台灣梅花鹿磨角行為對墾丁高位珊瑚礁森林樹木的危害
王相華1) 王玉婷1) 伍淑惠1) 蘇聲欣1,2)
摘 要
有蹄類動物對於森林生態系的影響為當前世界重要的保育議題之一,其中鹿科動物磨角行為對 樹木的危害是受到廣泛關注的主題。墾丁國家公園管理處復育的梅花鹿於1994年開始陸續野放,目前 已有鹿隻危害墾丁地區森林的研究報導。本研究以墾丁高位珊瑚礁森林動態樣區於2013年6~9月調查 記錄到的0.72 ha範圍,共3207株樹木(胸徑≥ 1 cm)為觀測樣木,於2014年3~5月調查梅花鹿磨角行為 所造成的危害現象。調查結果顯示,樣木樹皮受到梅花鹿磨角損害的單株比例為7.6%,樹皮被磨損 的高度集中在離地面30~90 cm處。就徑級而言,梅花鹿偏好選擇胸徑2~3.9 cm的小徑級單株來磨角,
胸徑≥ 8 cm的單株被磨角的機率明顯降低;就樹種而言,梅花鹿偏好選擇鐵色(Drypetes littoralis)、
土楠(Cryptocarya concinna)、石苓舅(Glycosmis citrifolia)的單株來磨角,紅柴(Aglaia formosana)、
毛柿(Diospyros philippensis)、白榕(Ficus benjamina)、正榕(Ficus microcarpa)、山欖(Planchonella obovata)被磨角的機率則明顯偏低。梅花鹿磨角的主要對象是森林中的稚樹,會對森林的天然更新產 生衝擊;此外,梅花鹿磨角行為有明顯的樹種選擇性,會影響森林的樹種組成。本文針對於梅花鹿磨 角行為的樹木選擇提出概要性的討論。
關鍵詞:台灣梅花鹿、磨角行為、樹木危害、選擇偏好、森林更新。
王相華、王玉婷、伍淑惠、蘇聲欣。2019。台灣梅花鹿磨角行為對墾丁高位珊瑚礁森林樹木的危害。
台灣林業科學34(3):207-15。
1)林業 試驗所 ,10066台北市南海路53號 Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan.
2)通訊作者 Corresponding author, e-mail:[email protected]
2018年12月送審 2019年7月通過 Received December 2018, Accepted July 2019.
Research note
Fraying Damage to Trees by Formosan Sika Deer in the Kenting Uplifted Coral Reef Forest
Hsiang-Hua Wang,1) Yu-Ting Wang,1) Shu-Hui Wu,1) Sheng-Hsin Su1,2)
【Summary】
Impacts of ungulates on forest ecosystems are an important conservation issue in the world today, including fraying damage by deer to trees. Formosan sika deer was reintroduced into Kent- ing National Park in 1994; and now damage to the forest by deer has been reported. In this study, 3207 trees (with a diameter at breast height (DBH) of ≥ 1 cm) recorded in a 0.72-ha forest plot in 2013 were used as sample individuals, and fraying damage was investigated in 2014. According to the survey, 7.6% of sample individuals had been damaged by fraying, and the height of the rubbed tree-bark was concentrated at 30~90 cm from the ground. In terms of tree diameter, sika deer pre- ferred saplings with a DBH of 2~3.9 cm for fraying, and the fraying ratio was significantly reduced on trees with a DBH of ≥ 8 cm. In terms of tree species, sika deer preferred Drypetes littoralis, Cryptocarya concinna, and Glycosmis citrifolia for fraying, while avoiding Aglaia formosana, Diospyros philippensis, Ficus benjamina, F. microcarpa, and Planchonella obovata. Fraying by sika deer causes serious damage to forest saplings and probably impacts the natural regeneration of forests. In addition, fraying damage to trees reveals obvious species preference, which may lead to changes in the tree species composition of forests. This article discusses tree selection for fraying by sika deer.
Key words: Formosan sika deer, fraying, tree damage, preference, forest regeneration.
Wang HH, Wang YT, Wu SH, Su SH. 2019. Fraying damage to trees by Formosan sika deer in the Kenting Uplifted Coral Reef Forest. Taiwan J For Sci 34(3):207-15.
大型鹿科動物對植物有咬食枝葉(browsing)、
剝皮(bark-stripping)及磨角(fraying)行為,其 族群變化會對植被及依賴植物維持的生態系產 生巨大影響,故有學者認為大型鹿科動物是生 態系的基石物種(keystone species) (Waller and Alverson 1997),因這些物種對保持生態群落 的結構起著重要的作用,它們影響著生態系統 中其他許多生物,並決定了群落中的群聚結構 及物種組成。梅花鹿(sika deer, Cervus nippon) 廣泛分布於亞洲大陸東側的西伯利亞、韓國、
中國東部,以及日本、台灣等西太平洋島嶼,
台灣梅花鹿(Formosan sika deer, Cervus nippon
taiouanus)則為其下的台灣特有亞種,野外族 群已在1969年左右滅絕(McCullough 1974)。墾 丁國家公園管理處自1984年進行台灣梅花鹿復 育,並於1994年開始在墾丁國家公園內陸續野 放。野放後鹿隻適應良好,在區域族群數量激 增下,近年來已陸續有梅花鹿對森林危害的研 究報告(Wu 2007, Wang et al. 2015, Lu 2018)。
位於梅花鹿野放區域範圍內的「墾丁高位珊瑚 礁自然保留區」於1994年依據文化資產保存法 公 告 設 立 , 其 目 的 為 保 護 台 灣 唯 一 鮮 少 人 為 干 擾 的 高 位 珊 瑚 森 林 生 態 系 。Wu ( 2 0 0 7 ) 年 曾 在 此 保 留 區 調 查 梅 花 鹿 族 群 密 度 及 對 林 木
幼 苗 的 危 害 , 發 現 鹿 隻 大 多 取 食 高 度 在60公 分以下苗木,苗木被折枝的數量不多,此研究 推論:「目前研究地內梅花鹿密度不高,對苗 木啃食仍未造成嚴重危害」。筆 者 先 前 在 此 地 進 行 森 林 植 被 動 態 的 研 究 , 發 現 到 森 林 的 幼 齡稚樹(sapling)在2001~2013年間之減少比例 高達52%,平均每年減少約4.3%,依據現場觀 察推測其可能原因來自於梅花鹿的咬食或磨角 行為(Wang et al. 2015)。Lu (2018)於2014年12 月在此保留區及周邊森林架設15個6×6 m的圍 籬組,並於周遭設置一個等面積的對照組。自 2015年1月至2017年10月,每隔三個月記錄樣 區內高度大於10公分的木本植物小苗其存活狀 況,結果顯示,圍籬組共有35種2333棵木本植 物小苗,對照組則有22種888棵,小苗的新增 及存活於圍籬內外具種間差異,評估台灣梅花 鹿 為 影 響 木 本 小 苗 動 態 的 重 要 因 子 。 上 述Wu (2007)及Lu (2018)等之研究著重於梅花鹿咬食 對樹木幼苗的危害,而Wang et al. (2015)的研究 根據墾丁森林動態樣區稚樹數量隨時間改變,
推論梅花鹿咬食、磨角行為已嚴重影響樹木的 更新,則僅為間接證據。本文為驗證前述Wang et al. (2015)的研究推論,遂於此保留區進一步 執行梅花鹿對森林樹木磨角的現象調查。
磨 角 行 為 是 指 公 鹿 經 由 鹿 角 摩 擦 樹 皮 方 式以去除鹿角上的絨毛、標記氣味,此一行為 常導致樹木受損、死亡(Gill 1992)。也有研究 者提及,梅花鹿有用鹿角尖端磨擦大樹的習性 (Larner 1977)。台灣梅花鹿公鹿的角約在4~6月 脫落,隨即長出新角,9~10月鹿角完整鈣化,
開 始 有 磨 角 行 為 、 脫 去 絨 毛 , 接 著11~1月進 入交配期,此時期以磨角標誌領域的行為會更 加明顯(Chen and Wang 1999, Mu-Sheng Tsai, personal communication)。Larner (1977)在愛 爾 蘭 一 處 森 林 研 究 顯 示 有 高 達3 0 % 樹 木 受 到 引進(introduced)梅花鹿的磨角危害,Schloeth (1968)則報導紅鹿(red deer, Cervus elaphus)造 成瑞士一處森林有3%的樹木遭受磨角危害,顯 示鹿科動物磨角會造成森林不同程度的危害。
Chen and Wang (1999)的調查記錄到墾丁地區 220株樹木遭受台灣梅花鹿磨角危害,為取樣數
量的1.68%,認為當時的野放梅花鹿族群對植被 的影響應屬有限;該研究同時指出,未曾發現 梅花鹿在崎嶇的高位珊瑚礁地形活動,因此並 沒有記錄到高位珊瑚礁森林樹木遭受到鹿隻磨 角的危害。但Wang et al. (2015)於墾丁高位珊 瑚礁森林的研究則顯示,該森林已明顯受到梅 花鹿擾動的影響。Gerhardt et al. (2013)分析鹿 科動物對歐洲森林危害的38篇研究報告發現,
較多文獻探討咬食枝葉的危害(82%)、剝皮危害 次之(26%),較少探討磨角危害(21%),顯示鹿 科動物的磨角危害較少受到關注。
為 了 解 高 位 珊 瑚 礁 森 林 受 梅 花 鹿 磨 角 的 危害情形,本研究在墾丁自然保留區的森林動 態樣區(forest dynamics plot;總面積10 ha,
400×250 m)內,挑選了4條跨越凸起岩塊、凹 陷谷地的180×10 m東西向樣帶,合計有72個 於2013年6~9月間完成每木調查的10×10 m小 區,做為本研究調查範圍。受到菲律賓板塊對 與亞洲大陸板塊擠壓的影響,高位珊瑚礁的凸 起岩塊、凹陷谷地多呈現南北走向,因此本研 究選擇可跨越不同地形(棲地)的東西向樣帶為調 查區域,以增加樣本的棲地代表性。
這72個小區於2013年共記錄到3207株胸 徑(diameter at breast height; DBH) ≥ 1 cm的單 株,其中包括兩種榕屬植物形成的支柱根:正 榕(Ficus microcarpa)支柱根83枝、白榕(Ficus benjamina)支柱根125枝,以及其他69種非支 柱 根 樹 種 的 樹 木 單 株2999株。由於此兩種榕 屬植物的支柱根生長形態近似於一般樹木的樹 幹,亦有可能遭受到梅花鹿的磨角,因此本研 究將支柱根視同於一般樹幹,逐一進行調查與 分析。本研究於2014年3~5月間重新調查上述 72個小區,記錄樣區內樹皮有被鹿隻磨損單 株的編號、種類、胸徑及磨損位置距離地面之 高度,包括樹幹磨損傷痕上緣高度、樹幹磨損 傷痕下緣高度。上述鹿隻磨損樹皮的危害只進 行一次調查,因此無法判定磨損的時間,痕跡 不 論 新 舊 皆 予 以 紀 錄 。 鹿 隻 對 樹 皮 的 危 害 包 括 磨 角 與 啃 咬 , 如 為 啃 咬 , 會 留 下 明 顯 的 齒 痕 或 深 淺 不 一 的 刮 痕 , 如 為 磨 角 , 會 留 下 長 約40~80 cm的平滑傷痕(M.-S. Tsai, personal
communication),本調查排除留有齒痕或深淺 不一刮痕的被啃咬樹木。
本 研 究 之 資 料 分 析 主 要 是 探 討 梅 花 鹿 磨 角行為是否與單株的樹種或胸徑具有關聯性。
我 們 採 取 的 分 析 方 式 是 將72個小區總合的單 株 組 成 資 料 視 為 當 地 森 林 之 「 背 景 組 成 」 , 再與小區內調查到的受危害單株組成資料做比 較,並以統計方法檢定兩者間的差異。在進行 分析前,先將單株的胸徑區分為1~1.9, 2~3.9, 4~7.9, 8~15.9, 16~31.9, ≥ 32 cm等6個徑級。
在 樹 種 分 析 方 面 , 雖 然72個小區共計調查到 60個樹種,但為避免稀少種資料不足所帶來的 不確定性,本研究僅針對單株數量≥ 30株的19 種進行分析。在統計方法上,同時採用卡方適 合度檢定(Chi-square goodness-of-fit test)以及 自助重抽法(bootstrap method)兩種檢定方法。
卡方檢定可比較受危害單株在不同樹種或胸徑 級之間的單株分配比例,與背景組成的分配比 例是否在整體上存有任何顯著差異;若有,即 表示單株受危害情形與樹種或胸徑級具有關聯 性。由於本研究的調查資料本質上是以樹木個 體(individuals)為單位的野外觀察結果,而非 經過實驗設計(experimental design)下的重覆樣 本(區)試驗結果,因此無法採行傳統的統計檢 定方法來進一步分析個別樹種或胸徑級是否具 有顯著性的數量差異。為此,本研究使用基於 重複抽樣方法(resampling)的自助重抽檢定,
以克服這項分析限制(Manly 2007)。本研究採 用的自助重抽檢定是從森林背景組成資料中的 3207單株重複進行隨機重置抽樣(sampling with replacement),每次抽取與實際調查到受危害單 株相等數量的自助重抽樣本(bootstrap sample),
再重複進行抽樣直至總計得到10,000個樣本。而 後計算自助重抽樣本分布(bootstrap distribution) 的2.5與97.5%百分位值(percentile),以估算 95%信賴區間(confidence interval),再與實際 觀測的受危害單株數量相比較(Manly 2007)。
若實際觀測數量偏離於95%信賴區間,則代表 梅 花 鹿 對 於 特 定 樹 種 或 胸 徑 級 具 有 磨 角 行 為 上 的 偏 好 或 迴 避 性 。 以 上 資 料 分 析 工 作 皆 是 以R 3.5.0統計語言(R Core Team 2018)完成。
研 究 結 果 顯 示 , 受 磨 損 樹 幹 的 傷 痕 下 緣 平 均 危 害 高 度(±standard deviation)為離 地 面3 0 . 0 ± 1 4 . 1 c m , 傷 痕 上 緣 的 平 均 危 害 高 度 為8 9 . 5 ± 9 . 2 c m 。 在 3 2 0 7 個 觀 測 樣 本 單 株 中 總 共 記 錄 到2 4 4 株 受 到 樹 皮 磨 損 的 危 害,平均受危害比率為7.6% (Table 1)。土楠 (Cryptocarya concinna)、石苓舅(Glycosmis citrifolia)的受危害比率超過20%,鐵色(Drypetes littoralis)、山柚(Champereia manillana)、
蟲屎(Melanolepis multiglandulosa)、皮孫木 (Pisonia umbellifera)的受危害比率高於10%,
但白榕、山欖(Planchonella obovata)、正榕、
毛柿(Diospyros philippensis)、茄冬(Bischofia javanica)、大葉山欖(Palaquium formosanum)等 6個樹種的受危害比率為0 (Table 1)。卡方檢定 的結果顯示,受危害單株不僅與森林背景組成的 徑級分配比例有明顯差異(X2 = 130.0, df = 5, p <
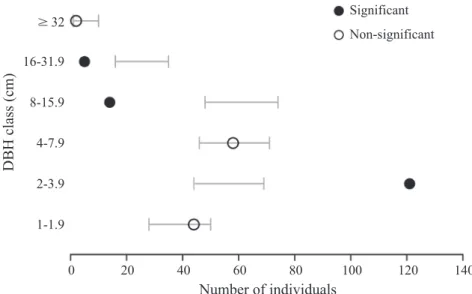
0.001),也與背景組成的樹種分配比例有明顯差 異(X2 = 137.5, df = 18, p < 0.001 )。自助重抽檢 定的結果顯示,梅花鹿磨角偏好選擇胸徑2~3.9 cm的單株,胸徑≥ 8 cm的單株被梅花鹿磨角危 害 的 機 率 則 明 顯 降 低(Fig. 1);就樹種而言,
梅花鹿喜歡選擇鐵色、土楠、石苓舅的單株來 磨角,紅柴(Aglaia formosana)、毛柿、白榕、
正榕、山欖被磨角的機率則明顯偏低(Fig. 2)。
梅 花 鹿 喜 好 選 擇2~3.9 cm徑級的單株磨 角,胸徑≥ 8 cm的單株明顯較不易受到磨角危 害。國內外也有許多研究指出,鹿科動物喜好 選擇較小徑級的單株磨角(Motta and Nola 1996, Vila et al. 2004, Ramos et al. 2006, White 2012, Lu et al. 2012, Charco et al. 2016),但並沒有探 討其原因。曾於墾丁國家公園管理處梅花鹿復 育中心任職現場管理員7年的蔡木生先生表示,
梅花鹿喜歡選擇樹幹有彈性的小樹,在磨樹時 梅花鹿會先將鹿角中下段頂住樹幹較下方處,
再朝向樹幹較上方處進行磨角;較大徑級的單 株因沒有彈性,導致磨角時樹幹並不會產生明 顯的搖動,因此不是梅花鹿喜愛選擇用來磨角 的對象(M.-S. Tsai, personal communication)。
本研究在野外觀察上也發現,梅花鹿針對少數 較大徑級的單株進行磨角,但主要是用鹿角尖
端 刮 磨 樹 皮 , 與 運 用 小 徑 級 單 株 磨 角 的 方 式 完全不同,Larner (1977)的研究也有類似的記 錄。
Wang et al. (2015)分析墾丁森林動態樣區 2001至2013年間3次每木調查資料,發現胸徑 1~4 cm的稚樹密度明顯減少,尤其是胸徑1~2 cm的幼齡稚樹由2001年約1600株/ha降至2013 年約770株/ha,減少之比例高達約52%;胸徑 2.1~4 cm的單株密度由2001年約1200株/ha降 至2013年約1040株/ha,減少之比例約為13%;
本研究則指出,梅花鹿磨角危害的單株集中在 2~3.9 cm的稚樹。對照以上兩項研究的結果,
並輔以本團隊常年在當地的野外觀察,我們推 斷梅花鹿對墾丁高位珊瑚礁森林的危害可能不 僅侷限於磨角行為,梅花鹿咬食枝葉行為對胸 徑1~2 cm幼齡稚樹所造成的危害可能尤勝於磨
角 行 為 對 胸 徑2~4 cm單株所產生的危害。蔡 木生於受訪時提及,梅花鹿通常可以輕易咬食 胸 徑1~2 cm小樹的枝葉,並在咬食枝葉時直 接扯斷胸徑1~2 cm小樹的樹幹,但胸徑> 2 cm 單株的枝葉高度通常已超過梅花鹿可直接咬食 的高度(M.-S. Tsai, personal communication)。
Renaud et al. (2003)的研究也發現,稚樹枝葉的 高度會明顯影響其受紅鹿危害的強度及頻度,
枝葉高度較低的稚樹通常較容易受到紅鹿咬食 的危害。綜合Wang et al. (2015)的研究、本研究 結果及訪談資料,梅花鹿對墾丁高位珊瑚礁森 林樹木的危害目前集中在胸徑1~4 cm的稚樹,
其中磨角行為主要造成對2~4 cm稚樹的危害,
而咬食枝葉行為則有可能是胸徑1~2 cm稚樹的 危害的主要原因。
本 研 究 的 結 果 也 發 現 梅 花 鹿 對 磨 角 樹 種 Table 1. Statistics on observed and frayed individuals of tree species recorded in a 0.72- ha sampled area in the Kenting Uplifted Coral Reef Forest Dynamics Plot. Only common species with ≥ 30 observed individuals are listed
Species Chinese Species No. of individuals No. of frayed Frayed name code observed individuals ratio (%)
Diospyros maritima 黃心柿 DiosMa 1548 110 7.1
Drypetes littoralis 鐵色 DrypLi 232 42 18.1
Aglaia formosana 紅柴 AglaFo 150 3 2.0
Ficus benjamina 白榕 FicuBe 125 0 0.0
Planchonella obovata 山欖 PlanOb 92 0 0.0
Ficus microcarpa 正榕 FicuMi 83 0 0.0
Champereia manillana 山柚 ChamMa 79 9 11.4
Diospyros eriantha 軟毛柿 DiosEr 78 4 5.1
Dendrocnide meyeniana 咬人狗 DendMe 66 3 4.5
Macaranga tanarius 血桐 MacaTa 65 3 4.6
Cryptocarya concinna 土楠 CrypCo 60 17 28.3
Diospyros philippensis 毛柿 DiosPh 59 0 0.0
Melanolepis multiglandulosa 蟲屎 MelaMu 48 7 14.6
Bischofia javanica 茄冬 BiscJa 44 0 0.0
Palaquium formosanum 大葉山欖 PalaFo 43 0 0.0
Antidesma pentandrum var. barbatum 枯里珍 AntiPe 33 3 9.1
Pisonia umbellifera 皮孫木 PisoUm 31 5 16.1
Glycosmis citrifolia 石苓舅 GlycCi 31 8 25.8
Leea guineensis 火筒樹 LeeaGu 30 0 0.0
Others 其他樹種 310 30 9.7
Total 3207 244 7.6
Fig. 1. Numbers of frayed tree individuals among different diameter at breast height (DBH) classes. Points represent the observed numbers of frayed individuals, and gray bars are the 95 confidence intervals estimated by the bootstrap method (n = 10,000).
Fig. 2. Numbers of frayed tree individuals among different tree species. Points represent the observed numbers of frayed individuals, and gray bars are the 95 confidence intervals estimated by the bootstrap method (n = 10,000). Note that the numbers of individuals are shown on a logarithmic scale for visual clarity. Complete species names for species codes are given in Table 1.
0 16-31.9
8-15.9
4-7.9
2-3.9
1-1.9
20 40 120
Number of individuals
DBH class (cm)
60 80 100 Significant Non-significant
140
PlanOb PalaFo LeeaGu FicuMi FicuBe DiosPh BiscJa MacaTa DendMe AntiPe AglaFo DiosEr PisoUm MelaMu GlycCi ChamMa CrypCo DrypLi DiosMa
Species
Significant Non-significant
1 2 5 10 20 40 80 160
log (Number of individuals + 1)
具有選擇性。其中,白榕、正榕支柱根雖然分 別調查到有125及83枝,但尚未發現有被梅花 鹿磨角危害的紀錄。此類榕屬植物支柱根的上 方與主幹側枝聯結、下方定著於土壤,導致鹿 隻若使用支柱根磨角時並不會產生類似同徑級 稚樹的主幹搖動現象,研判可能是造成梅花鹿 不喜愛選擇其當作磨角對象的原因(M.-S. Tsai, personal communication)。除了支柱根類型的榕 屬樹種外,本研究也發現山欖、毛柿、茄冬雖 然分別調查到有92, 59及44株,但也同樣未發現 有被梅花鹿磨角危害的紀錄;相對的,鐵色、
土楠、石苓舅則有較高的比例被梅花鹿選為磨 角的對象。國外的鹿科動物紅鹿、梅花鹿及麆 鹿(roe deer, Capreolus capreolus)在1960~1980 年 代 就 曾 被 研 究 報 導 對 磨 角 的 樹 種 有 選 擇 性 (Thompson 1969, Larner 1977, Langvatn 1982);
Vila et al. (2004)發現黑尾鹿(black-tailed deer, Odocoileus hemionus sitkensis)喜愛選擇美西側 柏(Thuja plicata)、柳屬(Salix spp.)、美國赤楊 (Alnus rubra)樹木磨角;Boulanger et al. (2009) 的研究則發現麆鹿磨角的樹種選擇與其木材密 度呈正相關;Gill (1992)也認為樹木物理特性 (physical characteristics)對鹿隻磨角選擇的影 響比樹木種類更為重要。雖然前述國外少量的 研究指出,鹿科動物磨角的樹種選擇似乎與樹 木的物理特性有關,但我們仍不清楚導致梅花 鹿磨角行為對墾丁高位珊瑚礁森林樹種具有選 擇性的確切影響因子,有待進一步的研究予以 釐清。例如可經由各種樹木不同徑級稚樹的木 材密度、木材彈性進行檢測,亦或是分析各種 樹木不同徑級稚樹的樹木形態差異,並經由合 理的試驗設計、分析,深入探討此一問題。此 外,我們有考量到樹種與胸徑這兩項變因之間 可能存有資料相關性,或是因子間的交互作用 (interaction)。但是由於本研究記錄到的磨角 現象包含了新、舊磨痕,我們無法確知磨痕發 生時的樹木胸徑確切大小,因此我們目前無法 以嚴謹的方式分析這兩項變因之間是否存有相 關性或交互作用。若要更進一步探討此間的關 係,還需要透過其他野外觀察設計或是進行更 多次的定期複查,取得更詳細的磨痕記錄資料
方可釐清。
鹿科動物在許多森林生態系中扮演基石物 種的角色,鹿隻的大量攝食也會造成植物社會 組成的改變(Abrams and Johnson 2012, Perea et al. 2014),間接對森林生態系造成系統性且跨 越營養階級的影響(Côté et al. 2004, Takatsuki 2009, Warren 2011)。本試驗之地點位於墾丁高 位珊瑚礁自然保留區內,綜合本研究及Wang et al. (2015)的研究發現,雖然截至2014年止野放 梅花鹿對墾丁高位珊瑚礁森林樹木的危害多侷 限於胸徑1~4 cm的稚樹,但此一胸徑級的單株 佔高位珊瑚礁森林動態樣區的全體單株比例高 達約50% (Wang et al. 2004),顯示野放梅花鹿 對此一受文化資產保存法保護之獨特森林已造 成明顯的危害。梅花鹿群長期對樹木的磨角與 咬食行為已經造成森林稚樹大量死亡,由現場 樹木幼苗被咬食至僅有少數殘存(H.-H. Wang, personal observation)的情形判斷,稚樹數量也 難以經由幼苗生長的過程予以補充,評估梅花 鹿群已對此地高位珊瑚礁森林的天然更新產生 重大衝擊;此外,因梅花鹿的磨角行為具有樹 種選擇性,導致不同樹種的族群數量產生差異 性的動態變化,評估也將會改變未來的森林樹 種組成與結構。若上述樹木結構、組成的改變 持續或擴大,將有可能造成高位珊瑚礁森林生 態系統的崩解,除應當進行更為深入的調查、
研究,瞭解不同強度的梅花鹿擾動對高位珊瑚 礁森林的系統性影響外,管理單位也必須及早 提出適切的族群數量管理方式。
謝 誌
本 研 究 由 農 委 會 林 業 試 驗 所 提 供 研 究 經 費、林業試驗所恆春研究中心同仁協助現場調 查及試驗樣區維護、管理,在此深表謝意。
引 用 文 獻
Abrams MD, Johnson SE. 2012. Long-term impacts of deer exclosures on mixed-oak forest composition at the Valley Forge National His-
torical Park, Pennsylvania, USA. J Torrey Bot Soc 139:167-80.
Boulanger V, Baltzinger C, Saïd S, Ballon P, Picard J, Dupouey JC. 2009. Ranking temperate woody species along a gradient of browsing by deer. For Ecol Manage 258:1397- 406.
Charco J, Perea R, Gil L, Nanos N. 2016.
Impact of deer rubbing on pine forests: im- plications for conservation and management of Pinus pinaster populations. Eur J For Res 135:719-29.
Chen SC, Wang Y. 1999. Rubbing of the For- mosan sika deer (Cervus nippon taiouanus) and its effect on trees in Kenting National Park. Biol Bull Natl Taiwan Norm Univ. 34(2):
151-62. [in Chinese with English abstract].
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM. 2004. Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113-47.
Gerhardt P, Arnold JM, Hackländer K, Hochbichler E. 2013. Determinants of deer impact in European forests–A systematic lit- erature analysis. For Ecol Manage 310:173-86.
Gill RMA. 1992. A review of damage by mammals in north temperate forests: 1. deer.
Forestry 65:145-69.
Langvatn R. 1982. An investigation of fray- ing damage on trees caused by red deer. Nor- wegian J For 90:270-81. [in Norwegian with English summary].
Larner JB. 1977. Sika deer damage to mature woodlands of southwestern Ireland. In: Pro- ceedings of the 13th International Congress of Game Biologists. Atlanta, GA: Wildlife Man- agement Institute. p 192-202.
Lu HF, Chen SC, Wang Y. 2012. Rubbing behavior of the Formosan sika deer (Cervus nippon taiouanus) in Kenting National Park. J Natl Park 22(3):22-30. [in Chinese with Eng- lish abstract].
Lu JW. 2018. Herbivory effects of Formosan
sika deer (Cervus nippon taiouanus) on com- munity composition of native woody seedlings in a tropical forest. Master’s thesis, Depart- ment of Life Science, Tunghai Univ. 62 p. [in Chinese with English abstract].
Manly BFJ. 2007. Randomization, bootstrap and Monte Carlo methods in biology. 3rd ed.
Boca Raton, FL: Chapman & Hall/CRC. 455 p.
McCullough DR. 1974. Status of larger mam- mals in Taiwan. Taipei, Taiwan: Tourism Bu- reau. 36 p.
Motta R, Nola P. 1996. Fraying damages in the subalpine forest of Paneveggio (Trento, Italy): a dendroecological approach. For Ecol Manage 88:81-6.
Perea R, Girardello M, San Miguel A. 2014.
Big game or big loss? High deer densities are threatening woody plant diversity and vegeta- tion dynamics. Biodivers Conserv 23:1303-18.
R Core Team. 2018. R: a language and envi- ronment for statistical computing, ver. 3.5.0.
Vienna, Austria: R Foundation for Statistical Computing. Available at http://www.r-project.
org.
Ramos JA, Bugalho MN, Cortez P, Iason GR. 2006. Selection of trees for rubbing by red and roe deer in forest plantations. For Ecol Manage 222:39-45.
Renaud PC, Verheyden-Tixier H, Dumont B.
2003. Damage to saplings by red deer (Cervus elaphus): effect of foliage height and structure.
For Ecol Manage 181:31-7.
Schloeth R. 1968. Analysis of rubbing and fraying by red deer in an alpine habitat (Swiss National Park). Swiss Comm Nat Sci 11:75.
Takatsuki S. 2009. Effects of sika deer on vegetation in Japan: a review. Biol Conserv 142:1922-9.
Thompson B. 1969. Fraying by roebucks at Glentress Forest. Deer 1:307-11.
Vila B, Guibal F, Torre F, Martin J. 2004.
Assessing spatial variation in browsing history by means of fraying scars. J Biogeogr 31:987-
95.
Waller DM, Alverson WS. 1997. The white- tailed deer: a keystone herbivore. Wildlife Soc B 25:217-26.
Wang HH, Sun IF, Chien CT, Pan FJ, Kuo CF, et al. 2004. Tree species composition and habitat types of a karst forest in Kenting, southern Taiwan. Taiwan J For Sci 19(4):323- 35. [in Chinese with English summary].
Wang HH, Wu SH, Sun IF, Chen KF. 2015.
Young sapling dramatically decreased during 2001 and 2013 in Kenting uplifted coral reef forest, southern Taiwan. J Natl Park 25(1):17- 23. [in Chinese with English abstract].
Warren RJ. 2011. Deer overabundance in the USA: recent advances in population control.
Anim Prod Sci 51:259-66.
White MA. 2012. Long-term effects of deer browsing: composition, structure and produc- tivity in a northeastern Minnesota old-growth forest. For Ecol Manage 269:222-8.
Wu YJ. 2007. Habitat use and sapling brows- ing by Formosan sika deer (Cervus nippon taiouanus) in karst forest, Kenting. Master’s thesis, Institute of Natural Resource Manage- ment, National Dong Hwa Univ. 60 p. [in Chi- nese with English abstract].