1
中文摘要
AP1 訊息傳導路徑和細胞的增生(proliferation)、轉化(transformation)以及細胞的程 式性死亡(apoptosis)有極大的相關性。探討鼻咽癌組織中 AP1 轉錄因子的表現情形有相 當的重要性:第一,放射線治療〈或化學治療〉主要乃是引起腫瘤細胞的程式性死亡。 而鼻咽癌的主要治療是放射線治療,或併用化學治療。第二,EB 病毒之致癌基因 LMP-1 已被證實會提高 AP1 的轉錄能力。此轉錄能力的上升,是否有程式性死亡調控的角色, 仍有待探索。在第一年的計劃中。我們發現了鼻咽癌組織中 AP-1 訊息傳導路徑的表現, 確實和病患接受放射線治療的預後有明確的關聯性。在鼻咽癌組織中,c-Jun 持續被活 化的病患,相較於腫瘤組織中 c-Jun 未被持續被活化的病患,有較佳的預後,且呈現統 計學上顯著差異。我們同時也用 Immunoprecipitation 的方法,證實了組織免疫染色的專 一性及可靠性。在第二年的計劃中,我們嘗試建立一個 in vitro model,來探討 AP-1 的表現與鼻咽 癌細胞接受放射線治療敏感性的關係。我們以帶有 LMP1 基因的 EBV-based vector,轉 殖到鼻咽癌細胞株 HNOE-1 中,試圖建立一個研究鼻咽癌的細胞模型。在篩選過程中, 選出的 stable clone 卻無法表現出明顯的 LMP1 蛋白質。但有趣的是,相較於 HNOE-1 母細胞株,僅表現 EBNA1 蛋白質的 vector-control 細胞株,不但生長速率較 HNOE-1 母 細胞株緩慢,也對 cisplatin 呈現極佳的敏感性。此外,對於較低劑量的放射線治療, vector-control 細胞株似乎也有較佳的敏感性。其可能機制與 AP-1 轉錄因子表現的相關 性,目前正進一步探討中。
2
英文摘要
AP-1 activates or represses the expression of certain target genes, which are mainly involved in cellular proliferation, transformation and apoptosis. The biological significance of AP-1 pathway deserves intensive study in NPC for two reasons. Firstly, in NPC, it is demonstrated that AP-1 transcriptional activity is up-regulated through JNK activation by the oncogenic LMP-1 (latent membrane protein –1) protein of EBV (Epstein-Barr virus). Secondly, the mainstay treatment of NPC is radiotherapy (or combined with chemotherapy), and it is generally believed that cancer treatment modalities, such as radiotherapy and chemotherapy, exert their effects mainly by induction of cellular apoptosis. In the first year of this study, we have demonstrated that AP-1 (c-jun) pathway is constitutively activated in more than half of the NPC specimens used in this study. Interestingly, persistent activation of AP-1 (c-jun) pathway correlates with better prognosis in NPC patients after radiotherapy. We are now establishing a cell mode to exploring the biological significance of persistent AP-1 activation in NPC.
In an effort to establishing an in vitro model, we introduce a hygromycin-resistant, LMP1-expressing EBV vector into HONE-1 cells to pick up stable clones. However, the selected stable clones failed to express detectable LMP1 protein. But interestingly, EBNA1-expressing vector-control clones retard the growth rate of parental HONE-1 cell line and sensitizes parental HONE-1 cells to cisplatin and low-dose irradiation. The detail mechanism and possible relationship to AP-1 expression is currently under extensive investigation.
3
結案報告內容
A. 研究背景及文獻探討AP-1(activator protein-1)轉錄因子乃是由 Jun 家族(包括 c-jun、junB、junD)或是加 上 fos 家族(包括 c-fos、fosB、Fra1、Fra2)兩兩結合而成。研究顯示,AP1 訊息傳導 路徑和細胞的增生(proliferation)、轉化(transformation)以及細胞的有極大的相關性1。 不同的 AP-1 家族成員結合,可能調控不同的基因,而影響細胞的功能2。不同的腫 瘤,可能使用不同的 AP-1 轉錄因子3, 4。同一組織或腫瘤不同的發展時期,也可能使 用不同的 AP-1 轉錄因子5-8。不同的 AP-1 轉錄因子也被證實和乳癌細胞的某些表現 型態有關9。甚至在某些情況下,AP-1 組成因子在功能上有互補的作用10, 11。 證據顯示,AP1 訊息傳導路徑導致細胞增生主要是來自於壓制抑癌基因(tumor suppressor gene)及促進細胞週期有關的基因(cell cycle regulator)1。AP-1 被證實在 NIH 3T3 細胞被 ras-轉化的過程中,扮演重要的角色12。而 EB 病毒之致癌基因 LMP-1(latent membrane protein-1)的轉化能力,也被認為和 LMP-1 能夠經由活化 JNK1 來提高 AP1
的轉錄能力有關13。另一方面,抑制 AP-1 訊息傳導路徑將導致多個訊息傳導路徑同 時被抑制,使得乳癌細胞停止生長14。這些證據顯示 AP-1 訊息傳導路徑和細胞的增 生(proliferation)和轉化(transformation)有極密切的關係。 有趣的是,AP-1 在細胞程式性死亡的調控上,也可能扮演重要角色。已知許多 外界來的刺激〈如放射線和化學治療藥物〉所引起的細胞程式性死亡,都同時伴隨 著 AP-1 轉錄能力的升高15-18。抑制NFκB訊息傳導路徑,會使得肝細胞對 TNF-α的 處理,特別容易進行細胞程式性死亡。此細胞程式性死亡,可以歸因於 JNK、c-Jun 的持續活化,以及 cytochrome c、細胞程式性死亡蛋白質釋放19。在人類胰臟癌組織 中,也發現 AP-1 和細胞程式性死亡的執行蛋白質 caspase3 有一致的表現20。AP-1
4 B. 研究方法 第一年:鼻咽癌組織中 AP-1 訊息傳導路徑的研究。 *第一部份:免疫組織染色法的研究 首先選取具有預後資料(prognosis data)的鼻咽癌病患治療前之鼻咽切片石 蠟包埋組織,進行免疫組織染色法的研究。在同一檢體上,實行 c-Jun 的免疫 組織染色。所得結果經判讀後,與病患之臨床資料作關聯性的探討,看不同 的 AP-1 蛋白質表現,是否與病患之治療預後有所相關。 * Immunohistochemical study
Serial 5-μm histologicalsectionswerecut, mounted on glass slides coated with 3-aminopropyltriethoxysilane, and air-dried overnight at room temperature. The sections were then deparaffinized in xylene and rehydrated in ethanol. Haematoxyline and eosin (H&E) stain was first performed in each specimen to confirm the presence of tumor cells. Endogenous peroxidase activity was then blocked by using methanol containing 3% H2O2 for 15 minutes. For all sections used in this study, antigen retrieval procedure was performed by immersing the slides in citrate buffer (PH 6.0) and then heating the slides in a microwave oven for10 minutes. The sections were then incubated with primary antibodies by using appropriate dilution for each anti-AP-1 Ab (anti-c-Jun, JunB, JunD, c-Fos, FosB, Fra1 and Fra2, all obtained from Santa Cruz) at 4℃ overnight, followed by staining with an Universal Immuno-peroxidase polymer (UIP) solution (Simple Stain MAX PO MULTI, Nichirei, Tokyo, Japan) for 30 minutes. The sections were finally reacted with an AEC (3-amino-9-ethylcarbazole) substrate solution (DAKO, Glostrup, Denmark) and then counterstained with haematoxylin before being mounted. Non-human reactive rabbit polyclonal IgG (DAKO, Glostrup, Denmark) was used as isotype negative controls.
*第二部份:AP-1 蛋白質的研究
此部份研究分為兩項:第一項乃是萃取冷凍鼻咽切片之蛋白質,並以西 方轉漬法(western blot)研究鼻咽癌組織中 AP-1 蛋白質的表現,並與免疫組織 染色法所得的結果互相印證。其次則是以 immunoprecipitation 的方法,證實 免疫組織染色法所使用之 c-Jun 抗體的專一性。
* Immunoprecipitation
5
and elute the immune complex. Add 20μl of the elute to a microcentrifuge tube. Equilibrate Sample Buffer to room temperature. Gently mix the Sample Buffer by inverting 5-10 times. Pipette 5 μl of the 5X Sample Buffer into the microcentrifuge tube. Heat the sample at 95-100°C for ~5 minutes. Allow the sample to cool to room temperature and apply to the gel for electrophoresis.
第二年:AP-1 訊息傳導路徑對鼻咽癌細胞的影響。
*第一部份: 建立一鼻咽癌研究的細胞模型
Construction of LMP1-expressing plasmid
In order to mimic the conditions of EBV latency, we used an EBV-based vector (pCEP4, Invitrogen, Carlsbad, CA, USA) to express LMP1 in HONE-1 cells. This vector contains the ori-P segment of EBV DNA and constitutively expresses the EBNA-1 protein, which is mandatory in maintaining episomal DNA with host chromosomes. Thus, this EBV-based vector can maintain an episomal existence in mammalian cells, just like EBV. LMP1 cDNA was purified and then ligated into a pCEP4 vector. The ligated LMP1-pCEP4 plasmid was purified and confirmed by direct sequencing.
Transfection and selection of LMP1-expressing HONE-1 cell clones
Transfection was performed by lipofectamine. The procedure was performed according to the manufacturer’srecommendations (Invitrogen). Briefly, DNA-liposome complex was prepared by mixing sol’n A (1μg ofplasmid DNA constructin 100μlof serum-free medium)and sol’n B (6μloflipofectaminediluted into 100μlofserum-free medium). The prepared DNA-liposome complex was incubated at room temperature for 30 min and then diluted with 0.8 ml serum-free medium. The diluted DNA-liposome complex was used to overlay 1×105 adherent HONE-1 cells in a 35-mm culture dish. After incubation for 5 hrs, 1 ml growth medium containing 20% FCS was added without removing the transfection mixture. The growth medium was refreshed 24 hrs after the start of transfection. At 72 hrs after transfection, cells were subcultured and selected in growth medium containing 400 μg/ml hygromycin (Invitrogen). Hygromycin-resistant clones were then selected and expanded for later usage. Vector-transfected HONE-1 clones were used as a control.
*第二部份: AP-1 訊息傳導路徑對鼻咽癌細胞治療抗性的影響
6 Growth curve determination
The growth curve of parental HONE-1 cells, LMP1-transfected HONE1 clones and vector-control HONE-1 cloneswere determined by standard protocol. Briefly, cells in mid-exponential growth were trypsinized and harvested in single cell suspension. After cell counting, 104 cells were plated into twelve 10-cm dishes for each cell clone. The cell numbers were counted for 9 consecutive days for each cell clone. Experiment was done in triplicate.
Clonogenic survival assay of cisplatin and radiation toxicity
Radiation and cisplatin toxicity studies were performed using a standard clonogenic survival assay. Cells in mid-exponential growth were trypsinized and harvested in single cell suspension. After cell counting, 500 cells were plated into individual 10-mm dishes. The plating efficiency was determined in preliminary experiments. After overnight incubation, the dishes were irradiated from 1 to 9 Gy using a cobalt-60 source or incubated with 2 to 12 μg/ml cisplatin (Platinol-AQ, Bristol-Myers Squibb, New York, NY, USA) for 1 hour. After a 10-day incubation, the plates were fixed with methanol/acetic acid (3:1), stained with crystal violet and observed under a dissecting microscope. Colonies larger than 50 cells were counted. Survival fraction was calculated by dividing colony numbers of treated plates by colony numbers of control plate. The survival fraction was determined in triplicate at all doses for each cell clone.
7
第二年:AP-1 訊息傳導路徑對鼻咽癌細胞的影響。
a. 建立一鼻咽癌研究的細胞模型
在此部份中,我們使用了 LMP1 expressing-EBV vector,嘗試把它轉殖到鼻 咽癌細胞株 HNOE-1 中,來建立一個研究鼻咽癌的細胞模型。然而,雖然我們 可以篩選出 stable clones(L1, L8),但 L1, L8 卻無明顯 LMP1 蛋白質或 EBNA1 蛋 白質表現的細胞株(圖 4)。
b. AP-1 訊息傳導路徑對鼻咽癌細胞治療抗性的影響
8
參考文獻
1. Shaulian E, Karin M. AP-1 as a regulator of cell life and death. Nat Cell Biol 2002;4(5):E131-6.
2. Bakiri L, Matsuo K, Wisniewska M, Wagner EF, Yaniv M. Promoter specificity and biological activity of tethered AP-1 dimers. Mol Cell Biol 2002;22(13):4952-64. 3. Mao X, Orchard G, Lillington DM, Russell-Jones R, Young BD, Whittaker SJ.
Amplification and overexpression of JUNB is associated with primary cutaneous T-cell lymphomas. Blood 2003;101(4):1513-9.
4. Mathas S, Hinz M, Anagnostopoulos I, et al. Aberrantly expressed c-Jun and JunB are a hallmark of Hodgkin lymphoma cells, stimulate proliferation and synergize with NF-kappa B. Embo J 2002;21(15):4104-13.
5. Vuong H, Patterson T, Adiseshaiah P, Shapiro P, Kalvakolanu DV, Reddy SP. JNK1 and AP-1 regulate PMA-inducible squamous differentiation marker expression in Clara-like H441 cells. Am J Physiol Lung Cell Mol Physiol 2002;282(2):L215-25.
6. Sheerin A, Thompson KS, Goyns MH. Altered composition of the AP-1 transcription factor in immortalized compared to normal proliferating cells. Cancer Lett 2002;177(1):83-7. 7. Serewko MM, Popa C, Dahler AL, et al. Alterations in gene expression and activity during
squamous cell carcinoma development. Cancer Res 2002;62(13):3759-65.
8. Andreucci JJ, Grant D, Cox DM, et al. Composition and function of AP-1 transcription complexes during muscle cell differentiation. J Biol Chem 2002;277(19):16426-32. 9. Bamberger AM, Methner C, Lisboa BW, et al. Expression pattern of the AP-1 family in
breast cancer: association of fosB expression with a well-differentiated, receptor-positive tumor phenotype. Int J Cancer 1999;84(5):533-8.
10. Passegue E, Jochum W, Behrens A, Ricci R, Wagner EF. JunB can substitute for Jun in mouse development and cell proliferation. Nat Genet 2002;30(2):158-66.
11. Wenzel A, Iseli HP, Fleischmann A, et al. Fra-1 substitutes for c-Fos in AP-1-mediated signal transduction in retinal apoptosis. J Neurochem 2002;80(6):1089-94.
12. Mechta F, Lallemand D, Pfarr CM, Yaniv M. Transformation by ras modifies AP1 composition and activity. Oncogene 1997;14(7):837-47.
13. Kieser A, Kilger E, Gires O, Ueffing M, Kolch W, Hammerschmidt W. Epstein-Barr virus latent membrane protein-1 triggers AP-1 activity via the c-Jun N-terminal kinase cascade.
Embo J 1997;16(21):6478-85.
14. Lu A, Zhang F, Gupta A, Liu J. Blockade of AP1 Transactivation Abrogates the Abnormal Expression of Breast Cancer-specific Gene 1 in Breast Cancer Cells. J Biol Chem
2002;277(35):31364-72.
15. Wanner R, Henseleit-Walter U, Wittig B, Kolde G. Proliferation-dependent induction of apoptosis by the retinoid CD437 in p53-mutated keratinocytes. J Mol Med 2002;80(1):61-7. 16. Takahashi K, Inanami O, Hayashi M, Kuwabara M. Protein synthesis-dependent apoptotic
9
2002;78(2):115-24.
17. Silvers AL, Bowden GT. UVA irradiation-induced activation of activator protein-1 is correlated with induced expression of AP-1 family members in the human keratinocyte cell line HaCaT. Photochem Photobiol 2002;75(3):302-10.
18. Li T, Dai W, Lu L. Ultraviolet-induced junD activation and apoptosis in myeloblastic leukemia ML-1 cells. J Biol Chem 2002;277(36):32668-76.
19. Liu H, Lo CR, Czaja MJ. NF-kappaB inhibition sensitizes hepatocytes to TNF-induced apoptosis through a sustained activation of JNK and c-Jun. Hepatology 2002;35(4):772-8. 20. Meggiato T, Calabrese F, De Cesare CM, Baliello E, Valente M, Del Favero G. C-JUN
and CPP32 (CASPASE 3) in human pancreatic cancer: relation to cell proliferation and death. Pancreas 2003;26(1):65-70.
21. Koo MS, Kwo YG, Park JH, Choi WJ, Billiar TR, Kim YM. Signaling and function of caspase and c-jun N-terminal kinase in cisplatin-induced apoptosis. Mol Cells
2002;13(2):194-201.
計劃成果自評
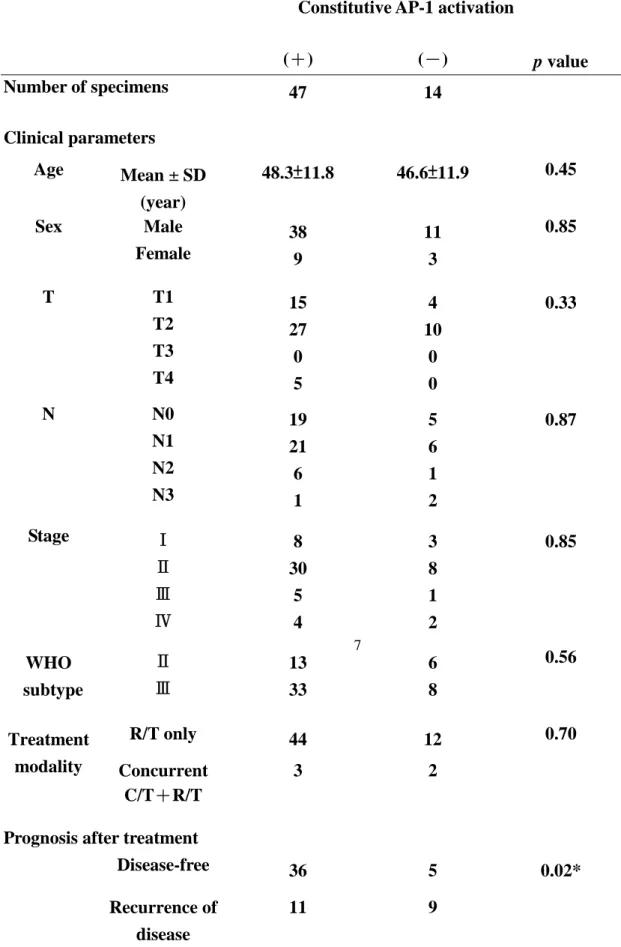
10 表 1. AP-1 的染色結果及病患臨床數據的相關性
Constitutive AP-1 activation
(+) (-) p value Number of specimens 47 14 Clinical parameters Age Mean ± SD (year) 48.3±11.8 46.6±11.9 0.45 Male 38 11 Sex Female 9 3 0.85 T1 15 4 T2 27 10 T3 0 0 T T4 5 0 0.33 N0 19 5 N1 21 6 N2 6 1 N N3 1 2 0.87 Stage Ⅰ 8 3 Ⅱ 30 8 Ⅲ 5 1 Ⅳ 4 2 0.85 Ⅱ 13 6 WHO subtype Ⅲ 33 8 0.56 R/T only 44 12 Treatment modality Concurrent C/T+R/T 3 2 0.70
Prognosis after treatment
Disease-free 36 5
Recurrence of disease
11 9
0.02*
a:(T1+T2 vs. T3+T4), b: (N0+N1 vs. N2+N3), c: (Stage I + II vs. Stage III + IV) * Statistically significant (p<0.05)
11
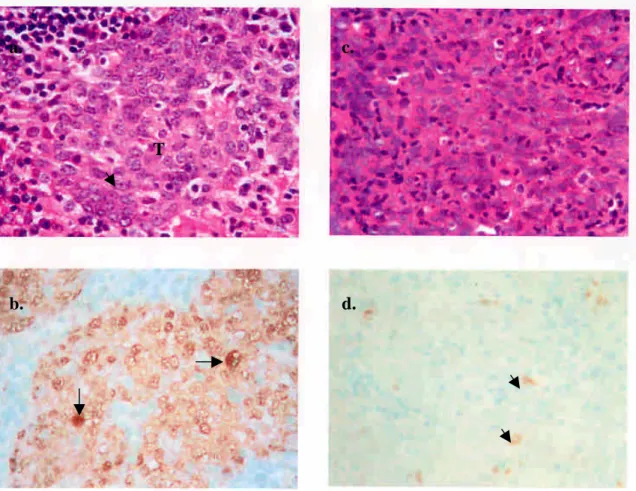
圖 1. 以免疫染色法偵測 AP-1 在鼻咽癌組織的表現情形
FIGURE 1. Staining of AP-1 on archival NPC specimens. (a) H&E stain. Large, vesicular
nucleus with prominent nucleolus (arrow) is noted in majority of the tumor cells (T). (b) same patient as in (a), showing strong intra-nuclear staining of AP-1 (arrows). (c) H&E stain from another patient. (d) same patient as in (c), showing occasional nuclear staining of AP-1 in tumor cells (short arrows) (original magnification × 400).
a.
b.
c.
12 圖 2. AP-1 在鼻咽癌組織的表現情形
2a. 鼻咽癌組織中表現出大量被活化(磷酸化)的 p-c-jun 蛋白質
HONE-1 cell lysate NPC tissue lysate
2b. 先用 anti-p-c-jun Ab 作 immunoprecipitation,再用另一個 polyclonal anti-c-jun Ab 來證實 anti-p-c-jun Ab 的專一性
13 圖 3. AP-1 表現與否和病患存活分析的關連性 3a. 無局部及頸部復發存活率分析 3b. 無遠處轉移存活率分析 0.000 0.250 0.500 0.750 1.000 0.0 40.0 80.0 120.0 160.0 200.0
LR-FREE SURVIVAL (months)
S U R V IV A L P R O B A B IL IT Y 0.000 0.250 0.500 0.750 1.000 0.0 40.0 80.0 120.0 160.0 200.0
META-FREE SURVIVAL (months)
S U R V IV A L P R O B A B IL IT Y 3c. 無病存活率分析 0.000 0.250 0.500 0.750 1.000 0.0 40.0 80.0 120.0 160.0 200.0
DISEASE-FREE SURVIVAL (months)
S U R V IV A L P R O B A B IL IT Y 3d. 總存活率分析 0.000 0.250 0.500 0.750 1.000 0.0 32.0 64.0 96.0 128.0 160.0
OVERALL SURVIVAL (months)
14
圖 4. 以 LMP1-expressing-EBV based vector transfect 到 HONE-1 細胞,其 stable clone L1, L8 並無 EBNA1 及 LMP1 表現 a. B95-8 HONE-1 L1 L1 L8 V1 V2 30 ug 50 ug 70 ug 50 ug 50 ug 50 ug 50 ug b. B95-8 HONE-1 L1 L8 B95-8 V1 V2 50 ug 750 ug 750 ug 750 ug 50 ug 50 ug 50 ug
FIGURE 4. Lack of LMP1 expression in LMP1-transfected HONE-1 cell clones (L1, L8) in
western blot. (a) Upper panel, EBNA1 protein can be detected in vector-control V1, V2 clones but not in LMP1-transfected HONE-1 cell clones (L1, L8). Lower panel, no LMP1 expression is detectable in L1, L8 cells. B95-8 cell lysate was used as positive control for both EBNA1 (~ 72 KD) and LMP1 (~ 65 KD) protein. (b) No EBNA1 or LMP1 protein could be detected in L1, L8 clones despite 750 ug protein lysate was used for western blot.
-15
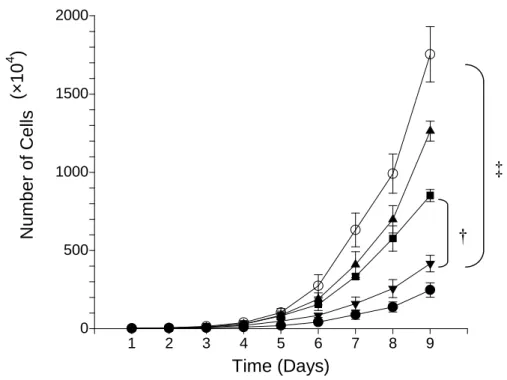
圖 4. EBNA1-expressing vector-control cell lines (V1, V2)和 parental HONE-1 cell line 的 生長情形 0 500 1000 1500 2000 1 2 3 4 5 6 7 8 9 Time (Days) N u m b e r o f C e lls
FIGURE 11. Growth curves for different cell clones. ○ parental HONE-1 cell, ▲ L1 cell clone (LMP1-transfected HONE-1 cells,clone-1), ■ L8 cell clone (LMP1-transfected HONE-1 cells,clone-8), ▼ V1 cell clone (vector-control cells, clone-1), ● V2 cell clone
(vector-control cells, clone-2). Bars indicate standard error of mean. †
p = 0.0026,‡p=0.0019, two-sample t-test, day 9.
16
圖 5. EBNA1-expressing vector-control cell lines (V1, V2)和 parental HONE-1 cell line 對 化學及放射線治療敏感性的探討 0 20 40 60 80 100 120 2 4 6 8 10 12
Cisplatin Dosage (ug/ml)
S u rv iv a l F ra c ti o n 0.0 25.0 50.0 75.0 100.0 1 3 5 7 9 R/T dose (Gy) S u rv iv a l F ra c ti o n
FIGURE 5. Clonogenic survival assay of cisplatin (CDDP) and irradiation toxicity for
different cell clones. ○ HONE-1 cell, ▲ L1 cell clone (LMP1-transfected HONE-1 cells,
clone-1), ■ L8 cell clone (LMP1-transfected HONE-1 cells,clone-8), ▼ V1 cell clone (vector-control cells, clone-1), ● V2 cell clone (vector-control cells, clone-2). Bars indicate
standard error of mean. Please note that parental HONE-1 cells, clone L1 and clone L8 are significantly more resistant to CDDP than vector-control V1, V2 clones. Bars indicate standard error of mean.
* Two-sample t-test for parental HONE-1 cells vs V1 clones at all CDDP doses:
p = 0.009 0.02 0.0017 0.07 0.298 0.128