!
!
!"#$%&'()&*+,&-./!
0123!
45678797:!;<!=;;>;?@!
A;>>:?:!;<!B8<:!CD8:5D:!
EF78;5F>!GF8HF5!I58J:K687@!
LF67:K!GM:686!
!
NOPQ Odysseus RSTUVWXYZ!
Ectopic expression of Odysseus causes mitotic defects
! [\]!
^:8_AM95!B85!
!
!!!!!!!!!!!!!!!!!!!!!!`abcdefg!!
!!!!!!!!!hiJ86;K!j!AMF9_G8!G85?!
!
!!klm!nopqr
s9>@t!uvvn!
Acknowledgements
wx%+,/yz{pX&|}~•T/S€~•‚ƒ„…†!
‡ˆ~‰„Š•X`abcefg‹Œ~•Ž••w0•‘&’“”•
–X—˜™š›œ•~žŸ•‰“X ¡¢£›œX¤¥~¦§¨©ª«¬- X®"†¯•p©X—°±~•&|²‰©~‰©‹Œb•X~³••Ÿ´² X†w›œXµ¶k~·¸%&X¹º»‹Œb•¼½™¾¢£¿ÀXÁÂà Ä~ÅÆÇÈÉÊXĘ̈©®Í~ÎÏ%&XÐ!Ñ‹Œb•Ò-ÓÔÕ’
³Ö×~¦§!Ø*XÙÚÛ‹Œ~w›œÜÝ‘X`a~‹ŒÞXßà~•
•TáâXS¢~‰„Š†•ŠŠãäå‹Œ~æç‹Œ~èéê‹Œ~³ë ìS‹Œ~íî~ïð¯ñ23~ÅÆwòó‘~T¨©`ô~••X23õ ö÷øXùQ†!
ŠŠ¯úûü~›œ•±ýþÿ•X!"S#†$$„ŠX%&'&
(~w›œ³òó‘~)b*•¨©~ÅÆV+ÿ•X,-~./012•†
ŠŠ34~•Ž5—°6•78~9:®Í~;w<!=>?••@ABXC DE†ŠŠ3F~GHIJXKL‰MN~;%‰OÊ•~ÅÆ.—žPQ R†ŠŠ«S~•TUw›œ•X‰©"V‘~W•XVY~ý•ö›œ³Z [••.\]~•W?•¨©M~^_`ûa²‰bXcdefŠŠg:J85WM ïð•X233ó~;hzi?W•jk‘x78~l²mnoô~»%pq _?f;TrwstX3u~v%³•Vw\xyX[z{'|zœ~}~?
¨©Q•—€~ŠŠ_†ŠŠÊ•~žP‚ƒ‚„ø…X†,¦§‡ô†ˆ~
!‰„)‰Š‹ŒfŠŠ•Ý‰Ž•~•‘‘X’e“”•–:@—˜™š›œX 1•“Z[ÈÈT6žTŸX Œ¡¢“•X£•"ó!de~¤¥¦¦X§
¨C7:©MF5:~¦§¬-›œ~ª%•;ž«²Z[”ŸX¬-~T?_ÞXý þ~-.''|÷®¯›“•fw¯±~•”{•X°e±²~³•´ŠŠ³ µ•~•Þ‰T¶·¸œ~•¹µ?~^º»}¼Ê콌f!
ÇÈ„Š¾¿À‹Œ³ÁÛ‹Œ~%_Þßà•Ã²TÄÅX-.Æ z~;TÇÈÉÊË-.•X%Ì~›wͩΞÏó•"•"„Š~%Ì)
·•‰ø~•T…ÐÑ•Ò¥Xûü~»X‰¼ÊfwÓ±••ÔÕ¨©~Ö
¬%×Û&€X`a~••&*?‚ƒ‚„X›œØÙ~‰„Š†!
\2ÚÛ—Ü~•$”„ŠX%•XÌΆ„Š•TÓÛøX¨¨~
v%Ýʳ•·[~Wà•ýþ•S€†„ŠÞÞ~·ß•XOʳ12à .°/~;*áâ³ZR[fÅÆpãÒ¥ËÌ•'|ä~ÞÞpq?†Š Šåå~ÈÈæç[OÊ•è%>†,éêP9•~ŠŠ_Þ~à_Þ¿
k~•ë²IJ%$bX†ŠŠìH~·_X„Š~³•³.í~ŠŠ_î ï;ýþ•ð›œ•~àPžT‰ñòó~••ô%•$õöT-X÷ø~
••wùúûüX—˜~Týþ=™~ŠŠ_†•ÿ_Þf! !
„Š‰!‘"²X!‰"Î~•%©ÛX#$~%õTáâXS¢†!
$}~•”&¯ñ23~'9•$(ÿX§§~;Twâ‘X))³*
+†!
,”
w-.&XÖô‘~/012`X%*RS,/°'3s4X12†
!"#$$%&$! 5!"$'6! %£•"78Q~*RS¢£X{"9:/~;<¢£³á=
>?¢£X@aABüC.DX12†EFDNAGHVI•¢JK~OdsH%L unc-48'12MNKO/P'~{Q)RT•"ST•UV~w<¢£/WX k~;<¢£³á=>?¢£XST•UVT“YZ0XQ[†EF\=X- .JK~ w<¢£k~OdsHÆ”PQwABS]X^_~¬`õ7abic L®Y^üX'Sd~¦Ë˜ABX'3d~ª%²e=ixVüfg;.h i†jÿØÙX™k~¢£X12`õfg~l¦½mUAS/Gal4n•~¦NO PQX惙•kVI†EFo",/XOdsHw<¢£pA'3n•PQX•
¢JK~qTá=>?¢£XOdsH*RSAB³pBS]'3rs8tNÈ~
<¢£³;<¢£XOdsHu.*†vwo",/XOdsHw¢£¿ÀPQ~)
*RSxMX8tYZ~RS3¿†EFÓÔ¦§y]¿ÀXÖ×~w<¢£
XOdsHPQ—~*zÕ{|}’~²•€~ÅÆ%w{|VW’P'YZ†2
•~abOdsH`õ¥mXVüfgi~‚ƒ{|}’kX{|VW†•„$…
w{|†XÖ×~8QOdsHl¦•„²ÃćkˆXV‰~OdsHw;<¢£
³á=>?¢£°RSXABüC.D~lõŠ„centromere-drive model†
!
!
!
!
!
Abstract
Genes that contribute to the reproductive isolation between species are known as “speciation genes’. Odysseus (OdsH) is a hybrid male sterility gene between Drosophila mauritiana and D. simulans. According to sequence analysis, OdsH duplicated from unc-4 recently in the Drosophila genome preserving the intron exon structure and the paired-like homeodomain. Among D. melaongaster subgroup, the homeodomain of OdsH in D. mauritiana, and D. simulans evolved rapidly. Previous studies showed that OdsH is highly expressed in the testes, and the function of OdsHmel is to enhance male fertility by accelerating the rate of sperm maturation.
Three OdsH alleles from D. mauritiana, D. simulans, and D. melanogaster were ectopically expressed in D. melanogaster with the Gal4/UAS system to uncover the molecular mechanism for the function of OdsH. In germline expression, OdsHmau caused developmental defect in the reproductive organs. Germline expression of OdsHmel in females caused the overlapping of nuclei in the embryos and embryonic lethality. In the eyes, all three alleles caused critical defect in the development resulting in small-eye phenotype. Results from embryos and eye discs indicate that cell cycle was perturbed at the mitotic phase with the expression of OdsHmel.
Therefore, the molecular mechanism of OdsH appears to play a role in the M phase of cell cycle regulation. Combined with the recent data in the cell culture indicating that OdsH can bind to pericentric satellite DNAs, the mechanism for OdsH to cause hybrid male sterility between D. mauritiana and D. simulans supports the centromere- drive model.
Keywords: Drosophila, Odysseus, gene duplication, homeodomain, ectopic expression, cell mitotic.
Contents
•Ž•#*•‘’……….i
Acknowledgements………...………...ii
Chinese abstracts ………...iv
English abstracts………....………...……...v
List of tables………...………...viii
List of figures………..…………...ix
Introduction………..…..………1
Materials and Methods………..……...7
Fly stocks………..………..………...7
Transgenic constructs……….…………7
Transgenic lines………..……...7
RNA extraction……….……….….…….10
RT-PCR……….….….…10
Embryo collection………...……….….…...11
Testes and ovary dissection……….………..….……..11
Scanning electron microscopy………...………..12
Immunostaining……….…..………12
Results………..……...14
The OdsH transgenic lines………...14
Ubiquitous and neuron expression of OdsH alleles………...…...14
Germlines expression of OdsH alleles……….…16
Eye disc expression of OdsH alleles………21
Morphology of adult eyes………21
The influence of OdsHmel in mitotic cells………23
Discussion………...…26
References………...…35
Appendix: Generate the germline available site specific transgenic vector……….…...40
Generate the unc-4 knockdown flies……….…..47
Recipe………...53
Row Data………...54
List of Tables
Table 1. The list of primers for expression analysis……….……….…..11
Table 2. The list of UAS-OdsH transgenic lines…………...……….……..14
Table 3. Phenotypes of ectopic expression of OdsH alleles………15
Table 4. The sequences for the oligos of mir6_unc-4mel………...….….48
Table 5. The list of UAS-mir_unc-4 transgenic lines………...……….50
Table 6. The sequences for the oligos of mir6_OdsHmel……….…52
List of figures
Fig. 1. Procedure of pP {5’-UAS::OdsHsim} and pP {5’-UAS::OdsHmau}...8
Fig. 2. The cross steps to get ectopic expression line on second chromosome……….9
Fig. 3. The cross steps to get ectopic expression line on the third chromosome…...9
Fig. 4. RT expression of OdsH in males and females……….………….16
Fig. 5. Gal4 inducible expression in the germline of the ovary………...17
Fig. 6. The morphology of the early embryos………..………...……..18
Fig. 7. Nucleus pattern in the embryos…….………...19
Fig. 8. Embryos in the gastrulation stage…………..………...19
Fig. 9. The morphology of the male reproductive organ…………...………...……...20
Fig. 10. The morphology of adult eye with ectopic expression different alleles of OdsH………22
Fig. 11. The width of eye with different OdsH alleles expressed...23
Fig. 12. The mitotic cell with OdsHmel expression………..24
Fig. 13. Quantification of the mitotic cells at the first and second mitotic waves…...24
Fig. 14. S phase cell in third-instar larva eye disc………...……….………...25
Fig. 15. The alignment of three OdsH alleles………...………29
Fig. 16. The centromere-drive model………..………33
Fig. 17. The pUAST transgenic vectors……….……….…….40
Fig. 18. The pUASP transgenic vectors………...41
Fig. 19. Site-specific integration mediated by !C31………….……….….42
Fig. 20. The site specific transgenic vector pUAST attB……….…....43
Fig. 21. Restriction enzyme digestion of pUASP attB by NheI and SpeI………45
Fig. 22. Procedure of the construct of pUASP attB…...………..………...46
Fig. 23. The target and structure prediction of two miRNA………....49
Fig. 24. RT expression of unc-4 ………..………51
Introduction
After the publication of “The Origin of Species”, the concept that all the life forms have evolved over time through the mechanism of evolution has become widely accepted. Nevertheless, the debate around the speciation developed into two theoretical concerns. First, how evolution initiates species evolution and second, how we define a “good” species? Those issues could only be discussed with the continued elaborating clarifying works upon the definition of species.
In Mayr’s biological species concept, he defined that “species are groups of actually or potentially interbreeding populations which are reproductively isolated from other such groups“ (Mayr 1963). Mayr's statement was the most widely applied concept among the existed species concepts. His proposed based on three advantages.
First, closely related species with little distinguishable characters can be easily identified. Second, it can explain the existence of discontinuous phenotypes among sexually reproducing organisms. Moreover, the most important reason is that the researchers can immediately verify the existence of the subject (Coyne and Orr 2004).
In the biological species concept, the mechanism of isolation are particularly important, in which species of sexual organisms are defined as derived from
reproductive isolation. Any biological difference or the reproductive characteristics between the populations that reduce gene exchange will cease gene flow between them and prevent species from fusing with another sibling species. There are many kinds of reproductive isolations, i.e. the pre-zygotic and the post-zygotic isolation.
Most of the prezygotic isolations are resulted from the pre-mating isolation, while under the instances of postmating, prezygotic isolation may have happened on some species. In the genetic view of speciation, genes that contribute to the reproductive isolation are ‘speciation genes”, and most in the form of hybrid inviability or sterility.
The molecular mechanism for the speciation genes to cause reproductive isolation is interested.
Until now, several speciation genes in Drosophila had been studied, including Hybrid male rescue (Hmr), Na/K-ATPase alpha subunit isoform 2 (JYAIpha),
Nucleoporins 96 (Nup96), and Odysseus (OdsH). Hmr is the one in Drosophila causes hybrids incompatibility; Hmr, the X-linked gene in Drosophila melanogaster, encodes a rapidly evolving protein with the sequence similar to the myb/SANT-like domain in Adf-1 (MADF) class of DNA binding proteins, undergo functionally diverge between D. melanogaster and its sibling species (Brideau et al. 2009).
Interspecific cross-mating of D. melanogaster females to the males of D. simulans resulted in lethality of the F1 hybrid males, because of the interaction between Hmr and the autosomal gene lethal hybrid rescue (Lhr), which localizes to heterochromatic regions of the genome in the D. simulans. Lhr also undergo to sequence diverged between these species.
JYAIpha was identified that resulting the hybrid sterility in Drosophila.
JYAIpha encodes a protein, in which has a functional motif of an alpha subunit of a Na+ and K+ adenosine triphosphatase. The specific activating site involved in ion exchange, which is essential for the sperm motility. In D. melanogaster, JYAIpha is located in the fourth chromosome but translocated to the third chromosome in D.
simulans lineage. Based on this long-term evolutionary transposition event, the hybrids males of D. melanogaster to D. simulans at last completely loss the JYAIpha gene and were sterile (Masly et al. 2006).
Nup96, which encodes a nuclear pore protein, was mapped as a hybrid inviability gene in hybrid males between D. melanogaster and D. simulans. In these two species lineage, Nup96 evolved by positive selection, thus the D. simulans Nup96
protein is no longer compatible with the interacting factor that encoded by the D.
melanogaster X chromosome. Later, another hybrid lethality gene nucleoporin Nup160kDa (Nup160) gene has been identified directly interact to Nup96 (Presgraves et al. 2003; Tang and Presgraves 2009).
Odysseus (OdsH) is also the defined speciation gene that result in hybrid male sterility. The OdsH locus was noticed in hybrid sterility associated with three X chromosome regions in D. mauritiana and D. simulans hybrids, when the D.
mauritiana allele introgressed into an appropriate D. simulans background (Coyne 1985). This sterility effect was closely linked to the fork (f) marker, and mapped the approximate location of this major effect hybrid sterility factor to 1.1cM on either side of f (Coyne and Charleswoth 1986). By ways of the chromosome introgression from D. mauritiana to D. simulans, the factor was further mapped between f and Bx, in cytological interval 16D and namely Odysseus (Ods) (Perez et al. 1993). Result from a series of recombinants, the function of molecular factor in Ods were
determined precisely. The longest fertile introgressions and the shortest sterile introgressions defined the minimized Ods gene within a genomic clone. Finally, Odysseus was cloned as a 340-amino-acid protein and predicted as a transcription factor with a paired-type homeodomain, a DNA binding motif, encoded by exon2 and 3 (Ting et al. 1998), therefore, it was named OdsH (for Ods-site homeobox gene).
According to previous investigation, OdsH is known to be a duplicated gene from a neuron gene unc-4, and is 13-kb apart in tandem with it (Ting et al. 2004).
Both OdsH and unc-4 consists of four exons with a paired-like type homeodomain on exons 2 and 3 sharing over 70% homology, and the structure of intron and exon are preserved (Ting et al. 2004). In Drosophila, both sequence and expression pattern of unc-4 is conserved. In Drosophila, unc-4 express in the
developing neuron system, female ovaries and male testes, while OdsH alleles are highly diverged not only in the sequential codon variation but also the expressing patterns among D. melanogaster subgroup including D. mauritiana, D. melanogaster, and D. simulans (Ting et al. 1998). In situ hybridization data showed that the OdsH is specifically expressed in the reproductive organs (Ting et al. 2004), and the
expression pattern of OdsH in hybrids was mis-regulated (Sun et al. 2004). The functional studies in D. melanogaster, revealing that loss of OdsH would cause defect in male fertility at younger age, 2 to 4 days old, suggested that the normal function of OdsH is to enhance the fertility of young males by accelerating the sperm maturing (Sun et al. 2004). Furthermore, overexpression of OdsH has been shown to promote male fertility that more offspring production than wild type do, and the null mutant of OdsH can reduce the male fertility (Chen 2005). Therefore, the function of OdsH was suggested to accelerate the sperm maturation and enhance the fertility of the males.
Even there have several studies in the OdsH, now still need to gain more understand about the molecular mechanism for the function of OdsH, to solve the questions that how does OdsH play a role in spermatogenesis and what’s the mechanism for OdsH to cause hybrid male sterile between D. mauritiana and D.
simulans.
To address this question, ectopic expression - gene express in a place where it is normally inactive, is a powerful technique to gain further understanding of the gene function. In Drosophila, ectopic expression can be done by introducing a transgene with a modified promoter into the target organism or using the Gal4/UAS system. By ectopic expression tests with the UAS/Gal4 system, I want to do the analysis between three OdsH alleles, and to clarify the molecular mechanism of OdsH gene. Several Gal4 lines were chosen to induce three OdsH alleles expression in different tissues, by
crossing with UAS-OdsHconstructs: elav-Gal4, expressed in the nervous system;
GMR-Gal4, expressed posterior to the eye disc morphogenetic furrow (MF) in the third instar larvae; nanos-Gal4, expressed in male and female germline; and the ubiquitous expression line, tub-Gal4. In order to avoid the position effect in the expression analysis and to express the OdsH in the germline, the site specific transgenic vector that workable in the germline is necessary to be generated (Appendix I).
All of these ectopic expression lines give phenotypes. Among them, there are two lines deserve to do further analysis. The first one is the nanos>OdsH lines that have OdsH expressed in the place that similar to the endogenous expression. Second, the GMR>OdsH line, since Drosophila eye development is a process between cell division and differentiation such as the spermatogenesis in testes. It is desired that these experiment results will provide insight to the molecular mechanism of OdsH gene and the mechanism for OdsH to cause hybrid male sterility between D.
mauritiana and D. simulans.
In addition to the molecular mechanism for the function of OdsH. As we have known that OdsH was duplicated from the unc-4 gene. Find out the normal function of OdsH and unc-4 will improve our understanding about the correlation between the duplication genes and speciation. unc-4 was first isolated as a novel Drosophila pair- liked homeobox gene, DPHD-1, the homeodomain of DPHD-1 showed 85% amino- acid similarity to the C. elegans Unc-4 protein. In situ hybridization data of embryos and third-instar larvae revealed that the DPHD-1 mRNA is localized in subsets of postmitotic neurons in the central nervous system (CNS) and in the developing epidermis with a segmentally repeated pattern (Tabuchi 1998). In Caenorhabditis elegans, UNC-4 homeoprotein and Groucho-like corepressor UNC-37 were showed
function together in VA motor neurons to repress VB-specific genes that specify synaptic choice motor neuron circuit (Winnier et al. 2007). However, the normal function of the unc-4 in the Drosophila is remained unknown. To comprehend the normal function, the microRNAs system in the Drosophila was applied to create the unc-4 mutant (Appendix II).
In this study, the first aim is to reveal the molecular mechanism for the function of OdsH. To deal with this, three alleles of OdsH were analysis by ectopic expression examination. This may provide enlightenment in understanding the male hybrid sterility defect between D. mauritiana and D. simulans. Second, I want to identify the normal function of unc-4 and to answer the question that whether OdsH have evolved novel function after the duplication event. With this, our understanding about these pair of duplication genes will be improved.
Materials and methods
Fly stocks
UAS-OdsHmel line carrying pP {5’-UAS::OdsHmel} construct was constructed by Yi-Lin Chen and Kah-Junn Tan (2005). Two attP insertion stocks, ZH-attP-51D (y w M{eGFP.vas-int.Dm}ZH-2A; M{RFP.attP}ZH-51D) and ZH-attP-86Fa (y w M{eGFP.vas-int.Dm}ZH-2A; +; M{RFP.attP}ZH-86Fa) were used to generate site specific transgenic lines (Bischof et al. 2007). Four Gal4 lines including elav-Gal4 P{w+mW.hs =GawB}elavC155 (Lin and Goodman 1994), GMR-Gal4: w*; P{w+mC
=GAL4-ninaE.GMR}12 (Freeman 1996), nanos-Gal4: w1118; P{w+mc =GAL4::VP16- nos.UTR}MVD1 (Van Doren et al. 1998), and tub-Gal4: y1 w*; P{w+mC =tubP-
GAL4}LL7/TM3, Sb1 (Lee and Luo 1999) were obtained from the Bloomington stock center. The multiple balancer line, w1118; Cyo/Sp; TM3/Sb, was used to set up the transgenic stocks. The His-2AvD GFP flies: P{His2AvT;Avic/GFP-S65T}62A, Df(3R)nm136 were used for expression analysis (Clarkson and Saint 1999). All stocks were kept at 25°C and raised on the standard cornmeal medium.
Transgenic constructs
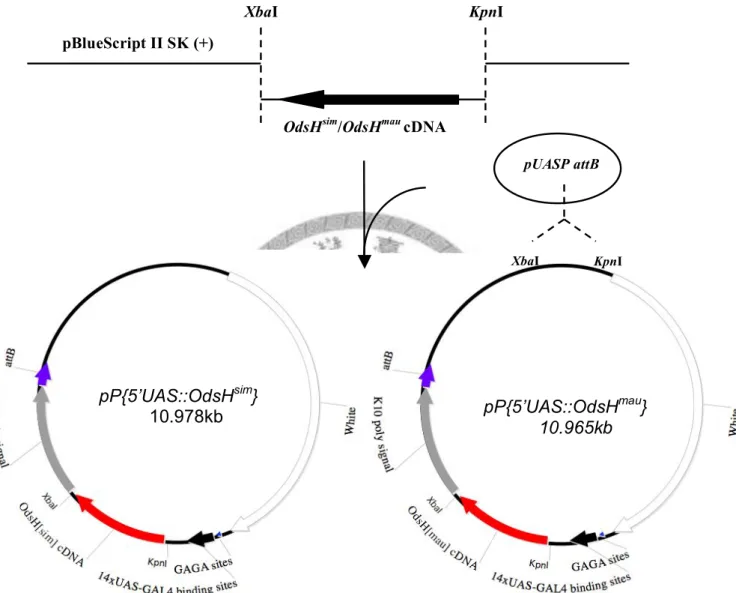
The cDNA fragments of OdsHsim and OdsHmau were digested from pBSKII (+)-OdsHsim and pBSKII (+)-OdsHmau by the restriction enzymes XbaI and KpnI, and subcloned into the transgenic vector pUASP attB. The transgenic constructs were named pP{5’-UAS::OdsHsim} and pP{5’-UAS::OdsHmau} respectively (Fig. 1).
Transgenic lines
Two constructs pP{5’-UAS::OdsHsim} and pP{5’-UAS::OdsHmau} with attB site, were microinjected into ZH-attP-51D and ZH-attP-86Fa respectively (thanks for help of Chiou-Yang Tang). After homozygous transgenic stocks were established on
the second or third chromosome, the transgenic flies were crossed to different Gal4 lines (Fig. 2 and Fig. 3).
Fig. 1.—Procedure of pP{5’-UAS::OdsHsim} and pP{5’-UAS::OdsHmau} constructs. Dashes mean the digestion by the nearby restriction enzymes. The cDNA of OdsHmau and OdsHsim were cut out with KpnI and XbaI from the vector pBluescript II SK(+) and cloned into the site specific integration vector pUASP attB to get the UAS constructs.
pP{5’UAS::OdsHsim}
10.978kb pP{5’UAS::OdsHmau}
10.965kb
XbaI KpnI
pBlueScript II SK (+)
OdsHsim/OdsHmau cDNA
XbaI KpnI pUASP attB
injection
!
G0 male or female x w1118 male or female
! G1 select red-eyed flies
+/P ; +/+
male or female
x double balancer male or female CyO/Sp ; TM3/Sb
!"
G2 select flies with CyO marker CyO/P ; TM3/+
male or female!
x CyO/P ; TM3/+
male or female
! !"!
G3 select flies without CyO marker P/P ; +/+
male or female!
x different Gal4 lines male or female
! #
For ectopic expression analysis
Fig. 2.— The cross steps to get ectopic expression line on second chromosome.
injection
!
G0 male or female x w1118 male or female
! G1 select red-eyed flies
+/+ ; +/P male or female
x double balancer male or female CyO/Sp ; TM3/Sb
!"
G2 select flies with TM3 marker +/CyO ; TM3/P
male or female!
x +/CyO ; TM3/P male or female
! !"!
G3 select flies without TM3 marker +/+ ; P/P
male or female!
x different Gal4 lines male or female
! #
For ectopic expression analysis
Fig. 3.—The cross steps to get ectopic expression line on the third chromosome.
RNA extraction
For the total RNA extraction, 0-5 day-old adult flies were collected. Before RNA extraction, flies were put into an empty vial to void most of yeast for at least 2 hours. Total RNA was extracted with the TRIzol® reagent followed the
manufacturer's instructions (Invitrogen). The extracted RNA were then treated with the DNase for 15 min at 25°C. After inactive the DNase by the EDTA for 10 min at 65°C, RNA was reverse-transcribed. The left RNA was stored in DEPC water at -80°C.
RT-PCR
For Reverse Transcription (RT), 5µg total RNA was used as the template for cDNA synthesis. The SuperScript II First-Strand Synthesis system with a mixture of Oligo-dT (Invitrogen), dNTPs (10 mM), RNA (5 μg), and DEPC- ddH2O was incubated for 5 min at 65°C. The RT contents were collected at the bottom by centrifuging, and 5X RT buffer, 25mM MgCl2, 0.1 M DTT, RNAse OUTTM, Superscript II RT were added. For PCR reactions, the reactions contained 8ng template, 1X reaction buffer, 1.5mM MgCl2, 16nM dNTP, 1 unit Taq polymerase, and 40nM primers each in a final volume of 20µl were amplified for 30 cycles of 30s at 95°C, 30s at 58°C, 1min 20s at 72°C, and the final extension at 72°C for 7min.
The PCR amplification products were analyzed by electrophoresis in 1.5% agarose gels. Drosophila ribosomal protein 49 encoding gene (rp49) was used as a loading control for the PCR reactions. Gene specific and rp49 primers for RT-PCR were listed in Table 1.
Table 1
The list of primers for expression analysis.
Gene Primer Direction1 Sequences
rp49 rp49F F 5’CAGTCGGATCGATATGCTAAGCTGT3’
rp49R R 5’TTACCGACCTTGGGCATCAGATACT3’
OdsH OdsH c0+ F 5’GCAAAAGCTAAGACGAAAATGGAT3’
U8 4524- R 5’TGCTTAGCTAACCACCCGAAATCA3’
unc-4 unc-4-552+ F 5’AGCCCGTCGTCGAAAGAAGC3’
unc-4-739- R 5’TTCGCCCTCGTCGTCGCTGTC3’
1“F”, forward primers; “R”, reverse primers.
Embryo collection
Adult flies were transfer into a plastic bottle with a grape juice agar plate. The prelaying time is 45mins each twice. After prelaying, change new plate for collecting embryos. Embryos were collected from the grape juice agar plate with dH2O into a Nitex nylon mesh (Cat. 3-100/47, Tetko Inc) and dechorionated with 50% bleach for about 5mins. These dechrionated embryos were washed with dH2O again and transfer into a glass bottle containing 1:1 n-heptane: 4 % paraformaldehyde for 20-30min fixation. Remove the bottom aqueous layer, and add 100% methanol with the same volume and shaked for 1 min vigorously. The devitellinized embryos will at the bottom of the bottle. Remove the heptane and wash several times with 100%
methanol. Embryos can be stored in the methanol and kept in -20°C (Theurkauf 1992).
Testes and ovary dissection
To observe the reproductive organs, the testes and ovaries were dissected in the 1XPBS, and fixed in the 2% Glutaraldehyde (Sigma) for 5min. After this,
mounting samples onto the glass slide in 1XPBS solution (White-Cooper et al. 1998).
Pictures were taken with the fluorescent microscopy: Zeiss Axio Imager A1.
Scanning electron microscopy (SEM)
Adult flies were fixed in 70% ethanol for 1hr, dissecting the heads
dehydration. Incubated the heads in serious dilution ethanol, including 85%, 90%, and 95% ethanol for 30mins each. Wash with 100% ethanol twice for 30mins each and acetone twice for 30mins each. Samples are dried after treating in critical point dry (CPD) specimen. After mounting samples on the studs, put them into the chamber of the sputter coater and coated with a thin golden film (Domínguez et al.
1998). These eyes were examined by a scanning electron microscope: FEI INSECT S. and images were acquired using the xT microscope server software.
Immunostaining
Dissecting eye-antennal discs from third instar larvae in 1X PBS, then transfer into fixation solution (4% paraformaldehyde in 1X PBS) for 15mins at room
temperature. Washing the fixed eye-antennal discs with 1XPBST (1X PBS+ 0.3%
Triton-100) three times for 5 mins each, then incubate in the 1XPBST contain 3%
BSA with rabbit anti-phosphohistone H3 (1:800) or rat anti-elav (1:500) antibody overnight at 4°C. Following, the samples were washed with 1X PBST three times and incubated with secondary antibodies conjugated with either FITC or Rodamin for 2hrs in dark at room tempterture. The discs were then washed 3 times with 1X PBST before mounting (Pai et al. 1998).
To label the S phase cell at the second mitotic wave, eye discs were dissected in 1XPBS and incubated in PBS containing 0.1 mg/ml of Bromodeoxyuridine (BrdU) for 40 mins. Discs were fixed with 4% para-formaldehyde for 30 mins, then treated with 3N HCl for 15 mins. After washing by PBST for three times and incubated with mouse anti-BrdU antibody in PBST containing 3% bovine serum albumin (BSA) at
4°C overnight, the BrdU signal was detected by secondary antibodies conjugated with FITC (Secombe et al. 1998).
All discs were mounting discs on the glass slide with their apical face up in mounting solution and pictures were taken with a confocal spectral microscope system: Lsica TCS SP5 and the LAS AF software.
Results
The OdsH transgenic lines
In order to study the molecular mechanism of the OdsH, set up OdsH transgenic flies for the subsequent ectopic expression analysis. Flies carrying UAS- OdsHmel construct was created by Yi-Lin Chen, and OdsHsim and OdsHmau each were cloned into the pUASP attB transgenic vector for germline expression. pP{5’-
UAS::OdsHsim} was injected into 29 embryos for each attP lines, and 11 larvae of ZH- attP-51D and 17 larvae of ZH-attP-86Fa were collected. Finally, one transformant on the second chromosome and three transformants on the third chromosome were obtained. For pP{5’-UAS::OdsHmau}, 30 embryos were injected for each lines and 11 larvae of ZH-attP-51D and 17 larvae of ZH-attP-86Fa attP lines were collected. Only one transformant of each chromosome was obtained (Table 2). The third
chromosome insertion lines for each UAS construct was used to cross with different Gal4 lines for ectopic expression analysis.
Table 2
The list of UAS-OdsH transgenic lines Numbers of
injected embryo Numbers of
hatched larvae Transformant UAS-OdsHsim
ZH-attP-51D 29 11 1
ZH-attP-86Fa 29 17 3
UAS-OdsHmau
ZH-attP-51D 30 11 1
ZH-attP-86Fa 30 17 1
Ubiquitous and neuron expression of OdsH alleles
According to previous investigation. OdsH is known to be a duplicated gene from a neuron gene unc-4. Expression sites of unc-4 can be in neuron cells, female ovaries and male testes. However, OdsH is notified highly expressed in testes but not
in neuron cells or female ovaries (Ting et al. 2004). Divergence function of OdsH from unc-4 is still unknown. In order to understand the molecular mechanism for the function of OdsH, different OdsH alleles from the Drosophila species: D. mauritiana, D. melanogaster, and D. simulans were overexpressed at several specific tissues by the UAS-Gal4 system here, and the result is showed in Table. 3.
Table 3
Phenotypes of ectopic expression of OdsH alleles.
Expression site UAS-OdsHmel UAS-OdsHmau UAS-OdsHsim elav-Gal4 neuron cell embryonic lethal no defect no defect GMR-Gal4 eye disc small and smooth
eye
small and rough eye
small and rough eye nanos-Gal4 germ line cell Female sterile
(maternal effect embryonic lethal) male fertile
female and male sterile
female and male fertile
tub-Gal4 ubiquitous larval stage lethal no defect no defect
First I used tub-Gal4 to drive three OdsH alleles for ubiquitious expression.
The phenotype of flies with tub>OdsHmel showed larval stage lethal. In addition, I used elav-Gal4 to make the OdsH alleles expressed in neuron cells. In elav>OdsHmel, all offspring died at the embryonic stage and no larvae and adults could be observed.
These results indicated that misexpress of OdsH would influence the early stage development of Drosophila.
In addition to the effect of OdsHmel, OdsHmau and OdsHsim were also analysis by two Gal4 lines, tub-Gal4 and elav-Gal4. Both the expressions of OdsHmau and OdsHsim have no clear effects in the developmental process and many adult flies can be obtained.
Germlines expression of OdsH alleles
OdsH is a rapid evolved gene that the amino acid is highly diverged among D.
mauritiana, D. melamogaster, and D. simulans. The endogenous OdsH is highly expressed in the male reproductive system, but loss of the expression in the female germline where is one of the expression site of unc-4. Here, I analyzed the influence of OdsH in both male and female germlines by using nanos-Gal4. Morever, it is also interesting to investigate the sequence divergence of the OdsH alleles in the germline system of D. melanogaster.
In order to check the germline expression by using the site specific transgenic vector pUASP attB, which I have modified from the pUAST attB. OdsH expression was detected by RT-PCR. Primers specific for OdsH were used. The result showed that OdsH was expressed in the females which carry either nanos> OdsHsim or nanos>
OdsHmau but no expression in the females from w1118 (Fig. 4). Therefore, the vector pUASP attB was successful to induce gene expressed in female germlines.
Fig. 4.—RT expression of OdsH in males and females. Males RNA were extracted from three different lines, including w1118 (w), nanos>OdsHmau (M), and
nanos>OdsHsim (S). While in females, only the flies with OdsH overexpression in germline cells have the OdsH product after RT-PCR procedure. rp49 was used as the control gene in all samples.
Females from three nanos>OdsH lines showed different phenotypes.
nanos>OdsHmel led to sterile phenotype of females, but the morphology of the ovary looked similar to the one from the w1118. Females with OdsHmel expression in the germline have normal form egg chamber, which contains ovum in various stages.
W M S W M S
Males Females
rp49 OdsH
More specifically, female manage to lay eggs, but all of them died at the embryonic stage. Females with OdsHsim expressed in germlines have no clear defect in fertility.
The morphology of egg chambers in the ovaries are also normal. However, females with OdsHmau expressed in germlines are sterile and cannot produce embryos.
Observation the ovaries dissected from these females under the microscope showed that they are much smaller than those in the adults carrying UAS-OdsHmel and UAS- OdsHsim (Fig. 5).
Fig. 5.—Gal4 inducible expression in the germline of the ovary. (A) The ovary from w1118 contains many egg chambers in it, and every female has two ovaries in the abdomen. The morphology of ovaries from females carrying with nanos>OdsHmel and nanos>OdsHsim are similar to w1118. (B) In nanos>OdsHmau famels, there are two ovaries can be seen but the size of the ovaries are much smaller than that in w1118. Besides, the structure are abnormal that without any egg chambers. Various stages of the ovum can be seen in the egg chambers from the ovaries of the females in w1118 (C), in nanos>OdsHsim (D), and in nanos>OdsHmel (E).
To figure out the defect in development leading to embryonic lethality in nanos>OdsHmel line, females with nanos>OdsHmel genotype were allowed to lay eggs for 30 minutes, and the embryos were collected. In general, embryos would go
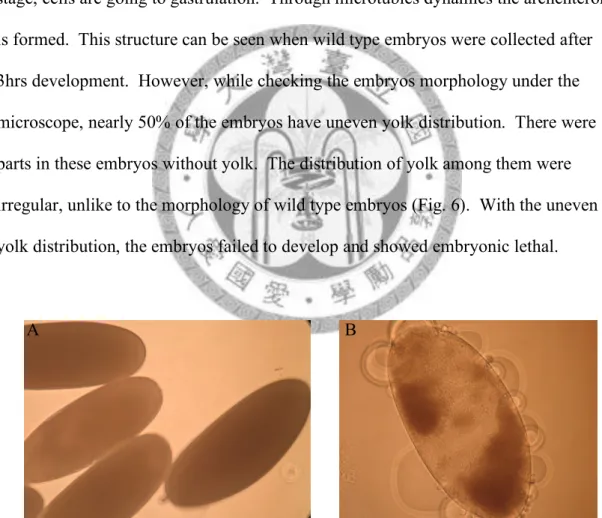
through the syncytial blastoderm stage at about 1hr 30min to 2hr 30min. In this stage, the nuclei in the embryos will divide four or more times syncytially. Immediately after the last division, the nuclei migrate to the surface and begin to partition into individual cells. Thus, when collecting the wild type embryos at 1 hr 30min, the individual nuclei can be seen clearly and the nuclei spaced evenly. Following this stage, cells are going to gastrulation. Through microtubles dynamics the archenteron is formed. This structure can be seen when wild type embryos were collected after 3hrs development. However, while checking the embryos morphology under the microscope, nearly 50% of the embryos have uneven yolk distribution. There were parts in these embryos without yolk. The distribution of yolk among them were irregular, unlike to the morphology of wild type embryos (Fig. 6). With the uneven yolk distribution, the embryos failed to develop and showed embryonic lethal.
Fig. 6.—The morphology of the early embryos. Embryos were collected in 30min.
(A) In w1118, all the embryos have even yolk distribution under microscope. (B) In nanos>OdsHmel , nearly 50% of the eggs produced from the females, with OdsHmel expressed in germline, have uneven yolk distribution. Part of the eggs are empty under microscope.
A B
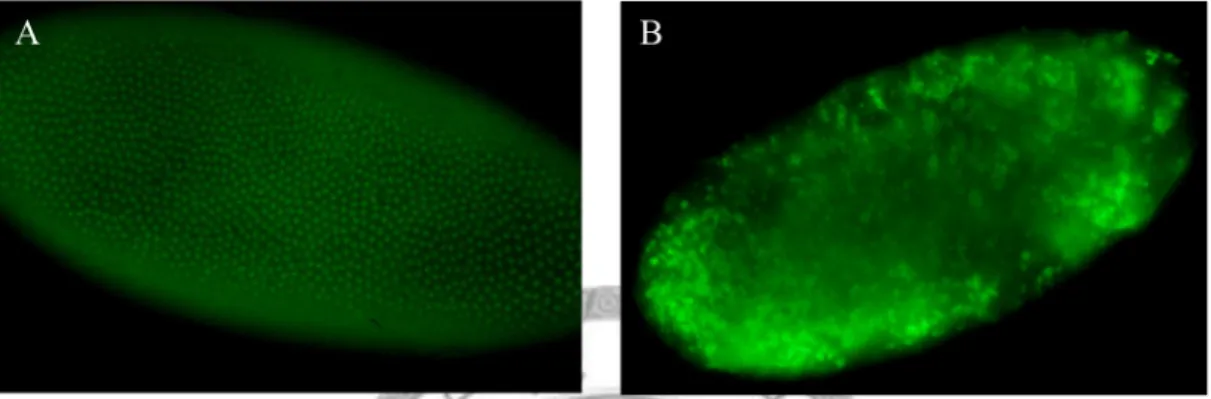
When the nuclei of the embryos were labeling by GFP, with the homozygous line of nanos>OdsHmel crossed to the His-2A GFP flies, the signal indicated that the nuclei are not spaced evenly. Many of the nuclei overlap with each other (Fig. 7).
Additionally, very high percentage of the mutant embryos could not form the archenteron successfully and caused a twisted phenotype (Fig. 8).
Fig. 7.—Nucleus pattern in the embryos. Embryos were collected in 1.5hr from:
w1118 and nanos>OdsHmel. (A) In w1118, the embryos are at the late time of the syncytial blastoderm (about stage 4) and begin moving into the cellularization stage.
Under the microscope, the individual nuclei are visible and distribution uniform. (B) In the embryos from the females with nanos>OdsHmel, the individual nuclei cannot be seen clearly since they overlap to each other.
Fig. 8.—Embryos in the gastrulation stage. Embryos were collected in 3hrs from:
w1118 and nanos>OdsHmel. (A) In w1118, the gastrulation stage of the embryos begin as soon as the cell formation finish, and form the structure of the archenteron. (B) In the embryos collected from the females with nanos>OdsHmel, the force from the microtubules dynamic cannot induce cell migration to form archenteron structure successfully and result in twist phenotype. Arrow shows the twist part in the embryo.
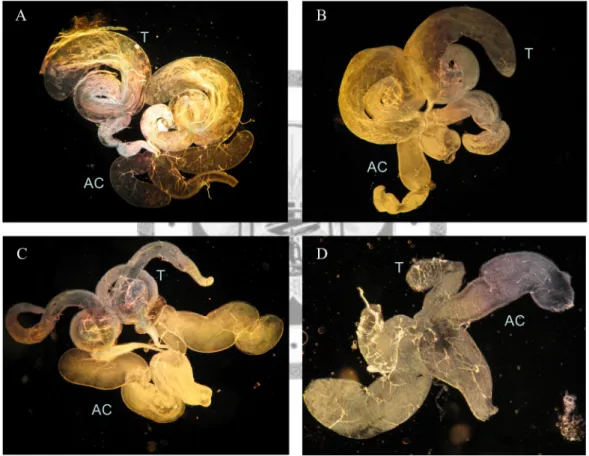
Males in nanos>OdsHmel and nanos>OdsHsin lines do not appear to have reproductive defects with the ability to produce offspring and both the testes and accessory gland structures were normal when compare to w1118. On the contrary, when males carrying nanos>OdsHmau cross to the w1118 females, there were no larvae due to the embryonic lethal phenotype. Besides, there are critical defect in the testes development with OdsHmau expression (Fig. 9).
Fig. 9.—The morphology of the male reproductive organ. Dark field of the male germline tissue. The testes (T) and accessory gland (AC) structures are clear to be seen in (A) w1118, (B) nanos>OdsHmel, and (C) nanos>OdsHsim. While the testes wither in all the males of nanos>OdsHmau in (D).
Eye disc expression of OdsH alleles
Drosophila eye development involves cell division and differentiation similar to spermatogenesis in the testes where OdsH is normally expressed. In addition, many signaling pathways that regulate eye development are well identified.
According to the observation in germline that the expression of OdsHmel in female germlines would cause embryonic lethal with the abnormal nuclei pattern.
Drosophila eye was applied to test whether OdsH plays a role in cell cycle regulation.
GMR-Gal4, which expresses posterior to the morphogenetic furrow in the eye discs of the third instar larva, was chosen to drive OdsH alleles expression.
Morphology of adult eyes
The transgenic flies with UAS-OdsHmel were crossed to GMR-Gal4 to induce the expression of OdsH during eye development. All the eyes of these flies have smaller size than the normal eye and have shuttle-liked phenotype. Besides, the eye surface is very smooth when compared to the three-dimensional compound eye structure in the w1118, as the control line. According to the Scanning electron
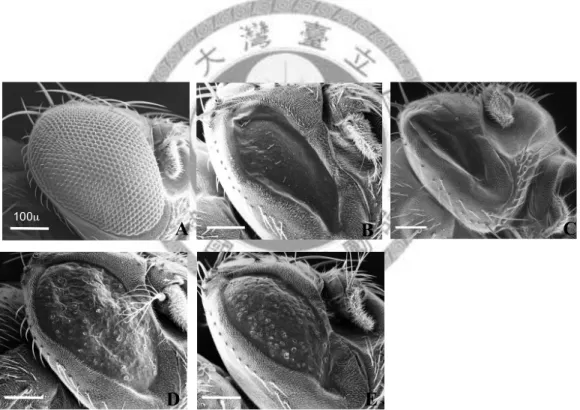
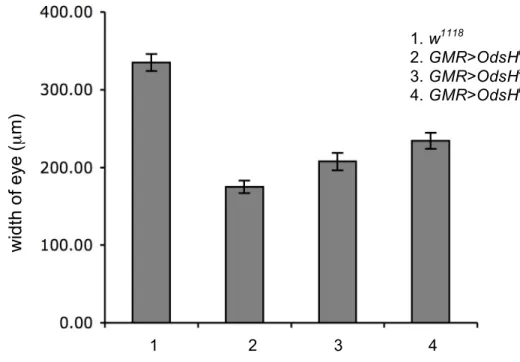
microscopy (SEM) pictures, the hexagon ommatidia that form the compound eye with uniform disturbed bristles were clear to be seen, and the width of the eye was
measured. The width range of the eye from w1118 is 334 µm and decrease severely to 175 µm in the adults with GMR>OdsHmel. Flies carrying homozygous OdsH
expression in the eye discs, the eye size will be reduce to 133 µm. In addition, there are not any ommatidia structures and fewer bristles contained in both heterozygous and homozygous lines (Fig. 10B and 10C).
In addition to the OdsHmel, the defects induced by different alleles of OdsH were also observed in the eye development. Driving OdsHmau and OdsHsim expression by GMR-Gal4 in the larva eye discs also showed the critical eye phenotypes. Both of
the eyes from GMR>OdsHmau and GMR>OdsHsim are smaller than normal ones.
Under the SEM, mean width of the eye been measured is about 224 µm from GMR>OdsHmau and 207 µm of the eye from GMR>OdsHsim. Both two alleles of OdsH cause the eyes become rough and lose the ommatidia structure. Some
individuals showed few bristles and some individuals have no bristles. Thus, the eye size is smallest with OdsHmel expressed in the third instars larva eye discs when
comparing to the eye from GMR>OdsHsim and GMR>OdsHmau (Fig. 10). Additionally, size of the eyes showed a different level decrease in three different GMR-OdsH lines (Fig. 11).
Fig. 10.—The morphology of adult eye with ectopic expression different alleles of OdsH. The eye typr in (A) w1118, (B) GMR>OdsHmel heterozygous strain, (C)
GMR>OdsHmel homozygous strain, (D) GMR>OdsHmau, and (E) GMR>OdsHsim are showed in SEM. Scale bar = 100µm.
D
100µ
m A B C
E
Fig. 11.—The width of eye with different OdsH alleles expressed. Bar graph showing the width of adult eyes with OdsH express (mean ! SE). The mean value in w1118 is 334µm, 175µm in GMR>OdsHmel, 207µm in GMR>OdsHsim, and 224µm in GMR>OdsHmau. All the statistical analysis were compared to GMR>OdsHmel (Student’s t-test, P<0.05).
The influence of OdsHmel in mitotic cells
According to the observation in the critical eye phenotype that caused by the ectopic expression of OdsHmel, I did the antibodies staining to see whether it was a consequence of the defect in the cell-cycle regulation. Neuron clusters and mitotic cells were detected by anti-Elav and anti-phosphohistone3 antibodies. Elav is to detect the neuron clusters and phosphohistone3 can label the cells that undergo mitosis. Comparing the eye discs from w1118 and GMR;OdsHmel, the mean columns and arrangement of the neuron clusters are not significant different. For my results, the mitotic cell numbers at the second mitotic wave (SMW) that posterior to the morphogenetic furrow were increase in GMR>OdsHmel (Fig. 12). The cell numbers at the SMW of the eye discs from GMR>OdsHmel contained about 65 (n=30) and there are about 51 cells (n=19) in the disc from w1118. According to the Student’s t-test, they showed statistically significant difference (Fig. 13).
width of eye (µm)
1. w1118
2. GMR>OdsHmel 3. GMR>OdsHsim 4. GMR>OdsHmau
1 2 3 4
Fig. 12.—The mitotic cell with OdsHmel expression. Anti-phospho-histone H3 antibody labeled the mitotic cells at the first and second mitotic wave, and anti-Elav antibody revealed the neuron clusters. The mitotic cell number at second mitotic wave in (A-B) w1118, and were increased in (C-D) GMR;OdsHmel. Arrows indicated the furrow. Scale bar = 50 µm.
Fig. 13.—Quantification of the mitotic cells at the first and second mitotic waves.
Bar graph shows the mitotic cell numbers (mean !"SE) at the second mitotic wave in the w1118 and GMR>OdsHmel. The mitotic cell numbers at the second mitotic wave are significant different between two lines (Student’s t-test, P<0.0001).
D
Elav PH3 C
PH3 B
Elav PH3 A
PH3
mitotic wave
number
*
GMR>OdsHmel w1118
The increase of the mitotic cells at the SMW could be due to the mitotic defects that happened in the SMW to cause the accumulation of arrested mitotic cells poster to the morphogenetic furrow, or multiple rounds of cell division occurring poster to the furrow. In order to distinguish between these two hypothesis,
Bromodeoxyuridine (BrdU) which is synthetic thymidine analog that can replace the space of thymidine and incorporated into the replicating DNA was applied to label the S phase cells at the SMW. The distribution and the number of the BrdU-labeling cells are similar between w1118 and GMR>OdsHmel (Fig. 14). This result indicated that there are not more cells undergoing multiple rounds of cell division in GMR>OdsHmel, and the mitotic cell numbers increase in the GMR>OdsHmel was caused by the mitotic defect.
Fig. 14.—S phase cell in third-instar larva eye disc. The cells were labeled with anti-BrdU antibody at the SMW in (A) w1118 and (B) GMR>OdsHmel eye discs.
Arrows indicated the morphogenetic furrow. Scale bar = 50 µm.
Discussion
In order to gain more insight in the molecular mechanism for the function of OdsH and find out the possible mechanism of the hybrid male sterility between D.
mauritiana and D. simulans. Ectopic expression analysis by the UAS/Gal4 system was applied. The site specific transgenic vector that was workable in the germline cells and named pUASP attB were generated first. This vector overcomes two limitations for the original transgenic vector pUAST: the insertion of the UAS construct is random that would have the position effect in the phenotypic analysis, and the activity of pUAST is poorly in the germline (Brand and Perrimon 1993). The modified vector called pUASP were generated and was shown to drive efficient expression in germline (Grossniklaus et al. 1989; Spardling 1993), but it still have the problem with the random insertion. The pUASP attB vector, which I have generated here was modified from the site specific transgenic vector called pUAST attB
(constructed by Johannes Bischof and Konrad Basler). The UAS constructs made with pUASP attB will have the specific integration site in the Drosophila genome when injected into the attP lines, by the recombination between the attB sequence in the vector and the attP sequence in the Drosophila genome (Fish et al. 2007). In site specific integration, the genome mapping procedure to check the insert site is
necessary, additionally, it can avoid the position effect when comparing different insertion lines. Second, pUASP attB works well in germline cells that it contains the important motifs from the traditional germline cells workable transgenic vector pUASP. UAS constructs with different alleles of OdsH in three Drosophila species including D. melanogaster, D. simulans, and D. mauritiana were then generated.
According to the ectopic expression analysis, the overexpression of OdsH in
this defect were not observed in the elav>OdsHsim or elav>OdsHmau embryos. In the germline expression, the OdsH allele from D. mauritiana caused defects of the reproductive organs and result in male and female sterility, while OdsHsim have no effect in the development of the testes and the ovaries. The females with OdsHmel expression were sterile with the embryonic lethal phenotype, but males with OdsHmel expression were fertile. In Drosophila eye development, all three alleles of the OdsH expression caused the decrease in the eye size with the gradient difference. The immunostaining results show that the mitotic cells were interfered with the OdsHmel expression.
OdsH in D. melanogaster germline cells
The expression of the OdsH allele from D. mauritiana causes the reproductive organs in both males and females withered in the D. melanogaster genetic
background, and there were no gametes produced. However, this phenotype was not appearing in the flies that expressed OdsH alleles from D. melanogaster or D.
simulans. These results suggested that the OdsH allele of the D. mauritiana is largely different from it in D. melanogaster and D. simulans.
OdsH, which contains two important motifs including homeodomain and Engrailed homology 1 (Eh1) domain, is a rapidly evolving gene that the sequences are highly diverged among D. mauritiana, D. melanogaster, and D. simulans (Ting et al.
1998). Among diverse texa, homeodomain is very conserved, but according to the sequence alignment of the OdsH alleles in the three species, many amino acids subtitution between three species. In the OdsHmau, there are eight amino acids different from OdsHmel and OdsHsim. Among these eight amino acids, four residues have changed the polarity, residues 152 and 157 have become non-polar amino acid, and residue 196 changed to the polar amino acid. This property is important in
protein structure and protein-protein interaction. Therefore, these amino acids can be the candidates contribute to the different phenotype in germline expression. On the other hand, Engrailed homology 1 (Eh1) domain may be another possible region to cause the variance.
Among Drosophila species, the Eh1 domain is conserved, OdsH from D.
mauritiana has a premature stop codon resulting in the loss of the Eh1 domain function (Fig. 15). Eh1 domain is the repression domain first identified in the
Drosophila Engrailed homeodomain protein, and it has been reported to interact with Groucho protein (Riz et al. 2009). Groucho proteins are widely expressed nuclear factors, which have no DNA-binding activity. By interacting with DNA-binding factor to influence its function, many patterning and differentiation events of the development are regulated (Buscarlet and Stifani 2007).
To synthesize, according to the phenotype of germlines expression and the information from the sequence alignment between D. melanogaster, D. simulans and D. mauritiana. There are three possible regions in OdsHmau to play important roles in the function of OdsH to affect the fertility in the D. melanogaster genetic background.
Furthermore, result in the developmental defect of the reproductive organs. The regions are: seven amino acids substitution in the homeodomain, and the Eh1 domain.
*
Fig. 15.—The alignment of three OdsH alleles. The homeodomain is labeled by the red under line, and the eh1 domain is labeled by the black under line.
30
In the nanos>OdsHmel lines, only females have fertility defect but not males.
The embryos produced from the females with the OdsHmel expression have developmental defect. In these embryos, the nuclei cannot divide to individual nucleolus in the syncytial blastoderm and overlap to each other. It suggested that the expression of the OdsHmel may influence the cell cycle procedure. While the
expression of OdsHalleles from D. simulans seems has no defect in the males and females fertility. In addition, the morphology of the reproductive organs are similar to the ones from w1118 in both males and females. It may due to the lower expression level that can be seen in the RT-PCR, and may be there are subtle fertility effect in nanos>OdsHsim.
According to the germlines analysis, OdsHmel may play a role in cell cycle regulation. To confirm this hypothesis, Drosophila eye development provides a good model system.
OdsH expression in Drosophila eye
The defect of the eye phenotype between heterozygous and homozygous strains with OdsHmel expression showed the correlation with the mutant eye
phenotype and the dosage of the OdsH allele. Only one copy of OdsH is sufficient to interference eye development and reduced adult eye size. As in Fig. 10.
GMR>OdsHsim and GMR>OdsHmau flies also have small eye phenotype and lose normal ommatidia structure. The degree of the eye defect, by measuring the width of the eyes from three ectopic expression lines, is correlated to the sequence divergence.
The nonsynonymous (Ka) and synonymous (Ks) ratio in OdsH of the five species in Drosophila shows that the ratio is 0.649 between D. melanogaster and D. simulans, 0.812 between D. melanogaster and D. mauritiana, and the highest ratio is between D.
D B C
Elav
A B C D
Elav
A
mauritiana and D. simulans, in a value of 2.542 (Ting et al. 2004). OdsH from D.
melanogaster have the strongest effect in eye development than D. simulans and the minor one from D. mauritiana. This result is consistent to previous observation in male fertility test. Under D. melanogaster background, OdsHsim and OdsHmau alleles also can increase male fertility like OdsHmel do, but the average influences are
OdsHmel > OdsHsim > OdsHmau (Chen 2005). Combine these analysis, the divergence of the sequence might influence the intensity of OdsH to implement its function.
OdsH influence the M phase of the cell cycle
The adult eyes of Drosophila are developed from monolayer epithelia, called eye imaginal discs. Beginning with third-instars larva, a wave of cells called
morphogenetic furrow (MF) sweeps across the eye disc from the posterior to the anterior. Cell posterior to the furrow divide and differentiate to form the ommatidial preclusters. The narrow band posterior to the MF arrest in G2, named second mitotic wave (SMW). In this wave, the cycling cells will enter the synchronized cell division process when receive the signal from ommatidia (Ready et al. 1976; Carthew 2007;
Wolff 1991). According to the immunostaining, the mitotic cell numbers at the second mitotic wave were increased, when expressed OdsHmel in the third-instar larva, compared to the index of w1118. Since the eye development of Drosophila is tightly regulated between cell division and differentiation. Genes that interfere with cell- cycle regulation will interrupt cell differentiation and resulting in different adult eye phenotypes.
In the normal eye development, it takes about 15 mins for cells at the SMW undergo syncitial cell mitotic. If GMR>OdsHmel inducing more cells to replicate, the mitotic cell index will increase. However, this is not the case in the OdsH expression.
The S phase cell numbers were similar in the eye discs no matter the OdsHmel express or not. Thus, the increase of the mitotic cell was caused by the mitotic defect.
Therefore, the molecular mechanism for the function of the OdsHmel is playing a role in cell cycle regulation and focus on the M phase.
There is an example that overexpression the endogenous homeodomain contain gene in the Drosophila eye caused mitotic defect. Pax6 is the members of a set of the transcription factors network that coexpress in the retinal precursor cells before cell differentiation to control the eye development. This gene contains two DNA-binding domain including the homeodomain and a transactivation domain. The splice variants of Pax6, Pax6p46 was observed to bind chromosome in the
precentromeric region and resulted in abnormal eye development because of the mitosis defect (Zaccarini et al. 2007).
Recent data in the D. simulans cell culture system showed that the OdsH of D.
mauritiana and D. simulans co-localized to the pericentric satellite DNAs, and OdsH of D. mauritiana bind to an extra locus (Bayes 2008). Combine the mitotic defect in the third-instar larva showed in ectopic expression flies, it is possible that OdsH participates in cell division by direct interaction with the chromosome. This might be the hint for us to think about the hybrid male sterility from the cross between D.
simluans and D. mauritiana. The spermatogenesis process in Drosophila begins with primordial germ cells. Following one time of cell mitotic, primary spermatocyte was produced from the spermatogonia, which developed from the germ cells. Primary spermatocyte will then undergo two times of meiosis to generate four spermatids and will finally develop into four sperms (Fuller 1998; Gönczy and DiNardo 1996; Lin et al. 1994). In the spermatogenesis of the F1 hybrid males between D. simluans and D.
mauritiana, some onion cells show a disparity in size between the nucleus and the
mitochondrial (Perez at al. 1993). With the understanding of the molecular mechanism and these information about OdsH, I proposed that the hybrid males sterility is caused by the problems in the division procedures in spermatogenesis.
Furthermore, this mechanism for speciation would support the “centromere-drive”
model (Malik and Henikoff 2002).
Henikoff and Malik brought up an idea that centromeric DNA sequences might play a special role in Haldane’s rule (Malik and Henikoff 2001; Malik et al.
2002). They suggested that the rapid evolution of centromeric DNA reflects an arms race. During female meiosis, homologous chromosomes compete with each other to be the only one of four meiotic products that arrives in an egg in most species.
Although centromere drive is a well known mechanism in mammals (de Villena and Sapienza 2001; Henikoff et al. 2001), it brings the cost— heterochromatic differences between paired chromosomes in Drosophila males at meiosis I would cause non- disjunction and result in unequal sex ratio or sterility (Fig. 16).
Fig. 16.—The centromere-drive model. There are two steps in this model. First, expansion in the satellite sequence of the centromere lead to more microtubule binding sites, which can result in a transmission advantage in female meiosis. This would cause deleterious effects in males, such as non-disjunction between the X–Y chromosomes. Second, a suppressor allele in centromeric histone H3 (CenH3) that can recover the equality of meiotic will be selectively favored. By expanding CenH3 binding and increasing microtubule attachments on the Y centromere the deleterious effects of centromere-drive will be lightened. (Malik et al. 2002)
Satellite
Expansion CenH3 mutates
satellit e
expansion CenH3 mutates
According to studies here, the molecular mechanism of OdsHmel was suggested to play a role in cell division. Whether hybrid male sterility between D.
mauritiana and D. simulans is resulted from the cell division defect in the
spermatogenesis process is now interested in further analysis. In addition, it is also important to understand the function of the amino acids substitution and the Eh1 domain in OdsHmel.
Reference
Bateman JR, Lee AM, Wu CT. 2006. Site-specific transformation of Drosophila via
!C31 integrase-mediated cassette exchange. Genetics. 173:769-777.
Baker NE. 2001. Cell proliferation, survival, and death in the Drosophila eye. Semin Cell Dev Biol. 12:499-507. Review.
Bayes JJ, Findlay GD, Malik HS. 2008. Cytological characterization of a hybrid sterility gene in Drosophila.
Bischof J, Maeda RK, Hediger M, Karch F, Basler K. 2007. An optimized transgenesis system for Drosophila using germ-line-specific !C31 integrases. Proc Natl Acad Sci U S A. 104:3312-3317
Brand AH, Perrimon N. 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 118:401-415.
Brennecke J, Hipfner DR, Stark A, Russell RB, Cohen SM. 2003. bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell. 113:25-36.
Brideau NJ, Flores HA, Wang J, Maheshwari S, Wang X, Barbash DA. 2006. Two Dobzhansky-Muller genes interact to cause hybrid lethality in Drosophila.
Science. 314:1292-1295.
Buscarlet M, Stifani S. 2007. The 'Marx' of Groucho on development and disease.
Trends Cell Biol. 17:353-361.
Carrington JC, Ambros V. 2003. Role of microRNAs in plant and animal development. Science. 301:336-338. Review.
Carthew RW. 2007. Pattern formation in the Drosophila eye. Curr Opin Genet Dev.
17:309-313.
Chen CH, Huang H, Ward CM, Su JT, Schaeffer LV, Guo M, Hay BA. 2007. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science. 316:597-600.
Chen YL. 2005. Functional diversity of Odysseus (OdsH) and its role in
spermatogenesis. Master thesis, National Tsing Hua University, Taiwan.
Clarkson M, Saint R. 1999. A His2AvDGFP fusion gene complements a lethal His2AvD mutant allele and provides an in vivo marker for Drosophila chromosome behavior. DNA Cell Biol. 18:457-462.
Coyne JA. 1985. The genetic basis of Haldane’s rule. Nature. 314:736-738.