國立臺灣大學理學院化學研究所 碩士論文
Department of Chemistry College of Science
National Taiwan University Master Thesis
利用單分子技術探討 DNA 重組酶 Rad51 與 Dmc1 核蛋白絲之成核動力學 Studying the Assembly Kinetics of Saccharomyces
cerevisiae Rad51 and Dmc1 Recombinases Using Single Molecule Methods
林聖堯 Sheng-Yao Lin
指導教授:李弘文 博士 Advisor: Hung-Wen Li, Ph.D.
中華民國 106 年 6 月 June, 2017
誌謝
首先要特別謝謝我的家人,自從上了大學之後與你們相處的時間就變得很少,
即使如此你們仍然在我需要的時候幫助我、成為我的心靈支柱。如果說我能在任何 方面有所成就,絕對是因為有你們的支持讓我可以跨越障礙。
再來要感謝弘文老師在這四年中提供了許多的實驗機會讓我嘗試並增進自己 無論實驗上或是面對障礙並嘗試找到突破口的可能性,還記得一開始身為專題生 時,還沒有對於自己的實驗有責任感,也理所當然實驗進度停滯不前。要謝謝老師 的諄諄教誨讓我開始理解到”要將實驗視為自己的責任”而不是做完敷衍了事的工 作而已。也要謝謝老師在實驗上總是熱情的與我們討論實驗的可能性,並提供許多 參加研討會的機會並提供經費上的支持,讓我從一開始容易怯場到現在可以比較 勇於與他人討論表達實驗結果。
感謝冀宏源老師在實驗上提供蛋白與時常與我們進行討論,共同推進實驗的 進度,並擔任我的口試委員。感謝李以仁與溫進德老師在實驗室的合作討論中提供 許多實驗思考的方向。謝謝范秀芳老師、王廷方老師、林敬哲老師、陳昭岑老師擔 任我的口試委員並提供許多珍貴的實驗討論。
再來要謝謝實驗室的大家,謝謝興雅學姐與靜如學姐在我剛進實驗室耐心指 導我實驗的技巧;謝謝黃韜學長、泓其學長與慧親學姐願意與剛進實驗室的我討 論實驗失敗的原因;謝謝文軒學長即使畢業後仍然會抽空與我討論實驗的內容;
謝謝泓儀學長與家傳學長與我討論實驗並提出建議;謝謝誌遠學長提供 ScDmc1 與 ScRad51 進行實驗並以巨觀實驗提供研究上的資訊;謝謝有耐心的宇軒與好相 處但小劇場有點多的庭慈無論在討論實驗或是生活瑣事上都讓我獲益良多;謝謝 好脾氣的煒翔在實驗上的幫助或是軍中生活的分享;謝謝聰明的致豪在 DNA 合 成上提供許多寶貴的意見、也謝謝在日本時常常陪我做實驗到晚上十點多;謝謝 認真的庭尉在實驗上的提問常常讓我反思與生活上的打屁聊天;謝謝翰霖、晏 展、林昀、邦瑀、許永、邦硯、李微為實驗室帶來需多歡笑。
謝謝在生命歷程中幫助過我的每個人,我能成為現在的我,都是因為有與你 們的相處累積而來的。
摘要
雙股斷裂的 DNA 損壞,可利用同源重組(homologous recombination)的方式,
準確地修復受損的 DNA。DNA 重組酶催化受損 DNA 與同源的雙股 DNA 配對,
進行股交換反應,進而利用完整的 DNA 模板進行修復。真核生物具有兩種同源重 組酶 Dmc1 與 Rad51,其序列及功能大多相似,但兩者在細胞週期出現的時間點 卻有不同。在細胞分裂時(mitosis)只需 Rad51 即可完成 DNA 修復,然而在減數分 裂(meiosis)的過程中,則同時需要 Rad51 與 Dmc1 兩者的參與。為何不同的細胞階 段對 DNA 重組酶的需求不同,其機制仍然未明。利用單分子栓球實驗(single molecule tethered particle motion),本研究直接比較 Saccharomyces cerevisiae 酵母 菌中 Rad51 與 Dmc1 兩個重組酶在形成核蛋白絲階段的動力學。我們觀察到 ScRad51 與 ScDmc1 在核蛋白絲的形成動力學明顯不同。在核蛋白絲的形成過程,
成核的階段是速率決定步驟,而 ScRad51 的成核速率比 ScDmc1 來的快,但兩者 在其核蛋白絲的延展及核蛋白絲的長度上,則沒有差別。不同的重組酶濃度實驗中 指出,ScRad51 與 ScDmc1 皆以二倍體(dimer)的形式進行成核,因此成核速率的差 異是來自於 ScRad51 對單股 DNA (ssDNA)有較高的親和力,這個現象也反應在 ScRad51 的成核速率隨著不同長度的單股 DNA 而明顯遞增。相對來說,ScDmc1 的成核速率在不同長度的單股 DNA 中則沒有明顯關聯,而是與單雙股交界處的連 接點(ds/ss DNA junction)處的成核速率相似。Rad51 與 Dmc1 核蛋白絲的形成動力 學上的差異,及其對於不同 DNA 構型的偏好性,在建構同源重組在減數分裂過程 中進行的分子模型時,可以提供這兩種 DNA 重組酶角色的重要基礎。
ABSTRACT
The double-stranded break (DSB) DNA damage can be repaired in high fidelity using homologous recombination (HR) pathway. DNA recombinases form nucleoprotein filaments and catalyze the pairing of the homologous DNA sequence and the exchange of DNA strands. So this allow DNA replication to repair the damaged DNA using the homologous DNA template. In eukaryotes, there exist two recombinases, Rad51 and Dmc1. Both of them share similar amino acid sequences and functions. However, while only Rad51 is required in mitosis, both Rad51 and Dmc1 are essential in meiosis recombination. The mechanism underlying the differential requirement is unknown. Here, we compared the kinetics of the nucleoprotein filament assembly of Saccharomyces cerevisiae ScRad51 and ScDmc1 using single-molecule tethered particle motion experiments (TPM). We found an apparent kinetics difference during nucleoprotein filament assembly for ScRad51 and ScDmc1. Forming recombinase nuclei in single- stranded (ss) DNA is the rate-limited step in the nucleoprotein filament assembly. In our real-time assembly measurement, we found that ScRad51 has much faster nucleation rate than ScDmc1, while the extension time and filament coverage are similar for both ScRad51 and ScDmc1. Study of the nucleation times at different recombinase concentrations showed that ScRad51 and ScDmc1 have the similar power-law dependence of ~ 2, suggesting that both form stable nuclei in dimers. Therefore, the faster nucleation of ScRad51 likely results from the higher ssDNA affinity. This is consistent with the observation that nucleation times of ScRad51 increases with DNA substrates of longer ssDNA lengths. However, nucleation times of ScDmc1 did not show apparent ssDNA length dependence. These kinetic differences in nucleoprotein filament assembly provide important molecular constrains in explaining biochemical roles of Rad51 and Dmc1.
Keywords: DNA Recombinases, Rad51, Dmc1, nucleorpotein filament, DNA homologous recombinational repair, single molecule Tethered Particle Motion
TABLE OF CONTENTS
摘要 ... i
ABSTRACT ... ii
TABLE OF CONTENTS ... iii
LIST OF FIGURES ... v
LIST OF TABLES ... vi
CHAPTER 1. INTRODUCTION ... 1
1-1 Homologous recombination (HR) ... 1
1-2 Dmc1 and Rad51 Recombinase ... 3
1-3 Calcium modulates the activity of Rad51 and Dmc1 ... 5
1-4 Motivation of this study ... 7
CHAPTER 2. MATERIALS AND METHODS ... 8
2-1 Rad51, Dmc1, Mei5-Sae3 protein purification ... 8
2-2 gap DNA dT200, AC263 synthesis method ... 8
2-2.1 AC263 gap DNA synthesis ... 8
2-2.2 dT gap synthesis ... 8
2-3 Buffer Recipe ... 10
2-4 Single-molecule Tethered Particle Motion (TPM) ... 11
2-4.1 microscope parameters ... 12
2-4.2 Reaction slide preparation ... 13
CHAPTER 3. RESULT ... 17
3-1.ScRad51 has fast nucleation rate than Dmc1 ... 17
3-2 dsDNA assembly control comparison between Rad51 and Dmc1 ... 19
3-3 Dmc1-DNA interaction is different from Rad51-DNA. ... 21
3-4 The nucleation rates of ScDmc1 and ScRad51 have the similar recombinase- concentration dependence... 22
3-5 Rad51 has higher assembly preference on ssDNA than Dmc1 ... 25
3-6 Calcium stimulates Dmc1’s nucleation ... 29
3-7 ScRad51 doesn’t stimulate the nucleation step of ScDmc1. ... 31
CHAPTER 4.CONCLUSION AND OUTLOOK ... 34
4-1 Conclusion ... 34
4-2 Outlook ... 37
Reference ... 39
Appendix ... 43
LIST OF FIGURES
Figure 1-1 Double-strand break repair by the homologous recombination... 2
Figure 1-2 Similarities and differences among recombinases from different species. ... 4
Figure 1-3 Calcium ions affect the hRad51 and hDmc1 joint molecule formation. ... 6
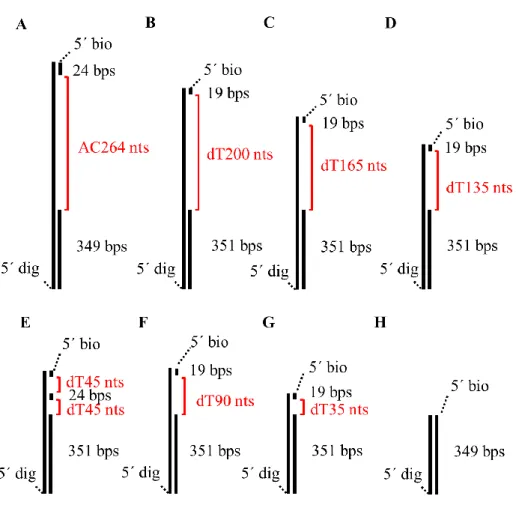
Figure 2-1 The DNA substrates were used in this study. ... 9
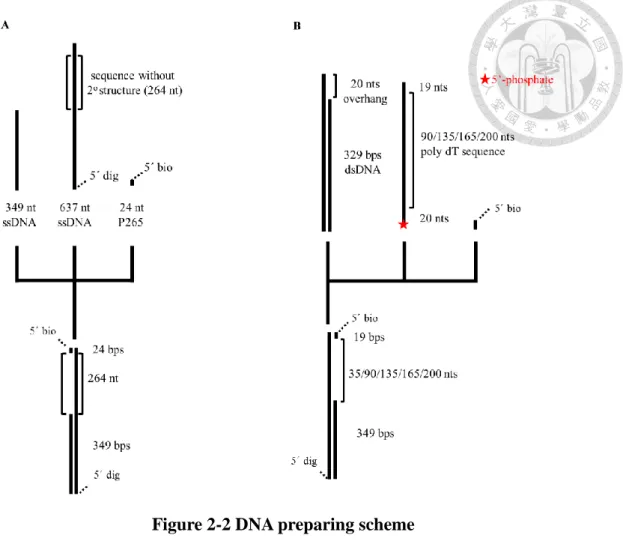
Figure 2-2 DNA preparing scheme ... 10
Figure 2-3 The Brownian motion value of DNA tethered beads varies linearly with DNA tether lengths. ... 12
Figure 2-4 recombinase assembly assay using TPM experiments ... 14
Figure 2-5 The nucleation time is determined by the cumulative histogram of time of individual time traces. ... 15
Figure 3-1 Dmc1 shows slower assembly kinetics than Rad51. ... 18
Figure 3-2 Four interaction modes of ScDmc1 and ScRad51. ... 20
Figure 3-3 The cumulative histograms of nucleation events indicate the nucleation reaction is dependent on the recombinase concentration. ... 23
Figure 3-4 Dmc1 and Rad51 show the similar reaction order in filament assembly. ... 24
Figure 3-5 The lengths of the ssDNA dependence in nucleation reaction. ... 26
Figure 3-6 ScDmc1 and ScRad51 show the different ssDNA affinity. ... 27
Figure 3-7 Ca2+ stimulates ScDmc1’s nucleation reaction. ... 31
Figure 3-8 Nucleation time comparison of Dmc1 with Rad51 ... 33
LIST OF TABLES
Table 2-2. Microscope parameters ... 12
Table 3-1 Nucleation times for each reaction condition in Fig. 3-4 ... 24
Table 3-2 The nucleation time to each gap DNA in Fig.3-6 ... 27
Table 3-3 nucleation rate comparison between ScRad51 and ScDmc1. ... 28
CHAPTER 1. INTRODUCTION
1-1 Homologous recombination (HR)
Homologous recombination (HR) is an important process for maintaining the gene integrity and producing the gene diversity. DNA in cell undergoes double strand break (DSB) when exposed to UV irradiation, chemical environment, or in the sister chromatid pairing in cell division1.
The DSB phenomena could lead to apoptosis without repairing. In HR, the DSB DNA will be degraded to generate a 3' single strand overhang by nuclease and helicase.
Then, the recombinases facilitate the homologous DNA sequence searching by forming presynaptic filament with the 3' overhang DNA. The presynaptic filament formation process can be divided into three steps. First, recombinases form a stable nucleus on ssDNA. Second, free recombinases assemble on the ssDNA from the nucleus and extend the filament. Finally, the filament covers most of the ssDNA and continue the homologous sequence searching. The first part, nucleus formation, is the rate determining step2. The synaptic filament surveys the homologous sequence and invades in it to become a displacement loop (D-loop) for the following strand exchange process. After searching the homologous DNA sequence, the injured DNA use the intact homologous DNA as a template to undergo the replication for the repairing purpose.
Figure 1-1 Double-strand break repair by the homologous recombination The DSB will first be degraded as 3' overhang. With the recombinases and accessory protein help, the injured DNA can search for the homologous sequence for replication and repair. The proposed products of the repaired DNA have two type: synthesis- dependent strand annealing (SDSA) and double-strand break repair (DSBR)3.
1-2 Dmc1 and Rad51 Recombinase
In prokaryote cell, only RecA plays a role of recombinase in HR. On the contrary, in eukaryote cell, there are two recombinases, Rad51 and Dmc1, processing in HR. Patrick Sung and Eric Greene have compared many types of the recombinases in the kinetic process of homologous DNA searching. For E.coli RecA, ScDmc1, and ScRad51, the smallest presynaptic filament searching the homologous sequence in HR process is about 8 nts in size, and all of them share the same stoichiometry ratio of 3 nucleotides to each recombinase4. For the homologous recombinases Rad51 and Dmc1, although the amino acid sequence is highly conserved3, 5, many biochemical assays show their difference.
In cell cycle, Rad51 can function in both meiotic and mitotic cell, while Dmc1 is meiotic-specific. The mechanism underlying these different requirements is still unknown.
During the homologous sequence searching in HR, Dmc1 has higher endurance in mismatch sequence pairing than Rad51 and RecA6, 7. Some previous studies indicate the different DNA-protein structures or functions between these two recombinases. In EM image, both ScDmc1-DNA and hDmc1-DNA interaction have two kinds of filament formation. One is the general extended filament structure as the RecA filament, and the other is a stack-ring structure5, 8, 9. However, the stack-ring structure is absent in Rad51- DNA interaction.
Based on those observations, Rad51 is considered to dominate the general repair process in HR because of the high fidelity of DNA sequence searching. However, owing
single molecule level.
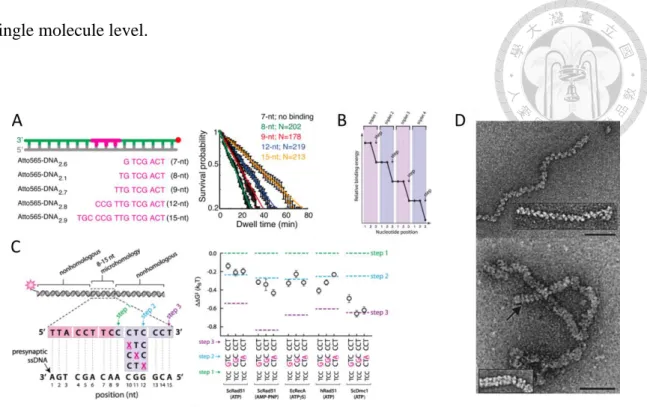
Figure 1-2 Similarities and differences among recombinases from different species.
(A) Measuring the dissociation rate with a fluorophore labeled presynaptic filaments which have different homology lengths DNA, the binding signal starts to appear when the homology presynaptic filament increases to 8 nts10. (B) The filament binding energy change every 3 nucleotides, hint each recombinase bind 3 nucleotides7.(C) With a single mismatch nucleotide, only Dmc1 can endure the mismatch in filament pairing7. (D) Top:
Electron microscopic visualization of hDmc1-ssDNA shows the filaments conformation with ATP. Bottom: Electron microscopic visualization of hDmc1-ssDNA shows the stack- ring conformation with ATP. The magnification bars represent 50 nm11.
1-3 Calcium modulates the activity of Rad51 and Dmc1
The calcium concentration is a signal for cell cycle. Especially in meiosis, the surge of calcium concentration is essential12. In the coincidence of Dmc1 existence and the increased calcium concentration, calcium might affect the activity of Dmc1. Both hDmc1, hRad51, and ScDmc1 D-loop formation and strand exchange can be improved by Ca2+
appearance13, 14, 15. Although the ATP-hydrolyzing activity of both hDmc1 and hRad51 are suppressed, the fact that the activation of D-loop assay has a higher dependence on free Ca2+ than Ca-ATP for hDmc1 indicates the possibility of the second Ca2+ binding site on Dmc114.
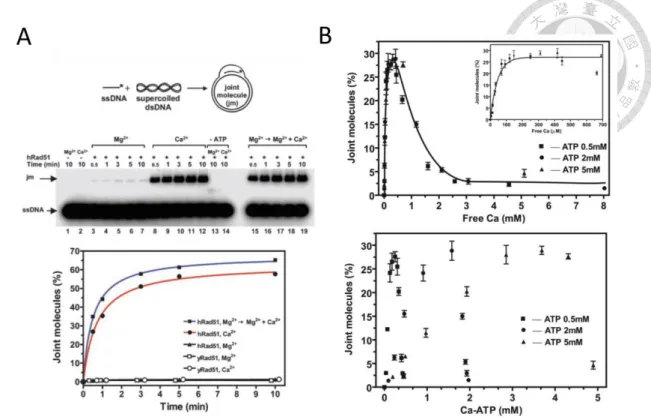
Figure 1-3 Calcium ions affect the hRad51 and hDmc1 joint molecule formation.
(A) For hRad51, the Calcium ion can improve the formation of the joint molecule. While the enhancement doesn’t appear on yRad5113.
(B) Top: At a certain ca concentration, the hDmc1 joint molecule has the highest signal of the product. Bottom: Although the Ca-ATP can suppress the ATP hydrolysis, there is no obvious tendency between the joint molecules formation and Ca-ATP concentration.
The above results indicate that Dmc1 might have the second ca binding site14.
1-4 Motivation of this study
The meiosis process needs the existing of both Rad51 and Dmc1. But the reason why the two recombinases need to coexist in the meiosis is still unknown.
Although many studies have worked on the overall HR reaction to find the difference between Dmc1 and Rad51, the important and essential step, presynaptic filament formation is rarely investigated. To illustrate the roles in the meiosis of the two homologous proteins, comparing the kinetic properties in the presynaptic filament formation which is the essential step in HR is a good starting spot.
The unpublished data from our collaboration lab indicates the ScRad51-ssDNA filament is more stable than ScDmc1’s. Here, we aim to compare the different kinetic properties between these two recombinases in filament formation by single molecule method- tethered particle motion (smTPM). Besides, we would like to confirm the interaction among ScRad51 and ScDmc1 whether can improve the presynaptic filament formation or not.
CHAPTER 2. MATERIALS AND METHODS
2-1 Rad51, Dmc1, Mei5-Sae3 protein purification
All the ScRad51, ScDmc1 and ScMei5-Sae3 are purified by Prof. Chi’s lab. To prevent thawing and refreezing repeatedly, those protein are diluted into lower concentration with T+150 buffer, aliquoted into small packages and stored at -80 oC.
2-2 gap DNA dT200, AC263 synthesis method
The DNA synthesis mainly separate into two part. One is AC263 gap DNA synthesis, second is dT200/165/135/90/35 gap DNA synthesis.
2-2.1 AC263 gap DNA synthesis
1. To synthesize AC263 gap DNA, first, we use polymerase chain reaction (PCR) to produce 637 double-strand DNA (dsDNA) and 349 dsDNA.
2. Owing to the primer design, the 637 and 349 dsDNA have one 5’ OH and 5’ phosphate end. The 5’phosphate end DNA can be degraded by λ exonuclease (NEB), while the 5’
OH end DNA will survive. After treating exonuclease to ds349 and ds637 and deactivating the exonuclease, the 637 and 349 ssDNA can be collected by gel extraction by Qiagen gel extraction kit.
3. Finally, we anneal the purified 637 ssDNA, 349 ssDNA and the 24 nt 5’-biotin labeled oligo with PCR machine, then extracting the gap product by gel extraction as previously mentioned. To prevent thawing the gap DNA repeatedly, DNA is aliquoted into small package and stored at -20 oC.
2-2.2 dT gap synthesis
1. The dT gap DNA synthesis separates into three steps. Using abasic PCR with Taq polymerase (NEB), the polymerase reaction will stop at the abasic site, fill the counter
site with Adenosine (dA) and produce the DNA with a 5’ overhang.
2. Then the 5’ overhang DNA links with the primer having Dt200/165/135/90/35 sequence by T4 DNA ligase to become 3' overhang DNA.
3. Separate the 3' overhang DNA product from reactant by gel extraction with Qiagen gel extraction kit. Annealing the 3' overhang DNA with 5' biotin-DNA oligo under room temperature. To prevent thawing and refreezing the gap DNA repeatedly, DNA is aliquoted into small package and stored at -20 oC.
Figure 2-1 The DNA substrates were used in this study.
Figure 2-2 DNA preparing scheme
(A) The AC264 DNA preparing scheme (B) The DT gap DNA preparing scheme.
2-3 Buffer Recipe
Anti-digoxigenin solution: 3 µg/ml anti-digoxigenin in H2O
T+150 buffer: [Tris-HCl]=25 mM, [KCl]=150 mM, [glycerol]=10%, [IGEPAL]
=0.01%, [beta-mercaptoethanol]=2 mM, [EDTA]=0.5 mM
Wash buffer: [Tris]=30 mM, [MgCl2]=2.5 mM, [KCl]= 50 mM, pH=7.5 , BSA= 0.1%
Assembly buffer: [Tris]=30 mM, [MgCl2]=2.5 mM, [KCl]= 50 mM, pH=7.5, BSA=
0.1%, [ATP]=1 mM, [DTT]= 1 mM
Assembly buffer with Calcium: [Tris]=30 mM, [MgCl2]=2.5 mM, [KCl]= 50 mM, pH=7.5, BSA= 0.1%, [ATP]=4 mM, [DTT]=1 mM, [CaCl2]=1 mM
2-4 Single-molecule Tethered Particle Motion (TPM)
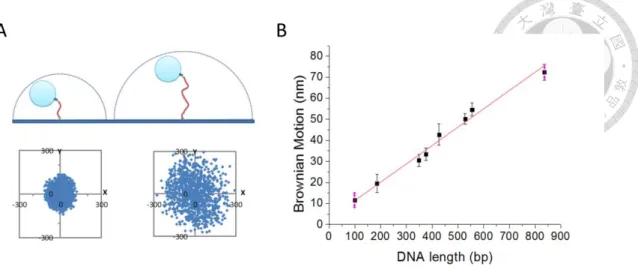
Tethered particle motion (TPM) is one method observing the DNA morphology change with protein-DNA interaction. In basically, one end of the linear DNA can be attached to the slide surface with digoxigenin / anti-digoxigenin interaction. The other end of the linear DNA can be labeled a nanoscale polystyrene bead modified with streptavidin (SA) by the biotin labeling at the end of the DNA. With the optical microscope, the scatter signal of the nanoscale bead can be tracked and analyzed. The value of the mean square displacement coming from Brownian-Motion of the scatter signal is defined as Brownian Motion. We can imagine that the DNA and the tethered bead are a rope and a ball. When the length of the rope increases, the ball has the broader region to explore. The circumstance reflects that the BM of the tethered bead with longer DNA increases in Fig. 2-2(B). Detecting the DNA morphology change in real-time make TPM a simple but powerful tool to investigate the kinetic process between protein and DNA interaction. This technique has been used to observe the protein-DNA interaction of helicase RecBCD translocation16, 17, the recombinase RecA18, and Cre protein19.
Figure 2-3 The Brownian motion value of DNA tethered beads varies linearly with DNA tether lengths.
(A) The tethered bead has broader region to navigate at longer DNA. (B) The relationship between the dsDNA lengths and the Brownian motion values is linear within 1000 base- pairs double-strand DNA. The formula of the regression line is y = 0.0925 x + 2.6478.
With this relationship, 1 nm of Brownian motion corresponds to 10.8 base pairs of dsDNA.
2-4.1 microscope parameters
Table 2-1. Microscope parameters
2-4.2 Reaction slide preparation
The 20 X 40 mm slide is prepared as the following three steps.
1. The slide sinks in the order of 2M KOH, ethanol and distilled deionized water and sonicate for 10 minutes in each solution.
2. The slide is dried with nitrogen first, then each slide can be separated into three reaction channels with parafilm and covered with 20X20 mm slide.
3. The prepared slide is heated with hot plate at 90 oC until the parafilm under cover slide become slightly transparent with the tweezer pressing.
The DNA sample is prepared as the following steps.
1. Introduce 3 µ g/ml anti- digoxigenin solution into the prepared slide for 30 minutes at room temperature.
2. Wash the sink with 100 µl wash buffer and wait for 30 minutes to block the slide surface with BSA.
3. Introduce 1 nM DNA solution into the reaction sink and wait for 30 minutes to fix the DNA on slide surface with digoxigenin / anti-digoxigenin interaction.
4. Remove free DNA substrates with 100 µl wash buffer. Then move the slide to the optical microscope. To trace the DNA fixed on the slide surface, introduce the 1-fold SA bead solution. When the field is filled with tethered bead, wash the free SA bead with the wash buffer.
2-5 Data Analysis
Analyzing the change of the center position of the tethered beads, we can attain the BM value of the tethered bead.
2-5.2 Nucleation time-trace analysis
The nucleation time-course can be separated into 3 periods of times by two state model fitting. The two state fitting is accomplished by Microsoft Excel.
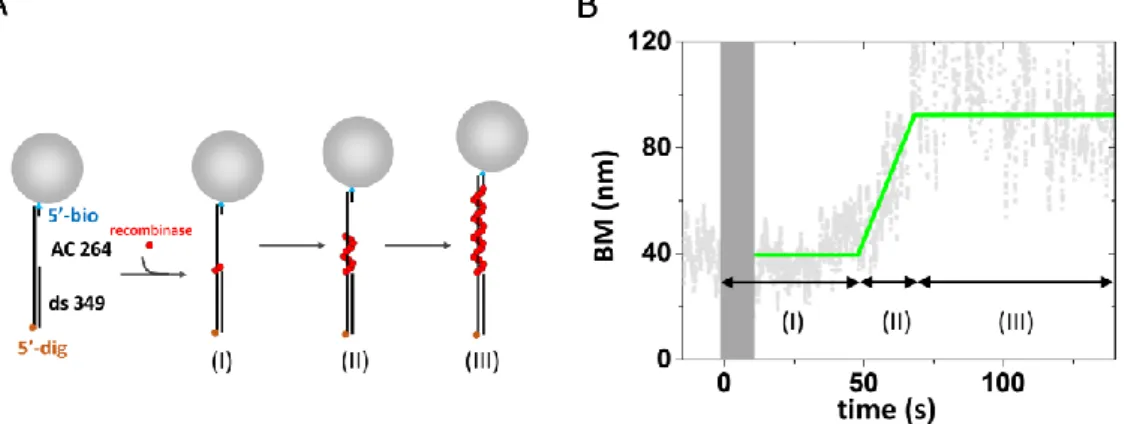
After the fitting process, we can get the typical trace in Fig. 2-3(B). the time-course can reveal three kinetic parameters of the nucleation process. (I) the time after the recombinases solution being introduced into reaction channel and before the BM starting increasing was defined as nucleation time (s). (II) the time between BM value starting to increase and BM value reaching to plateau can be defined as the extension time. The change of BM motion divide by the extension time can offer the filament extension rate (nm/s). (III) the plateau of the filament BM can offer the comparing standard for recombines DNA filament length.
Figure 2-4 recombinase assembly assay using TPM experiments
(A)The scheme represents the recombinase assembly assay.(B) The experimental time- course of the assembly assay, the time region of (I),(II) and (III) is corresponding to the
(I),(II) and (III) in Fig. 2-3(A).
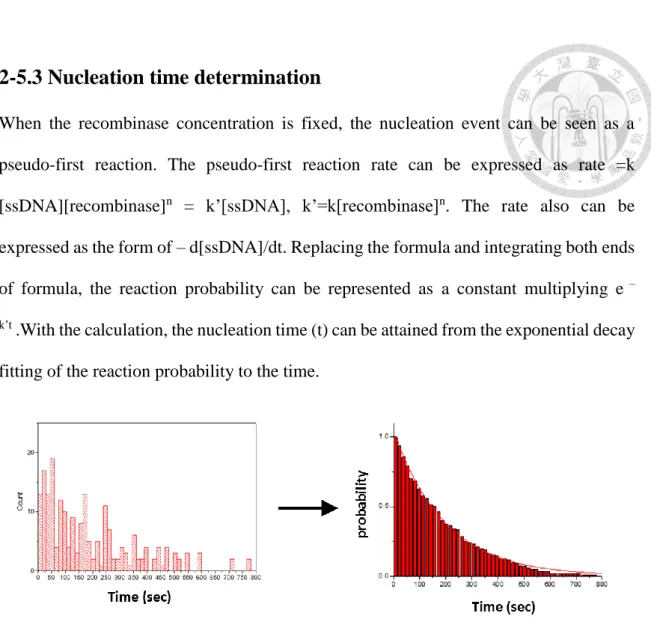
2-5.3 Nucleation time determination
When the recombinase concentration is fixed, the nucleation event can be seen as a pseudo-first reaction. The pseudo-first reaction rate can be expressed as rate =k [ssDNA][recombinase]n = k’[ssDNA], k’=k[recombinase]n. The rate also can be expressed as the form of – d[ssDNA]/dt. Replacing the formula and integrating both ends of formula, the reaction probability can be represented as a constant multiplying e –
k’t .With the calculation, the nucleation time (t) can be attained from the exponential decay fitting of the reaction probability to the time.
Figure 2-5 The nucleation time is determined by the cumulative histogram of time of individual time traces.
The histogram of the nucleation (left figure) is plotted as the cumulative histogram (right figure). Cumulative histograms of nucleation time can be fitted to single exponential decays to return the nucleation time.
2-5.4 TPM snapshots
condensation form, the BM region is separated into 3 regions, condensation (20 nm -35 nm), no-interaction (35 nm-50 nm), and filament formation (50 nm-100 nm).
CHAPTER 3. RESULT
3-1.ScRad51 has fast nucleation rate than Dmc1
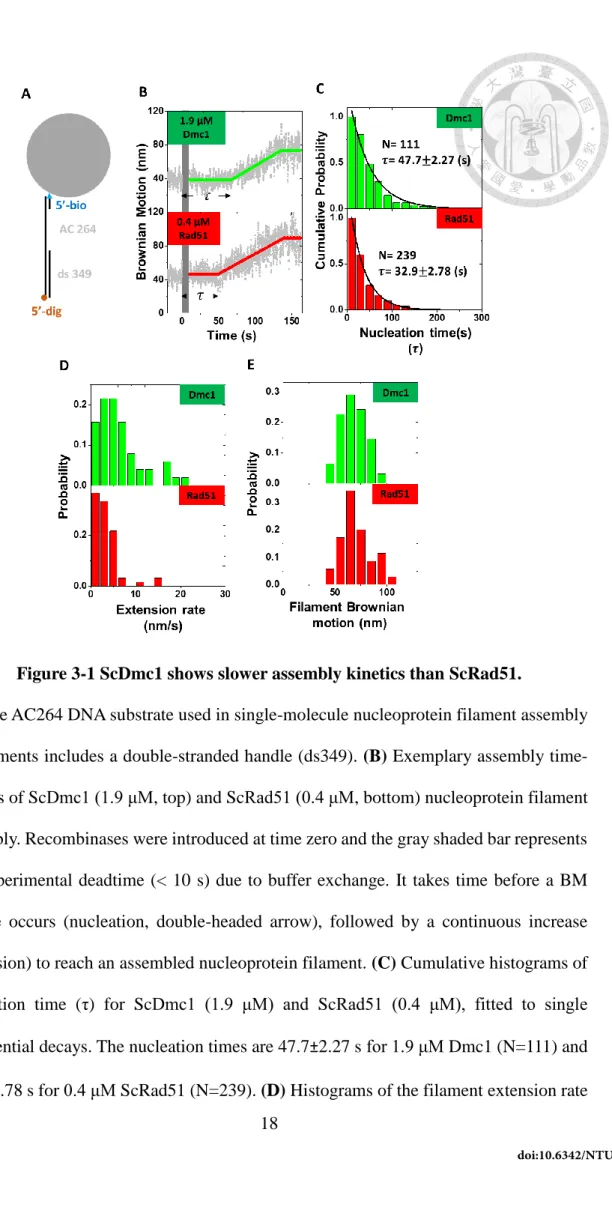
We utilized single-molecule tethered particle motion (smTPM) experiments to monitor the nucleoprotein filament assembly on gap ssDNA substrates in real-time.
During the recombinase nucleoprotein filament assembly process, recombinase binding to DNA leads to the apparent DNA tether length increase due to the lengthening and increased stiffness of DNA substrates2, 20.
For excluding the interruption of DNA secondary structure, a secondary-structure- free, AC-only ssDNA (AC264) was selected to do the assembly assay mentioned in the Method. After analyzing the typical time-course, we get nucleation time, extension rate, and filament lengths for each type of recombinases. Among these three kinetic parameters, cumulative histograms of extension rates and filament BM are similar for ScDmc1 and ScRad51 (Fig. 3-1(D)-(E)). Surprising, as evident in Fig. 3-1(B), the nucleation time of 0.4 μM ScRad51 is faster (32.9s) than that of 1.9 μM ScDmc1 (47.7 s). Considering the nucleation depends on recombinase concentration, lower 0.4 μM ScRad51 nucleates faster than 1.9 μM of ScRad51 suggests that ScRad51 has a higher nucleation rate than ScDmc1. In fact, attempts to use the same ScRad51 and ScDmc1 concentrations on the assembly experiments failed, since the dramatic difference in nucleation time between ScRad51 and ScDmc1 makes experiments unfeasible.
Figure 3-1 ScDmc1 shows slower assembly kinetics than ScRad51.
(A) The AC264 DNA substrate used in single-molecule nucleoprotein filament assembly experiments includes a double-stranded handle (ds349). (B) Exemplary assembly time- courses of ScDmc1 (1.9 μM, top) and ScRad51 (0.4 μM, bottom) nucleoprotein filament assembly. Recombinases were introduced at time zero and the gray shaded bar represents the experimental deadtime (< 10 s) due to buffer exchange. It takes time before a BM change occurs (nucleation, double-headed arrow), followed by a continuous increase (extension) to reach an assembled nucleoprotein filament. (C) Cumulative histograms of nucleation time (τ) for ScDmc1 (1.9 μM) and ScRad51 (0.4 μM), fitted to single exponential decays. The nucleation times are 47.7±2.27 s for 1.9 μM Dmc1 (N=111) and 32.9±2.78 s for 0.4 μM ScRad51 (N=239). (D) Histograms of the filament extension rate
shown in nm/s. (E) Histograms of extended BM value, indicative of the nucleoprotein filament length at the end of the first assembly event. The BM data were collected when the first stable BM was observed, as dictated by the BM plateau after 140 s in (B).
To solve why the ScRad51 have higher nucleation rate than Dmc1, we consider the nucleus formation as a single step reaction, the rate of nucleus formation can be simply expressed as knucleation ×[recombinase]n, where knucleation is assembly rate constant, and n is the fundamental unit of the stable nucleus. Obviously, the nucleation rate difference could come from larger nucleation unit and/or larger assembly constant of ScRad51.
3-2 dsDNA assembly control comparison between Rad51 and Dmc1
To confirm most of the BM increasing events come from filament formation initiating from ssDNA part, we compare the interaction result of recombinase and gap DNA or dsDNA (Fig. 3-2). Control experiments utilizing fully duplex DNA substrates with excess recombinase (Fig. 3-2(C)) return that less than 5% tethers with filament formation over the same experimental time scale (for Dmc1:3.3% no reaction, 51.1%
condense, 4.40% extension, 41% resurge; for ScRad51: 100% no reaction, 0% extension).
However, utilizing gap DNA substrates returns 30-100% extension events in ScRad51 and ScDmc1 individually. (for Dmc1: 0% no reaction, 22.4% condense, 32.6% extension,
increase events, observed here occurred through the initiation on the ssDNA region.
Figure 3-2 Four interaction modes of ScDmc1 and ScRad51.
(A) Four kinds of typical time-courses were observed in smTPM. (B)(C) Upper: The structures represent of the AC263 gap substrate and the double-stranded DNA. Both of them has 5’digoxygenin end and 5’ biotin end to be fixed on the slide and labeled with a tethered bead. Bottom: The histogram shows the ratio to each mode of reaction. The histogram indicates that the extension (type I) events depend on ssDNA existing.
3-3 ScDmc1-DNA interaction is different from ScRad51-DNA.
However, we have found some Dmc1-dependent phenomenon inducing the BM decrease no matter with dsDNA or gap DNA. This BM decrease is never noticed in the ScRad51 filament formation. If the BM decrease ends up with a surging up which leads to that BM value is higher than the pure gap DNA, these events are categorized as resurgences. Without the BM surge, the BM decreasing events are defined as condensation. Because both the condensation and the resurgence are observed with general BSA surface and hydrophobic siloxane surface, these phenomenon is surface independent. Owing to the BM of the tethered bead reflect the DNA length change, the decrease of BM is speculated that the DNA is condensed. Based on previous studies, Dmc1 have filament and ring structure no matter with dsDNA or ssDNA11, 21. Considering that we also observe that Dmc1 has interaction modes differing from presynaptic filament with gap DNA or dsDNA, the BM decrease could result from DNA condense-ring structure observed under electron microscopy8, 11.
Although previous EM data indicated the condense ring structure and filament structure seems to be modulated with the cofactor for ATP or ADP9. However, in our experiment, the four modes of interaction can be recorded under a single field of microscopy. Although the resurgence or condensation in biochemical meaning still unknown, the interaction is existing even in single molecule level. Here we focus on comparing the different properties between ScRad51 and ScDmc1 extension
3-4 The nucleation rates of ScDmc1 and ScRad51 have the similar recombinase-concentration dependence.
To address which one is the dominant factor leading to difference nucleation rate between ScRad51 and Dmc1, we titrate different recombinase concentration and measure the nucleation rate to study how many recombinases (n) are necessary for the nucleation step and nucleation rate constant (k nucleation). Average nucleation times for six different recombinase concentration of ScRad51 and ScDmc1 were determined individually with the same AC263 gap DNA substrate, with each experiments containing more than 50 qualified events. Note that the nucleation times of ScRad51 are much shorter than ScDmc1, so different concentration ranges are used. The fitting of nucleation rate to recombinases concentration shows that the reaction order is 1.46 for ScDmc1 and 1.95 for ScRad51. This result suggests that nucleation on ssDNA required a minimum of two clusters for both ScDmc1 and ScRad51, within our experimental resolution. As previously mentioned, the nucleus formation rate can be seen as pseudo-first order reaction of n recombinase + ssDNA → stable nucleus and expressed as rate = 𝑘𝑎𝑠𝑠𝑒𝑚𝑏𝑙𝑦× [𝑟𝑒𝑐𝑜𝑚𝑏𝑖𝑛𝑎𝑠𝑒]𝑛.The similar nucleation units(n) of these two recombinases indicates that the major difference in nucleation rate mentioned in Fig. 3-1(C) comes from the different level of nucleation rate constant (𝑘𝑛𝑢𝑐𝑙𝑒𝑎𝑡𝑖𝑜𝑛) between ScRad51 and ScDmc1.
Actually, for this AC263 gap substrate, the assembly rate constant for ScRad51 is 0.165 µM-1.95s-1, contrary to that the case of Dmc1 is 0.01 µM-1.46s-1. The ScRad51 nucleation rate constant is quantitatively 10 times greater than Dmc1’s. Obviously, the similar nucleation sizes don’t play a dominant role in the different assembly rate between the two recombinases. On the contrary, the affinity, or the nucleation rate constant could be the factor leading to the kinetic difference.
Figure 3-3 The cumulative histograms of nucleation events indicate the nucleation reaction is dependent on the recombinase concentration.
(A) Nucleation times of Dmc1 nucleoprotein filaments formation measured under various recombinase concentrations using AC263 gap DNA substrates and 1 mM ATP. Take the nucleation reaction as single step reaction, the survival function of nucleation times at
Figure 3-4 ScDmc1 and ScRad51 show the similar reaction order in filament assembly.
Nucleation rates show a similar power-law of recombinase concentration dependence with n=1.46±0.11 for ScDmc1 (error bar is the fitting error), and n=1.95±0.34 for ScRad51 with the AC263 gap DNA substrate.
Table 3-1 Nucleation times for each reaction condition in Fig. 3-4
0 1 2 3
0.02 0.04 0.06 0.08
nuc leation rate (s
-1)
[Recombinase] ()
[ScRad51]
1.95[ScDmc1]
1.45Previous studies have proved the nucleus size for hRad51 is 2.422. Here we demonstrate that in Saccharomyces cerevisiae, the nucleation size of ScRad51 (nRad51=1.95) is similar to that of ScDmc1 (nDmc1=1.46), which indicate both these recombinases form stable nucleus as two cluster conformation. The similar nucleation size between human and yeast recombinases also confirm our results. Based on the DNA substrates used, the significant different in nucleation rate constant between Dmc1 and Rad51 could be attributed to a distinct preference of DNA assembling.
3-5 ScRad51 has higher assembly preference on ssDNA than ScDmc1
To investigate whether ScRad51 and ScDmc1 have different substrate preference during filament assembly process, we determined the assembly kinetics using gap DNA substrates with different ssDNA lengths or with similar DNA length but the different number of DNA gap. These substrates have the same dsDNA handle (349 bp) but different ssDNA dT lengths (349/(dT)n, n= 35, 90, 135, 165, and 200). As we previously showed that ScRad51’s or ScDmc1’s presynaptic filament doesn’t initiate from dsDNA over the same experimental time scale in Fig. 3-2, recombinases could form nucleus at either ss/dsDNA junction or free ssDNA.
The gap DNA substrates contain fixed number of junction site but many ssDNA sites for recombinase assembly to take place. Under the same protein concentration, it is
Figure 3-5 The lengths of the ssDNA dependence in nucleation reaction.
(A) Nucleation times of Dmc1 nucleoprotein filaments formation measured under [ScDmc1]= 1.3 µM using different DNA substrates and 1 mM ATP. Nucleation times at each length of dT gap DNA are single-exponential fits of more than 40 assembled events from at least 5 independent experiments. (B) The similar cumulative histograms shows in the case of [ScDmc1]= 1.9 µM. (C) The similar cumulative histograms shows in the case of [ScRad51]= 0.4 µM.
Table 3-2 The nucleation time to each gap DNA in Fig.3-6
Figure 3-6 ScDmc1 and ScRad51 show the different ssDNA affinity.
The nucleation rates of ScDmc1 and ScRad51 depend on the ssDNA lengths for 349/35,349/90, 349/135, 349/165, 349/200 and 349/(45+45) polydT DNA substrates.
Nucleation rates at each DNA lengths were determined from the nucleation time histograms of more than 75 molecules in the assembly experiments at the 1.9 μM ScDmc1,
Table 3-3 nucleation rate comparison between ScRad51 and ScDmc1.
Divided by concentration factor, the affinity can be quantitatively compared. For each three nucleotides sites, ScRad51’s ssDNA affinity is as 20 times as high as the case of Dmc1’s.
Since nucleation rate also depends on the concentration of recombinase’s nucleation unit, to exclude the concentration effect, we divide the nucleation rate constant over the nth-power of the recombinase concentration, where n is the nucleation unit that we determined in Fig. 3-3. To verify the nucleation size for Dmc1 which is 1.46, we utilizes different concentration of ScDmc1 (1.3 µM and 1.9µM). If the nucleation size determined from Fig. 3-4 is correct, then the value of rate/[recombinase]n will be similar even with different protein concentration. The overlap of the value of rate/[recombinase]n of 1.3 μM and 1.9 μM ScDmc1 with dT90 and dT200 gap DNA make sure the nucleation size determined in Fig. 3-4.
The fitted linear relation between the modified nucleation rate and ssDNA lengths returns a slope and an intercept, where the magnitude of the slope represents the assembly preference on single-strand DNA nucleation site, and the magnitude of intercept represents the affinity to ss/ds junction site. Owing to the previous study shows that the BM increase mainly comes from ssDNA but not dsDNA, we can neglect the contribution of nucleation events on dsDNA here. The apparent flat slope and distinct intercept of
ScDmc1 indicate that Dmc1 nucleation preferentially occurs in ds/ss junction. To ensure that the nucleation of ScDmc1 prefers to occur on DNA junctions, we compare the nucleation time of 349/90 single gap DNA and 349/(45+45) double gap DNA with 1.3 µM ScDmc1(Fig. 3-6). The major difference between 349/(45+45) gap DNA and 349/90 is the number of DNA gap. The result that the nucleation time of dT45+45 gap DNA is about two times of the one of dT90 gap DNA also confirms the junction nucleation preference of ScDmc1. However, in the case of ScRad51, as the dT sequence extends from 35 nucleotides to 200 nucleotides, the modified nucleation rate increases significantly, demonstrating that ScRad51 assembles mainly on ssDNA rather than ds/ss junction site. By comparing the slopes of the nucleation rate at available assembly sites (Table 3-1.), ScRad51 is 20 times faster than ScDmc1. Besides, this again confirms our earlier observation on their nucleation rates (Fig. 3-1) and clarify that the higher ssDNA affinity of ScRad51 leads to the obviously fast nucleation rate in contrast to the case of ScDmc1.
3-6 Calcium stimulates Dmc1’s nucleation
Previous studies showed that human DMC1 forms a much stable nucleoprotein filament in the presence of calcium14. Consistently, Ca2+ stimulates Saccharomyces cerevisiae Dmc1-mediated homologous DNA pairing and strand exchange activity15, 23. Could this calcium stimulation result from the Dmc1 assembly step? To address this
also bind ATP, we increased ATP concentration from 1 mM to 4 mM to make ATP in excess, so the comparison can be made. Increasing ATP concentration from 1 mM to 4 mM without calcium ions does not change the nucleation time of the same ScDmc1 concentration (Fig. 3-5, dark blue). ScDmc1 nucleation time in the presence of calcium ion is shorter (39.7 s), compared to the reaction without calcium (25.7 s). While the extension rate and equilibrium Brownian motion value are similar, calcium ions show apparent stimulation on the nucleation rate of ScDmc1.
In the meiotic cell, it has been shown that the concentration of calcium ion would increase when meiotic cells become matured. Furthermore, calcium ion is also an indicator that if amounts of DNA double strand break occur inside the cell, the concentration of calcium ion would increase. Based on previous knowledge, calcium ion should be a regulator of homologous recombination inside the cell. For hRAD51, calcium can stabilize filament by suppressing the ATP hydrolysis activity, which results in increasing the strand exchange activity13.
While in the case of hDMC1, enhancement from calcium effect doesn’t come only from the ATP hydrolysis attenuation24.Based on the above study, the stimulation mechanism from calcium for RAD51 and DMC1 could be different.
Figure 3-7 Ca2+ stimulates ScDmc1’s nucleation reaction.
Single-molecule experiments of 1.9 μM Dmc1 assembling on the 349/264 DNA substrates in the absence of Ca2+ (top, in dark blue) and in the presence of 1 mM Ca2+
(bottom, in purple). Reactions take place with 4 mM ATP. (A) Exemplary time-courses.
(B) Histograms of nucleation time fitted to single exponential decays, returning 39.7±2.75 s and 25.7±1.44 s for reaction without and with calcium ions. (C)-(D) Histograms of extension rate and filament Brownian motion show no significant difference.
Although the Ca2+ enhancement doesn’t show up in vitro biochemical assay of ScRad5113, it has been confirmed that calcium ion can stimulate the D-loop formation and DNA binding stability of ScDmc1 by ATP hydrolysis inhibition23. In our work, we observe calcium ion can accelerate the assembly step of Dmc1 with ATP. The result supports that the Calcium enhancement of the filament assembly step of ScDmc1.
and Gasior has found the Rad51 mutant can recover the number of Dmc1 foci while no Rad51 foci are observed25, 26. This result not only hints that Rad51 could be an accessory factor of Dmc1, but also indicates that the interaction of Rad51 and Dmc1 exists in solution. Here, based on the unpublished data in collabo lab indicating that ScRad51 has physical interaction with ScDmc1 under 30oC, we use the assembly assay under 30oC to confirm whether the interaction change kinetics in the presynaptic filament formation or not.
Owing to ScRad51 has higher nucleation rate than ScDmc1, we used a low concentration of ScRad51 incubated with ScDmc1. By comparing the 0.6 µM Dmc1 nucleation time (33.1 s) and 0.6 µM Dmc1 plus 0.1 µM Rad51 (34.6 s), the nucleation time is the same. We think that there some reasons can explain the circumstances. One is that the previous in vitro assay utilizes the mutant type Rad51 as Dmc1 cofactor, the other possibility is that the interaction between Rad51 and Dmc1 doesn’t contribute to the assembly process.
Figure 3-8 Nucleation time comparison of ScDmc1 with ScRad51
Under 30oC, the nucleation time of 0.6 µM ScDmc1 is nearly the same as the nucleation time of ScRad51. (A)(C)The timetrace for [ScDmc1]=0.6 µM and [ScDmc1]:[ScRad51]=0.6µM:0.1µM. (B)(D) The nucleation time fitting histogram for for [ScDmc1]=0.6 µM and [ScDmc1]:[ScRad51]=0.6µM:0.1µM.
CHAPTER 4.CONCLUSION AND OUTLOOK
4-1 Conclusion
ScRad51 and ScDmc1 have different kinetic properties on filament formation and different preference of DNA substrates.
With smTPM, we analyze the presynaptic filament formation on ssDNA with the recombinases. The kinetic process of ScRad51 presynaptic filament is much faster than the one of ScDmc1. We have known that ScRad51 is responsible for the DNA damage repair, while Dmc1 mainly focus on creating gene diversity in normal cells. Considering the DSB in mitotic cells are urgent to be renovated, Rad51 must have high fidelity and efficiency to repair the injured DNA. The difference in the nucleation rate between ScRad51 and ScDmc1 is reasonable. However, because the meiosis cell is aimed at producing the gene diversity for evolution, maintaining the gene integrity is not the first priority for Dmc1. Therefore, Dmc1 does not need to proceed the HR as fast and efficient as Rad51. This might explain their difference of nucleation rate and DNA binding affinity.
By matching the recombination titration and the nucleation rate, the reaction order of the nucleation reaction or the nucleus size is determined. Interestingly, both ScRad51 and ScDmc1 share similar nucleus size between mono-cluster and di-cluster.
ScRad51 and ScDmc1 have different preference nucleation sites
The different nucleation rate mainly results from ScRad51’s higher affinity of ssDNA than ScDmc1. However, ScDmc1 shows special affinity on the ds/ss junction.
The junction preference of ScDmc1 have been verified with two sets of experiment. With 1. DNA substrates with different lengths of ssDNA and the same number of DNA gap and 2. DNA substrates with the same DNA lengths but different number of DNA gap, we
confirm the special affinity of ScDmc1 and DNA junctions. The dramatic kinetic difference in nucleoprotein filament assembly might be responsible for the requirement of both Rad51 and Dmc1 in meiotic homologous recombination seen in vivo.
ScDmc1-DNA interaction has different modes comparing with ScRad51-DNA.
In single molecular scale, we find that ScDmc1 has the unique behaviors, condensation, and resurgence, in contrast to the typical filament formation of ScRad51.
Because that the Brownian Motion decrease of ScDmc1 can be differentiated from the stuck bead events and that the phenomena are independent of the surface, the BM decrease indicates the special interaction between ScDmc1 and DNA. Considering that the Dmc1 stack-ring structure observed under the electronic microscope, this condensation in smTPM could be related to the special structure in EM. Besides, because the BM surge after the resurgence is similar to the typical presynaptic filament, the condensation could be an intermediate state of the filament formation.
Ca2+facilitate the assembly of ScDmc1 nucleus.
In the meiotic cell, it has been shown that the concentration of calcium ion would increase when meiotic cells become matured. Furthermore, calcium ion is also an indicator that if a huge amount of DNA double-strand break occur inside the cell, the concentration of calcium ion would increase. Based on previous knowledge, calcium ion should be a regulator of homologous recombination inside the cell. It has been supposed
with ATP. Although the result can’t confirm the second binding site for Ca2+ in ScDmc1, this observation could ensure that modulation from Ca2+ promotes the assembly step of ScDmc1 presynaptic filament formation.
The interaction between Rad51 and Dmc1 doesn’t affect the kinetic process of nucleation.
Although previous works have found Rad51 and Dmc1 have interaction both in vivo10 and in vitro27, 28, we didn’t see significant acceleration on Dmc1’s filament formation with Rad51 even at 30oC, which is the temperature that ScDmc1 has physical interaction with ScRad51(data not shown). One possible reason is that the interaction between these two similar proteins doesn’t work on the filament formation. The other is that Rad51 can be an accessory protein of Dmc1 only when the recombinase activity of Rad51 is suppressed, considering that the enhancement from Rad51 is observed only when the suppressor protein Hed1 or the mutant type Rad51 show up27, 28.
Figure 4-1 The hypothesis scheme represents the recombinase presynaptic filament formation.
When a lot of 3' overhang DNA are exposed in meiosis cells, both ScRad51 and ScDmc1 form stable nuclei on ssDNA in dimer. However, ScRad51 prefers to nucleate on the ssDNA part, while Dmc1 prefer to nucleate on the ds/ss junction. In contrast to that ScRad51 directly construct the presynaptic filaments, Dmc1 and DNA were observed to have a meta-stabled state, the condense-ring, before the presynaptic filament formation.
Ca2+ can accelerate the nucleation step of Dmc1.
4-2 Outlook
Dmc1 and Rad51 are important in the mitotic and meiotic process in Eukaryotic cells.
However, the biological meaning of these two similar protein existence still waited to be investigated. Here, we compare ScRad51 and ScDmc1’s kinetic process in presynaptic
overhang DNA with FRET pair on the ssDNA. When the filament formation on DNA overhang, FRET efficiency will decrease owing to the distance between dye pair increasing from the filament DNA stretching. Titrating the length of ssDNA overhang, when the length of the ssDNA is longer than the smallest nucleation size of recombinases, the probability of FRET decrease will incline. With this experiment, we can make sure the nucleation size of recombinases.
Although in kinetics analysis ScRad51 doesn’t enhance ScDmc1 filament formation, the situation might result from that the interaction might not directly affect the filament formation process. To evaluate the viewpoint, both optical tweezer and fluorescent microscopy are powerful tools. With optical tweezer, the real-time force change along the DNA structure can be attained29. Owing to the persistence length and contour length of DNA can reflect the structure of DNA, the optical tweezer might examine the difference between Rad51 filament and Dmc1 filament or the existence of Rad51-Dmc1 co-filament.
The optical tweezer also can measure the stability of filament with offering different force to the filament. Concerning the physical interaction among proteins might not affect the result of functional assays, the fluorescent microscope is the proper tool to see the protein physical interaction by detecting the location of fluorophore-labeled protein. Observing the co-localization of the fluorescent dye from different kinds of protein and the existing time spot of the dye, the protein interaction can be studied. With the fluorescent microscope, the interaction of ScRad51, ScDmc1 and ScMei5-Sae3 can be investigated.
Reference
1. Sonoda, E.; Sasaki, M. S.; Buerstedde, J. M.; Bezzubova, O.; Shinohara, A.; Ogawa, H.; Takata, M.; Yamaguchi-Iwai, Y.; Takeda, S., Rad51-deficient vertebrate cells accumulate chromosomal breaks prior to cell death. EMBO J 1998, 17 (2), 598-608.
2. Hsu, H. F.; Ngo, K. V.; Chitteni-Pattu, S.; Cox, M. M.; Li, H. W., Investigating Deinococcus radiodurans RecA protein filament formation on dsDNA by a real-time single-molecule approach. Biochemistry 2011, 50 (39), 8270-80.
3. Sung, P.; Klein, H., Mechanism of homologous recombination: mediators and helicases take on regulatory functions. Nat Rev Mol Cell Biol 2006, 7 (10), 739-50.
4. Qi, Z.; Redding, S.; Lee, J. Y.; Gibb, B.; Kwon, Y. H.; Niu, H.; Gaines, W. A.; Sung, P.; Greene, E. C., DNA sequence alignment by microhomology sampling during homologous recombination. Cell 2015, 160 (5), 856-69.
5. Masson, J. Y.; West, S. C., The Rad51 and Dmc1 recombinases: a non-identical twin relationship. Trends Biochem Sci 2001, 26 (2), 131-6.
6. Borgogno, M. V.; Monti, M. R.; Zhao, W.; Sung, P.; Argarana, C. E.; Pezza, R. J., Tolerance of DNA Mismatches in Dmc1 Recombinase-mediated DNA Strand Exchange.
J Biol Chem 2016, 291 (10), 4928-38.
7. Lee, J. Y.; Terakawa, T.; Qi, Z.; Steinfeld, J. B.; Redding, S.; Kwon, Y. H.; Gaines, W. A.; Zhao, W.; Sung, P.; Greene, E. C., Base triplet stepping by the Rad51/RecA family of recombinases. Science 2015, 349 (6251), 977-81.
recombinase Dmc1 promotes ATP-dependent homologous DNA strand exchange. Nature 2004, 429 (6990), 433-437.
10. Brown, M. S.; Grubb, J.; Zhang, A.; Rust, M. J.; Bishop, D. K., small Rad51 and Dmc1 complexes often Co-occupy both ends of a meiotic DNA double strand break.
PLOS Genetics 2016, 11 (12), e1005653.
11. Sauvageau, S.; Stasiak, A. Z.; Banville, I.; Ploquin, M.; Stasiak, A.; Masson, J.-Y., fission yeast Rad51 and Dmc1, two efficient DNA recombinases forming helical nucleoprotein filaments. Mol. Cell. Biol. 2005, 25 (11), 4377-4387.
12. Machaca, K., Ca(2+) signaling, genes and the cell cycle. Cell Calcium 2010, 48 (5), 243-50.
13. Bugreev, D. V.; Mazin, A. V., Ca2+ activates human homologous recombination protein Rad51 by modulating its ATPase activity. Proc. Natl. Acad. Sci. U. S. A. 2004, 101 (27), 9988-93.
14. Bugreev, D. V.; Golub, E. I.; Stasiak, A. Z.; Stasiak, A.; Mazin, A. V., Activation of human meiosis-specific recombinase Dmc1 by Ca2+. J Biol Chem 2005, 280 (29), 26886- 95.
15. Lee, M. H.; Chang, Y. C.; Hong, E. L.; Grubb, J.; Chang, C. S.; Bishop, D. K.; Wang, T. F., Calcium ion promotes yeast Dmc1 activity via formation of long and fine helical filaments with single-stranded DNA. J Biol Chem 2005, 280 (49), 40980-4.
16. Chung, C.; Li, H.-W., Direct observation of RecBCD helicase as single-stranded DNA translocases. J. Am. Chem. Soc. 2013, 135 (24), 8920-8925.
17. Fan, H. F.; Li, H. W., Studying RecBCD helicase translocation along Chi-DNA using tethered particle motion with a stretching force. Biophys J 2009, 96 (5), 1875-83.
18. Fan, H. F.; Cox, M. M.; Li, H. W., Developing single-molecule TPM experiments for direct observation of successful RecA-mediated strand exchange reaction. PLOS ONE
2011, 6 (7), e21359.
19. Fan, H. F.; Cheng, Y. S.; Ma, C. H.; Jayaram, M., Single molecule TPM analysis of the catalytic pentad mutants of Cre and Flp site-specific recombinases: contributions of the pentad residues to the pre-chemical steps of recombination. Nucleic Acids Res. 2015, 43 (6), 3237-55.
20. Piechura, J. R.; Tseng, T. L.; Hsu, H. F.; Byrne, R. T.; Windgassen, T. A.; Chitteni- Pattu, S.; Battista, J. R.; Li, H. W.; Cox, M. M., Biochemical characterization of RecA variants that contribute to extreme resistance to ionizing radiation. DNA Repair (Amst) 2015, 26, 30-43.
21. Yu, X.; Egelman, E. H., Helical filaments of human Dmc1 protein on single-stranded DNA: a cautionary tale. J Mol Biol 2010, 401 (3), 544-51.
22. Hilario, J.; Amitani, I.; Baskin, R. J.; Kowalczykowski, S. C., Direct imaging of human Rad51 nucleoprotein dynamics on individual DNA molecules. Proc. Natl. Acad.
Sci. U. S. A. 2009, 106 (2), 361-368.
23. Busygina, V.; Gaines, W. A.; Xu, Y.; Kwon, Y.; Williams, G. J.; Lin, S. W.; Chang, H. Y.; Chi, P.; Wang, H. W.; Sung, P., Functional attributes of the Saccharomyces cerevisiae meiotic recombinase Dmc1. DNA Repair (Amst) 2013, 12 (9), 707-12.
24. Chang, H. Y.; Liao, C. Y.; Su, G. C.; Lin, S. W.; Wang, H. W.; Chi, P., Functional relationship of ATP hydrolysis, presynaptic filament stability, and homologous DNA pairing activity of the human meiotic recombinase DMC1. J Biol Chem 2015, 290 (32), 19863-73.
recombination complexes. Genes Dev 1998, 12 (14), 2208-21.
27. Busygina, V.; Sehorn, M. G.; Shi, I. Y.; Tsubouchi, H.; Roeder, G. S.; Sung, P., Hed1 regulates Rad51-mediated recombination via a novel mechanism. Genes Dev 2008, 22 (6), 786-95.
28. Cloud, V.; Chan, Y. L.; Grubb, J.; Budke, B.; Bishop, D. K., Dmc1 catalyzes interhomolog joint molecule formation in meiosis with Rad51 and Mei5-Sae3 as accessory factors. Science 2012, 337 (6099), 1222-5.
29. Neuman, K. C.; Nagy, A., Single-molecule force spectroscopy: optical tweezers, magnetic tweezers and atomic force microscopy. Nat. Methods 2008, 5 (6), 491-505.