國立台灣大學生物資源暨農學院森林環境暨資源所 碩士論文
School of Forestry and Resource Conservation College of Bioresources and Agriculture
National Taiwan University Master Thesis
水黃皮之微體繁殖及體胚誘導初步探討
Preliminary study on the micropropagation and somatic embryogenesis induction of Pongamia pinnata
研究生:張文齊 Wen-Chi Chang
指導教授:王亞男 教授 Advisor:Dr. Ya-Nan Wang
中華民國 102 年 1 月
January, 2013
i
誌謝
本論文得以付梓完成,承蒙指導教授 王亞男博士之悉心教誨,在論文研究寫 作期間,不吝地給予指導與協助,使我了解做學問及待人接物應有的態度,委實 獲益良多,受用不盡。
口試期間感謝 李明仁老師、張淑華老師、林敏宜老師及林蘭東老師於百忙中 撥冗審閱論文及殷切指正,並賜予許多寶貴意見,使本論文更臻充實完備;特別 感謝 汪淮教授願意撥空修改英文摘要。
其次,要感謝實驗室的玲薇學姊、學文學長、天培、巍中、忠諺、佳紋、亭 潔及唯恩,謝謝大家的陪伴及包容,尤其同窗好友的禹安及俊佑,一路上互相打 氣與砥礪,讓我的研究生涯因你們增添不少笑聲,更讓我有重新出發的動力;還 有宜俊學長,在我剛進入台大校園,面對新的學校、新的同學及新的生活之際,
援助懵懂的我得以順利加入這個溫暖的大家庭;亦由衷感謝忠義學長,我們實驗 室的小家長,總是熱心的提供學術上的經驗及生活上的關心。
最後,僅以本論文獻給摰愛的家人,感謝一路上無論精神或生活上,無怨無 悔支持我的父母,有你們的後援我才能徜徉肆恣於浩瀚的學術領域中,更能無後 顧之憂完成我的碩士學業,回憶往事種種,感激之情,溢於言表;謝謝哥哥祐翔 不時的談天鼓勵及提供相機的協助;另感謝親愛的雙胞胎姊姊文慈,這段時間對 我無私的付出與關愛,包容我的任性,無論開心傷心的每一天,因為有你的陪伴,
都是有意義的每一天;最後是我們家最可愛的德國狼犬 mini,感謝有你安撫我的 心情;這段期間有你們默默的付與和關心,我才能努力不懈,很開心能和你們成 為一家人,我永遠愛你們。
其中感謝曾經幫助過我的貴人們,因為有你們不吝嗇的伸出援手,我才遂以 邁入新的人生階段,未來也希望大家都能平安順心,身體健康。
張文齊 謹誌於 台灣大學森林環境暨資源研究所 2013 年
ii
中文摘要
本試驗目的為建立水黃皮組織培養之微體繁殖及體胚誘導體系。
本試驗以水黃皮之帶節莖段為培殖體。將帶節莖段先以流水處理2 h,後以稀 釋300之安期消毒液震盪30 sec,再以70%之酒精震盪30 sec,最後在5%NaOCl水溶 液 (含約0.1% (v/v) Tween 20 展著劑) 之超音波中震盪30 min,無污染率可達65
%。
在器官形成方面:以MS或WPM中添加1 mg/L BA無菌苗之結果較佳。以水黃 皮無菌種子苗之子葉節、帶節莖段、莖段及下胚軸為材料,體胚誘導結果以子葉 節較佳,以添加1 mg/L BA前處理之子葉節培養1個月後,於MS添加2 mg/L BA及 0.1 mg/L NAA中,每個培殖體平均可產生5.42個芽體。以水黃皮母樹萌蘗之枝條為 培殖體,取春季正要突破休眠的春芽,芽體誘導在MS添加1-2 mg/L BA及0.1 mg/L NAA中較佳。芽體伸長結果以在MS添加1 mg/L GA3及0.2 mg/L BA中最佳(平均 1.18 cm)。將單一芽體切下,培養於1/2MS或MS添加IBA之發根培養基中誘導發 根,以在1/2MS添加1 mg/L IBA中之處理較佳,達發根率74.55%、平均產生2.96個 根,且平均根長度約1.33 cm。以子葉為培殖體進行體胚誘導,於MS添加3 mg/L 2,4-D與0.5 mg/L Kn2個月後,體胚誘導率為20.83%,且平均體胚數為3個。
發根的小苗經馴化結果,馴化6週的成活率為85.88%,且莖伸長約0.78 cm。
關鍵字:水黃皮、微體繁殖、體胚誘導、馴化
iii
Abstract
The main objectives of this study are to purpose and establish the micropropagation and somatic embryogenesis system in the tissue culture of Pongamia pinnata.
Results of nodal stem segments of epicormics culture of P. pinnata were as followed:nodal stem segments were first treated with running water for two hr. , shaking in 1/300 (v/v) anticeptol solution (Benzel thyonium chloride U.S.P 10﹪ (w/v) , Alkyl aryl polyether alcohol 10﹪(w/v)) for 30 sec , then shaking in 70﹪ethanol solution for 30 seconds, then soaked in 5﹪NaOCl (supplemented with 0.1﹪ (v/v) Tween 20) and treated with ultrasonic shaking for 30 min and the nodal stem segments obtained 65﹪
non contamination.
In the aspect of organ formation : the medium of 1 mg/L BA in MS or WPM gained the best results of aseptic seedlings. Micropropagation of P. pinnata from cotyledonary nodes, nodal stem, epicotyls and hypocotyls explants derived from in vitro seedlings. Multiple shoots induction from cotyledonary node explants gained the best result of that from other explants. The greatest shoots were induced from the 1 mg/L BA pretreatmenting of cotyledonary nodes explants was achieved when MS was added with 2 mg/L BA and 0.1 mg/L NAA, average of 5.42 shoots per explants were formed in one month. Spring buds at the stage of ready to burst from dormancy and shoot explants in spring were collected from the coppice shoots of P. pinnata wild trees. MS medium containing 1-2 mg/L BA and 0.1 mg/L NAA was the best for inducing more shoots.
Shoot elongation of these multiple shoots was good at 1 mg/L GA3 and 0.2 mg/L BA in MS medium. Individual shoot were rooted on 1/2MS or MS supplemented with IBA.
iv
The 1 mg/L IBA in 1/2MS medium gave the highest percentage of rooting (74.55%) with root number of 2.96 roots/shoot and root length of 1.33 cm. Cotyledons were induced with the highest percentage of 20.83% and with average of 3 somatic embryo/per explant of somatic embryogenesis in media composed of MS medium supplemented with 2,4-D 3 mg/L and Kn 0.5 mg/L after two months.
Shoots with roots were suitable for acclimatization. The plantlets survival rates were 85.88% with average shoot length of 0.78 cm after six weeks of acclimatization.
【 Key words 】 Pongamia pinnata, micropropagation, somatic embryogenesis, acclimatization
v
目錄
誌謝 ...i
中文摘要 ... ii
英文摘要 ... iii
目錄 ... v
表目次 ...ix
圖目次 ...xi
壹、前言 ( Introduction ) ... 1
貳、前人研究 (Literature Review) ... 3
一、水黃皮之介紹 ... 3
(一) 生質能源 ... 3
(二) 植生復育 ... 3
(三) 生藥功能 ... 4
二、微體繁殖的定義 ... 6
三、微體繁殖的步驟 ... 6
四、水黃皮之微體繁殖 ... 7
(一) 培養基 ... 7
(二) 培殖體 ... 7
(三) 器官發生 ... 7
(四) 體胚發生 ... 12
參、材料與方法 (Materials and Methods) ... 16
一、材料 ... 16
vi
(一) 種子 ... 16
(二) 萌蘗枝條 ... 16
(三) 水黃皮之微體繁殖 ... 16
(四) 水黃皮組培苗之馴化 ... 17
(五) 水黃皮體胚之誘導 ... 17
二、方法 ... 17
(一) 種子處理 ... 17
(二) 試驗項目 ... 18
(三) 培養基 ... 22
(四) 培養環境 ... 22
(五) 形態觀察 ... 23
(六) 統計分析 ... 23
肆、結果 (Results) ... 28
一、水黃皮野外帶節莖段之表面消毒 ... 28
二、水黃皮無菌種子苗之培養與生長 ... 30
三、水黃皮之芽體誘導試驗 ... 34
(一) 種子苗材料 ... 34
(二) 野外材料 ... 49
四、水黃皮之芽體伸長試驗 ... 53
五、水黃皮芽體誘導發根之試驗 ... 55
六、水黃皮切片觀察結果 ... 57
(一) 芽體試驗 ... 57
(二) 芽體伸長試驗 ... 57
vii
(三) 發根試驗 ... 57
七、水黃皮組培苗之馴化 ... 60
八、水黃皮體胚誘導之初步探討 ... 61
(一) 培殖體特性 ... 61
(二) 各植物生長調節劑誘導 ... 61
伍、討論 (Discussion) ... 66
一、水黃皮野外帶節莖段之表面消毒 ... 66
二、水黃皮無菌種子苗之培養與生長 ... 66
(一) 發芽率 ... 66
(二) 平均根長度 ... 67
(三) 平均莖長度 ... 67
三、水黃皮芽體誘導試驗 ... 68
(一) 種子苗材料 ... 68
(二) 野外材料 ... 70
四、水黃皮之芽體伸長試驗 ... 71
(一) 培殖體差異 ... 71
(二) GA3與 BA 兩種植物生長調節劑之影響 ... 71
五、水黃皮芽體誘導發根之試驗 ... 72
(一) 基礎培養基 ... 72
(二) IBA 對發根之影響 ... 72
六、水黃皮組培苗之馴化 ... 73
七、水黃皮體胚誘導之初步探討 ... 73
(一) 培殖體特性 ... 73
(二) 各植物生長調節劑誘導 ... 74
viii
陸、結論 (Conclusion) ... 75 柒、參考文獻 ( References ) ... 76 捌、附錄 (Appendix) ... 87
ix
表目次
表 1 水黃皮之藥用功能 ... 5
表 2 水黃皮莖段誘導之結果 ... 9
表 3 水黃皮莖段伸長之結果 ... 11
表 4 水黃皮根的誘導結果 ... 12
表 5 豆科樹種體胚誘導之結果 ... 13
表 6 水黃皮帶節莖段之表面消毒試驗組合 ... 18
表 7 水黃皮無菌種子苗誘導在 MS、1/2MS 及 WPM 之組合 ... 19
表 8 水黃皮芽體伸長在 MS 添加 BA 與 GA3之試驗組合 ... 20
表 9 水黃皮發根試驗在 MS 或 1/2MS 添加 IBA 之組合 ... 21
表 10 水黃皮體胚誘導在 MS 添加 Kn、BA 與 2,4-D 之組合 ... 22
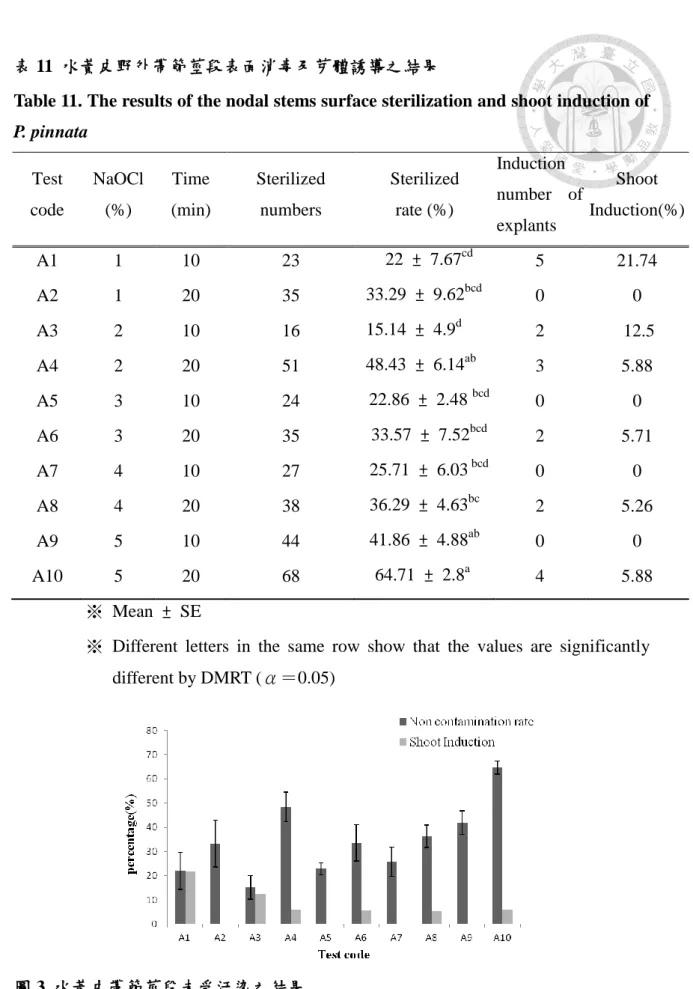
表 11 水黃皮野外帶節莖段表面消毒及芽體誘導之結果 ... 29
表 12 水黃皮無菌種子苗誘導與培養 2 個月之枝條生長結果 ... 31
表 13 水黃皮之子葉節與帶節莖段在 WPM 前處理於 BA 與 NAA 不同組合對芽體 之誘導結果 ... 40
表 14 水黃皮之子葉節與帶節莖段在 GA3前處理下於 BA 與 NAA 不同組合對芽體 誘導之結果 ... 41
表 15 水黃皮之子葉節與帶節莖段在 BA 前處理於 BA 與 NAA 不同組合對芽體誘 導之結果 ... 42
表 16 水黃皮之子葉節與帶節莖段在 BA 前處理於 BA 與 IBA 不同組合對芽體誘導 之結果 ... 43
x
表 17 水黃皮之子葉節與帶節莖段在 TDZ 不同濃度對芽體誘導之結果 ... 44
表 18 水黃皮生長點在不同 BA 及 NAA 組合下枝條生長之結果 ... 50
表 19 水黃皮子葉節及帶節莖段芽體伸長試驗之結果 ... 54
表 20 水黃皮發根試驗之結果 ... 55
表 21 水黃皮未成熟胚誘導體胚之結果 ... 62
表 22 水黃皮未成熟胚與子葉誘導體胚之結果 ... 63
xi
圖目次
圖 1 水黃皮 ... 24
圖 2 水黃皮種子發育之各階段 ... 26
圖 3 水黃皮帶節莖段未受汙染之結果 ... 29
圖 4 水黃皮之種子苗培養 2 個月 ... 32
圖 5 水黃皮在 MS 添加 2 mg/L BA 及 0.1 mg/L NAA 之芽體誘導結果 ... 37
圖 6 水黃皮前處理子葉節與帶節莖段在 2 mg/L BA 及 0.1 mg/L NAA 中芽體誘導之 結果 ... 39
圖 7 水黃皮子葉節在 BA 前處理與 BA 及 0.1 mg/LNAA 中 1 個月芽體數誘導之結 果 ... 39
圖 8 水黃皮以莖段及下胚軸為培殖體培養於不同培養基誘導之芽體及癒合組織 45 圖 9 水黃皮芽體誘導之結果 ... 51
圖 10 水黃皮野外帶節莖段經表面消毒後芽體誘導之結果 ... 56
圖 11 水黃皮在 0.2 mg/L BA 及 1 mg/L GA3中 1 個月芽體伸長之結果 ... 56
圖 12 水黃皮子葉節芽體在 1/2MS 添加 1 mg/L IBA 中 1 個月發根之結果 ... 56
圖 13 水黃皮切片之結果 ... 58
圖 14 水黃皮組培苗馴化之結果 ... 60
圖 15 水黃皮體胚誘導 2 個月之結果 ... 64
1
壹、前言 ( Introduction )
水黃皮 (Pongamia pinnata) 為豆科 (Fabaceae) 蝶形花亞科 (Faboideae) ,原 產於印度的中型喬木,具有景觀樹木、防風樹種、木材利用、飼料,葉與種子經 榨油後的種餅 (seed cake) 可作為牛及禽畜的飼料,且種餅經醱酵後所產生的生物 氣 (biogas) 亦可用於瓦斯燃料 (Sugla et al., 2007) 等功能。其種子油可作為生質 能源,豆莢與柴薪亦可供第二代生質能源纖維乙醇 (cellulosic ethanol) 之原料,另 民俗療法,可用其治療風濕病、皮膚病等;樹葉可作綠肥 (徐聖等,2010;應紹舜,
2011) 。
面對有限之石化燃料、原油價格高漲及溫室氣體日增等問題,尋找永續、具 再生性且對環境友善的新能源已成為一個國家賴以生存及發展經濟所必頇努力的 目標。因油料作物生長過程中所吸收的二氧化碳可抵銷使用過程中二氧化碳的釋 放,不會造成大氣中二氧化碳濃度的上升,故生質柴油被視為碳中性 (carbon neutral) 的綠色能源 (Kumar and Sharma, 2011) ,如此不僅可減少石化燃料之使 用,並能降低溫室氣體的排放。
現今面臨石化原料逐漸耗竭以及環保意識形成,台灣目前對於重金屬污染土 壤之處理採取休耕策略,如能於休耕或受重金屬污染之土地中種植能源作物,在 所種植植物生產之生質能合乎法規要求之前提下,利用此生質能源,不但達到受 污染土地再利用的目的,對於環境品質而言更可以達到減少溫室氣體之目的 (賴鴻 裕及盧至人,2007) ,另外藉由植物生長也可將土讓中重金屬吸收,累積一定濃度 之重金屬,以幫助土壤復育。
鑒於上述水黃皮之效用,若其成為經濟造林木並大量推廣栽植,常受下列因 素影響: (1) 種子活性低; (2) 種子儲藏時因真菌感染而使種子發芽率降低而匱 乏; (3) 種苗易受根腐病菌 (Rhizoctonia hiemalis) 之感染,會有未成熟落葉、枯 萎和生長不良等症狀; (4) 因種皮過厚而難以發芽 (Sugla et al., 2007) 。水黃皮可 利用扦插、嫁接進行無性繁殖,但利用此方式種植出之植株扎根不深,因此非常 容易傾倒 (Azam et al., 2005) ,故無法在貧瘠和邊際土壤進行栽植。
水黃皮在國內並未有組織培養方面相關的研究,本研究期望藉由水黃皮培養 之種子苗其子葉節、莖段、下胚軸及帶節莖段,以及野外萌蘗生長點和葉片為材
2
料,誘導產生水黃皮芽體,並建立其微體繁殖體系 (micropropagation system) ; 利 用 3 種 發 育 階 段 之 未 成 熟 胚 與 成 熟 胚 之 胚 與 子 葉 , 誘 導 水 黃 皮 體 胚 發 生 (somatic embryogenesis) ,體胚培養主要有以下優點:(1) 根芽同步發生,可減少 繼代次數與多次操作受污染之風險; (2) 體胚體積小可高密度培養; (3) 無需切 割分離,簡化操作流程並減少對培殖體之需求; (4) 可製成人工種子,利於貯藏 運送; (5) 便於進行基因轉殖以達品種改良之目的 (林敏宜,2001;王亞男,
2006 ) 。以期將來大量栽植時能節省成本、縮短收穫時間,且收穫之時間容易預 期等優勢,期望提供未來學術研究及商業應用上參考。
3
貳、前人研究 (Literature Review)
一、 水黃皮之介紹
水黃皮花冠蝶形呈淡紫色,總狀花序腋生,花期為春秋兩季,約4-5月及9-11 月 (圖1-A) ;葉為奇數羽狀複葉 (圖1-B) ;果為莢果呈橢圓形,內含種子一粒,
鮮有兩粒;種子富含油質,長10-20 cm (Kesari and Rangan, 2010) 。水黃皮適合生 長於年降雨量500至2,500 mm之潮濕及亞熱帶環境 (Kumar and Sharma, 2011) 。
(一) 生質能源
水黃皮於栽植4-7後年開始結果,且單株林木的種子產量約為9-90 kg,推估種 植密度為100株/ha水黃皮,則有900-9,000 kg/ha的種子產量潛力 (Karmee and Chadha, 2005) 。水黃皮含油量隨著地區及品種的不同會略有變化,有報告指出採 集自印度德干高原西部之瑪哈拉希特拉省 (Maharashtra state) 種子含油量高達 49.8% (Manjare et al., 2003) ,油質分析方面,其脂肪酸甲酯 (fatty acid methyl ester, FAME) 的性能符合美國 (ASTM D6751) 、德國 (DIN 51606) 及歐盟 (EN 14214) 生物柴油標準規範,適合作為生物柴油 (Azam et al., 2005;Karmee and Chadha, 2005) ,這些性能包括:碘價 (iodine value) 為74.78-100.98,表示油品長期存放的 穩定性;十六烷值 (cetane number) 為50.85-59.11表示其具燃料燃燒能力 (Mukta et al., 2009) 。
水黃皮做為能源樹種的優勢為: (1) 不屬於農作物,因此不會產生糧食危機 的問題 (徐聖等,2010) ; (2) 水黃皮可忍受半乾燥邊陲地帶,種子苗能種植在邊 際性土地 (marginal land) ; (3) 種子高產量、高油量且品質優良; (4) 屬於豆科 植物,能行固氮作用 (nitrogen fixation) ,可降低氮肥使用量。因此商業化栽培水 黃皮已成為有利可圖之產業 (Karmee and Chadha, 2005) 。
(二) 植生復育
Shirbhate及Malode (2012) 以水黃皮實生苗栽植於受重金屬汙染之土壤中2個 月,分析其重金屬含量,結果顯著降低了銅、鋅、鉻及錳等重金屬含量,水黃皮 根可耐鹽在鹹水中生長良好,並可種植在退化、荒地或休耕的土地 (Tomar and Gupta, 1985;Kumar and Sharma, 2011) ,其具有改善土質的優點 (徐聖等,2010) ,
4
為具植生復育潛力之樹種。如其建立無菌體系後,可以嘗試基因轉殖工程,利用 基因轉殖形成金屬螯合素 (phytochelatin, PC) ,利用金屬螯合素可以緊緊的把重 金屬抓住,以植生復育達到去除重金屬之目的 (林梓銘,2005) 。
(三) 生藥功能
近幾年來也有越來越多研究指出水黃皮之藥用潛力 (表1) ,其根的皮分離出 pongabiflavone及karanjabiflavone,有治療牛皮癬的研究 (Ghosh et al., 2011) ,並使 用乙醇萃取根部,取得水黄皮根總黄酮 (puerariae radix flavones, PRF) ,可能與減 少自由基生成、抑制脂質過氧化反應與增強氧化能力,以干預聚糖硫酸鈉 (dextran sulfate sodium, DSS) 誘導潰瘍性結腸炎小鼠作用顯著 (李靖等,2008) ,乙酸乙酯 萃取物 (acetic ether extract from P. pinnata roots, PRA) 可抑制胃粘膜組織超氧歧化 酶 (superoxide dismutase, SOD) 活性降低、丙二醛 (malondialdehyde, MDA) 生成 增加及一氧化氮減少,可抗小鼠實驗性胃潰瘍及抗炎鎮痛(劉可雲等,2007;劉可 雲和朱毅,2010) ;花的萃取物對順雙氨雙氯鉑 (cisplatin) 和全達素 (gentamicin) 所引起的腎損傷具有抗氧化活性,對腎臟有保護作用 (Shirwaikar et al., 2003) 並抗 糖尿病 (Punitha and Manoharan, 2006) ;果之種子油可抗菌,且利用乙醇萃取後,
水黃皮醇及水黃皮素可降血糖 (Tamrakar et al., 2008;Kesari et al., 2010;Tamrakar et al., 2011) ,果萃取物可抗血脂異常及抗病毒 (Elanchezhiyan et al., 1993;Bhatia et al., 2008;Vadivel and Biesalski, 2011) ;葉子乙醇萃取物有抗血脂及抗氧化 (Essa and Subramanian, 2006;Essa et al., 2010) ,可作為合成抗蝨劑 (Sunilson et al., 2009) 等。
5
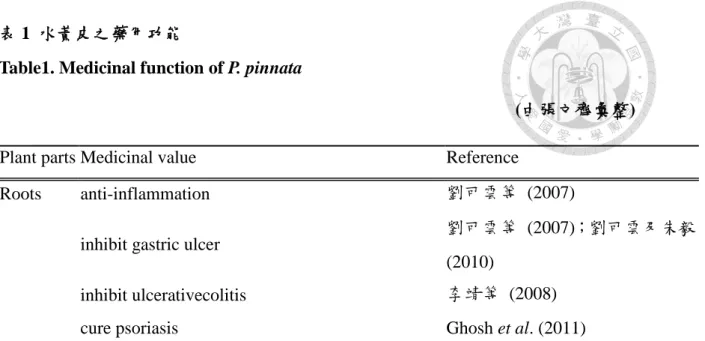
表 1 水黃皮之藥用功能
Table1. Medicinal function of P. pinnata
(由張文齊彙整) Plant parts Medicinal value Reference
Roots anti-inflammation 劉可雲等 (2007)
inhibit gastric ulcer
劉可雲等 (2007);劉可雲及朱毅 (2010)
inhibit ulcerativecolitis 李靖等 (2008)
cure psoriasis Ghosh et al. (2011)
Flower protective renal Shirwaikar et al. (2003)
inhibit alloxan induced diabetic Punitha and Manoharan (2006) Seed antihyperglycemic and antilipidperoxidative Elanchezhiyan et al. (1993)
anti-dyslipidemic activity Bhatia et al. (2008) antihyperglycemic activity
Tamrakar et al. (2008);Tamrakar et al. (2011)
antimicrobial activity Kesari et al. (2010) against herpes simplex virus HSV-1 and
HSV-2 Vadivel and Biesalski (2011)
Leaf against cholera and enteroinvasive bacterial Brijesh et al. (2006) anti-hyperammonemic and antioxidant Essa and Subramanian (2006) inhibited White Spot Syndrome Virus Rameshthangam and Ramasamy
(2007)
antimicrobial agent Bajpai1et al. (2009) resistance to synthetic anti-lice agents Sunilson et al. (2009) hypolipidaemic and antihyperlipidaemic Essa et al. (2010)
anticonvulsant activity Manigauha and Patel (2010)
6
二、微體繁殖的定義
微體繁殖為組織培養範疇之一,利用植物細胞分化全能性 (totipotency) ,即 每個細胞都具有該植物遺傳信息的特性,離體細胞在一定培養條件下,經過逆分 化 (dedifferentiation) 和 再 分 化 (redifferentiation) 的 步 驟 , 藉 由 器 官 發 生 (organogenesis) 或體胚發生兩條途徑,重新合成具再生完整能力的個體,以達到大 量繁殖的目的。
三、微體繁殖的步驟
最初由 Murashige (1974) 將微體繁殖概分為無菌苗的建立、誘導並增殖芽體 或體胚、發根及馴化等三個階段;至今 Debergh 及 Maene (1981) 為求商業生產效 率及經濟考量所需,將其分成五個階段:第零階段為培殖體準備階段:母株或培 殖體在人為控制環境下培養,或給予適當的處理,以獲得更健康且均質之培殖體。
第壹階段為無菌體系之建立:此階段培殖體需達無菌條件外,還頇考慮額外添加 劑 (如:抗生素或抗氧化劑) 對培殖體存活率及再生率之影響,Robert 及 Dennis (2000) 指出能成功建立第壹階段的四個因素,分別為: (1) 培殖體的生理時間; (2) 培殖體的取材位置; (3) 培殖體的大小; (4) 培殖體的褐化問題。第貳階段為增 殖階段:在適當的培養方式下,先產生器官發生並促進芽體大量生長,而器官發 生途徑有二,一為直接器官發生,即由培殖體直接誘導器官形成;二為間接器官 發生,先由培殖體產生癒合組織 (callus),再進行誘導器官體系形成,但根據 Cecchini 等 (1992) 利用荷蘭豆 (Pisum sativum) 芽體作為培殖體誘導癒合組織及 間接產生芽體測試 DNA 含量,結果兩者在 DNA 含量皆降低,顯示癒合組織在長 時間繼代培養下對 DNA 含量可能產生的變異,對種原保存及大量繁殖苗木而言,
並不是一種理想繁殖方式。第參階段為芽體的伸長、根原體誘導及根系發育階段,
使其可成功移植到土壤環境中生長所準備,許多木本植物在此階段以添加細胞生 長素 (auxin) 類之植物生長調節劑(plant growth regulators, PGRs),在適當的植物生 長調節劑種類與濃度下有助於植物體之發根。第肆階段為移植階段:剛自植株培 養在無菌容器移出於自然環境時,主要將其營養來源由混營性 (mixtrophy) 或異營 性 (heterotrophy) 生長,達到進行光自營性 (photoautophy) 生長之能力,藉由馴
7
化 (acclimatization) 及健化 (hardening) 的步驟,使植株能適應外界水分逆境、病 蟲危害及溫度、光度等變化,使組織培養苗可面對自然環境的劇烈變化,能存活 且健康的生長。
四、水黃皮之微體繁殖 (一) 培養基
一般植物較常採用的是 Murashige and Skoog (MS)培養基,而木本植物以 woody plant medium (WPM) 培養基 (Lloyd and McCown, 1980) 培養基最常見。
MS 培養基之鹽類配方在芽體誘導及開始階段較佳,但過高的鹽類濃度較不適合大 多數木本植物之芽體培養 (Tonon et al., 2001) ,故許多木本植物會以調整過鹽類 較低的 1/2MS 或 WPM 基本培養基進行試驗。
為了培殖體的誘導反應,我們會在培養基上添加各種不同的植物生長調節 劑 ,常用的植物生長調節劑分為細胞生長素 (auxins, a) 、細胞分裂素 (cytokinins, c) 和激勃素 (gibberellins) 三大類;在植物組織和器官培養的應用上,細胞生長素 和細胞分裂素是目前調節生長和形態發生最重要的調節劑。前人研究發現細胞生 長素與細胞分裂素的比率 (a/c) 降低時,可促進芽的分化;比率提高時,則可促進 根及癒合組織的形成 (Sita, 1979) 。
(二) 培殖體
利用植物體之組織或器官作為培養材料稱為培殖體,包括植物根、莖、葉、
芽及胚等。培殖體會因來源部位、生理狀態等條件而影響試驗結果 (George and Sherrington, 1984) 。 水 黃 皮 芽 體 誘 導 最 常 被 使 用 的 培 殖 體 就 是 子 葉 節 (cotyledonary nodes), Bailey (1956) 指出子葉節位於子葉之著生點,與維管束 (vascular strands) 相接連,且為幼苗之原始部分。
(三) 器官發生
器官發生為培殖體經誘導後再生成新的再生體 (regenerant) ,為形態發生的途 徑之一,不同於體胚發生為具有形態雙極性,即在發育的早期階段從方向相反的
8
兩端分化出莖端及根端,而器官發生需要透過各種植物生長調節劑,使其進一步 再分化出芽原體 (primordial shoot) ,原生莖段不具封閉型維管束,並埋於培殖體 組 織 內 , 其 伸 長 出 成 莖 段 , 再 經 由 莖 段 基 部 發 根 成 獨 立 植 株 (George and Sherrington, 1984) 。
1.芽體誘導
Sugla 等 (2007) 指出水黃皮子葉節在未添加 PGRs 的MS或 WPM基本培養基 中,即無莖段增生現象,若添加相同濃度N6-benzylaminopurine (BA) 於 WPM 及 MS 培養基,則在 MS 培養基中有較佳芽體誘導的效果,水黃皮的微體繁殖中,以 MS 培養基較常被使用 (Purohit and Singhvi, 1998) 。
至於添加的植物生長調節劑,Murthy等 (1998) 認為在培養基內添加高濃度的 細胞分裂素能促進芽體大量增殖,但低濃度的thidiazuron (TDZ) 就能促進組織生
長,且芽體的再生能力與其他細胞分裂素相同。Sugla等 (2007) 利用對水黃皮子
葉節誘導芽體之結果發現BA比kinetin (Kn) 更具影響且細胞分裂素的種類與濃度 對芽體生長具深遠的影響。TDZ在水黃皮的微體繁殖中有雙重作用,將其選擇性 的加強分生組織細胞以產生更多具分化成地上部的芽 (caulogenic buds) 細胞,但 同時抑制這些細胞及分生組織分化成莖段 (Sujatha and Hazra, 2007;Shrivastava and Kant, 2010) ,此也表示在高濃度會明顯抑制分生細胞快速的增殖 (Sujatha and Hazra, 2007) 。大體而論,水黃皮以子葉節、子葉及帶節莖段為培殖體,主要誘導 芽體的培養基為TDZ或BA並輔以低濃度細胞生長素 (表2) 。
Belide 等 (2010) 指出在含有 1 mg/L BA 的 MS 培養基中先進行種子萌芽之前 處理,可觀察到比對照組更大且發育更完整的葉原體 (leaf primordial) ,並可誘導 出約兩倍的芽體數,將其子葉節培養於 1 mg/LTDZ 及 200 mg/L adenine sulfate (ADS) 的 MS 培養基中,可誘導出最多芽體數,以證明 ADS 具有促進細胞分裂活 性,添加於培養基中可促進芽體生長或加強反應。Sugla 等 (2007) 對於水黃皮的 種子苗誘導,得到將去種皮種子培養於添加1.0 μM gibberellic acid (GA3) 的 MS 培 養基中,94%再生單一芽體,不添加 PGRs 的 MS 培養基之再生芽體則為 28%,
顯示植物生長調節劑有利於水黃皮去種皮之種子萌芽。
9
表 2 水黃皮莖段誘導之結果
Table 2 . The inducted results of shoot of P. pinnata
(由張文齊彙整)
Explants MS medium + PGRs Induction
time(days) Results Reference Cotyledonary
nodes
7.5 μM BA 12 6.8 shoots / explant Sugla et al. (2007)
shoot length : 0.67 cm
87% shoot induction
Nodal explant 4.54 μM TDZ 28 10.7shoots /explant Sujatha and Hazra (2007)
shoot length : 0.83 cm
66% shoot induction
Cotyledon 200 mg/L ADS 20 93% shoot induction Sujatha et al.
(2008) 11.35 μM TDZ 205 buds /explants
66.9% bud induction Cotyledonary
nodes
1 mg/L BA pretreatmenting
28 32.2 shoots /explant Belide et al.
(2010) 1 mg/L TDZ
Cotyledonary nodes
8.8 μM BA+0.53 μM NAA
28 2.2 shoots /explant Shrivastava and Kant (2010) shoot length : 5.4 cm
80% shoot induction
10
2.芽體伸長
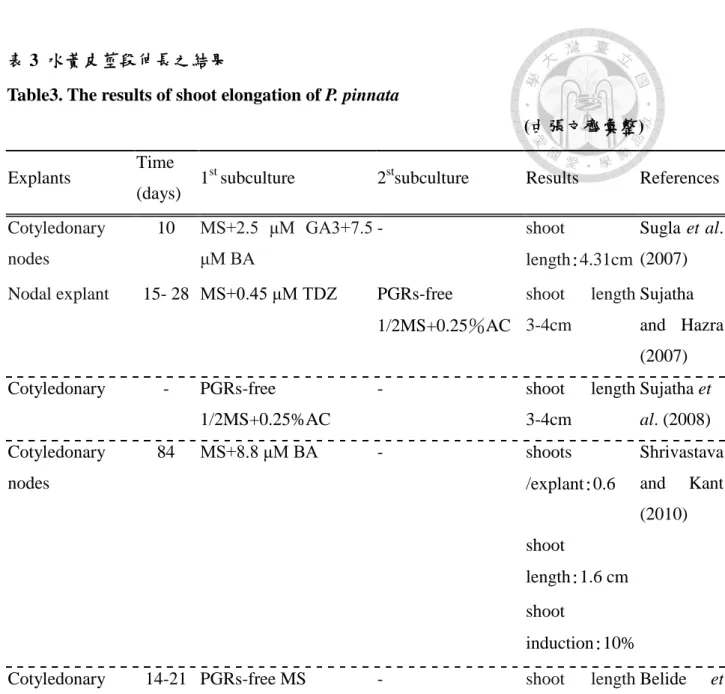
自培殖體誘導最佳莖段的培養條件建立後,當培養基養分不足或植物已可自 行合成 PGRs 時則需改變 PGRs,即需離體繼代在新鮮培養基或減少細胞分裂素濃 度,以伸長芽體。水黃皮芽體伸長之配方,主要以減少細胞分裂素濃度或輔以低 濃度 GA3的培養基作為繼代培養配方 (表 3) 。Sugla 等 (2007) 指出少量的 GA3
能有效影響水黃皮芽體的伸長,且添加 BA 於培養基中具促進發育不良的芽體伸長 的效果。Sujatha 等 (2008) 以水黃皮子葉誘導芽體於 TDZ 誘導中發現,雖然 TDZ 可誘導芽體但不能使其同時分化,子葉組織所誘導的芽沒有分化及伸長現象,故 在無 TDZ 的 MS 基礎培養基下繼代培養,再培養於 1/2MS 添加 0.25%活性碳作芽 體伸長。
同樣 Shrivastava 及 Kant (2010)以子葉節作為培殖體的則離體繼代於去除 α-naphathaleneacetic acid (NAA) 的培養基,可產生莖段團並進一步減少癒合組織 及促進芽體伸長。隨後 Belide 等 (2010) 甚至將子葉節重複繼代至不含 PGRs之培 養基中。
Sujatha 及 Hazra (2007) 自野外成熟的水黃皮取得帶節莖段作為培殖體,發現 含 TDZ 的培養基會抑制帶節莖段分生組織之分化,在高濃度 TDZ 誘導莖段需歷經
多次繼代且延遲發根階段,故先降低 TDZ 濃度以維持芽體數,而後移轉至無 PGRs
的 1/2MS 培養基中,使芽原體分化為芽體。
11
表 3 水黃皮莖段伸長之結果
Table3. The results of shoot elongation of P. pinnata
(由張文齊彙整)
Explants Time
(days) 1st subculture 2stsubculture Results References Cotyledonary
nodes
10 MS+2.5 μM GA3+7.5 μM BA
- shoot
length:4.31cm
Sugla et al.
(2007) Nodal explant 15- 28 MS+0.45 μM TDZ PGRs-free
1/2MS+0.25%AC
shoot length 3-4cm
Sujatha and Hazra (2007) Cotyledonary - PGRs-free
1/2MS+0.25%AC
- shoot length
3-4cm
Sujatha et al. (2008) Cotyledonary
nodes
84 MS+8.8 μM BA - shoots
/explant:0.6
Shrivastava and Kant (2010) shoot
length:1.6 cm
shoot
induction:10%
Cotyledonary nodes
14-21 PGRs-free MS - shoot length 3-5cm
Belide et al. (2010)
12
3.根之誘導
添加細胞生長素於培養基對芽體誘導發根是必要的。Sugla 等 (2007) 添加不 同濃度的 indole-3-butyric acid (IBA) 、indole-3-acetic acid (IAA) 及 NAA 於全量或 1/2MS 培養基中對子葉節所誘導芽體的發根量,MS 添加 1 μM IBA 的結果較佳,
顯示 IBA 對誘導發根有影響。在水黃皮組織培養研究中,多以添加 IBA 誘導發根 (表 4) ,且由於植物的莖頂會自動合成細胞生長素並誘導根部的發展,因此也有將 已伸長的芽體轉移至不添加任何植物生長調節劑的 MS 或 1/2MS 培養基中成功發 展出根部 (Sujatha and Hazra, 2007;Sujatha et al., 2008) 。
表 4 水黃皮根的誘導結果
Table 4. The inducted results of root of P. pinnata
(由張文齊彙整)
Medium Induction time (days)
Results References
MS+1 μM IBA 7-9 92% rooting Sugla et al. (2007) 3.4 roots/shoot
MS 28 70% rooting Sujatha and Hazra (2007)
1/2MS +0.25%AC 28 87.5% rooting Sujatha et al. (2008)
MS+9.8 μM IBA 28 93% rooting Shrivastava and Kant (2010) 2.5 roots/shoot
root length : 4.3 cm
1/2MS+1 mg/L IBA 28 64.5% rooting Belide et al. (2010) 5.3 roots / shoot
(四) 體胚發生
體胚發生為體細胞 (somatic cell) 在適當的誘導下,朝向具有胚性細胞發育,
生長分化後成體胚 (somatic embryo),為具形態雙極性之獨立個體,形成胚的發育 程序是從原胚開始,進入球胚期、心臟期、魚雷期與子葉期,而發芽為完整植株,
13
體胚發生的過程與結合子胚發育的過程相當類似,但未經過細胞融合階段,為無 性胚。
Gaj (2004) 指出,80%以上的研究只單獨用細胞生長素誘導體胚發生或結合細 胞分裂素,可能因細胞生長素及細胞分裂素高度參與細胞週期之調控與細胞分 裂,故成為體胚發生的關鍵因素,其中Raemakers等 (1995) 提出各細胞生長素中,
使用最多的是2, 4-dichlorophenoxyacetic acid (2,4-D) (49%) ,接著是NAA (27%) 、 IAA (6%) 、IBA (6%) 、picloram (5%)與dicamba (5%) ;細胞分裂素則以BA是最 常被使用 (57%) ,再來是Kn (37%)、zeatin (3%) 與TDZ (3%) 。
豆科植物之體胚誘導,培殖體以年輕之分生組織為主,如未成熟胚或葉子居 多,Aitken-Christie 等 (1985) 指出放射松 (Pinus radiata) 子葉之年齡,直接影響 芽體形成,可能是較老的子葉,所含內生性細胞分裂素和激勃素量少,不易形成 分生組織。誘導配方則以細胞生長素搭配細胞分裂素,較少單獨添加細胞分裂素 作誘導配方,以細胞生長素而言,根據物種及細胞生長素種類具有顯著差異性,
豆科植物多使用細胞生長素 2,4-D,能產生高頻率的體胚再生,但可能造成形態異 常及轉換植株上的困難 (Weaver and Trigiano, 1991;Lakshmanan and Taji, 2000) , 由於水黃皮之體胚誘導方面尚無研究發表,故本章僅以豆科木本植物之體胚誘導 研究結果作為參考 (如下表 5) 。
表 5 豆科樹種體胚誘導之結果
Table 5 . The inducted results of somatic embryogenesis of legumes trees
(由張文齊彙整)
Species Explant Medium Induction pathway
Induction time (days)
Results References
Acacia arabica
immature embryo
MS+Kn 0.5mg/L+
2,4-D 3 mg/L indirect 28
72.5%
Somatic
Embryogenesis
Das (2011) Acacia
catechu
immature embryo
MS+ Kn 1.5mg/L+
2,4-D 2 mg/L indirect 28
66.2%
Somatic
Embryogenesis
Das (2011)
14
表 5 (續) 豆科樹種體胚誘導之結果
Table5 (continued). The inducted results of somatic embryogenesis of legumes trees
Species Explant Medium Induction pathway
Induction time (days)
Results References Acacia
nilotica
immature endosperm
1st MS+2,4-D 2.5μM+BA 5μM
indirect 77 7% Somatic Embryogenesis
Garg et al. (1996) 2st MS+2,4-D
10μM+BA 25 μM Albizia
richardiana hypocoty MS+BA 10-5M indirect 40 1.4 Somatic Embryos/explan
Tomar and Gupta (1988)
Cajanus
cajan cotyledon MS+TDZ 10 μM indirect 42 33.8 Somatic Embryos/explant
Sreenivasu et al.
(1998) 45.3% Somatic
Embryogenesis leaf MS+TDZ 10 μM indirect 42 47.8 Somatic
Embryos/explan 73.6% Somatic Embryogenesis
cotyledon EC6+TDZ 4.54
μM direct 28
38.4 Somatic Embryos/explan
Mohan and Krishnamurthy (2002)
97% Somatic Embryogenesi cotyledon MS+TDZ 2 mg/L direct 42 73.34% Somatic
Embryogenesi Aboshama (2011) Calliandra
tweedii internode MS+2iP 10 μM
direct 90
22.33 Somatic Embryos/explan
Kumar et al.
(2002) 85% Somatic
Embryogenesis petiole MS+2iP 5 μM 26.47 Somatic Embryos/explan
69% Somatic
Embryogenesis Cassia
angustifolia cotyledon
MS+2,4-D 10μM+ BA 2.5μM
indirect 60 14.36 Somatic Embryos/explan
Agrawal and Sardar (2007)
70% Somatic
Embryogenesis Ceratonia
siliqua
somatic embryos
MS+IBA 0.5μM
+BA4.4 μM direct 84 33.8% Somatic Embryogenesi
Canhoto et al.
(2006)
15
表 5 (續) 豆科樹種體胚誘導之結果
Table5 (continued). The inducted results of somatic embryogenesis of legumes trees
Species Explant Medium Induction pathway
Induction time (days)
Results References
Cercis
canadensis ovule SH+2,4- D 9μM direct 42 4.6 Somatic Embryos/explants
Buckley and Trigiano (1994)
33% Somatic
Embryogenesis Cladrastis
lutea
immature
embryo SH+2,4-D 9 μM direct 42 46% Somatic Embryogenesi
Weaver and Trigiano (1991) Dalbergia
latifolia
immature embryo
MS+2,4-D 2 mg/L+ Kn 0.1 mg/L
direct 28 32 Somatic Embryos/explants
Rao and Lakshmisita (1996)
94.5% Somatic
Embryogenesis Dalbergia
sissoo cotyledon MS+ 2,4-D 9.04
µM + Kn0.46 µM indirect 40-50 26.5 Somatic Embryos/explants
Singh and Chand (2003) 55% Somatic
Embryogenesis immature
embryo
MS+ Kn 1.5mg/L+
2,4-D 2 mg/L indirect 28 76.2% Somatic
Embryogenesis Das (2011) Delonix
regia
immature zygotic
embryos MS+2,4-D 2 mg/L +BA0.25mg/L
indirect 28-35 45% Somatic
Embryogenesis Abdi and Hedayat (2011) immature
embryo
46% Somatic Embryogenesis Hardwickia
binata
immature cotyledon
MS+ Kn 4.64μM+
NAA5.37 μM indirect 21 56.8 Somatic Embryos/explants
Das et al.
(1995)
semi-mature embryos
MS+2,4-D 2.26μM direct 49
18.3 Somatic Embryos/explants
Chand and Singh (2001) 72.3% Somatic
Embryogenesis immature
embryo
MS+ Kn 0.5mg/L+
2,4-D 3 mg/L indirect 28 78.4% Somatic
Embryogenesis Das (2011) Robinia
pseudoacacia
immature seed
FM+ 2,4-D
45μM+ BA2.2μM direct 60 26.8Somatic Embryos/explan
Arrillaga et al. (1994)
16
參、材料與方法 (Materials and Methods)
一、材料 (一) 種子
本試驗在 2011 及 2012 年春季 (約 4、5 月) 及夏季 (約 7、8 月) ,在台灣大 學校本部隨機採集 8 棵水黃皮 (圖 1-C) 之成熟但外表呈綠色之莢果 (圖 1-D、E) 。
(二) 萌蘗枝條
本試驗在 2011 年約 5、6 月隨機採集 8 棵水黃皮,選取健康無病蟲害之萌蘗 (圖 1-F) ,採集天氣皆為晴天,將其切成 2-4 cm 之帶節莖段,每一帶節莖段皆含一個 節作為培殖體。
(三) 水黃皮之微體繁殖
1.水黃皮芽體誘導試驗 (1) 無菌苗為培殖體
由近成熟的綠色果莢 (圖2-A) 所建立之無菌苗於3種不同培養基 (WPM、
WPM+1 mg/L BA及WPM+0.35 mg/L GA3) 培養下,取其帶節莖段、莖段、子葉節 及下胚軸為誘導芽體之培殖體。
a、莖段、帶節莖段
無菌苗長至3-5 cm時,取長約0.5-1 cm不帶節之莖段及帶一個節之帶節莖段,
進行培養。
b、子葉節及下胚軸
無菌苗長至0.5至1 cm時,取約0.5-1 cm長之下胚軸,及各帶1/3之子葉之子葉 節,進行培養。
(2) 野外材料
本實驗在 2012 年春季 (5、6 月) ,在台灣大學校本部隨機採集 8 棵水黃皮之 萌蘗,取葉片及側芽進行培養。
a、葉片
以解剖刀將葉片切成大小約1.5×1.5 cm2,進行培養。
17
b、生長點
以解剖刀將芽切開,並用鑷子小心挑出約0.2-0.3 cm之生長點作培養。
(3) 水黃皮之芽體伸長試驗
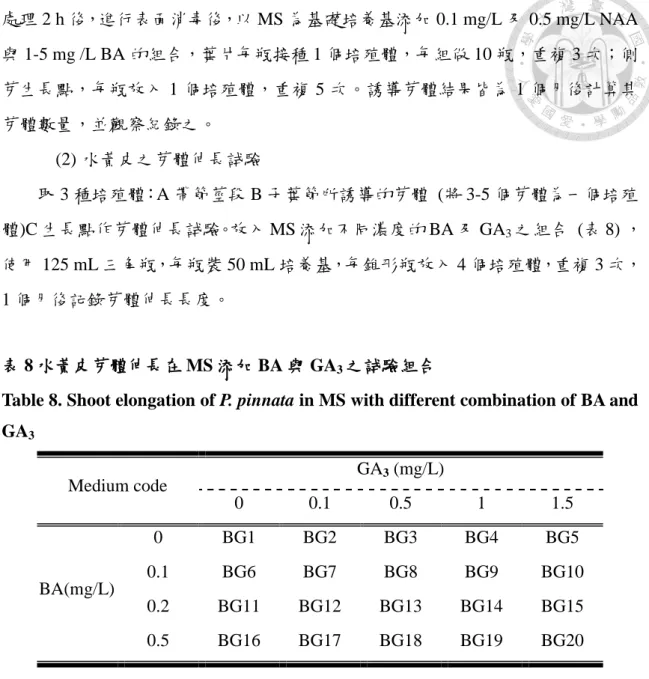
取 3 種培殖體作芽體伸長試驗,分別為帶節莖段及子葉節所誘導的芽體 (將 3-5 個芽體為一個培殖體)及生長點。
(4) 水黃皮芽體誘導發根之試驗
以子葉節誘導出之芽體取已抽長約 2-3 cm 為培殖體。
(四) 水黃皮組培苗之馴化
選取發根試驗中植株完全展葉且根長達 2-4 cm 時作出瓶馴化。
(五) 水黃皮體胚之誘導
本試驗在2011及2012年6-7月,於台灣大學校本部所種植8棵水黃皮隨機採集不 同成熟程度之種子。
1.帶胚乳之未成熟胚
當水黃皮花期結束約1個月後 (約6月) 採集莢果,參考張淑華等 (2002) 不同 發育階段牛樟 (Cinnamomum kanehirae) 種子的胚培養,依子葉佔種子內部體積的 比例,將胚的成熟度分成3種階段:未成熟胚A:子葉佔種子內部體積75%以上,
其餘為胚乳 (圖2-B) 。未成熟胚B:子葉佔種子體積之50-75% (圖2-C) 。未成熟胚 C:子葉佔種子體積50%以下 (圖2-D) ,取上述3種不同發育情況進行黑暗培養。
2.無胚乳未成熟胚
當水黃皮花期結束後約2月個後 (約7月) 之莢果,取種子佔整個莢果大小約 50-70% (圖2-E) ,並切大小約1×1×1 cm3之子葉及胚,進行培養。
二、方法
(一) 種子處理
建立離體培養時汙染為主要遭遇的問題,儘管經過幾個殺菌劑的反覆試驗和 各種消毒程序,高微生物及酚滲出物使成熟種子難建立種子苗,為了減少這些限 制,需在落果之前取得近成熟的綠色果莢,選取大小、形狀相似的種子作為發芽
18
試驗培殖體 (Sujatha et al., 2008) ,採集後的成熟莢果以封口袋密封後置於 15℃冰 箱冷藏,所有種子在 1 個月內使用完畢,並參考 Kumar 等 (2007) 以新鮮水黃皮 種子於 60℃熱水處理 15 min,可使休眠種子發芽達 100%,以此作為破除種子因 冷藏後可能造成種子休眠之問題。水黃皮種子表面消毒以安期消毒液稀釋三百倍 置於超音波震盪器震盪 10 min,然後再於 70%酒精震盪 5 min,兩步驟皆以二次水 沖洗 3 次,而後以 2%次氯酸鈉 (NaOCl) 消毒 (含 0.1% (v/v) tween20) ,最後移 入無菌操作台,以無菌水沖洗 3 次。
(二) 試驗項目
1.水黃皮野外帶節莖段之表面消毒
先以流水處理2 h再以安期消毒液稀釋300倍置於超音波震盪器震盪30 sec,然 後再於70%酒精震盪30 sec,兩步驟皆以二次蒸餾水沖洗4次,而後以次氯酸鈉 (NaOCl) 消毒。比較次氯酸鈉溶液 (含0.1% (v/v) tween 20) 1-5%濃度於10及20 min的消毒效果 (表6) ,於培養第10天記錄汙染率,且考慮到消毒藥劑過量可能造 成細胞喪失誘導活性,故於1個月後記錄芽體誘導率,共10個處理,每瓶放1個培 殖體,重複7次。
表 6 水黃皮帶節莖段之表面消毒試驗組合
Table6. Surface sterilization of nodal stems of P. pinnata by using different combined test for disinfectants
Test code NaOCl Time (%) (min)
A1 1 10
A2 1 20
A3 2 10
A4 2 20
A5 3 10
A6 3 20
A7 4 10
A8 4 20
A9 5 10
A10 5 20
19
2.無菌種子苗之培養與生長
以MS、1/2MS及WPM為基礎培養基,並以MS或WPM添加1 mg/L BA或0.35 mg/L GA3的組合進行試驗,如下表 (表7) ,以125 mL三角瓶分裝,每瓶50 mL。2 個月後記錄發芽率、發根率與根莖平均長度,7種處理,每瓶放入1個培殖體,重 複5次。
表 7 水黃皮無菌種子苗誘導在 MS、1/2MS 及 WPM 之組合
Table7. Aseptic seedlings induction of P. pinnata in MS 、 1/2MS and WPM compositions
Medium\PGRs Test code
1/2MS 1
MS 2
WPM 3
MS+0.35 mg/L GA3 4
WPM+0.35 mg/L GA3 5
MS+1 mg/L BA 6
WPM+1 mg/L BA 7
3.水黃皮之微體繁殖
(1) 水黃皮芽體誘導試驗 a、無菌苗為培殖體
Purohit 及 Singhvi 於 1998 指出水黃皮的微體繁殖,以 MS 培養基較常被使用,
故以 MS 為基礎培養基添加不同濃度的 TDZ:0.1、1、2、3 mg/L 及 BA:0、0.5、
1、2 mg/L 並另外添加 NAA:0、0.5、1、2 mg/L,並以較佳結果再以 MS 加入 IBA:
0、0.5、1、2 mg/L 的組合誘導芽體,每錐形瓶放入 4 個培殖體,重複 3 次。
b、野外材料
為維持萌蘗活性,材料皆為當天早上約七點採回直接使用,選取健康無病蟲 害之野外萌蘗,以清水沖洗表面並將萌蘗切成約 1 cm 帶腋芽之莖段及葉片,流水
20
處理 2 h 後,進行表面消毒後,以 MS 為基礎培養基添加 0.1 mg/L 及 0.5 mg/L NAA 與 1-5 mg /L BA 的組合,葉片每瓶接種 1 個培殖體,每組做 10 瓶,重複 3 次;側 芽生長點,每瓶放入 1 個培殖體,重複 5 次。誘導芽體結果皆為 1 個月後計算其 芽體數量,並觀察紀錄之。
(2) 水黃皮之芽體伸長試驗
取 3 種培殖體:A 帶節莖段 B 子葉節所誘導的芽體 (將 3-5 個芽體為一個培殖 體)C 生長點作芽體伸長試驗。放入 MS 添加不同濃度的 BA 及 GA3之組合 (表 8) , 使用 125 mL 三角瓶,每瓶裝 50 mL 培養基,每錐形瓶放入 4 個培殖體,重複 3 次,
1 個月後記錄芽體伸長長度。
表 8 水黃皮芽體伸長在 MS 添加 BA 與 GA3之試驗組合
Table 8. Shoot elongation of P. pinnata in MS with different combination of BA and GA3
Medium code
GA3 (mg/L)
0 0.1 0.5 1 1.5
BA(mg/L)
0 BG1 BG2 BG3 BG4 BG5
0.1 BG6 BG7 BG8 BG9 BG10
0.2 BG11 BG12 BG13 BG14 BG15 0.5 BG16 BG17 BG18 BG19 BG20
(3) 水黃皮芽體誘導發根之試驗
以子葉節誘導出之芽體為培殖體,將已抽長約 2-3 cm 的芽體放入 1/2MS 或 MS 添加 IBA 組合 (表 9) 進行發根試驗,以 100 mL 試管分裝,每管含 25 mL 培 養基,共 8 個處理,每瓶放 1 個培殖體,重複 11 次。1 個月後記錄發根數及發根 長度。
21
表 9 水黃皮發根試驗在 MS 或 1/2MS 添加 IBA 之組合
Table 9. Root induction induction of P. pinnata in MS or 1/2MS with compositions of IBA
Medium (strength) + auxin (mg/L) Test code
MS (½ ) 1
MS 2
MS (½ ) + IBA (0.5) 3
MS (½ ) + IBA (1) 4
MS (½ ) + IBA (1.5) 5
MS + IBA (0.5) 6
MS + IBA (1) 7
MS + IBA (1.5) 8
(4) 水黃皮組培苗之馴化
選取發根試驗中已完全展葉且根長達 2-4 cm 植株作出瓶馴化,先以自來水徹 底清洗後再移植到經過高溫高壓殺菌的蛭石/泥炭土/土壤=1:1:1 的混合介質中。
介質保持完全濕潤而不積水,培殖容器置於簡易網室,並將容器以透明塑膠袋罩 住,每 2 週剪去塑膠袋一邊的角落,於第 6 週移除塑膠袋並記錄其存活率及莖平 均長度,共馴化 85 盆植株。
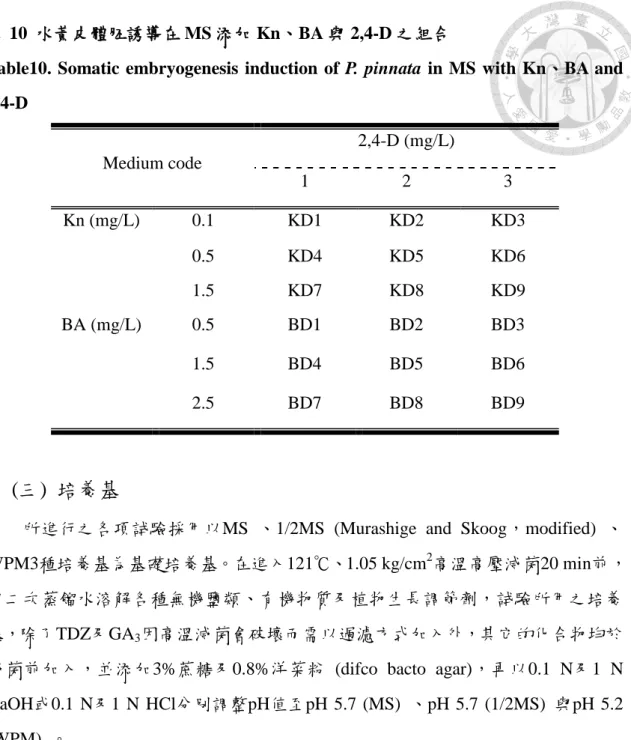
4.水黃皮體胚之誘導
考慮種子為未成熟階段,不宜使用熱水 (60℃) 處理種子,故試驗材料材料皆 為當天早上約七點採回直接使用,將各種發育階段的種子進行表面消毒後,以解 剖刀及鑷子去除種皮,以未成熟胚之帶子葉之胚及半成熟胚之胚及子葉為培殖 體,培養於 Kn 及 BA 與 2,4-D 之組合表 (表 10) 及 TDZ:0.5、1、2 及 3 mg/L 中 進行培養。以 125 mL 三角瓶分裝,每瓶 50 mL,共 22 種處理,每錐形瓶放入 8 個培殖體,重複 3 次,2 個月後記錄誘導結果。
22
表 10 水黃皮體胚誘導在 MS 添加 Kn、BA 與 2,4-D 之組合
Table10. Somatic embryogenesis induction of P. pinnata in MS with Kn、BA and 2,4-D
Medium code
2,4-D (mg/L)
1 2 3
Kn (mg/L) 0.1 KD1 KD2 KD3
0.5 KD4 KD5 KD6
1.5 KD7 KD8 KD9
BA (mg/L) 0.5 BD1 BD2 BD3
1.5 BD4 BD5 BD6
2.5 BD7 BD8 BD9
(三) 培養基
所進行之各項試驗採用以MS 、1/2MS (Murashige and Skoog,modified) 、 WPM3種培養基為基礎培養基。在進入121℃、1.05 kg/cm2高溫高壓滅菌20 min前,
以二次蒸餾水溶解各種無機鹽類、有機物質及植物生長調節劑,試驗所用之培養
基,除了TDZ及GA3因高溫滅菌會破壞而需以過濾方式加入外,其它的化合物均於
滅菌前加入,並添加3%蔗糖及0.8%洋菜粉 (difco bacto agar),再以0.1 N及1 N NaOH或0.1 N及1 N HCl分別調整pH值至pH 5.7 (MS) 、pH 5.7 (1/2MS) 與pH 5.2 (WPM) 。
本試驗所使用的各種無機鹽類、有機物質、植物生長調節劑、洋菜粉及蔗糖 皆為Sigma Chemical (Mo., U.S.A. ) 。
(四) 培養環境
培養室溫度25±1 ℃恆溫,光源為光度50±5 µmole/ m2s之螢光燈 (Philips),光 照16 h;暗處理則培養於培養室之紙箱完全黑暗環境下。
23
(五) 形態觀察
1.以複式顯微鏡觀察外表形態並拍照記錄。
2.石蠟切片及徒手切片。
(1) 石蠟切片
為觀察水黃皮芽體發生之情形,將誘導之芽體埋於蠟塊內,連同蠟塊一起切 片,除去石蠟再經染色和封片而成。製片過程如下 (蔡淑華,1975) :
a、 固定、洗滌:以2.5% Karnovsky固定,固定後用15-50%酒精沖洗約10 min。
b、脫水:以乙醇混合液 (TBA-series) 脫水劑(蒸餾水+乙醇+三級丁醇將 組織細胞內的水份置換、透明化,先用50%浸漬數小時後,更換數次 濃度逐漸提高之TBA,直到100%。
c、浸蠟:依序間隔2 h,將組織依序浸入石蠟。
d、埋蠟:將組織及液狀蠟倒入模型,置於冷水中使之快速冷卻凝固,形 成固體蠟塊。
e、 切片:把多餘的蠟塊剔除,固定在小木塊上,利用切片機切片。
f、 脫水、染色:先用黏附劑張貼切片,將蠟帶用二甲苯溶解後,用高濃 度的酒精逐漸移至染料(Eosin)溶液中進行染色。
(2) 徒手切片
為觀察水黃皮芽體伸長、發根及體胚發生之情形,利用切片直接觀察或染色 觀察,染色處理使用雙重染色,先以Safranin-O染色1 min後,用清水清洗,後使用 Stevenel's Blue染色1 min,再用清水清洗。
(六) 統計分析
1.以完全逢機設計,進行變異數分析。
2.若處理間差異達到顯著水準 (α=0.05) 時,則使用鄧肯氏新多變域檢定法 (duncan’s new multiple range test, DMRT) 比較。
24
圖 1 水黃皮
Fig1. The P. pinnata
圖1-A. 開花期之水黃皮。
Fig1-A. The flowering stage of P. pinnata.
圖1-B. 水黃皮之奇數羽狀複葉。
Fig1-B. The odd-pinnately compound leaf of P. pinnata.
圖1- C. 台灣大學校本部電機二館旁的水黃皮,樹高約4 m。
Fig1-C. The site of four meters high P. pinnata is located beside the second building of the Graduate Institute of Electrical Engineering, National Taiwan University
圖1-C. 水黃皮約花期結束後1個月 (6月)之莢果。
Fig1-C. The pods ripen about one month after flowering stage (in June) of P. pinnata.
圖1-D. 水黃皮約花期結束後2個月(7月)之莢果。
Fig1-D. The pods ripen about two months after flowering stage (in July) of P. pinnata.
圖1-F. 水黃皮萌蘗之枝條。
Fig1-F. The burst from coppice shoots of P. pinnata .
25
1-A
A Fi
Fig A
1-B
A Fi
Fig A
1-F
A Fi
Fig A 1-E
A Fi
Fig A
1-D
A Fi
Fig A 1-C
A Fi
Fig A
26
圖 2 水黃皮種子發育之各階段
Fig2. Different development stage of the seeds of P. pinnata
圖 2-A.水黃皮花期後約 3 個月 (8 月) 成熟之種子。 (Bar = 2 cm)
Fig2-A. The mature seeds of P. pinnata were collected about three months after flowering (in August). (Bar = 2 cm)
圖 2-B.水黃皮未成熟之種子,子葉佔種子內部體積 75%以上。 (Bar = 1 cm) Fig2-B. The immature seeds of P. pinnata with a cotyledon more than 75 percent of the seed volume. (Bar = 1cm)
圖 2-C.水黃皮未成熟之種子,子葉佔種子體積之 50-75% 。 (Bar = 5 mm)
Fig2-C. The immature seeds of P. pinnata with a cotyledon between 50 - 75 percent of the seed volume. (Bar=5 mm)
圖 2-D.水黃皮未成熟之種子,子葉佔種子體積 50%以下。(Bar = 5 mm)
Fig2-D. The immature seeds of P. pinnata with a cotyledon less than 50 percent of the seed volume. (Bar = 5 mm)
圖 2-E.水黃皮花期後約 2 個月 (7 月) 未成熟之種子。 (Bar = 1 cm)
Fig2-E. The immature seeds of P. pinnata were collected about two months after flowering (in July). (Bar=1cm)
27
1 cm 2 cm
1 cm
5 mm 5 mm
2-B
A Fi
Fig A 2-A
A Fi
Fig A
2-E
A Fi
Fig A
2-D
A Fi
Fig A 2-C
A Fi
Fig A
28
肆、結果 (Results)
一、水黃皮野外帶節莖段之表面消毒
帶節莖段經培養約 5-7 天為發霉的最高峰,根據蔡國書 (2005) 在大葉桉 (Eucalyptus robusta) 表面消毒之培殖體消毒後 10 天內會因消毒不完全而發生霉 變,10 天後發霉可能是因封瓶不夠緊密所造成,故污染率計算應以第 10 天計算。
本試驗利用不同濃度NaOCl(1-5%)配合不同消毒時間 (10或20 min) 處理消 毒水黃皮帶節莖段,調查其存活率及污染率。帶節莖段培養約1週後,可觀察培殖 體周圍與其接觸之培養基邊緣,開始出現汙染之現象,通常有發霉及乳白色或黃 色的分泌物感染較為常見,培養中期 (4、5天) 汙染多屬前者,培養末期 (7、8天) 甚至培養10天以上,多出現後者之感染徵狀。
試驗結果自野外採回之材料,經表面消毒後污染率仍高 (表 11),試驗中所測 試之處理時間,並無法達到正面之消毒成效,延長處理時間比不上提高 NaOCl 濃 度效果好,單就討論消毒時間而論,都是消毒 20 min 的污染率低於消毒 10 min (圖 3) ,但在統計上並無差異性。NaOCl 濃度 5%消毒 20 min 污染率較低 (64.76%) , 污染的時間多發生在培養末期,消毒時間與 NaOCl 濃度越大,帶節莖段兩端白化 的範圍也會增加,於操作台中修切時易造成較大的傷口,褐化率較高;1% NaOCl 消毒 10 min 污染率較高,並於培養初期易造成全面性的污染狀況,褐化程度較輕 微。
培殖體誘導結果都不高,1%NaOCl 消毒10 min有較高的誘導率 (22%) ,但污 染也高,誘導多為側芽,而NaOCl濃度以5%消毒20 min污染率雖低,卻會降低誘 導率,無芽體誘導之帶節莖段多褐化死亡,或產生微量乳黃色癒合組織,且癒合 組織並無進一步產生逆分化。
29
表 11 水黃皮野外帶節莖段表面消毒及芽體誘導之結果
Table 11. The results of the nodal stems surface sterilization and shoot induction of P. pinnata
Test code
NaOCl (%)
Time (min)
Sterilized numbers
Sterilized rate (%)
Induction number of explants
Shoot Induction(%) A1 1 10 23 22 ± 7.67cd 5 21.74
A2 1 20 35 33.29 ± 9.62bcd 0 0
A3 2 10 16 15.14 ± 4.9d 2 12.5 A4 2 20 51 48.43 ± 6.14ab 3 5.88
A5 3 10 24 22.86 ± 2.48 bcd 0 0
A6 3 20 35 33.57 ± 7.52bcd 2 5.71 A7 4 10 27 25.71 ± 6.03 bcd 0 0 A8 4 20 38 36.29 ± 4.63bc 2 5.26
A9 5 10 44 41.86 ± 4.88ab 0 0
A10 5 20 68 64.71 ± 2.8a 4 5.88
※ Mean ± SE
※ Different letters in the same row show that the values are significantly different by DMRT (α=0.05)
圖 3 水黃皮帶節莖段未受汙染之結果
Fig 3. The results of the the nodal stems with sterilized rate of P. pinnata
30
二、水黃皮無菌種子苗之培養與生長
此試驗目的為培養健康生長之無菌種子苗,提供微體繁殖的材料,本次試驗 採 MS、1/2MS 及 WPM 為基礎培養基,再以 MS 及 WPM 培養基添加 1 mg/L BA 或 0.35 mg/L GA3探討無菌種子苗培養情形,結果在發芽方面,以 WPM 添加 1 mg/L BA 可達高約 79.97%的發芽率,其次為 MS 添加 1 mg/L BA (79.74%) ,發芽率較 低分別為 WPM、MS 及 1/2MS,發芽率依序為 42.75%、36.38%與 33.55% (表 12) , 但在種子苗莖部生長狀況則以 WPM 培養基較良好,莖部肥大,葉片茂盛、顏色呈 現墨綠色 (圖 4-A) ;而添加 BA 植物生長調節劑處理結果,於種子苗子葉節位置 誘導 2-3 根側芽,其生長狀況較細且葉片較小 (圖 4-B) ;添加 GA3處理結果,莖 段更為細長,以 WPM 添加 GA3為例莖段長達 5.56 cm,且葉片較小、顏色淡 (圖 4-C) ,小苗也較容易掉葉。
根部生長情形,則以 WPM 培養基及添加 GA3處理下,種子苗之主根明顯,
長度約 7-8 cm,側根茂盛 (圖 4-D、E) ;BA 處理下,主根不明顯,長度僅上述根 之一半 (3 cm) ,側根較少發生 (圖 4-F)。
31
表 12 水黃皮無菌種子苗誘導與培養 2 個月之枝條生長結果
Table 12. The results of shoot growth of P. pinnata from aseptic seeds as induced and cultured in vitro for two months
Medium Number of explants
Germination rate (%)
Mean length of root (cm)
Mean length of shoot (cm) 1/2MS 120 33.55±2.47c 8.52±1.18a 2.44±0.7bc
MS 120 36.38±1.85c 7.41±1.5a 1.4±0.62c WPM 120 42.75±6.52c 8.26±0.8a 2.8±1.17bc MS+0.35 mg/L GA3 120 46.41±5.44c 7.36±0.29a 3.9±0.41ab WPM+0.35 mg/L GA3 120 54.17±6.85bc 9.38±0.33a 5.56±0.83a MS+1 mg/L BA 120 79.74±6.76a 3.29±0.14b 3.09±0.07bc WPM+1 mg/L BA 120 79.97±5.59ab 3.2±0.15b 3.97±0.13ab
※ Mean ± SE
※ Different letters in the same row show that the values are significantly different by DMRT (α=0.05)
※ The number of repeat:5
32
圖 4 水黃皮之種子苗培養 2 個月
Fig4. Aseptic seedling of P. pinnata cultured for two months
圖 4-A. 水黃皮種子苗在 WPM 培養基中莖之形態。 (Ba r = 1 cm)
Fig4-A. The morphology of stem from aseptic seeds of P. pinnata cultured on WPM medium. (Bar = 1cm)
圖 4-B. 水黃皮種子苗在 WPM 添加 0.35 mg/L GA3中莖之形態。 (Bar = 2 cm) Fig4-B. The morphology of stem from aseptic seeds of P. pinnata cultured on WPM with 0.35 mg/L GA3 treatment. (Bar = 2 cm)
圖 4-C. 水黃皮種子苗在 WPM 添加 1 mg/L BA 中莖之形態。 (Bar = 1 cm)
Fig4-C. The morphology of stem from aseptic seeds of P. pinnata cultured on WPM with 1 mg/L BA treatment. (Bar = 1 cm)
圖 4-D. 水黃皮種子苗在 WPM 培養基中根之形態。 (Bar = 1 cm)
Fig4-D. The morphology of root from aseptic seeds of P. pinnata cultured on WPM medium. (Bar = 1cm)
圖 4-E. 水黃皮種子苗在 WPM 添加 0.35 mg/L GA3中根之形態。 (Bar = 1 cm) Fig4-E. The morphology of root from aseptic seeds of P. pinnata cultured on WPM with 0.35 mg/L GA3 treatment. (Bar = 1 cm)
圖 4-F. 水黃皮種子苗在 WPM 添加 1 mg/L BA 中根之形態。 (Bar = 1 cm)
Fig4-F. The morphology of root from aseptic seeds of P. pinnata cultured on WPM with 1 mg/L BA treatment. (Bar = 1 cm)
33
1 cm
1 cm 1 cm
1 cm 1 cm
2 cm 4-B
A Fi
Fig A 4-A
A Fi
Fig A
4-E
A Fi
Fig A
4-F
A Fi
Fig A 4-D
A Fi
Fig A 4-C
A Fi
Fig A
34
三、水黃皮之芽體誘導試驗 (一) 種子苗材料
經無菌培養試驗,選生長及發芽率較佳之 WPM 作為基礎培養基,或添加 BA
與 GA3兩種生長調節劑,取其作為培殖體誘導之芽體,探討經此 3 種不同培養基
之種子苗培養之培殖體於芽體誘導下彼此間之影響。
1.培殖體部位與芽體發生之關係
無菌種子苗長至0.5-1.0 cm或3-5 cm時,各別取子葉節及下胚軸或莖段和帶節 莖段進行培養,子葉節與帶節莖段之芽體誘導率最高皆可達100%,然莖段及下胚 軸培殖體誘導率僅20%以下,莖段及下胚軸多誘導出癒合組織,僅少數由培殖體表 面產生芽體;子葉節自子葉間處產生芽體;而帶節莖段部分,芽體或源於原本存 在於葉腋之潛伏芽、或自節間處產生。
2.前處理間的差異
3種前處理 (WPM、WPM+1 mg/L BA及WPM+0.35 mg/L GA3) ,帶節莖段之 芽體形態並無太大差異 (圖5-A、B、C) ;子葉節誘導芽體形態以WPM及GA3處理 結果之芽體數較少但較長 (圖5-D、E) (表13、14),BA處理之芽體較多但較短 (圖 5-F) ;芽體數誘導結果於3種前處理下雖無顯著差異,但以BA前處理結果芽體數 較高 (圖6) 。癒合組織形成之形態和誘導率與前處理間之差異亦不明顯,下胚軸 及莖段為培殖體主要產生癒合組織,僅少數芽體產生,WPM前處理下皆有芽體產 生,GA3及BA前處理僅莖段部分有芽體產生。空白試驗 (對照組) 在BA前處理下,
僅帶節莖段有誘導出少量之芽體 (33.33%) 。 3.各植物生長調節劑誘導結果
(1) BA 與 NAA 不同濃度組合誘導芽體結果
試驗結果,子葉節與帶節莖段都以 BA 前處理下添加 2 mg/L BA 及 0.1 mg/L NAA 的誘導下結果較佳 (表 15) ,且平均芽體數隨 BA 濃度增加而提高 (0-2 mg/L) ,當 BA 濃度高於 2 mg/L 時,芽體數達最大值且趨勢線趨於平緩 (圖 7) 。 子葉節誘導芽體數較高,平均為 5.42 個芽,單一培殖體最高芽體數 15 個,且芽體 發生於子葉節之分生組織,其次為帶節莖段 (平均 3.17 個芽) ,單一培殖體最高芽
35
體數為 7 個,芽體多自側芽誘導出其他芽體;莖段與下胚軸,於 GA3前處理以莖
段為培殖體添加 0.5 mg/L BA 及 1 mg/L NAA 平均誘導 0.25 個芽體數 (誘導率 13.33%) (圖 8-A) 及 WPM 前處理下添加 2 mg/L BA 及 2 mg/L NAA 平均誘導 0.83 個芽體數 (誘導率 20%) (圖 8-B) ;下胚軸僅在 WPM 前處理下,添加 2 mg/L BA 及 0.1 mg/L NAA 可平均誘導 0.33 個芽體數 (誘導率 8.33%) (圖 8-C) 。
(2) BA 與 IBA 不同濃度組合誘導芽體之結果
鑑於 BA 添加 NAA 與各前處理誘導結果,BA 前處理芽體數較高,故於此階 段試驗以 BA 前處理為主。BA 添加與 NAA 相同濃度之 IBA 誘導結果,以子葉節 培殖體較佳配方為 2 mg/L BA 添加 0.5 mg/L IBA,平均誘導 4.33 個芽體;帶節莖 段平均誘導 3.42 個芽體 (表 16) ;莖段與下胚軸無任何芽體產生,僅癒合組織發 生。
(3) TDZ 不同濃度誘導芽體結果
試驗結果,子葉節與帶節莖段皆以 BA 前處理下添加 0.1 mg/L TDZ 的誘導下 結果較佳 (表 17) ,子葉節平均誘導 3.86 個芽體,隨 TDZ 濃度增加 (0.1-3 mg/L) , 不僅降低芽體數,並於子葉節切口處伴隨更多之癒合組織;帶節莖段平均誘導 2.5 個芽體;0.1 mg/L TDZ 以莖段為培殖體以 WPM 前處理下平均產生 4.5 個芽體 (誘 導率 13.33%) (圖 8-D) ,GA3前處理可誘導平均 3 個芽體 (誘導率 13.33%) (圖 8-E、
F) ;下胚軸為培殖體在 1 mg/L TDZ 誘導下以 BA 前處理可產生平均 2 個芽體 (誘 導率 6.67%) (圖 8-G),WPM 前則誘導平均 3 個芽體 (誘導率 13.33%) (圖 8-H)。
試驗觀察中發現,不添加任何植物生長調節劑,帶節莖段僅經過 BA 前處理下 有芽體誘導;子葉節無任何生長反應;下胚軸及莖段則於傷口處有褐化及黑化的 現象,亦無芽體或癒合組織的產生。
IBA 或 NAA 與 BA 交叉組合下誘導芽體或癒合組織形態上無太大差異,芽體 誘導數以 BA 和 NAA 組合結果較 IBA 高;子葉節和帶節莖段培殖體與培養基接觸 之處會有癒合組織發生;下胚軸與莖段培殖體之癒合組織則形成自上方之一面,
或自破脹之表皮處,使整個培殖體狀似鼓槌 (圖 8-I、J) ,故發現,水黃皮莖段與 下胚殖培殖體,雖平躺於培養基上,其生理反應仍具極性。癒合組織呈黃色且形 質鬆軟,部分子葉節培殖體癒合組織之表層會形成一層白色絨毛狀之細胞,當提