國立臺灣大學生物資源暨農學院森林環境暨資源學系 博士論文
Department of Forestry and Resource Conservation College of BioResources and Agriculture
National Taiwan University Doctoral Dissertation
臺灣中海拔青背山雀繁殖生物學研究
Breeding ecology of Green-backed Tits in mid-elevation of Taiwan
蕭明堂 Ming-Tang Shiao
指導教授:袁孝維 博士 Advisor: Hsiao-Wei Yuan, Ph.D.
中華民國 107 年 6 月
June 2018
誌謝
感謝神帶領我經歷 9 年的豐富,進到論文最後的致謝篇章。感謝王穎老師自 碩士班以來的引領,帶領我踏入學術的殿堂,從此山雀與巢箱的研究進入了我的 生活。每每和王老師碰面,老師總像長輩般關心我的生活、職場和家庭,然後分 享著他最近好慘的近況,以及從各種地方帶回來的奇特食物。感謝袁孝維老師在 我初進博士班時,啟蒙我研究的靈感與方向,關於人工林的鈣源對森林鳥類的影 響以及穩定同位素的應用等,成為我著手博士研究的起點。在研究的過程,袁老 師總是信任與支持,放手讓我做各樣的嘗試,並在最後畢業前的許多行政程序與 規定上,義不容辭地給予大力的幫忙與鼓勵。謝謝劉小如老師、徐堉峰老師及丁 宗蘇老師,撥空審查很不成熟的論文初稿,細心地提供許多文詞用語上和資料分 析上的寶貴建議,讓這本論文更加可讀並完整。
能以在職的身份進修博士班,感謝服務單位雪霸處的幾任處長,包括陳茂春 處長、林青處長、李秋芳處長與鍾銘山處長,他們總給我的支持和肯定,並讓我 在興趣專長的山雀研究上任意驅馳。感謝保育課于淑芬課長這些日子以來的支持 與包容,無論是每年自行研究計畫的提案或研究設備耗材的添購,常是二話不說 地支持。每年鳥類繁殖季的 3–8 月,我總是分身乏術於每週至少一次的野外調查 與辦公室工作,為此,謝謝于課長與這些日子保育課的潘振彰、傅國銘、陳家鴻、
李佩如等同仁,在工作上的分擔與包容。在保育課的工作也讓我常有機會認識不 同研究學門,這些經歷擴大了我研究構思的範疇與思維,並嘗試跨領域間的整合。
這個研究跨入了許多領域學門,完全不可能靠自己獨力完成。除了我自己熟 悉的山雀外,還包括土壤化學、陸生蝸牛、植群、鱗翅目幼蟲等面向,並還有一 些做過卻來不及呈現在論文內的工作項目,如雨水化學分析等。感謝中興大學森 林系的顏江河教授和研究室團隊,協助土壤化學的採樣與分析工作,特別是顏老 師聽我說不知怎樣判斷土壤層時,親自帶著研究室學生上山指導土樣的採集,並 協助完成大量樣本的後續分析工作。陸生蝸牛的採樣與鑑定上,有賴熊大維和楊 信得在起初的協助與引領,並且提供我實驗操作添加蝸牛殼的技術上建議,使得 實驗操作得以進行;謝謝廖婉均協助從土樣及枯枝落葉中,細心挑出蝸牛,這繁 瑣無趣的工作卻是研究的重要環節。各年繁殖鳥巢的例行監測、成鳥與雛鳥的繫 放、針孔錄影器材的架設與電瓶更換等龐大的工作,惟有賴各年的野外幫手協助,
才使得這 9 年的鳥巢繁殖物候的資料得以累積。早年的工作有賴師大野生動物實 驗室的黃書彥、潘玉潔、許詩涵、呂翊維、林玉珮等,以及臺大森林系實驗室的 黃婉如、謝惠冰、陳映嵐等,特別是書彥與詩涵,在大家逐步邁入各自職場生涯 時,仍多次擠出時間專程從臺南、從臺北跑來協助,點滴在心。此外,非常謝謝 蔣功國、姚牧君、陳柏因等幾任助理幫手,包容我許多任性和急躁的要求,在總 是風風雨雨的春夏鳥巢繁殖季,開著 march 小車奔走在危險的大鹿林道上。樣區 基礎植群資料的建立有賴林政道、陳柏因等協助,使得後續跨營養層級的研究工 作得以推行。謝謝中興大學昆蟲系的葉文斌教授與李蕙宜協助由雛鳥糞囊鑑定昆 蟲殘骸,建立雛鳥的食譜,這是非常困難又珍貴的分析工作。謝謝特生中心薛美 莉組長協助雨水收集裝置之架設及後續水樣的離子組成分析,讓我對樣區的背景 環境能有更完整的看見。謝謝歸國的蔡佳淳學姊百忙中仍讓我打擾,提供統計上 的諮詢,更讓我見識到國外研究不同的層級。
這 9 年的過程,有幸有 2–3 年能在雪霸的觀霧管理站服務,走出辦公室就是 中海拔的研究樣區,不知道是多少研究人員的憧憬。在觀霧服務的日子,感謝每 年管理站上的替代役熱血:江浩宇、陳冠豪、黃瀚嶢、蘇民弦、陳品翰、張乃中、
劉秉青、劉鎮源,協助研究工作上各樣的大小雜事、苦力等。謝謝你們的幫忙,
讓我能放手嘗試以沒人做過的龐大規模,進行臺灣中海拔樹冠昆蟲的生物量定量 與飼養工作。每次抖樹至少需要 3 個人才能進行,所以沒你們的幫忙,就沒有這 樣的資料累積,並且在每兩星期的抖樹採樣後,幼蟲的飼養須以每 3 天的頻率更 換食草,直到幼蟲化蛹,謝謝你們耐心幫忙當中繁雜的工作。另外,特別謝謝瀚 嶢,貢獻自己的興趣所長,協助鱗翅目成蟲陷阱採集後的大量鑑定工作。雖然部 分資料未能及時整合、呈現於論文內,然而我相信這仍是臺灣中海拔蛾類研究的 寶貴紀錄。謝謝中研院吳士緯博士協助鱗翅目成蟲與幼蟲鑑定上的諮詢,成為我 們在蛾類鑑定上最大的後援。另外,所抖落的昆蟲,扣除飼養的鱗翅目幼蟲外,
其餘大量的蜘蛛和其他無脊椎昆蟲等樣本的整理與初步分類工作,既耗時又傷眼 力,有賴廖佩柔耐心又細心的的協助,拼上了樹冠層昆蟲的最後一片拼圖。此外,
謝謝解說志工楊瓊珠與郭鳳嬌,在研究期間的許多鼓勵與協助,為冰冷的科學研 究帶入人的溫度。
最後,感謝我的妻子美真這些日子的陪伴。長年的研究工作已和我們彼此的 人生相交融,從交往、結婚、安禧的出生直到她快滿三歲的日子。謝謝妳這些日 子的包容、陪伴、忍耐與全力的支持,妳最常說的話就是:「你希望我怎麼幫忙?
怎麼配合?」謝謝妳的這句話,以及為了支持而背後經歷的許多辛酸,努力換班、
不計代價地讓班,來配合我上山巡巢、操作測量蛋或雛鳥、繫放親鳥,盡量分擔 繁殖季爆炸多的工作壓力。在探討鈣與蝸牛議題之那幾年,下班後妳常挑燈幫忙 從土樣和枯枝落葉中找出小小的蝸牛;在執行雨水化學研究的那 2 年,則是常常 刷著氣象局網站,看著氣象啥時下雨、啥時雨停,以便在雨停後立刻上山,回收 新鮮收集的水樣;在進行樹冠昆蟲與蛾類調查的那 2 年,安禧已經在肚子內了,
妳仍然三不五時陪我上山抖樹,幫忙更換幼蟲食草,秤著常常是幾百隻的蛾類成 蟲的生物量,然後一隻隻裝袋;在安禧甫出生的那年,則努力將上下山的路程安 插入安禧的作息、喝奶的時間,並在當中的空檔協助雛鳥的繫放。直到安禧 2–3 歲的現在,整天要爸爸、爸爸的時候,妳仍努力地 hold 住,讓我能撥空專心寫 paper、
寫論文。謝謝你包容許可我任性地做這樣的研究,並且完全的支持;另外也謝謝 國恩多次上山的協助,彼此雙方家人時刻的關心和代禱。
感謝神帶領我經歷這 9 年的豐富。剛踏入博士班時,或因著碩士時期的研究 剛刊登在 AUK 期刊上,我帶著些許的自信挑戰新的研究。然而,由碩士研究的動 物行為學、個體層級,跳到博士研究的族群或群落層級,是不同的領域;並且儘 管同樣是青背山雀,不同地區的族群對干擾壓力有著不同的反應;在職與全職不 同的身份更是不可同日而語。這許多的不同,讓我在博士班的初期如同經歷聖經 哈巴谷書那沒羊、沒牛、樹不結果、田無土產,什麼都沒有的光景。但就在那看 似失控不如預期下,卻讓我體認神創造的豐富與寬廣,遠超過我原本的自以為。
在祂的恩典下,保守每次調查合適的天氣、人力需求的供應,到最後能有地利之 便,完成一些沒人做的嘗試挑戰,所獲得資料的豐富,更超越博士論文的門檻。
感謝神揭露祂奇妙創造的一丁點,讓我經歷也享受於其中。在這不算短的年日,
感謝神讓我時刻處在科學研究、邏輯思考的雲端,與經營管理應用層面的現實之 間,雖然煎熬又精神分裂,卻也不斷地促使我思考科學的價值、研究的目的、生 命努力的目標與應抱持的態度。感謝神在 9 年來的恩典與帶領,願榮耀都歸於祂。
中文摘要
於 2009–2017 年間在觀霧中海拔針闊葉混淆林與柳杉針葉林,吾人監測使用 巢箱繁殖的青背山雀(Parus monticolus)之繁殖表現。研究工作主要由三部分所組成,
第一部分為檢驗環境中富鈣物質的多寡(亦即陸生蝸牛密度)有無限制雌鳥的生殖 產出。結果顯示,儘管兩種棲地的土壤均是高度酸性,研究族群的雌鳥產下的蛋,
蛋殼並無缺陷的症狀,並且額外提供鈣源未增加雌鳥的生殖產出。顯示現階段鈣 並非限制青背山雀雌鳥產蛋的限制營養。第二部分為調查在兩種棲地上,不同樹 種所提供的不同幼蟲如何影響青背山雀雛鳥的生長。吾人藉由抖落法定量各主要 樹種上的鱗翅目毛蟲與膜翅目幼蟲組成,以及幼蟲生物量於季節上的變化。結果 顯示,闊葉棲地具多樣的樹種,構成較寬的幼蟲生物量高峰。幼蟲生物量於初春 時急遽上升而後逐漸下降。相反的,針葉棲地的幼蟲生物量為隨著季節緩慢增加,
並且其物種組成中有高比例的有毛苔蛾。兩種棲地具有不同的幼蟲生物量之季節 性動態,並影響青背山雀繁殖的第一窩與第二窩雛鳥體重。第三部分係以多年的 監測資料來檢視氣候因子與青背山雀繁殖起始之關聯,並檢測影響鳥巢存活率的 重要因子。研究結果顯示,不同年間,青背山雀產蛋繁殖的早晚與三月均溫呈顯 著正相關,與 2–3 月春雨雨量無關。在較溫暖的初春,雌鳥傾向較早開始產蛋繁 殖。繁殖期間的大雨事件則對鳥巢的存活有負面影響。最後,吾人整合青背山雀 的生活史參數,並估算族群成長係數。藉由情境模擬顯示,亞成鳥存活率、成鳥 存活率與每巢離巢雛鳥數為影響年間族群波動的最重要因子。
關鍵詞:鈣限制、青背山雀、鳥巢存活、物候、生殖產出
Abstract
The use of nest boxes by Green-backed tit (Parus monticolus) was monitored in a mosaic landscape of mixed-oak forest and Cryptomeria plantation in the mid-altitude area of Guanwu between 2009 and 2017. Field work falls into three sections that
together provide insight and knowledge of breeding behavior of hole-nestling passerines.
The first section was to test whether the availability of calcium-rich materials limits reproduction output of birds. Our results found that despite the high acidity of the soil in two habitats, none of the birds in the study population produced eggs with defective shells. The provision of supplementary calcium did not appear to influence reproductive output of birds. Calcium is not currently a limiting micronutrient for egg production by Green-backed Tits. The second section examined how differential larval phenology supported by different tree species in the two habitats affects nestling mass. The larval abundance (Lepidoptera and Hymenoptera sawfly larvae) of dominant tree species was measured via the tree-shaking method. The results showed that the diversity of trees in the broadleaf habitat supported a broader food peak overall that spiked in early spring and gradually declined. In contrast, the coniferous habitat had a lower larval biomass that increased slightly over the season and included a high percentage of hairy lithosiines. Habitat-specific seasonality in larval abundance significantly affected nestling mass in early- and late-season broods. The third section determined what weather cues montane birds use to fine-tune breeding behavior and what factors affects their breeding success from multi-year monitoring data. Our data indicated that March temperatures, but not February–March rainfall affected the laying date for
Green-backed Tits. Females shifted their laying dates earlier in response to earlier warm spring weather. Heavy rainfall during the breeding period negatively affects their nest survival. Finally, several life-history traits of the Green-backed tits were estimated and discussed.
Keywords: calcium limitation, Green-backed Tit, nest survival, phenology, reproductive output
目 錄
口試委員會審定書………i
誌謝………...ii
中文摘要………...v
英文摘要……….... .vi
圖目錄………...…...ix
表目錄………...x
第一章 緒論……….. 1
1.1 前言……… 1
1.2 研究樣區與巢箱設置……… 1
1.3 研究地區氣候……… 2
1.4 繁殖鳥巢例行監測……… 3
1.5 參考文獻……… 3
第二章 產蛋期雌鳥生殖產出………...8
2.1 前言……….8
2.1 研究方法……….9
2.3 結果………...12
2.4 討論………...13
2.5 參考文獻………...15
第三章 育雛期雛鳥生長……….25
3.1 前言………...25
3.1 研究方法………...27
3.3 結果………...29
3.4 討論………...31
3.5 參考文獻………...34
第四章 繁殖季的起始與鳥巢存活……….45
4.1 前言………...45
4.1 研究方法………...46
4.3 結果………...50
4.4 討論………...52
4.5 參考文獻………...55
第五章 族群成長係數與總結……….76
5.1 前言………...76
5.1 研究方法………...77
5.3 結果………...78
5.4 討論………...79
5.5 參考文獻………...80
圖 目 錄
圖 1.1、2009–2017 年觀霧地區鳥巢箱設置樣區於空間之配置。………...6 圖 1.2、2009–2017 年中央氣象局 C0D550 自動測站記錄之各月均溫± SD 與平均降
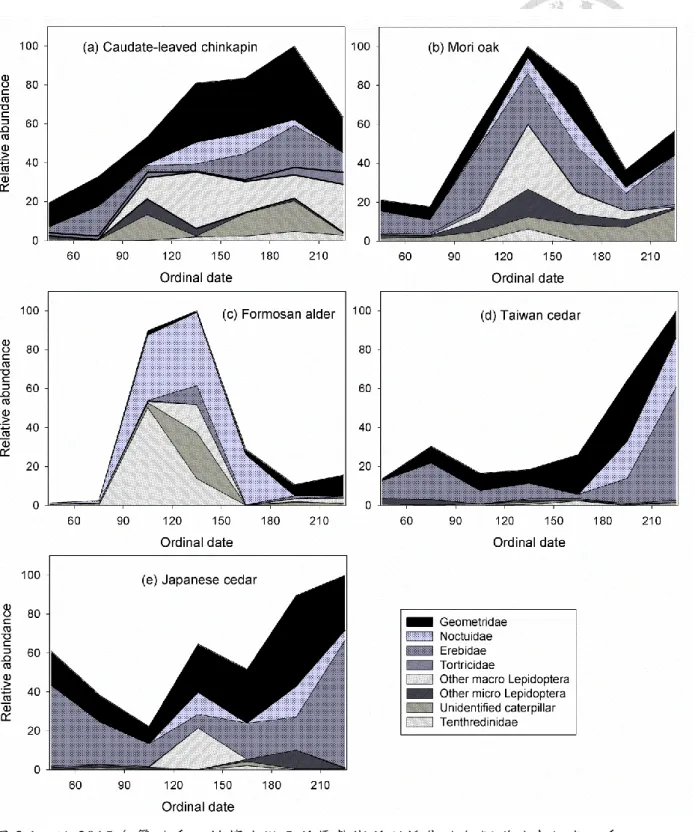
水量+ SD。……….………..…………...………7 圖 3.1、於 2015 年繁殖季以抖落法從 5 種優勢樹種所採集的各類群幼蟲組成之季
節性變化。……….………..…………...………40 圖 3.2、於 2015 年繁殖季以抖落法兩週乙次之採樣頻率,由 5 種優勢樹種所採集
之幼蟲生物量於季節上的分布。………..………..………41 圖 3.3、預測的闊葉棲地和針葉棲地之幼蟲生物量分布。………42 圖 3.4、於闊葉棲地與針葉棲地繁殖之青背山雀第一窩和第二窩繁殖巢,(a)於雛鳥
需求高峰時期,可獲得之總幼蟲生物量(毫克 ± SE)和(b)平均雛鳥體重(克 ± SE)。..………..………..………..………..………..………43 圖 3.5、於 2015 年闊葉棲地與針葉棲地繁殖之青背山雀,於雛鳥需求高峰時期(第
10–21 天日齡),可獲得之總幼蟲生物量和雛鳥體重殘差之關係。…………44 圖 4.1、2009–2017 年觀霧地區利用巢箱之青背山雀卵數及雛數分布。……….68–69 圖 4.2、2009–2017 年觀霧地區利用巢箱之棕面鶯卵數及雛數分布。………...70–71 圖 4.3、於 2010–2017 年觀霧地區青背山雀與棕面鶯第一窩繁殖巢平均產蛋日之比
較。..…..………..………..72 圖 4.4、於 2010–2017 年觀霧地區青背山雀(實心圓)與棕面鶯(空心圓)第一窩繁殖巢
之平均產蛋日與三月均溫之關係。…..………..………..73 圖 4.5、於 2009–2017 年觀霧地區青背山雀鳥巢之週期存活率與繁殖嘗試、有無大
雨事件之關係。…..………..………..…………..………..74 圖 4.6、於 2009–2017 年觀霧地區棕面鶯逐日存活率與鳥巢日齡之關係。…..…..75 圖 5.1、青背山雀生活史參數與其和影響因子關係之概念圖。…..………..………...86 圖 5.2、青背山雀繁殖族群密度與各項生活史參數之關聯。…..………..………...87 圖 5.3、2010–2017 年各年度青背山雀之繁殖密度(實心圓)與預測繁殖密度(空心圓)
之關係…..………..………..……..………..………..……..………..…...88
表 目 錄
表 1.1、2009–2017 年觀霧地區針闊葉混淆林與柳杉針葉林之有效巢箱數。...5
表 2.1、觀霧地區針闊葉混淆林與柳杉針葉林枯枝落葉及土壤參數之比較。...21
表 2.2、模式分析 2011–2014 年中海拔觀霧地區青背山雀之(a)蛋形質特徵、(b)窩 卵數和(c)孵化成功率。...22
表 2.3、各研究調查森林陸生蝸牛密度之比較。...24
表 3.1、以抖落法所採集的 5 種優勢樹種上,幼蟲生物相組成之百分比。...39
表 4.1、2009–2017 年觀霧地區青背山雀與棕面鶯之鳥巢存活率模式分析,各候選 模式所涵蓋的變數和假說。...63
表 4.2、2009–2017 年觀霧地區青背山雀鳥巢存活分析之模式篩選結果。...64
表 4.3、2009–2017 年觀霧地區青背山雀鳥巢存活之最佳支持模式之各參數估計值 與 95%信賴區間。...65
表 4.4、2009–2017 年觀霧地區棕面鶯鳥巢存活分析之模式篩選結果。...66
表 4.5、2009–2017 年觀霧地區棕面鶯鳥巢存活之最佳支持模式之各參數估計值與 95%信賴區間。...67
表 5.1、2009–2017 年青背山雀成鳥年存活率之模式選擇結果。...83
表 5.2、2009–2017 年青背山雀生活史參數之估計。...84
表 5.3、模擬不同繁殖參數之情境下,對青背山雀族群成長係數之影響。...85
第一章 緒論 1.1 前言
鳥類繁殖過程的各階段受不同因子所限制,並形塑出其繁殖表現,各階段的 繁殖表現則影響整體生活史的參數與族群成長。本研究以青背山雀(Parus
monticolus)為標的物種,由產蛋期雌鳥的生殖產出、育雛期雛鳥生長、繁殖季的起 始與鳥巢存活,以及生活史族群參數估算等四個面向,逐步檢視影響青背山雀繁 殖過程的重要因子。
青背山雀為小型的森林食蟲鳥類(體重約 12 公克),是溫帶模式物種─大山雀 (Parus major)的相近種,為臺灣特有亞種;它普遍分布於臺灣中低海拔(1,000–2,500 公尺)的闊葉林及造林地,每個繁殖季繁殖 1–2 巢(兼行兩巢,facultative double breeders),且能利用巢箱繁殖(Severinghaus et al. 2012),適合做為高山森林鳥類長 期監測及試驗操作的標的。吾人於臺灣北部中海拔針闊葉混淆林與柳杉針葉林鑲 嵌環境設置人工巢箱,針闊葉混淆林為臺灣高山地區具代表性的原始植群類型,
而柳杉針葉林則是臺灣重要的造林林型,占全臺造林地 11%(Qiu et al. 2015)。本研 究分成四個章節,各章節的研究目的與資料期程分述如下:
一、 產蛋期雌鳥的生殖產出(2011–2014 年):調查二林相的蝸牛密度,探討產蛋 期間,環境中富鈣物質的多寡對雌鳥生殖產出之影響;
二、 育雛期雛鳥生長(2015 年):調查二種林相的重要樹種上,幼蟲生物量(鱗翅 目毛蟲與膜翅目蠕蟲)的季節性變化,並檢視幼蟲生物量與育雛期雛鳥體重 之關聯;另以多年資料(2010–2017 年)比較二林相之繁殖特徵差異。
三、 繁殖季的起始與鳥巢存活(2009–2017 年):綜觀所有監測資料,檢視氣候因 子對於青背山雀繁殖起始時間之關聯,以及對整體鳥巢存活率之影響;另 外,一併檢視並比較青背山雀與共域物種─棕面鶯(Abroscopus albogularis) 的差異。
四、 族群成長參數估算(2009–2017 年):綜觀所有監測資料,估算青背山雀的各 項繁殖參數與族群成長參數,並檢視影響年間族群波動的重要因子。
1.2 研究樣區與巢箱設置
研究地點位於亞熱帶臺灣北部,中海拔霧林帶的觀霧地區(24° 30′ N,121° 06′
E,海拔高為 1,900–2,200 公尺),位於雪霸國家公園觀霧遊憩區及鄰近聯外道路的 兩側林地。林地坡度為 10–40 度,為針闊葉混淆林(mixed-oak forest)與柳杉針葉林 (Cryptomeria plantation)的鑲嵌地景。吾人於 2012 年針對針闊葉混淆林和柳杉針葉 林分別選取 21 個和 22 個 10 x 10 m 樣方進行植群調查,並計算各樹種的相對優勢 度(relative dominance,RD),以該樹種胸高斷面積之總和除以所有樹種胸高斷面積 之總和。針闊葉混淆林的森林內部優勢樹種為森氏櫟(Cyclobalanopsis morii;RD:
19%)、卡氏櫧(Castanopsis cuspidate;RD:17%),森林邊緣的主要樹種則為火燒 或崩坍後的先驅樹種,包括臺灣赤楊(Alnus formosana;RD:11%)、臺灣二葉松(Pinus taiwanensis;RD:26%)等。柳杉針葉林為天然林皆伐後,重新栽植 30-40 年。主 要的造林樹種為日本柳杉(Cryptomeria japonica;RD:54%)、臺灣杉(Taiwania cryptomerioides;RD: 25%)和杉木(Cunninghamia lanceolata;RD:13%)。
吾人選擇數塊針闊葉混淆林與針葉林林地設置巢箱(圖 1.1),每塊林地約 1–2 公頃。研究前期(2009–2012 年)設置的巢箱主要採網格狀配置,然因鳥類繁殖領域 的排他性,以致於某區塊一旦有鳥進駐使用後,周遭巢箱常不再被其他個體所利 用。為擴大鳥巢樣本數,後期(2014–2015 年)增設的巢箱改採用帶狀配置。針葉林 巢箱樣區共包括 2009 年設置 3 處網格狀樣區(共 90 個巢箱;1.1A 區、C 區)、2014 年增設的 1 處帶狀配置樣區(共 6 個巢箱;圖 1.1F 區),以及 2015 年增設的 2 處帶 狀配置樣區(共 12 個巢箱;圖 1.1E 區、R 區);闊葉林巢箱樣區共包括 2010 年設 置 1 處帶狀樣區(共 15 個巢箱;圖 1.1B 區)、2011 年、2012 年增設的 2 處網格配 置樣區(共 50 個巢箱;圖 1.1D 區),以及 2014 年增設 1 處帶狀配置樣區(共 9 個巢 箱;圖 1.1F 區)。各年度可利用巢箱數與設置樣區面積如表 1。
巢箱與巢箱間的間距約 15–30 公尺。所有的巢箱均釘在 3 公尺高度的樹上,
若是可能,洞口盡可能朝南或西南。巢箱規格設計供小型雀型目使用,洞口直徑 為 3 公分,底面積為 10.5 × 12.5 平方公分,箱底具 4 個孔洞可供排水,巢箱深度 由從洞口至底板為 11.3 公分。
1.3 研究地區氣候
研究地點屬於亞熱帶的潮濕氣候,鄰近的氣象測站位於巢箱樣區 F 區(圖 1.1),
為中央氣象局所設置的 C0D550 雪霸自動測站(24°31′N37′′N,121°06′58′′E),氣象
站海拔高 1,956 公尺,設立時間為 2007 年 10 月起。測站資料顯示本區的溫度與降 水具明顯的年週期變化,自 1 月起至 7 月期間為持續增溫(圖 1.2)。於 2009–2017 年研究期間,4–8 月的平均溫度± SD 為 16.9 ± 2.3 °C,範圍介於 5.7–23.3 °C。每年 的 10 月至隔年 1 月為枯水期,2–3 月的降雨,通稱春雨(spring rainfall;Chen et al.
2003, Chang et al. 2013 和本研究)和 4–8 月的繁殖季降雨具明顯的年間變異。於 2009–2017 年研究期間,2–3 月春雨之平均降水量± SD 為 337.6 ± 157.1 毫米,範圍 介於 198 毫米(2011 年)和 656 毫米(2016 年)之間;4–8 月繁殖季降雨之平均降水量
± SD 為 1,565 ± 578 毫米,範圍介於 1,007 毫米(2016 年)和 2,806 毫米(2013 年)之 間。
1.4 繁殖鳥巢例行監測
於 2009 至 2017 年間,在 3 月中旬至 8 月期間每周至少 1 次巡巢,紀錄巢箱 的內容物(蛋數和雛鳥數),所有的鳥巢在產蛋期或孵蛋前期就被發現。吾人假設雌 鳥每天產一個蛋,由巡巢時紀錄的蛋數來回推產第一枚蛋日(laying date),並以四 月日(April date)來表示(1 = 4 月 1 日)。新生雛鳥於孵化後第一次巡巢發現時秤重或 在那之後盡快進行,以電子口袋秤(JYA–150,Jin Yuan,臺灣)秤重量測至 0.1 公克,
並由平均雛鳥生長曲線(Huang 1996, Shiao 2006)來回推其日齡。鳥巢於育雛中至後 期,選擇性架設針孔錄影,紀錄巢內活動與離巢日期(方法詳見第三章),雛鳥於離 巢前將上鋁環和色環繫放;繁殖成鳥於育雛期間透過雙筒萬遠鏡於野外觀察或鳥 巢內的針孔錄影進行個體辨識,無腳環的個體將於育雛中至後期進行捕捉繫放,
上色環與鋁環。其餘各年度額外的蛋或雛鳥的測量、操作試驗及環境調查於各章 節分述之。
1.5 參考文獻
Chiou, C.-R., T.-Y. Chen, H.-Y. Liu, J.-C. Wang, C.-L. Yeh, and C.-F. Hsieh (2009).
Atlas of natural vegetation in Taiwan. Forestry Bureau, Council of Agriculture.
Taipei, Taiwan (In Chinese).
Huang, C.-L. (1996). Breeding biology of birds using nest boxes in Taroko National Park. M.S. thesis, National Taiwan Normal University, Taiwan (In Chinese).
Qiu, L.-W., Q.-X. Huang, C.-C. Wu, and H.-T. Hsieh (2015) Summary of the fourth national forest resource inventory report. Taiwan Forestry Journal 41:3–13 (In Chinese).
Severinghaus, L. L., T.-S. Ding, W.-H. Fang, W.-H. Lin, M.-C. Tsai, and C.-W. Yen (2012). The avifauna of Taiwan. 2nd edition. Forest Bureau, Council of Agriculture, Taipei, Taiwan.
Shiao, M.-T. (2006). Study on the food allocation of the parental Green-backed Tit (Parus monticolus) during fledging period at Guanyuan. M.S. thesis, National Taiwan Normal University, Taiwan (In Chinese with English abstract).
表 1.1、2009–2017 年觀霧地區針闊葉混淆林與柳杉針葉林之有效巢箱數
針闊葉混淆林 柳杉針葉林
年度 有效巢箱數 面積(公頃) 有效巢箱數 面積(公頃)
2009 - - 90 3
2010 15 1 90 3
2011 45 2 90 3
2012 63 3 90 3
2013 64 3 90 3
2014 74 5 96 4
2015 74 5 107 9
2016 74 5 107 9
2017 71 5 108 9
圖 1.1、2009–2017 年觀霧地區鳥巢箱設置樣區於空間之配置。底圖為 Chiou(2009) 臺灣現生天然植群圖集,淺藍色為人工林、淺橘色為山地針闊葉混淆林、淺綠色 為山地常綠闊葉林,粉紅色為果園耕地。
圖 1.2、2009–2017 年中央氣象局 C0D550 自動測站記錄之各月均溫± SD(實心圓) 與平均降水量+ SD(長條圖)。
第二章 產蛋期雌鳥生殖產出 2.1 前言
鈣是鳥類生殖的重要營養,產蛋的雌鳥需要大量的鈣來形成蛋殼,鳥蛋的乾 重有 10–15%為鈣所組成(Gilbert 1971, Graveland and van Gijzen 1994)。許多鳥類的 跗趾髓骨(medullary bone)能進行短期的儲鈣(Simkiss 1967, Gilbert 1983, Reynolds 2003),,然而雌鳥仍有賴產蛋期間每日增加對富鈣物質的攝取與吸收,以滿足它們 產蛋所需的鈣(Reynolds and Perrins 2010)。蝸牛殼和許多富鈣的無脊椎動物,如鼠 婦(Isopoda spp.)和馬陸(Diplopoda spp.),是許多食蟲雀型目鳥類的主要鈣源
(Graveland and van Gijzen 1994, Graveland 1996, Ramsay and Houston 1999,
Reynolds and Perrins 2010)。因此,鳥類繁殖產蛋的棲地須提供足夠的富鈣食物。
蝸牛及其他富鈣的無脊椎動物的密度受環境土壤及枯枝落葉的化學特性,如 總鈣、土壤 pH、交換性鈣離子、陽離子交換能力(cation exchange capacity, CEC) 所影響(Gärdenfors et al. 1995, Graveland and van der Wal 1996, Johannsenn and Solhøy 2001, Martin and Sommer 2004)。局部地區的鈣含量是土壤特性、植群與外 在因子間交互作用下的產物。人為酸性汙染物的沉降(酸沉降)為溫帶地區土壤酸化 的主要原因(De Vries and Breeuwsma 1986),導致表層土壤鈣離子的流失,並進一 步減少蝸牛、馬陸等富鈣無脊椎動物的密度(Graveland and van der Wal 1996, Graveland 1998, Rusek and Marshall 2000)。在高度酸化的地區,例如挪威的
Buunderkamp 森林,大山雀和其他山雀無法獲得足夠的鈣來形成蛋殼,而產下具有 缺陷、蛋殼過薄的蛋,並造成高比例的棄巢或空巢(empty nests; Graveland et al. 1994, Graveland 1998)。鈣的可獲得性除了限制鳥類的生殖產出,也可能改變它們活動的 時間分配(time budgets)。搜尋並攝取充足的鈣比滿足能量需求更為耗時(Turner 1982),因而鈣缺乏的雌鳥可能會耗費較多時間在搜尋鈣,並減少休息或其他的活 動(Graveland and Berends 1997)。
土地利用也影響地區性的鈣含量。相較於天然森林,人工針葉林通常被視為 缺鈣的棲地,因為一些針葉樹種,如歐洲赤松(Pinus sylvestris)、歐洲黑松(Pinus nigra)、美國黑松(Pinus contorta)和花旗松(Pseudotsuga menziesii)等的枯枝落葉所含 的交換性塩基陽離子少於闊葉樹,因而造成土壤酸化並且分解慢(Zinke 1962, Sawyer 1993, Reich et al. 2005)。這些人工林的土壤動物相之多樣性及密度相對較低
(Cameron 1978, Mänd et al. 2000a, Tattersfield 2001, Bonham et al. 2002),在這類棲 地繁殖的鳥類傾向產下較小的窩卵數和較輕、較小的蛋(Tilgar et al. 1999, 2002)。
不過,樹種的枯枝落葉之化學特性為內生性(intrinsic)、具種間差異的特徵(Reich et al. 2005)。部分柏科(Curpressaceae),如日本柳杉的枯枝落葉含有高達闊葉樹種三 倍以上的鈣(Xue and Luo 2002; Reich et al. 2005; Ohta et al. 2014a),因而在日本柳杉 生長的棲地具有較高的交換性鈣離子和豐富的富鈣甲殼類(crustaceans; Ohta et al.
2014b)。不過,至今我們對於鳥類在富鈣人工林環境的繁殖表現所知有限。
臺灣為位在東亞的多山島嶼,鄰近中國東南沿岸僅 220 公里。臺北北部及中 部的高山森林受到西部沿岸平原的工業活動和來自中國的長程傳輸所帶來的酸沉 降輸入(SO42-_S:10–48 kg ha-1yr-1、NO3-_N 和 NH4+_N:9–73 kg ha-1yr-1;Lin et al. 2000, Hsiao et al. 2007, Liang et al. 2009)。酸沉降造成的土壤酸化效應(De Vries and Breeuwsma 1986, Graveland 1998),伴隨山區的高降雨量(年降雨量 2,000–3,000 mm) 加速土壤的侵蝕,使得臺灣北部山區成為酸沉降臨界負荷敏感的區域(Chang et al.
2009),並可能進一步影響鳥類的生殖產出。
本研究目的為調查在北臺灣高山地區針闊葉混淆林與柳杉針葉林鑲嵌環境下,
青背山雀的生殖產出是否受鈣的可獲性所影響。針闊葉混淆林為高山地區具代表 性的原始植群類型,而柳杉針葉林則是臺灣重要的造林林型,其占全臺造林地 11%(Qiu et al. 2015)。青背山雀與其它山雀相似,以蝸牛殼為主要的鈣源(Graveland 1996, Reynolds and Perrins 2010)。是以,吾人調查二種棲地類型中的土壤養分及小 型蝸牛的密度。同時,在四年的研究期間(2011–2014 年),針對在二種棲地繁殖的 部分青背山雀鳥巢,於產蛋期間供給補充鈣源。倘若任一棲地的繁殖雌鳥,其產 蛋受鈣的可獲性所限制,提供額外鈣源將有助於生殖產出的增加。研究將比較二 種棲地間,以及操作組與控制組間,窩卵數、蛋的型質及孵卵率等項目,並預期(1) 在枯枝落葉含鈣量、土壤 pH、交換性鈣離子以及蝸牛密度等相關參數上,柳杉針 葉林應高於針闊葉混淆林;(2)在針闊葉混淆林的繁殖雌鳥受鈣缺乏的影響可能高 於在柳杉針葉林的繁殖個體。
2.2 研究方法
2.2.1 添加鈣試驗與蛋形質測量
2011 年至 2014 年期間,於繁殖季每星期巡巢中,一旦發現鳥巢進入內襯築巢 期(nest lining;在苔蘚結構上出現羽毛或絨毛),便開始進行添加鈣的試驗,直到雌 鳥產完蛋。為了減少產蛋時間對雌鳥繁殖表現的可能影響,所有鳥巢在內襯築巢 期間都依據其築巢時間的先後進行配對,兩組中逢機一個鳥巢作為添加鈣的操作 組,另一個做為控制組。配對的過程分別在闊葉棲地與針葉棲地兩個林相內各自 進行。控制組與操作組在各年間的樣本數配置分別為:2011 年 10 巢和 8 巢、2012 年 10 巢和 8 巢、2013 年 6 巢和 6 巢、2014 年 5 巢和 3 巢。
添加鈣的操作組鳥巢,於每個巢箱內側靠近洞口處設置一個小型塑膠盒,裡 面放置 200 毫克的蝸牛殼碎片以及 200 毫克的家雞(Gallus domesticus)蛋殼碎片。
在大山雀的研究發現,雄鳥極少攝取補充鈣(Graveland and Berends 1997)。又,在 築巢和產蛋期間,親鳥會持續地防衛牠們的領域,驅趕任何接近牠們鳥巢的個體,
因此假設所設置的添加鈣源僅能被該巢的雌鳥所取食,很難被目標以外的其他個 體所利用。塑膠盒及所添加的鈣源於每星期巡巢時更換新的一份,俾使得添加的 鈣源在操作期間能被親鳥任意取食。並回收前次未取食的殘餘,若有任何掉落在 巢材上的鈣源碎片,視為未被雌鳥所取食,並一併回收。所回收的塑膠盒及殘餘 鈣源將置於 60 度的烘箱(STD–30A,Cherng Huei Co., Ltd.,臺灣)烘乾,然後秤重,
以計算雌鳥實際攝取添加鈣源的量。
青背山雀通常一季繁殖 1–2 巢。在雌鳥產完蛋後,紀錄每巢的窩卵數,以電 子口袋秤(JYA–150,Jin Yuan,臺灣)測量所有蛋的蛋重,量測至 0.1 公克;以電子 游標卡尺(CD–6” CSX,Mitutoyo,日本)測量所有蛋的長(L)和寬(B),量測至 0.01 毫米,用以計算蛋形指數(egg shape,B ∕ L;Hoyt 1979)、蛋體積(egg volume,0.51
× L × B2;Hoyt 1979)及整窩蛋體積(clutch volume)。整窩蛋體積定義為整窩蛋的體 積之總和(Mänd et al. 2000b),並包括無卵黃的侏儒蛋(dwarf eggs)。另外,計算孵 化成功率,以雛鳥數除以蛋數之比例來表示。
2.2.2 蝸牛密度與土壤化學
在 2013 年,於設置巢箱的針闊葉混淆林與柳杉針葉林林地內(圖 1.1,A–D 區),
每塊林地內選擇 3–9 個採樣點,採樣點的數量係依據林地大小以及該地區繁殖鳥 巢的資料數,總計於針闊葉混淆林和柳杉針葉林內分別共選擇 20 和 16 個採樣點。
於每個採樣點上,選取 2 塊相鄰、25 × 25 平方公分的方形採樣樣方(sampling plot),
並各自從中採樣枯枝落葉及表層土壤樣本,一份樣本用於檢視陸生蝸牛,另一份 用於土壤化學分析。枯枝落葉的採樣係採取樣方地表上的所有枯枝落葉,而土壤 的採樣則是採取樣方上的土壤腐質層。不同採樣點的土壤腐質層深度不同,土樣 採樣的平均深度(±SD)為 2 ± 1.5 公分(n = 36)。陸生蝸牛密度的估算係以手挑選肉 眼尋找枯枝落葉及土壤樣本中的蝸牛殼並計數(隻數/ 25 × 25 平方公分)。土壤化學 分析則是委由國立中興大學森林系森林土壤及林木菌根研究室進行。分析項目包 括枯枝落葉的含鈣量、土壤層的 pH-H2O、pH-KCl、交換性鈣離子及陽離子交換能 力等。
2.2.3 統計分析
所有的統計分析均是在 R version 3.4.1 平台上執行(R Development Core Team 2017)。吾人先以 Shapiro-Wilk test 檢測土壤化學的各項參數有無符合常態分布。符 合常態分佈的資料,如 pH-KCl、陽離子交換能力、枯枝落葉的含鈣量等項目,以 one-way ANOVA 進行組間比較;不符合常態分佈的資料,如 pH-H2O 和交換性鈣 離子等項目,則以 Mann-Whitney U test 進行組間比較。陸生蝸牛的分布在棲地內 變化極大,導致資料呈現過度分散(overdispersion),且過多的資料筆數為零,故吾 人以 zero-inflated negative binomial (ZINB) regression (function zeroinfl in package pscl)來比較二棲地間蝸牛密度的差異。ZINB 迴歸適用於過度分散且具有大量計數 為零的資料,在 ZINB 迴歸中,將”棲地”以自變數納入考量,並以 likelihood ratio test 比較在涵蓋”棲地”因子的完整模式(full model)和不涵蓋”棲地”因子的簡化模式 (reduced model)兩者間的差異,來判斷”棲地”因子是否為模式中的顯著重要因子。
另外,使用線性模型(linear model,LM;function lm)來檢視蝸牛密度與土壤化學參 數間的可能關係。
鳥巢的繁殖表現資料為連續 4 年的資料(2011–2014),當中少部分個體在不同 年間被重複測量。為了解釋觀察值之間並非完全獨立的問題,吾人選擇用混合模 型的方法(mixed modeling approach)。在模式中納入”成鳥個體 ID”及”年”為隨機效 應(random effects),以便估計其他的固定效應(fixed effects)。為比較”添加蝸牛殼被 攝食比例”以及”添加蛋殼被攝食比例”在不同棲地間有無差異,吾人進行兩個具二
項誤差結構(binomial error structure)的廣義線性混合模型(generalized linear
mixed-effects models,GLMMs;function glmer in package lme4)。其中,所添加蝸 牛殼 / 蛋殼被攝食比例係以雌鳥攝取量除以總供給量來表示,並設為模式的應變 數,棲地類型則為模式的自變數。在比較棲地間及操作組別間的繁殖參數上,吾 人依據資料的結構來建立模式。對於連續性數值資料,如蛋重、蛋形指數、蛋體 積及整窩蛋體積等四個參數,吾人分別建立四個具高斯誤差結構(Gaussian error structure)的線性混合模型(linear mixed models,LMMs;function lmer in package lme4、
package lmerTest);對於百分比資料(孵化成功率),吾人建立具高斯誤差結構的廣 義線性混合模型(GLMM);對於屬計數資料(count data)的窩卵數,因其窩卵數的分 布範圍小,介於 4–7 之間,且樣本數有限,以至於在 GLMM 的建模過程,二個隨 機效應(成鳥個體 ID 和年)之變異數均為零。為此,吾人將這兩個隨機效應刪除 (Bolker et al. 2009),並建立具卜瓦松誤差結構(Poisson error structure)的廣義線性模 型(generalized linear model,GLM;function glm in package lme4)。上述模式的建立 係透過逐步向後法(stepwise backward procedure),棲地類型、操作組別及它們的交 互作用起初均納入模式中,並逐步刪除不顯著的交互作用或因子。除非特別說明,
否則所有數值均以平均值 ± SE 來表示。統計的閥值設為 0.05。
2.3 結果
2.3.1 土壤養分和陸生蝸牛
柳杉針葉林枯枝落葉的含鈣量略高於針闊葉混淆林,但統計上未達顯著差異 (表 2.1)。二林相的土壤均呈現酸性,針闊葉混淆林的土壤 pH-HCl 最低為 2.61,而 柳杉針葉林的土壤 pH-HCl 最低為 2.78。相較於針闊葉混淆林土壤,柳杉針葉林土 壤有顯著較高的 pH、顯著較高的交換性鈣離子以及顯著較低的陽離子交換能力(表 2.1)。
二林相的蝸牛分布均為不均勻分布,並有多數為零的紀錄,造成資料分布的 正偏(positive skew)。然而整體而言,柳杉針葉林的蝸牛密度(平均值 ± SD:4.3 ± 7.8 隻蝸牛 / 25 × 25 平方公分,中位數:2.5 隻蝸牛 / 25 × 25 平方公分,範圍:0–30 隻蝸牛 / 25 × 25 平方公分)高於針闊葉混淆林(平均值 ± SD:2.8 ± 2.7 隻蝸牛 / 25
× 25 平方公分,中位數:1.0 隻蝸牛 / 25 × 25 平方公分,範圍:0–9 隻蝸牛 / 25 ×
25 平方公分)。然而,在 ZINB 迴歸分析中,納入棲地變數的完整模式並未比簡化 模式顯著增加其對於蝸牛密度變異的解釋(likelihood ratio test:χ2 = 2.09,df = 1,P
= 0.15),亦即代表蝸牛密度在二林相間未達統計上的顯著差異。另外,蝸牛密度與 土壤 pH-H2O (r2 = 0.14,P = 0.02,n = 36)、交換性鈣離子(r2 = 0.52,P < 0.001,n =35),
及枯枝落葉的含鈣量(r2 = 0.21,P = 0.005,n = 36)均呈顯著正相關。
2.3.2 添加鈣的攝取
從內襯築巢期至產完蛋的操作期間,針闊葉混淆林的操作組雌鳥(n = 13)攝取 了所添加蝸牛殼的 66.6 ± 15.0 (SD) %,以及所添加蛋殼的 63.8 ± 22.4 (SD) %。相 對的,柳杉針葉林的操作組雌鳥(n = 12)攝取了所添加蝸牛殼的 52.8 ± 23.8 SD %,
以及所添加蛋殼的 46.6 ± 22.7 SD %。在蝸牛殼和蛋殼被攝取比例的 GLMM 模式 中,棲地均非顯著的因子(snail intake model:β estimate = 0.87 ± 1.36,z = 0.64,P = 0.52;eggshell intake model:β estimate = 1.64 ± 1.78,z = 0.92,P = 0.36),亦即表 示蝸牛殼和蛋殼被雌鳥攝取的比例在兩個棲地間無顯著差異。
2.3.3 繁殖表現
於 2011 年至 2014 年期間,於針闊葉混淆林與柳杉針葉林分別紀錄 30 和 26 巢繁殖巢。於 56 巢 292 顆蛋中,只紀錄 1 顆無卵黃的侏儒蛋,其為針闊葉混淆林 的繁殖雌鳥所產下。在研究族群中,並未發現任何沒有殼或薄殼易破裂的蛋。窩 卵數、蛋重、卵形指數、蛋體積、整窩蛋體積以及孵化成功率在二棲地間和二操 作組別間並無顯著差異(表 2.2)。
2.4 討論
本研究證實在亞熱帶臺灣的高山地區,青背山雀的生殖產出未顯著受鈣所限 制。研究地區的土壤和臺灣北部的其他研究相似,均為高度酸性(Horng and Chang 1996)。儘管如此,研究族群的雌鳥產下的蛋,蛋殼並無缺陷的症狀。在二林相繁 殖的鳥巢其繁殖特徵亦無顯著差異,並且人為添加額外的鈣源並不影響鳥巢的生 殖產出。添加鈣對於青背山雀繁殖上的影響效應不高,推測為繁殖親鳥在它們自 然環境中已能獲得足夠的鈣源,因此額外的添加鈣源無法進一步提升繁殖的表現,
這樣的結果與 Johnson and Barclay (1996)的結果一致。添加鈣的操作組雌鳥其獲益 可能反應在其他方面,例如:添加鈣減輕它們的營養或能量需求,降低它們的時 間分配壓力(time budget stress),因而使得雌鳥有更佳的身體狀況(Mänd and Tilgar 2003);或者如在歐洲絲雀(Serinus serinus)的研究,在添加食物操作下產生較多的 偶外配對子代(Hoi-Leitner et al. 1999, also see Reynolds et al. 2017)。
在人為圈養下,產蛋期的大山雀雌鳥每日產下一顆蛋需攝取約 65 毫克的鈣,
相當約 197 毫克的蝸牛殼(Graveland and Berends 1997)。假設青背山雀雌鳥(成鳥體 重:12–15 公克,蛋重:1.2–1.5 公克)的產蛋需求和大山雀(成鳥體重:18–21 公克,
蛋重:1.4–1.6 公克;Graveland and van Gijzen 1994)相似。又,在產蛋期的大山雀 (Betts 1955)與藍山雀(Parus caeruleus;Ramsay and Houston 1999)雌鳥嗉囊內容物 分析中發現,小型陸蝸,如 Discus rotundatus (1.1–4.4 毫米)為它們主要的鈣源。倘 若青背山雀也是依賴小型的陸蝸(<5 毫米,約 1–4 毫克乾重)為它們主要的鈣源(此 類陸蝸也是研究樣區中主要的蝸牛大小),那麼產蛋期的雌鳥每天需攝食 50–200 個小型蝸牛,才能產下一個蛋。二林相之蝸牛密度平均為 40–70 隻蝸牛/平方公尺。
青背山雀平均領域面積約 10,000 平方公尺(Ming-Tang Shiao personal observation),
與大山雀(Wilkin et al. 2006)相近。因此就青背山雀而言,蝸牛可能非現階段的限制 資源。前述的估算是相對保守的,因為鳥類比人更善於搜尋蝸牛(Ramsay and Houston 1999),並且它們除了取食枯枝落葉上的蝸牛,還能取食樹棲性蝸牛 (arboreal snails),而樹棲性蝸牛不易量化調查,並在目前的研究中未納入考量。另 外,產蛋期的雌鳥也可能飛到領域外去尋找並獲取足夠的鈣(Wilkin et al. 2009)。不 過,對鳥類而言,搜尋並攝取許多蝸牛是耗時的活動(Turner 1982, Graveland and Berends 1997),因而可能在時間分配下受限(time budget constraints)。
相較於過往溫帶森林的研究(Graveland and van der Wal 1996, Mänd et al.
2000a),本研究棲地的鈣可獲性介於在富鈣與缺鈣棲地之間。本研究二林相的土壤 pH 均高於 Buunderkamp 森林中的缺鈣棲地,而在該棲地繁殖的鳥巢有 10–60 %已 出現蛋殼缺陷的情況(Graveland and van der Wal 1996)。相反的,本研究的土壤 pH 低於該研究(Graveland and van der Wal 1996)所指的富鈣棲地,並且本研究二林相的 蝸牛密度接近前人研究所指的富鈣棲地之最低閥值(Graveland and van der Wal 1996, Tilgar et al. 1999, Mänd et al. 2000a;表 2.3),低於這個閥值以下,鳥巢的生
殖產出就可能受鈣所限制。不過,青背山雀產下的窩卵數(4–6 顆蛋,極少數為 7 顆蛋)少於歐洲的大山雀(5–12 顆蛋;Perrins 1979),因此青背山雀產下整窩蛋所需 的鈣少於溫帶森林的大山雀,因此對於鈣的缺乏,可能也相對較不敏感。
在受人為干擾的地景上,鳥類可能占據非它們傳統使用的棲地(Newton 1998)。
倘若鳥類在非原生棲地上,其生殖產出仍舊依循它在原始棲地的表現,而未依據 實際棲地的品質來調整,那麼他們的繁殖表現可能變成不適應的。例如:產下高 於(Virkkala 1990, Rytkönen and Orell 2001)或低於(Mänd et al. 2005)現存環境所能 支持的蛋數。研究結果顯示,柳杉針葉林的蝸牛密度與針闊葉混淆林相近,甚至 更高。並且,研究發現柳杉針葉林的雌鳥產下的窩卵數與針闊葉混淆林相當。因 此,鈣並非是在柳杉針葉林繁殖的青背山雀於產蛋上的限制營養。然而,相較於 在原始森林繁殖的個體,在柳杉針葉林繁殖的青背山雀可能是較不適應的,因為 日本柳杉針葉林所能提供的毛蟲生物量低於原始的闊葉森林所能供給的食物量 (Mizutani and Hijii 2002, 本研究第三章)。因此,在柳杉針葉林的青背山雀產下和 針闊葉混淆林族群相似的窩卵數可能是較不利的,而較低的食物豐度可能不利於 雛鳥的生長及離巢後的存活(i.e., Virkkala 1990, Rytkönen and Orell 2001, Tremblay et al. 2003)。
總結而言,由研究結果與鈣需求的計算,推測鈣並非現階段臺灣山區青背山 雀產蛋的限制營養。原始林(針闊葉混淆林)與干擾後棲地(柳杉針葉林)均能提供足 夠的鈣給產蛋期的雌鳥。兩種林相在枯枝落葉品質上的差異影響其土壤化學性質。
柳杉針葉林所能支持的蝸牛密度與針闊葉混淆林相似、甚至更高,是因為柳杉針 葉林的枯枝落葉具有較多的鈣,導致土壤有較高的 pH 及交換性鈣離子。不過,研 究地區仍然受酸沉降的威脅,倘若本區土壤酸化開始影響蝸牛密度,將導致鈣可 獲性的減少,並進一步限制青背山雀的生殖產出。為此,持續監測土壤養分、蝸 牛密度及青背山雀繁殖參數之變化,將可提供酸沉降對亞熱帶臺灣之高山森林影 響之重要資訊。
2.5 參考文獻
Betts, M. M. (1955). The food of titmice in oak woodland. Journal of Animal Ecology 24:282–323.
Bolker, B.M., M. E. Brooks, C. J. Clark, S. W. Geange, J. R. Poulsen, M. H. H. Stevens, and J.-S. S.White (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology and Evolution 24:127–135.
Bonham, K. J., R. Mesibov, and R. Bashford (2002). Diversity and abundance of some ground-dwelling invertebrates in plantation vs. native forests in Tasmania, Australia.
Forest Ecology and Management 158:237–247.
Cameron, R. A. D. (1978). Terrestrial snail faunas of the Malham area. Field studies 4:715–728.
Chang , C.-T., T.-C. Lin, and N.-H. Lin (2009). Estimating the critical load and the environmental and economic impact of acid deposition in Taiwan. Journal of Geographical Science 56:39–58 (In Chinese with English abstract).
De Vries, W. and A. Breeuwsma (1986). Relative importance of natural and anthropogenic proton sources in soils in The Netherlands. Water, Air, and Soil Pollution 28:173–184.
Gärdenfors, U., H. W. Waldén, and I. Wäreborn (1995). Effects of soil acidification on forest land snails. Ecological Bulletins 44:259–270.
Gilbert, A. B. (1971). The egg: its physical and chemical aspects. In Physiology and Biochemistry of the Domestic Fowl, Vol. 3 (D. J. Bell and B. M. Freeman, Editors).
Academic Press, NY, USA.
Gilbert, A. B. (1983). Calcium and reproductive function in the hen. Proceedings of the Nutrition Society 42:195–212.
Graveland, J. (1996). Avian eggshell formation in calcium-rich and calcium-poor habitats: importance of snail shells and anthropogenic calcium sources. Canadian Journal of Zoology 74:1035–1044.
Graveland, J. (1998). Effects of acid rain on bird populations. Environmental Reviews 6:41–54.
Graveland, J., and A. E. Berends (1997). Timing of the calcium intake and effect of calcium deficiency on behaviour and egg laying in captive Great Tits, Parus major.
Physiological Zoology 70:74–84.
Graveland, J., and T. van Gijzen (1994). Arthropods and seeds are not sufficient as calcium sources for shell formation and skeletal growth in passerines. Ardea 82:299–314.
Graveland, J., and R. van der Wal (1996). Decline in snail abundance due to soil acidification causes eggshell defects in forest passerines. Oecologia 105:351–360.
Graveland, J., R. van der Wal, J. H. van Balen, and A. J. van Noordwijk (1994) Poor reproduction in forest passerines from decline of snail abundance on acidified soils.
Nature 368:446–448.
Hoi-Leitner, M., H. Hoi, M. Romero-Pujante, and F. Valera (1999). Female extra-pair behaviour and environmental quality in the Serin (Serinus serinus): a test of the
‘constrained female hypothesis’. Proceedings of the Royal Society of London, Series B 266:1021–1026.
Horng, F.-W., and W.-E. Chang (1996). Soil nutrient pool and available nutrient dynamics in the Fushan mixed hardwood forest ecosystem. Taiwan Journal of Forest Science 11:465–473 (In Chinese with English abstract).
Hoyt, D. F. (1979). Practical methods of estimating volume and fresh weight of bird eggs. The Auk 96:73–77.
Hsiao, H.-M., T.-C. Lin, J.-L. Hwong, C.-C. Huang, and N.-H. Lin (2007). Precipitation chemistry at the Lienhuachi Experimental Forest in central Taiwan. Taiwan Journal of Forest Science 22:1–13.
Johannessen, L. E., and T. Solhøy (2001). Effects of experimentally increased calcium levels in the litter on terrestrial snail populations. Pedobiologia 45:234–242.
Johnson, L.S., and R. M. R. Barclay (1996). Effects of supplemental calcium on the reproductive output of a small passerine bird, the House Wren (Troglodytes aedon).
Canadian Journal of Zoology 74:278–282.
Lin, T.-C., S.-P. Hamburg, H.-B. King, and Y.-J. Hsia (2000). Throughfall patterns in a subtropical rain forest of northeastern Taiwan. Journal of Environmental Quality 29:1186–1193.
Liang, Y.-L., T.-C. Lin, J.-L. Hwong, N.-H. Lin, and C.-P. Wang (2009). Fog and precipitation chemistry at a mid-land forest in central Taiwan. Journal of Environmental Quality 38:627–636.
Mänd, R., and V. Tilgar (2003). Does supplementary calcium reduce the cost of reproduction in the Pied Flycatcher Ficedula hypoleuca? Ibis 145:67–77.
Mänd, R., V. Tilgar, and A. Leivits (2000a). Calcium, snails, and birds: a case study.
Web Ecology 1:63–69.
Mänd, R., V. Tilgar, and A. Leivits (2000b). Reproductive response of Great Tits, Parus major, in a naturally base-poor forest habitat to calcium supplementation. Canadian Journal of Zoology 78:689–695.
Mänd, R., V. Tilgar, A. Lõhmus, and A. Leivits (2005). Providing nest boxes for hole-nesting birds – Does habitat matter? Biodiversity and Conservation 14:1823–1840.
Martin, K., and M. Sommer (2004). Relationships between land snail assemblage patterns and soil properties in temperate-humid forest ecosystems. Journal of Biogeography 31:531–545.
Mizutani, M., and N. Hijii (2002). The effects of arthropod abundance and size on the nestling diet of two Parus species. Ornithological Science 1:71–80.
Newton, I. (1998) Population limitation in birds. Academic Press, London, UK.
Ohta, T., S. Niwa, and T. Hiura (2014a). Calcium concentration in leaf litter affects the abundance and survival of crustaceans in streams draining warm-temperate forests.
Freshwater biology 59:748–760.
Ohta, T., S. Niwa, N. Agetsuma, and T. Hiura (2014b). Calcium concentration in leaf litter alters the community composition of soil invertebrates in warm-temperate forests. Pedobiologia 57:257–262.
Perrins, C. M. (1979). British tits. William Collins Sons & Co, London, UK.
Qiu, L.-W., Q.-X. Huang, C.-C. Wu, and H.-T. Hsieh (2015) Summary of the fourth national forest resource inventory report. Taiwan Forestry Journal 41:3–13 (In Chinese).
R Development Core Team. (2017). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://
www.r-project.org/.
Ramsay, S. L., and D. C. Houston (1999). Do acid rain and calcium supply limit eggshell formation for Blue Tits (Parus caeruleus) in the U.K.? Journal of Zoology 247:121–125.
Reich, P. B., J. Oleksyn, J. Modrzynski, P. Mrozinski, S. E. Hobbie, D. M. Eissenstat, J.
Chorover, O. A. Chadwick, C. M. Hale, and M. G. Tjoelker (2005). Linking litter calcium, earthworms and soil properties: a common garden test with 14 tree species.
Ecology Letters 8:811–818.
Reynolds, S. J. (2003). Mineral retention, medullary bone formation, and reproduction in the White-tailed Ptarmigan (Lagopus leucurus): a critique of Larison et al. (2001).
The Auk 120:224–228.
Reynolds, S. J., and C. M. Perrins (2010). Dietary calcium availability and reproduction in birds. Current Ornithology 17:31–74.
Reynolds, S.J., J. A. Galbraith, J. A. Smith, and D. N. Jones (2017). Garden bird feeding: insights and prospects from a North-South comparison of this global urban phenomenon. Frontiers in Ecology and Evolution 5: Article 24.
Rusek, J., and V. G. Marshall (2000). Impacts of airborne pollutants on soil fauna.
Annual Review of Ecology and Systematics 31:395–423.
Rytkönen, S., and M. Orell (2001). Great Tits, Parus major, lay too many eggs:
experimental evidence in mid-boreal habitats. Oikos 93:439–450.
Sawyer, J. (1993). Plantation in the tropics: environmental concerns. IUCN, Gland, Switzerland and Cambridge, UK.
Simkiss, K. (1967). Calcium in reproductive physiology: a comparative study of vertebrates. Chapman & Hall, London, UK.
Tattersfield, P., M. B. Seddon, and C. N. Lange (2001). Land-snail faunas in indigenous rainforest and commercial forestry plantations in Kakamega Forest, western Kenya.
Biodiversity and Conservation 10:1809–1829.
Tilgar, V., R. Mänd, and A. Leivits (1999). Effect of calcium availability and habitat quality on reproduction in Pied Flycatcher Ficedula hypoleuca and Great Tit Parus major. Journal of Avian Biology 30:383–391.
Tilgar, V., R. Mänd, and M. Mägi (2002). Calcium shortage as a constraint on
reproduction in Great Tits Parus major: a field experiment. Journal of Avian Biology 33:407–413.
Tremblay, I., D. W. Thomas, M. M. Lambrechts, J. Blondel, and P. Perret (2003).
Variation in Blue Tit breeding performance across gradients in habitat richness.
Ecology 84:3033–3043.
Turner, A. K. (1982). Timing of laying by Swallows (Hirundo rustica) and Sand Martins (Riparia riparia). Journal of Animal Ecology 51:29–46.
Virkkala, R. (1990). Ecology of the Siberian Tit Parus cinctus in relation to habitat quality: effects of forest management. Ornis Scandinavica 21:139–146.
Wilkin, T.A., D. Garant, A. G. Gosler, and B. C. Sheldon (2006). Density effects on life-history traits in a wild population of the Great Tit Parus major: analyses of long-term data with GIS techniques. Journal of Animal Ecology 75:604–615.
Wilkin, T. A., A. G. Gosler, D. Garant, S. J. Reynolds, and B. C. Sheldon (2009).
Calcium effects on life-history traits in a wild population of the Great Tit (Parus major): analysis of long-term data at several spatial scales. Oecologia 159:463–472.
Xue, L., and S. Luo (2002). Seasonal changes in the nutrient concentrations of leaves and leaf litter in a young Cryptomeria japonica stand. Scandinavian Journal of Forest Research 17:495–500.
Zinke, P. J. (1962). The pattern of influence of individual forest trees on soil properties.
Ecology 43:130–133.
表 2.1、觀霧地區針闊葉混淆林與柳杉針葉林枯枝落葉及土壤參數之比較
針闊葉混淆林 柳杉針葉林
P value mean ± SD n mean ± SD n
Litter
Calcium concentration (mg/g) 4.90 ± 2.17 20 6.33 ± 2.78 16 0.090a Soil
pH-H2O 3.92 ± 0.30 20 4.62 ± 0.81 16 0.005b pH-KCl 3.14 ± 0.34 20 3.48 ± 0.38 16 0.009a Exchangeable Ca2+ (cmol/kg) 1.69 ± 1.44 19 4.47 ± 5.45 16 0.028b Cation exchange capacity (cmol/kg) 37.74 ± 7.30 20 31.49 ± 6.43 16 0.011a
a One-way ANOVA test; bShapiro-Wilk test
表 2.2、模式分析 2011–2014 年中海拔觀霧地區青背山雀之(a)蛋形質特徵(linear mixed model,LMM)、(b)窩卵數(generalized linear model,GLM)和(c)孵化成功率 (generalized linear mixed-effects model,GLMM)。成鳥個體 ID 及年均納入 LMMs 和 GLMM 中作為隨機效應。各項變數之係數及顯著性係在逐步向後法建模的過程,
簡化模式加上該項變數下,該項變數之數值
β ± SE Wald statisticsa P value a. Egg traits
Egg mass (n = 56 nests)
Intercept 1.43 ± 0.01 95.43 < 0.001 Habitatb -0.01 ± 0.03 -0.39 0.70 Treatmentc 0.02 ± 0.02 0.72 0.47 Habitat x treatment 0.004 ± 0.050 0.08 0.94 Egg shape index (n = 56 nests)
Intercept 0.75 ± 0.01 137.6 < 0.001
Habitatb 0.01 ± 0.01 0.95 0.35
Treatmentc -0.004 ± 0.007 -0.59 0.56 Habitat x treatment 0.01 ± 0.01 0.98 0.34 Egg volume (n = 56 nests)
Intercept 1408.2 ± 16.3 86.28 < 0.001
Habitatb -4.8 ± 30.9 -0.16 0.88
Treatmentc 15.1 ± 25.5 0.59 0.56 Habitat x treatment -22.7 ± 51.5 -0.44 0.66 Clutch volume (n = 56 nests)
Intercept 7254.5 ± 211.3 34.3 < 0.001 Habitatb -432.5 ± 341.5 -1.27 0.21 Treatmentc -34.1 ± 291.1 -0.12 0.91 Habitat x treatment -506.3 ± 582.0 -0.87 0.39 b. Clutch size (n = 56 nests)
Intercept 1.65 ± 0.06 28.22 < 0.001 Habitatb -0.05 ± 0.12 -0.40 0.69 Treatmentc -0.03 ± 0.12 -0.29 0.77
Habitat x treatment -0.06 ± 0.23 -0.24 0.81 c. Hatching success (n = 46 nests)
Intercept 2.39 ± 0.51 4.72 < 0.001 Habitatb -0.29 ± 0.57 -0.52 0.61 Treatmentc 0.18 ± 0.51 0.35 0.73 Habitat x treatment 1.70 ± 1.00 1.71 0.09
a Wald t test 為 LMMs 分析,Wald z test 為 GLM 和 GLMM 分析。
b棲地(Habitat)為類別變數(針闊葉混淆林或柳杉針葉林),並以柳杉針葉林為參考 值。
c操作(Treatment) 為類別變數(添加鈣操作或控制組),並以控制組為參考值。
表 2.3、各研究調查森林陸生蝸牛密度之比較
Country Forest type Year Snail density
(ind./m2) Reference Calcium-poor forests
Estonia Pine forests 1995 0–2
Mänd et al. 1997 Pine forests 1995 10–48
Pine forests 1995, 1999 13–75 Tilgar et al. 1999, Mänd et al. 2000 Netherlands Pine forests 1970 34–81
Graveland and van der Wal 1996 Pine forests 1992 7–39
Mixed coniferous
-deciduous forests 1992 0–826 Taiwan Mixed-oak forest 2013 43a
This study Cryptomeria plantation 2013 69a
Calcium-rich forests
Netherlands Ash forests 1973 35–304
Graveland and van der Wal 1996 Ash forests 1992 72–206
Oak forests 1992 58–1581 Estonia Deciduous/ mixed
forests 1995 69–219 Mänd et al. 1997 Deciduous forests 1995, 1999 92–214 Tilgar et al. 1999,
Mänd et al. 2000 Deciduous/ mixed
forests 1995 146–427 Mänd et al. 1997
a 以該林相內所有採樣樣方之平均密度換算之。
第三章 育雛期雛鳥生長 3.1 前言
食物豐度是影響鳥類繁殖表現的重要近因(Lack 1968)。對於季節性繁殖的鳥類,
繁殖特徵受季節性食物變化所形塑,當雛鳥食物需求的高峰與食物豐度高峰相一 致時,將有最大的適存值(reproductive fitness;Verboven et al. 2001, Tremblay et al.
2003)。雖然鳥類與它們獵物間營養階層的交互作用(trophic interaction)在溫帶地區 的多種棲地已被廣泛地研究(van Balen 1973, Rytkönen and Orell 2001, Massa et al.
2004, Ziane et al. 2006, Lambrechts et al. 2008),然而在低緯度的生態群落卻少有研 究。低緯度的生態群落,如副熱帶季風林生態系仍具一定程度的季節性,包括乾 濕季週期、溫度變化及光週期等,因而影響鳥類的繁殖時間(Tye 1992, Wikelski et al.
2000, Shiao et al. 2015)。並且,低緯度的生態群落較複雜且多樣化,因而其營養階 層間的交互作用可能也不同於物種數相對較少的溫帶地區(Şekercioğlu et al. 2012)。
為此,欲探討食物資源對於低緯度地區鳥類繁殖的影響程度,須調查棲地的食物 資源季節性變化,並與鳥類的繁殖表現相連結。
棲地類型決定植被的種類,並影響食蟲性鳥類最主要的食物─鱗翅目與膜翅目 幼蟲(Lepidoptera and Hymenoptera larvae)的豐度與多樣性(van Balen 1973)。在溫帶 地區,落葉性樹種的幼蟲生物量在初夏呈現一個短暫、急遽變化的高峰(Verboven et al. 2001, Veen et al. 2010, Smith et al. 2011);相對地,松科樹種的幼蟲生物量則隨 著季節呈穩定緩慢地增加,其高峰較低且較晚(van Balen 1973, Veen et al. 2010)。
相較於溫帶地區狹窄的食物豐度高峰,低緯度的生態群落具有較高的樹種多樣性,
因而假設將形成較寬的幼蟲豐度高峰(Tremblay et al. 2003)。不過,目前未有研究 證實這個假說。此外,不同棲地的幼蟲在體型、外觀及營養含量上可能不同(Isaksson and Andersson 2007, Arnold et al. 2010),因而影響食物的品質和鳥類的覓食選擇 (e.g., Naef-Daenzer et al. 2000, Tremblay et al. 2005, García-Navas and Sanz 2011)。例 如:大山雀偏好大型的夜蛾(Noctuidae)幼蟲,而藍山雀則不吃多毛的枯葉蛾 (Lasiocampidae)幼蟲(García-Navas et al. 2013)。為此,在評估不同棲地的鳥類的食 物資源上,幼蟲的量和品質均須納入考量。
食物的可獲得性限制鳥類的繁殖表現(Martin 1987)。在食物豐富的環境,親鳥
能耗費較少的時間與能量在覓食活動,並通常帶回較大的獵物(Naef-Daenzer and Keller 1999, Tremblay et al. 2005);親鳥通常會較早開始產蛋,並產下比較大的窩卵 數(Blondel et al. 1993, Massa et al. 2004, Sanz et al. 2010),雛鳥的體重較重,離巢幼 鳥數也較高(Verboven et al. 2001, Tremblay et al. 2003)。因此,雛鳥食物需求高峰與 幼蟲豐度高峰相吻合對鳥類的繁殖是有益的(van Noordwijk et al. 1995, Thomas et al. 2001)。然而,對於兼行兩巢的鳥種,親鳥需在第一窩與第二窩繁殖間最適化它 們的生殖投資,而幼蟲豐度亦隨著棲地類型與季節變化而有不同。van Balen (1973) 在落葉橡樹林和松科造林地上,觀察發現大山雀在不同棲地類型具不同的繁殖表 現(Habitat-specific breeding performances)。落葉橡樹林的幼蟲豐度高峰使得大山雀 第一窩雛鳥有較重的體重,而後幼蟲豐度逐漸減少,第二窩雛鳥體重也隨著降低;
在松科造林地則反映出相反的趨勢,幼蟲高峰出現較晚,而第二窩的雛鳥亦較第 一窩略重。然而,少有研究比較不同棲地的食物季節性變化,並與親鳥的繁殖表 現相結合(e.g., Mägi and Mänd 2004, Lambrechts et al. 2008, Mägi et al. 2009),而其 結果因棲地類型而異。
本研究的目標為瞭解在針闊葉混淆林(以下簡稱闊葉棲地)與柳杉針葉林(以下 簡稱針葉棲地)兩種棲地下,幼蟲季節性豐度變化與其多樣性對食蟲性青背山雀的 繁殖表現之影響。青背山雀雛鳥的食譜主要以鱗翅目與膜翅目的幼蟲為主,占 71%(Chuang 2006)。全臺森林總面積有 21%為人工林,而柳杉針葉林是臺灣重要 的造林林型,占全臺人工林面積之 11%(Qiu et al. 2015)。人工林的結構相對簡單,
林齡均一,植被多樣性少,以至其食物可能較缺乏,鳥類而言為較差的棲地。在 天然與人工林鑲嵌的地景下,鳥類對這非原生地造林地之反應,並未有深入的研 究,而它們的繁殖特徵也未可知。為此,本研究於 2015 年(1) 調查兩種林相中 5 種優勢樹種:2 種常綠橡樹和 1 種落葉榿木(闊葉棲地),以及 2 種杉(針葉棲地)的 鱗翅目及膜翅目幼蟲之季節性動態以及多樣性組成,同時(2) 調查雛鳥的生長,並 估計在每巢雛鳥需求高峰時,環境的幼蟲生物量,接著檢視其是否可解釋雛鳥的 體重變異;最後,(3) 檢視多年(2010–2017)的繁殖資料,比較在闊葉棲地與針葉棲 地其各項繁殖特徵,包括年繁殖密度、第二窩比例、產蛋日、窩卵數等。吾人預 測幼蟲生物量的季節性變化和組成在不同樹種及棲地間不同,如同 Tremblay et al.
(2003)所預測的,闊葉棲地具較寬的幼蟲高峰,而針葉棲地的幼蟲生物量及品質均
較差,並且不同棲地的幼蟲生物量之物候變化將影響雛鳥的生長。
3.2 研究方法
3.2.1 雛鳥狀況評估與資料定義
除例行性每週至少一次的巡巢、產蛋日及新生雛鳥日齡之推估、離巢前雛鳥 和無腳環親鳥的繫放外,在進行幼蟲物候調查的 2015 年,雛鳥於 13–14 日齡時,
以口袋電子秤(JYA–150,Jin Yuan,臺灣)秤重,量測至 0.1 公克,計算整巢之平均 雛鳥體重作為該巢雛鳥狀況之指標;另外,以電子游標卡尺(CD–6” CSX,Mitutoyo,
日本)測量雛鳥跗趾長,量測至 0.01 毫米。
兩種棲地之年繁殖密度係以該年同時繁殖鳥巢之最大值除以棲地面積來估算。
青背山雀一年產 1–2 巢。依據 van Noordwijk et al. (1995)和吾人野外經驗將繁殖嘗 試(breeding attempt)分成:(1)第一窩,其產蛋日位在最早繁殖鳥巢之產蛋日往後 30 天以內的區間;以及(2)第二窩,其產蛋日位在 30 天以後的區間,第二窩繁殖巢也 包括第一窩繁殖失敗後的補充巢(replacement brood),它們和第二窩處在相同時空 環境。另計算二林相內,第二窩繁殖巢數相對於第一窩繁殖巢數之比例,定義為 產第二窩比例(second brood proportion)。
3.2.2 幼蟲生物量物候和組成
吾人於 2015 年 2 月 10 日至 8 月 24 日期間,針對二林相內相對優勢度最高的 樹種,以抖落法(tree-shaking method;Hsu 2010)調查其幼蟲組成及生物量變化。在 針闊葉混淆林內選取常綠闊葉樹的森氏櫟(RD:19%)和卡氏櫧(RD:17%)、以及落 葉闊葉樹的臺灣赤楊(RD:11%);於柳杉針葉林內,則選取常綠針葉樹的柳杉(RD:
54%)和臺灣杉(RD:25%;詳見第一章 1.2 研究樣區與巢箱設置)。吾人未使用國外 常用的蟲糞掉落法之原因,為蟲糞法較適用在昆蟲種類少且形成單一短暫的蟲糞 高峰的溫帶地區,而不適用於具重濕(perhumid)、多雨氣候(Tinbergen and Dietz 1994) 的觀霧地區。抖落法係針對幼蟲受驚嚇時,掉落裝死的特性,以採集掉落的幼蟲。
少數種類如刺蛾科(Limacodidae)幼蟲為爬行黏附在植物枝葉,不易以抖落法採集,
然因刺蛾本身具毒性,非鳥類取食的獵物,故對於本研究標的(探討幼蟲生物量與 雛鳥之關聯)之影響不大。