A. japonica ; Cheng and Tzeng 1996). General patterns of current systems in the western North Pacific and central Indonesian Seas (adapted from Nitani 1972, Lukas et al. 1991, and Revendin et al. 1994) and spawning grounds of A. japonica (gray circle with the letter J) and A. marmorata (black circle with the letter M) (Kuroki et al. 2009, Tsukamoto et al. 2011) are also shown. Sampling locations:
Tungkang River (T) and Shuangshi River (S), Taiwan; Mingchiang River (M), Chyantarng River (C), and Yalu River (Y), China; Ichinomiya River (I), Japan; Cagayan River (P), the Philippines;
Hsiukuluan River (H), Taiwan; and Kurio River (J), Japan. NEC, North Equatorial Current; KC, Kuroshio Current; OC, Oyashio Current; TS, Tsushima Current; CCC, China Coastal Current;
SCSWC, South China Sea Warm Current; IK, Intruded Kuroshio; MC, Mindanao Current;
NECC, North Equatorial Counter Current; SEC, South Equatorial Current.
26
Table 2. Sampling information and sample sizes of A. japonica and A. marmorata glass eel collection in various rivers and estuaries in East Asia. Values inside the parentheses indicate the number of individuals used for otolith analyses.
*from Cheng and Tzeng (1996)
Hsiukuluan River, Taiwan Kurio River, Japan
45 (13) 19 May 2008
168 (41) 37 (15) 86 (13) 6 Jun. 1996
20 May 2008 A. marmorata Cagayan River, Philippines
24 Mar. 1993 Shuangshi River, Taiwan 20 Dec. 1992 17 Feb. 1993
240 (131) 30 (20) 30 (23) 30 (23) 30 (10) Mingchiang River, China
Chyantarng River, China Yalu River, China
Ichinomiya River, Japan 10 Jan. 1994 03 May 1993 17 Feb. 1993 1 Mar. 1993
30 (13) 30 (12) 30 (14)
Species Sampling site Sampling date n
Tungkang River, Taiwan 30 Dec. 1992 30 (16)
A. japonica*
27 2.2 Juvenile eel collection
Juvenile A. marmorata were collected in Aguang River in eastern Luzon, Philippines (Fig. 10) from June-August 2012 to determine the migration pattern and habitat use. The eels were caught using an electro-fishing gear with the help of some local fishermen in the area (Fig.
11). For comparison, samples from an aquaculture farm in Sanya, southern Hainan province, China was included in the analyses.
Fig. 10. Sampling location of the adult eels in a river system in Aurora province in eastern Luzon, Philippines.
28
Fig. 11. Electro-fishing gear (a) used to catch A. marmorata juveniles in a river system (b) in Aurora province, eastern Luzon, Philippines.
29
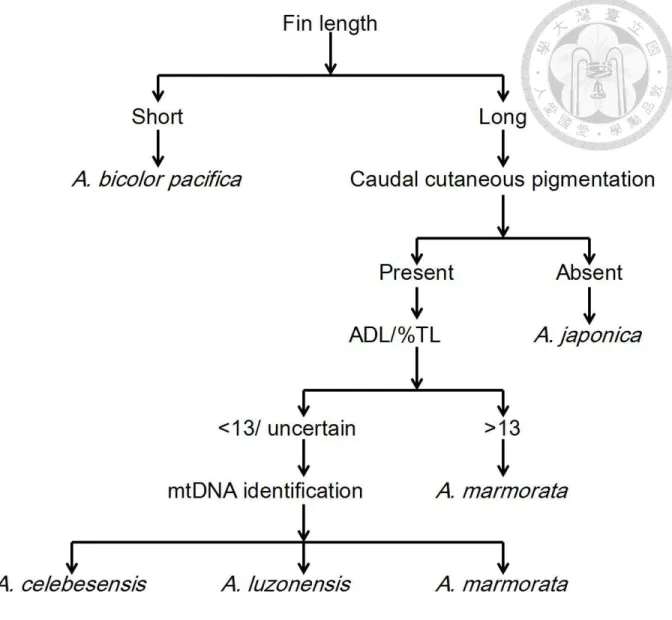
2.3 Glass eel species identification and morphometric measurements
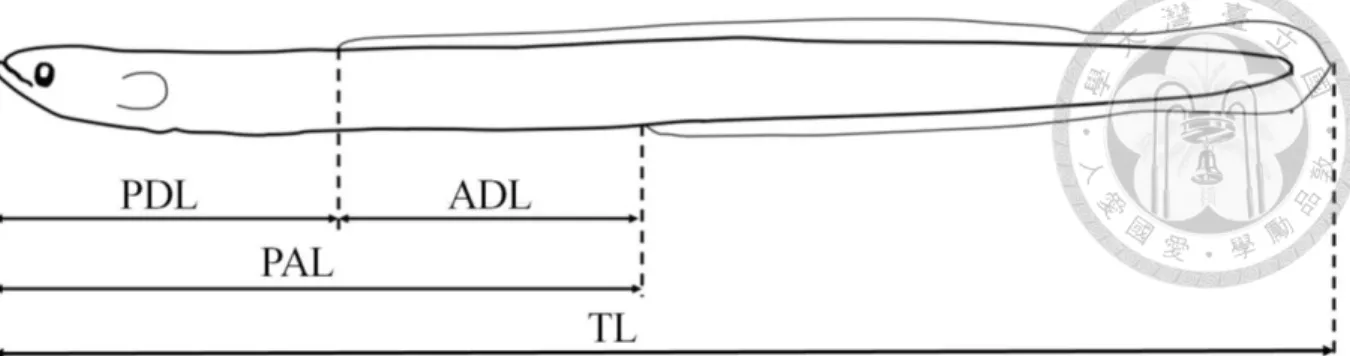
Glass eel species were identified using morphological characters and pigmentation patterns summarized in Fig. 12 which was modified from Tzeng (1982, 1983a) and Tzeng and Tabeta (1983). Morphological characters, including total length, pre-dorsal fin length (PDL), pre-anal fin length (PAL), and ADL, were measured to the nearest 0.1 mm (Fig. 13). Total length (TL) was determined by measuring the distance between the tip of the snout and the end of the tail, while the PDL was determined by measuring the distance from the tip of the snout to the origin of the dorsal fin. The PAL was determined by measuring the distance from the tip of the snout to the origin of the anal fin. The ADL, on the other hand, was the difference in distance between the origin of the dorsal and anal fins in percent of the total length (ADL/%TL). In addition, tail bud and caudal fin cutaneous pigmentation patterns, which appear during the glass eel pigmentation process, were also used for species identification.
Glass eels were classified into long- and short-finned types according to the value of ADL/%TL. Individuals with ADL/%TL values of < 5% were classified as short-finned eels (the pre-dorsal fin origin is closer to the anus than jaw), while individuals with AD/%TL values of >
5% were classified as long-finned eels (the pre-dorsal fin origin is closer to the jaw than anus) (Ege 1939, Tesch 2003). Anguilla bicolor pacifica is a short-finned eel with pigmentation in the tail bud that extends to the caudal fin, while the rest are long-finned eels. The 3 long-finned eel species were then separated according to the cutaneous pigmentation patterns on the posterior part of the body. Anguilla japonica has no pigmentation at this stage, while both A. marmorata and A. luzonensis and/or A. celebesensis have more or less the same pigmentation patterns in the tail bud. Anguilla marmorata and A. luzonensis and/or A. celebesensis were then separated according to ADL/%TL values. The ADL/%TL was reported to differ between these species but
30
there is some degree of overlap (Tzeng 1982, Teng et al. 2009, Watanabe et al. 2009).
Individuals with ADL/%TL values of > 13 were classified as A. marmorata, while those with values of < 13 were classified as A. luzonensis and/or A. celebesensis. Furthermore, the reliability of using ADL/%TL to discriminate A. marmorata, A. luzonensis, and/or A. celebesensis was tested by molecular identification. In total, 6 individuals with minimum, medium, and maximum values of ADL/%TL were chosen for the DNA analysis.
The morphometric datasets were subjected to normality and equal variance tests because of the unequal sample sizes. If the dataset passed the test, significant differences were examined using a one-way analysis of variance (ANOVA) followed by pairwise multiple comparisons using the Holm-Sidak method. On the other hand, if the dataset failed the test, significant differences were examined using a Kruskal-Wallis ANOVA on ranks followed by multiple comparisons using Dunn’s method. Holm-Sidak and Dunn’s post-hoc tests were conducted to detect pairwise differences between species with an overall alpha level of 0.05. All statistical analyses were carried out using SigmaStat software vers. 3.5 (Systat Software, San Jose, CA, USA).
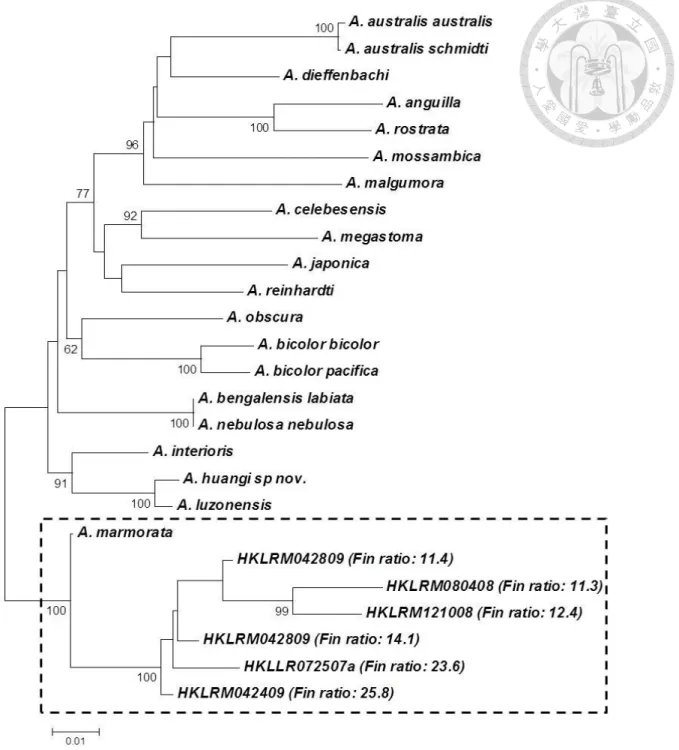
2.4 DNA extraction, polymerase chain reaction (PCR) amplification and phylogenetic analysis
Total genomic DNA was extracted from muscle tissues of individuals with minimum, medium, and maximum values of ADL/%TL using a DNA purification and extraction kit. A pair of oligonucleotide primers, H15341 (5’-TGCTAACGATGCCCTAGTGG-3’) and L151341 (5’-CTAGTCAACCTACTAATGGG-3’) was used to amplify a fragment of Cyt b using PCR amplification (Han et al. 2002). PCR amplification was carried out in a 25-µl reaction mixture containing 0.5 µl template DNA, 2.5 µl 10x reaction buffer, 0.5 µl dNTP, 1 µl of each forward
31
and reverse oligonucleotide primers, 0.25 µl DNA Taq polymerase, and 19.25 µl double-distilled water. The thermal profile consisted of initial denaturation at 94°C for 3 min followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 50-55°C for 1 min, and extension at 72°C for 30 s, with a final extension at 72°C for 10 min. PCR products were electrophoresed on a 1%
agarose gel and stained with ethidium bromide (EtBr) for band characterization via ultraviolet trans-illumination. Sequencing reactions were performed using an ABI PRISM 377 Auto DNA Sequencer (Applied Biosystems, Foster City, CA, USA).
The generated sequences were compared to the mitochondrial (mt)DNA Cyt b sequences of all known species and subspecies of Anguilla retrieved from GenBank (accession nos.:
AB038556, AB469437, and AP007233-49) to determine their phylogenetic relationships using the Neighbor-joining (NJ) method with the Kimura two-parameter model as implemented in MEGA 4.1 (Tamura et al. 2007). The resultant topology was assessed by bootstrapping with 1000 replications.
32
Fig. 12. Schematic diagram of the methods used for anguillid glass eel species identification.
33
Fig. 13. Diagram showing morphometric measurements of the glass eel. PD, pre-dorsal length;
AD, ano-dorsal length, PA, pre-anal length and TL, total length. Glass eels were classified into long- and short-finned types according to the value of ADL/%TL.
2.5 Determination of glass eel developmental stages
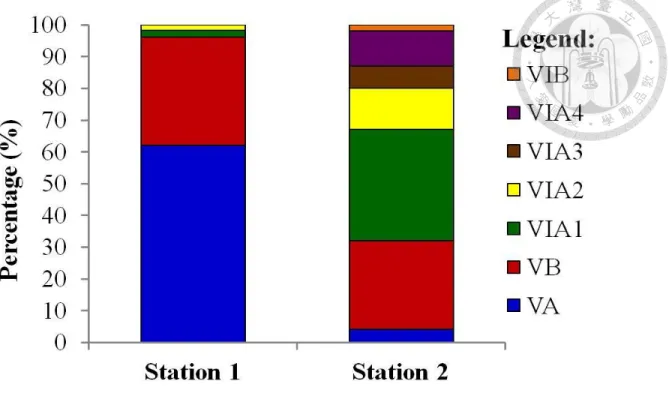
Developmental stages from glass eel to elver were also determined according to the extent (or absence) of skin pigmentation over the head, tail, and other body regions following methods described by Strubberg (1913), Bertin (1956), and Tesch (1977, 2003) (Fig. 14a-b, Table 1). Post-metamorphic juveniles were sub-classified into stages VA, VB, VIA1, VIA2, VIA3, VIA4, VIB,and VII. Juveniles up to stage VIA2 were classified as glass eels, while those in stages VIA3 and VIA4 were in the transition stage to elvers, which become fully pigmented at stage VIB stage (Fukuda 2010). Stage VIB indicates the end of pigmentation, while stage VII represents the fully pigmented, benthic elver (Tabeta and Mochioka 2003).
34
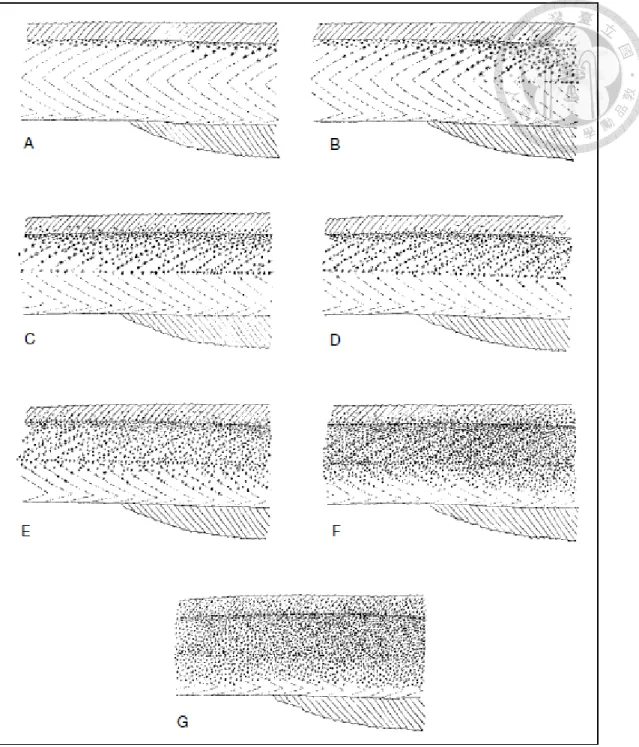
Fig 14a. Progressive sub-epidermal pigmentation in the anal region of the glass eel used to determine the different ontogenetic stages (after Strubberg, 1913; Bertin 1956; Tesch 2003). A:
Stage VIA1; B:Stage VIA2; C: Stage VIA3; D,E,F: Stage VIA4; G: Stage VIB.
35
Fig. 14b. Developmental stages (VIA1 to VIB) in Japanese eels showing pigmentation process from glass eel to the fully pigmented elver stage at VIb. Arrows indicates the location of the unique pigmentation characteristic of each stage (adapted from Fukuda et al. 2013)
36 Table 3. Development of pigmentation in anguillid eels (Tesch 2003).
Stage Characteristics
I Larva, fully grown leptocephalus
II Semilarva, pigmentation on the posterior end of the spinal chord
III Semilarva, pigmentation on the nerve chord becomes more extensive, skin pigment also seen at the tip of the caudal fin
IV Semilarva, pigmentation on the nerve chord reaches the head
VA Metamorphosis complete, eel-like form, no external pigment (glass eel) except the caudal spot
VB No pigment on the back, body or tail region, except for the skull, caudal spot and some rostral pigment
VIA1 Development of pigmentation along the whole dorsum, post-anal dorsolateral pigment develops, post anal, no clear mediolateral melanophores (Fig.14b-a)
VIA2 No pre-anal ventrolateral pigment. Post-anal development mediolateral pigment (Fig. 14b-b)
VIA3 No pre-anal ventrolateral pigment. Clear pre-anal development of mediolateral pigment, post-anally over almost entire dorsum, pigment rows along the myosepta and in places doubling of the mediolateral melanophores (Fig. 14b- c)
VIA4
Clear development of pre-anal ventrolateral pigmentation. Initially, in places, a doubling of the mediolateral
melanophores in the pre-anal region (Fig. 14b-d), post-anal pigment between the myosepta in the ventral region (Fig.
9b-e) and finally, similar changes in the pre-anal region (Fig. 14b-f)
VIB Pigment rows along the myosepta becoming indistinct. Lateral line still recognizable, as are the individual melanophores on the head, cheek, behind and below the eyes and on the lower jaw (Fig. 14b-g)
37 2.6 Morphometric measurements of adult eels
Morphological characters, including total length, pre-dorsal fin length (PDL), pre-anal fin length (PAL), and ADL, were measured to the nearest 0.1 mm (Fig. 15). Total length (TL) was determined by measuring the distance between the tip of the snout and the end of the tail, while the PDL was determined by measuring the distance from the tip of the snout to the origin of the dorsal fin. The PAL was determined by measuring the distance from the tip of the snout to the origin of the anal fin. The ADL, on the other hand, was the difference in distance between the origin of the dorsal and anal fins in percent of the total length (ADL/%TL).
38
Fig. 15. Diagram showing morphometric measurements of the adult eel. PDL, pre-dorsal length; ADL, ano-dorsal length, PAL, pre- anal length, SL, standard length, and TL, total length.