博士論文
Institute of Oceanography College of Science
National Taiwan University Doctoral Dissertation
亞熱帶水庫系統鹼性磷酸酶活性降尺度研究 Downscaling Alkaline Phosphatase Activity
in a Subtropical Reservoir
曾于芳 Yu-Fang Tseng
指導教授:夏復國 博士 Advisor: Fuh-Kwo Shiah, Ph.D.
中華民國 一百 年 七 月
July, 2011
Downscaling Alkaline Phosphatase Activity in a Subtropical Reservoir
- i -
的論文得以完成。首先由衷感謝指導教授夏復國博士啟蒙了學生在生物海洋學範 疇上的研究熱誠,感謝老師開明的引導態度讓學生培養獨立思考的研究精神;感 謝老師對學生諄諄不倦的教導,當學生感到困惑時,老師總能用最好的方式讓學 生豁然;更要感謝老師在學生徬徨的時刻,給予最有力量的支持與協助;而老師 費心對於學生論文的修訂,學生深表感激。感謝各位辛苦的口試委員,中山大學 李 玉玲教授、海洋大學 蔣國平教授與張正教授、師範大學 陳仲吉教授、台灣大學 謝 志豪教授,謝謝你們為學生論文細心的審閱並提供寶貴的建議。感謝中央研究院 環境變遷中心 高樹基博士研究室的協助。
感謝夏家軍所有曾經一起努力的夥伴與學弟妹們,宗岳學長、家祿學長、佩 蓉、國源、芷彤 (慧雯)、宗翰、香宜、欽洲、昭成、旭仁、其芳、丁元、庭彰、
季謙、靜英、家榕、旺緯、芷嫻、怡雯、谷威、至希、玟欣、Ariani 與 Inah,真 的謝謝你們,這本論文是大家一起努力的成果。感謝曾在海洋研究船上共事的各 位大哥大姐們,勇伯、曹 sir、文慧姐、何文華大哥與林嘉向大哥,還有總是幫我 們很多的盧老闆。感謝曾經一起出海的好夥伴,嘉文、虹君、思穎,謝謝你們讓 每一段航海旅程成了最美麗的回憶。謝謝高中好同學湘怡與琍瑋,總是聆聽當我 需要被聆聽的時候。謝謝大學同學們與研究所同學們的關心,熱情的你們讓小紅 帽一直覺得不孤單。謝謝哈阿貝健隆,帶我體驗研究以外的台客生活。謝謝楠傑 在論文最後階段給予很多很多的協助,相信灰色的天空總掛著一道幸福的彩虹。
最後,衷心感謝一路守護著我的最親愛的家人們,謝謝爸爸媽媽永遠尊重脾 氣倔強的女兒,謝謝哥哥與妹妹對我的包容,謝謝可愛無比的曾小胖。謝謝你們 在我快樂與悲傷的每個時候,陪伴在我身邊和我一起走過。謹將此論文獻給最親 愛最親愛的你們。
- ii -
本論文首先藉由降尺度法 (群聚-族群-細胞-生理層次) 於 2006 至 2009 年進 行亞熱帶翡翠水庫系統浮游生物缺磷指標 - 鹼性磷酸酶活性之系統性分析,以探 討水體中浮游生物之缺磷狀態。野外調查結果顯示,水庫上層水體 (0~20 m) 有
高的鹼性磷酸酶活性 (1.6~95.2 nM h-1) ,且顯示明顯的季節性變化,推測此系統
浮游生物呈現季節性缺磷狀態。多元迴歸分析結果顯示,混合層深度 (即水體磷 酸鹽可利用度指標) 為主要影響水體鹼性磷酸酶活性季節性變動的調控因子。利 用分層過濾探討不同浮游生物階層對水體鹼性磷酸酶活性貢獻度發現,磷酸酶主 要來源為超微型浮游生物。進一步利用螢光標定酵素分析法輔以螢光顯微鏡觀察 發現,細菌為主要鹼性磷酸酶的供應者,推測在亞熱帶磷缺乏系統中,異營性細 菌對於整體磷循環扮演相當重要的角色。光強度佐以營養鹽操控實驗結果顯示,
此系統中超微浮游生物的生長機制受不同環境因子調控,其中細菌生長主要受到 磷酸鹽調控,而超微藍綠藻生長則主要受到光的調控。另外,操控實驗結果亦證 實在磷缺乏系統中,高的光照強度會刺激超微藍綠藻的生長,甚而勝過細菌的生 長,推測光強度為影響超微藍綠藻與細菌競爭磷酸鹽的主要決定因子。此外本論 文首先觀察到夏季強烈颱風事件為影響水體鹼性磷酸酶活性年間變異的主要調控 因子,證實強烈擾動事件 (颱風與強烈降雨) 對水體所造成的物理化學因子變 動,將直接影響水體浮游生物生理缺磷狀態。未來研究將著重於探討亞熱帶磷缺 乏系統中,颱風事件的強度與頻度對水體鹼性磷酸酶活性表現的調控機制,以探 討強烈水體擾動伴隨營養鹽注入對於浮游生物缺磷程度的影響。
關鍵字: 鹼性磷酸酶、磷缺乏、超微型浮游生物、颱風事件、翡翠水庫。
- iii -
(P)-deficient status of different plankton and the role of alkaline phosphatase activity (APA) in subtropical Feitsui Reservoir. Results from field survey showed that bulk APA (1.6~95.2 nM h-1) was widely observed in the epilimnion (0~20 m) with an apparent seasonal variations, suggesting that plankton in the system were subjected to P-deficient seasonally. Mixed layer depth (an index of phosphate availability) is the major factor influencing the variation of bulk APA and specific APA (124~1,253 nmol mg C-1 h-1), based on multiple linear regression analysis. Size-fractionated APA assays showed that picoplankton (size 0.2~3 μm) contributed most of the bulk APA in the system. In addition, single-cell APA detected by enzyme-labeled fluorescence (ELF) assay indicated that heterotrophic bacteria are the major contributors of APA. Thus, we can infer that bacteria play an important role in accelerating P-cycle within P-deficient systems. Light/nutrient manipulation bioassays showed that bacterial growth was directly controlled by phosphate, while picocyanobacterial growth is controlled by light and can out-compete bacteria under P-limited condition with the aid of light.
Further analysis revealed that the strength of summer typhoon is a factor responsible for the inter-annual variability of bulk and specific APA. APA study demonstrated the episodic events (e.g. strong typhoon and extreme precipitation) had significant influence on APA variability in sub-tropical to tropical aquatic ecosystems. Hence, the results herein will allow future studies on monitoring typhoon disturbance (intensity and frequency) as well as the APA of plankton during summer-to-autumn in subtropical systems.
Key words: alkaline phosphatase activity, phosphorus deficiency, picoplankton, typhoon event, Feitsui Reservoir.
- iv -
Figures... 12
Chapter 2 Temporal Variations of Alkaline Phosphatase Activity in a Subtropical Reservoir... 13
2.1 Introduction... 15
2.2 Materials and Methods... 16
2.3 Results... 19
2.4 Discussion... 24
2.5 Conclusion & References... 28
2.6 Tables and Figures... 34
Chapter 3 Temporal Variations of Alkaline Phosphatase Activity in Four Size Fractions in a Subtropical Reservoir... 48
3.1 Introduction... 50
3.2 Materials and Methods... 52
3.3 Results... 54
3.4 Discussion... 56
3.5 Conclusion & References... 58
3.6 Tables and Figures... 62
Chapter 4 A comparison of Alkaline Phosphatase Activity of Osmotrophs by Enzyme-Labeled Fluorescence (ELF) Method... 65
4.1 Introduction... 67
4.2 Materials and Methods... 69
4.3 Results... 71
4.4 Discussion... 73
4.5 Conclusion & References... 75
4.6 Tables and Figures... 80
Chapter 5 Light/Nutrient Effects on the Osmotrophs Behaviors in a Subtropical Reservoir... 84
5.1 Introduction... 86
5.2 Materials and Methods... 88
5.3 Results... 91
5.4 Discussion... 94
5.5 Conclusion & References... 97
5.6 Tables and Figures... 101
Chapter 6 Conclusion... 111
- v -
Table 2.2. Multiple linear regression analysis of APA over other parameters.
Table 2.3. Inter-annual comparison of the parameters during typhoon seasons.
Table 2.4. Linear correlation matrix of parameters collected from typhoon seasons.
Table 2.5. A comparison of bulk APA derived from this and other aquatic ecosystems.
Table 2.6. The level of specific APA as an indicator for P-starvation in studies.
Table 3.1. Multiple linear regression analysis for size-fractionated APA vs. parameters.
Table 3.2. Multiple linear regression analysis for size-fractionated APA vs. bulk APA.
Table 3.3. One-way ANOVA analysis for year-to-year comparison of the parameters.
Table 4.1. In situ ranges of environmental factors and ELF measurements.
Table 4.2. Correlation matrix of the measurements derived from field survey.
Table 5.1. In situ conditions of environmental factors of bioassay 1.
Table 5.2. In situ conditions of parameters and the experiment setup of bioassay 2.
Table 5.3. The increase percentage of picophytoplankton production in bioassay 1.
Table 5.4. The increase percentage of bacterial production in bioassay 1.
Table 5.5. Initial picoplankton APA and percentage of APA inhibition in bioassay 1.
Table 5.6. A list of the turnover rates of picocyanobacteria and bacteria in bioassay 2.
- vi -
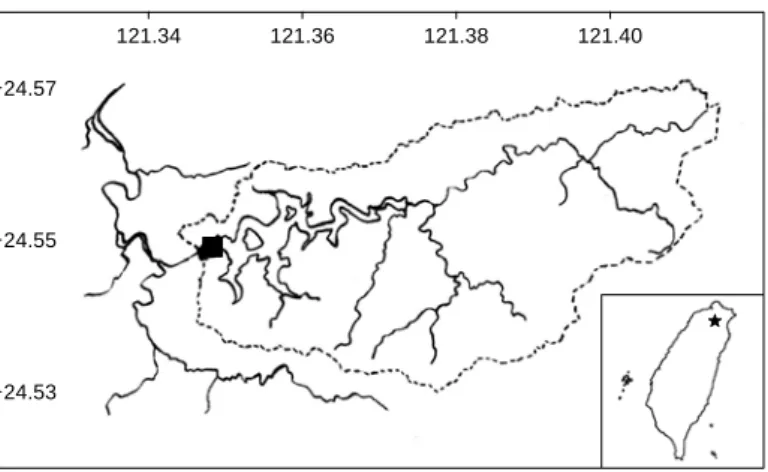
Fig. 2.1. Sampling dam site in the Feitsui Reservoir, north of Taiwan.
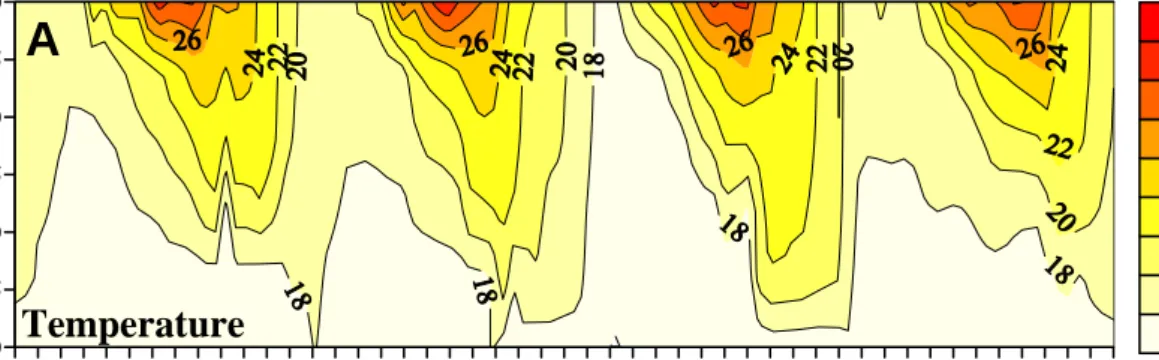
Fig. 2.2. Depth contours of water temperature, SRP, and APA.
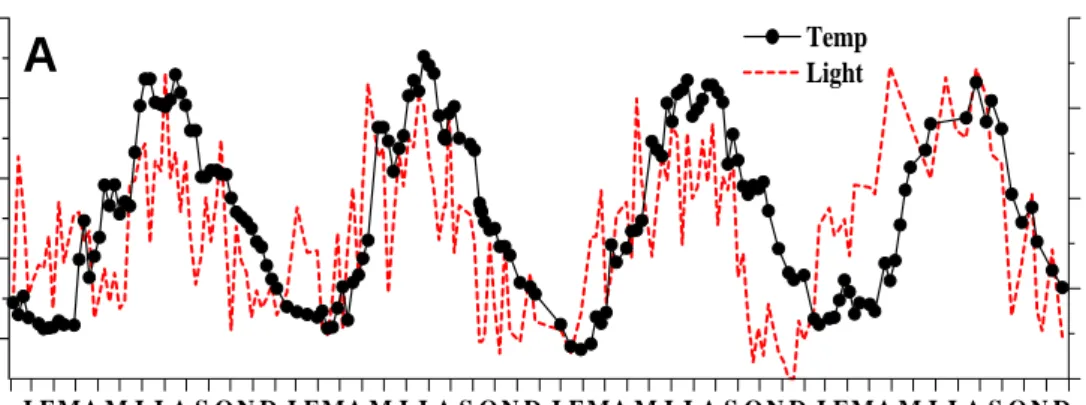
Fig. 2.3. Time series of surface temperature vs. light intensity, mixed layer depth, and daily precipitation vs. typhoon index.
Fig. 2.4. Time series of DIN, SRP, and N/P ratio in epilimnion and hypolimnion.
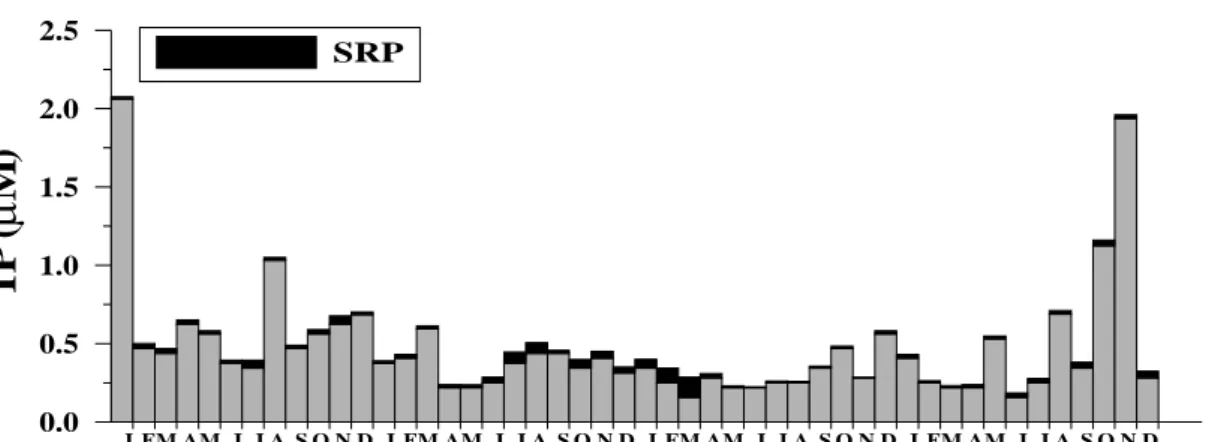
Fig. 2.5. Time series of total phosphorus during 2006~2009.
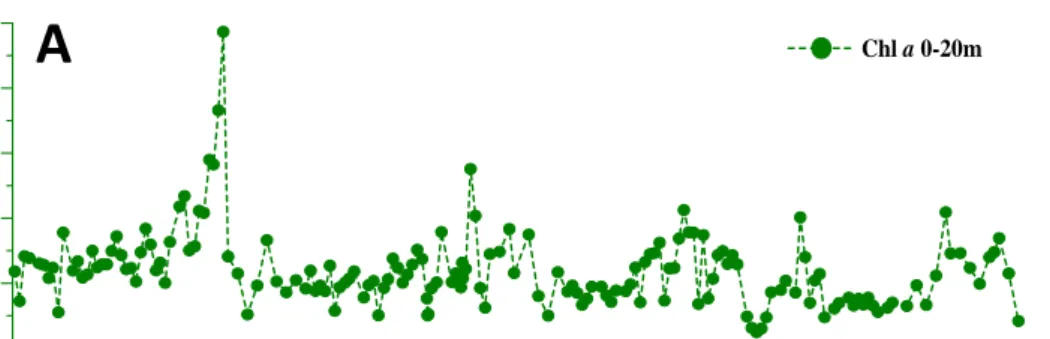
Fig. 2.6. Time series of Chl a, CYA vs.BA, and bulk APA vs. specific APA.
Fig. 2.7. Time series of phytoplankton composition percentage (%).
Fig. 2.8. Scatter plots of Ln transformed bulk APA vs. temperature.
Fig. 3.1. Relative contribution of each size class to the bulk chlorophyll a.
Fig. 3.2. Relative contribution of each size class to the bulk APA.
Fig. 4.1. Scatter plots of picoplankton APA vs. phosphate concentrations and picoplankton APA vs. abundance of ELF-positive picoplankton.
Fig. 4.2. The percentage of ELF-labeled cells to the total cell counts.
Fig. 5.1. The changes of picocyanobacteria abundance, bacteria abundance, and picoplankton APA of Exp. #1 conducted in January 2008.
Fig. 5.2. The changes of picocyanobacteria abundance, bacteria abundance, and picoplankton APA of Exp. #2 conducted in June 2008.
Fig. 5.3. The changes of picocyanobacteria abundance, bacteria abundance, and phosphate concentrations of Exp. #3 conducted in September 2008.
Fig. 5.4. The changes of picocyanobacteria abundance and bacteria abundance of Exp. #4 conducted in October 2008.
Chapter 1
Introduction
Phosphorus (P) is an essential element for all living organisms (for a review, see Karl 2000). A growing amount of research has indicated that P plays an important role in controlling plankton growth and production in aquatic environments, especially in freshwater systems (Schindler 1977, Coveney & Wetzel 1992, Hudson et al. 2000, Guildford et al. 2005). Among various forms of P in water, orthophosphate (i.e.
phosphate) is the preferred form for microbial growth (Cotner & Wetzel 1992). However, ambient phosphate concentrations in freshwater systems are often low and insufficient to satisfy plankton demand (Hudson et al. 2000, Karl 2000). This perceptible lack of phosphate has encouraged research to focus on the role of dissolved organic phosphorus (DOP) playing in the biogeochemical cycles of aquatic ecosystems.
In oligotrophic freshwater systems, DOP concentrations generally exceed phosphate concentrations and comprise a significant proportion of total phosphorus (TP). For instance, Karl & Yanagi (1997) observed that phosphate constituted <25% of the TP in an oligotrophic system, and the remaining 75% occurred in the forms of DOP and inorganic polyphosphate compounds. Consequently, the cycling of DOP could potentially control P-availability, and affect biomass and production of natural plankton communities (Sebastian et al. 2004). This has led to an emphasis on the study of extracellular phosphatase activity due to its important roles in regulating P regeneration from DOP and in increasing the concentration of bio-available P.
Among extracellular phosphatase groups, alkaline phosphatase (APase) is considered as the most important enzyme in controlling DOP degradation (Cotner &
Wetzel 1992, Huang & Hong 1999, Labry et al. 2005, Dyhrman & Ruttenberg 2006, Ivancic et al. 2009). A brief diagram of the ecological importance of APase in aquatic ecosystems is shown in Fig. 1.1. Numerous literatures indicated that when ambient
phosphate is sparse, microorganisms can produce extracellular APase to hydrolyze DOP in compensating for their P-deficiency (Berman 1970, Jansson 1976, Pettersson 1980, Hashimoto et al. 1985, Chrost & Overbeck 1987, Istvanovics et al. 1992, Nausch 1998, Strojsova et al. 2003, Sebastian et al. 2004, Labry et al. 2005, Gao et al. 2006). Further, APase activity (APA) was found repressed when phosphate became available (Perry 1972, Elser & Kimmel 1985, Jamet et al. 1997, Dignum et al. 2004, Labry et al. 2005, Cao et al.
2010). In addition, the performance of APase was inversely correlated to extracellular and intracellular phosphate concentrations (Jansson et al. 1988, Dyhrman & Palenik 1999). Therefore, APA has been used in many aquatic studies as an indicator of P-deficiency of natural plankton populations (Jansson et al. 1988, Istvanovics et al. 1992, Kalinowska 1997, Rose & Axler 1998, Ammerman & Glover 2000, Hoppe 2003, Cao et
al. 2005).
Feitsui Reservoir is an artificially build reservoir with a surface area of 10.24 km2 and a mean depth of 39.6 m. The dam-site is the deepest place with a depth of ~100 m.
As a major source of drinking water for mega Taipei City, Feitsui Reservoir has been less influenced by anthropogenic impact since 1976. According to the long-term records of Carlson's TSI (range from 38 to 46; Chou et al., 2007), Feitsui Reservoir has been considered as a mesotrophic system. However, many investigations have indicated that this system has had high (>200) molar ratios of dissolved inorganic nitrogen to phosphorus (N/P ratios) all year round (Chang & Wen 1997). The unbalanced N/P ratios for microbial requirements (N/P ratio = 16 for phytoplankton and 9 for bacteria) (Redfield 1958, Chrzanowski et al. 1996, Vrede et al. 2002) and low concentrations of phosphate lead to an assumption that P is the limiting element for microbial growth in Feitsui Reservoir.
However, these cannot be used to explain the real nutrient status of plankton in Feitsui Reservoir for the following reasons: Firstly, the analytical determination of phosphate concentrations does not account for the organic form, which could be utilized by living plankton (Karl & Yanagi 1997, Baldwin 1998, Benitez-Nelson 2000). Secondly, low phosphate concentrations do not necessarily represent deficiency under fast regeneration and efficient utilization (Hudson et al. 2000, Karl 2000). APA method is one of direct determinations of the P-status of plankton community since it is a physiological phenomenon induced or repressed by extracellular or intracellular phosphate pools (Chrost & Overbeck 1987). In another word, APA is the method portraying more details about the bio-cycling of DOP in aquatic ecosystems.
Many APA methods have been used (Perry 1972, Hoppe 1983, Scanlan & Wilson 1999, Ammerman & Glover 2000, Sebastian & Niell 2004). Among them, the fluorometric technique (Perry 1972, Gonzalez-Gil et al. 1998) of using artificial soluble organic phosphate substrates (3-0-methylfluorescein phosphate; 3-0-MFP) has been the most popular one.
Bulk APA assay reveals the total amount of enzyme being produced by microorganisms in a sample (Istvanovics et al. 1992, Newman & Reddy 1993, Gao et al.
2006), which provides general information about the P-status at the community level.
However, bulk APA assay is not able to distinguish the activities of APase between particulate and dissolved fractions (Solorzano & Sharp 1980). Moreover, the assay is incapable to differentiate the sources of APA from different plankton, including phytoplankton, bacterioplankton, and zooplankton. Measuring APA by size-fractionation method may allow one to differentiate “loosely” the signals come from particles of different size and dissolved fraction. However, the applicability of this method could be
doubtful for plankton groups (i.e. picophytoplankton and bacterioplankton) that are in the same size range.
To overcome the problems mentioned above, a single-cell Enzyme-Labeled Fluorescence (i.e. ELF) method has been developed to detect the P-status information at the individual taxon level. It involves an addition of a phosphomonoesters substrate (ELF-97 phosphatase substrate; Molecular Probes) to samples; however, instead of releasing soluble products to the medium, fluorescent precipitates only forms at the site where APase hydrolysis occurred, which gives this method a high cell-specific capacity.
The ELF method has been applied widely to the studies of natural plankton communities (Rengefors et al. 2003, Strojsova et al. 2003, Lomas et al. 2004, Ranhofer et al. 2009, Cao et al. 2010), and P-status between classes or even between individual cells of the same species can be determined (Dyhrman & Palenik 1999, Dyhrman et al. 2002).
Based on the introduction mentioned above, the P-status of different plankton and their roles in P-cycling were “systematically” (see the last paragraph) studied in a subtropical reservoir. This dissertation entitled “Downscaling Alkaline Phosphatase Activity in a Subtropical Reservoir” has six chapters in it, and organized as follow:
Chapter 1: Introduction
Chapter 2: Temporal variations of alkaline phosphatase activity in a subtropical reservoir Chapter 3: Temporal variations of alkaline phosphatase activity in four size fractions in a
subtropical reservoir
Chapter 4: A comparison of alkaline phosphatase activity of osmotrophs by Enzyme-Labeled Fluorescence (ELF) Method
Chapter 5: Light/nutrient effects on the osmotrophs behaviors in a subtropical reservoir Chapter 6: Conclusions
The term “downscaling” means to study a phenomenon first at larger or broader scales, then its results are used as a boundary condition for the continuing study at smaller or narrower scales, and so on. The major purposes form Chapters 2 to 5 were in a hierarchy of this logic. The main purposes of Chapter 2 were to define bulk APA behavior in Feitsui Reservoir and to find out the controlling mechanisms for its seasonal and inert-annual variations. Using the finding of bulk APA as a boundary, the main purpose of Chapter 3 was to identify which size fraction of plankton APA determined bulk APA variation in general. And it turned out to be the pico-fraction (0.2~3 μm) which was composed mainly by osmotrophs. The ELF method was adopted in Chapter 4 to make out the relative contribution of osmotrophs to the APA of the pico-fraction. In Chapter 5, a series of light/nutrient manipulation experiments were performed to examine their effects on osmotrophs’ growth.
References
Ammerman JW, Glover WB (2000) Continuous underway measurement of microbial ectoenzyme activities in aquatic ecosystems. Mar. Ecol.-Prog. Ser. 201:1-12.
Baldwin DS (1998) Reactive "organic" phosphorus revisited. Water Res. 32:2265-2270.
Benitez-Nelson CR (2000) The biogeochemical cycling of phosphorus in marine systems.
Earth-Sci. Rev. 51:109-135.
Berman T (1970) Alkaline phosphatase and phosphorus availability in Lake Kinneret Limnol. Oceanogr. 15:663-674.
Cao XY, Song CL, Zhou YY (2010) Limitations of using extracellular alkaline phosphatase activities as a general indicator for describing P deficiency of phytoplankton in Chinese shallow lakes. J. Appl. Phycol. 22:33-41.
Cao XY, Strojsova A, Znachor P, Zapomelova E, Liu GX, Vrba J, Zhou YY (2005) Detection of extracellular phosphatases in natural spring phytoplankton of a shallow eutrophic lake (Donghu, China). European Journal of Phycology 40:251-258.
Chang SP, Wen CG (1997) Changes in water quality in the newly impounded subtropical Feitsui Reservoir, Taiwan. J. Am. Water Resour. Assoc. 33:343-357.
Chou WS, Lee TC, Lin JY, Yu SL (2007) Phosphorus load reduction goals for Feitsui Reservoir watershed, Taiwan. Environ. Monit. Assess. 131:395-408.
Chrost RJ, Overbeck J (1987) Kinetics of alkaline phosphatase activity and phosphorus availability for phytoplankton and bacterioplankton in Lake Plusssee (North-German Eutrophic Lake). Microb. Ecol. 13:229-248.
Chrost RJ, Siuda W (2002) Ecology of microbial enzymes in lake ecosystems. Enzymes in the environment: activity, ecology, and applications, 1st edn. Marcel Dekker, New York.
Chrzanowski TH, Kyle M, Elser JJ, Sterner RW (1996) Element ratios and growth dynamics of bacteria in an oligotrophic Canadian shield lake. Aquat. Microb.
Ecol. 11:119-125.
Cotner JB, Wetzel RG (1992) Uptake of dissolved inorganic and organic phosphorus compounds by phytoplankton and bacterioplankton. Limnol. Oceanogr.
37:232-243.
Coveney MF, Wetzel RG (1992) Effects of nutrients on specific growth rate of bacterioplankton in oligotrophic lake water cultures Appl. Environ. Microbiol.
58:150-156.
Dignum M, Hoogveld HL, Matthijs HCP, Laanbroek HJ, Pel R (2004) Detecting the phosphate status of phytoplankton by enzyme-labelled fluorescence and flow cytometry. Fems Microbiology Ecology 48:29-38.
Dyhrman ST, Palenik B (1999) Phosphate stress in cultures and field populations of the dinoflagellate Prorucentrum minimum detected by a single-cell alkaline phosphatase assay. Appl. Environ. Microbiol. 65:3205-3212.
Dyhrman ST, Ruttenberg KC (2006) Presence and regulation of alkaline phosphatase activity in eukaryotic phytoplankton from the coastal ocean: Implications for dissolved organic phosphorus remineralization. Limnol. Oceanogr.
51:1381-1390.
Dyhrman ST, Webb EA, Anderson DM, Moffett JW, Waterbury JB (2002) Cell-specific detection of phosphorus stress in Trichodesmium from the western north Atlantic.
Limnol. Oceanogr. 47:1832-1836.
Elser JJ, Kimmel BL (1985) Nutrient Availability for Phytoplankton Production in a Multiple-Impoundment Series. Can. J. Fish. Aquat. Sci. 42:1359-1370.
Gao G, Zhu GW, Qin BQ, Chen J, Wang K (2006) Alkaline phosphatase activity and the phosphorus mineralization rate of Lake Taihu. Science in China Series D-Earth Sciences 49:176-185.
Gonzalez-Gil S, Keafer BA, Jovine RVM, Aguilera A, Lu SH, Anderson DM (1998) Detection and quantification of alkaline phosphatase in single cells of phosphorus-starved marine phytoplankton. Mar. Ecol.-Prog. Ser. 164:21-35.
Guildford SJ, Hecky RE, Smith REH, Taylor WD, Charlton MN, Barlow-Busch L, North RL (2005) Phytoplankton nutrient status in Lake Erie in 1997. Journal of Great Lakes Research 31:72-88.
Hashimoto S, Fujiwara K, Fuwa K (1985)
Relationship between alkaline-phosphatase activity and ortho-phosphate in the Present Tokyo Bay
.Journal of Environmental Science and Health Part a-Environmental Science and Engineering & Toxic and Hazardous Substance Control
20:781-809.Hoppe HG (1983) Significance of xxoenzymatic activities in the ecology of Brackish Water - Measurements by means of methylumbelliferyl-Substrates. Mar.
Ecol.-Prog. Ser. 11:299-308.
Hoppe HG (2003) Phosphatase activity in the sea. Hydrobiologia 493:187-200.
Huang BQ, Hong HS (1999) Alkaline phosphatase activity and utilization of dissolved organic phosphorus by algae in subtropical coastal waters. Marine Pollution Bulletin 39:205-211.
Hudson JJ, Taylor WD, Schindler DW (2000) Phosphate concentrations in lakes. Nature 406:54-56.
Istvanovics V, Pettersson K, Pierson D, Bell R (1992) Evaluation of phosphorus deficiency indicators for summer phytoplankton in Lake Erken. Limnol.
Oceanogr. 37:890-900.
Ivancic I, Radic T, Lyons DM, Fuks D, Precali R, Kraus R (2009) Alkaline phosphatase activity in relation to nutrient status in the northern Adriatic Sea. Mar. Ecol.-Prog.
Ser. 378:27-35.
Jamet D, Amblard C, Devaux J (1997) Seasonal changes in alkaline phosphatase activity of bacteria and microalgae in Lake Pavin (Massif Central, France). Hydrobiologia 347:185-195.
Jansson M (1976) Phosphatases in lake water-Characterization of enzymes from phytoplankton and zooplankton by gel-filtration. Science 194:320-321.
Jansson M, Olsson H, Pettersson K (1988) Phosphatases - origin, characteristics and function in lakes. Hydrobiologia 170:157-175.
Kalinowska K (1997) Eutrophication processes in a shallow, macrophyte dominated lake - Alkaline phosphatase activity in Lake Luknajno (Poland). Hydrobiologia 342:395-399.
Karl DM (2000) Aquatic ecology - Phosphorus, the staff of life. Nature 406:31-33.
Karl DM, Yanagi K (1997) Partial characterization of the dissolved organic phosphorus pool in the oligotrophic North Pacific Ocean. Limnol. Oceanogr. 42:1398-1405.
Labry C, Delmas D, Herbland A (2005) Phytoplankton and bacterial alkaline phosphatase activities in relation to phosphate and DOP availability within the Gironde plume waters (Bay of Biscay). Journal of Experimental Marine Biology and Ecology 318:213-225.
Lomas MW, Swain A, Shelton R, Ammerman JW (2004) Taxonomic variability of phosphorus stress in Sargasso Sea phytoplankton. Limnol. Oceanogr.
49:2303-2310.
Nausch M (1998) Alkaline phosphatase activities and the relationship to inorganic phosphate in the Pomeranian Bight (southern Baltic Sea). Aquat. Microb. Ecol.
16:87-94.
Newman S, Reddy KR (1993) Alkaline-phosphatase activity in the sediment-water column of a hypereutrophic lake. Journal of Environmental Quality 22:832-838.
Perry MJ (1972) Alkaline phosphatase activity in subtropical Central North Pacific waters using a sensitive fluorometric method. Marine Biology 15:113-119.
Pettersson K (1980) Alkaline phosphatase activity and algal surplus phosphorus as phosphorus deficiency indicators in lake Erken. Archiv Fur Hydrobiologie 89:54-87.
Ranhofer ML, Lawrenz E, Pinckney JL, Benitez-Nelson CR, Richardson TL (2009) Cell-specific alkaline phosphatase expression by phytoplankton from Winyah Bay, South Carolina, USA. Estuaries and Coasts 32:943-957.
Redfield AC (1958) The biological control of chemical factors in the environment American Scientist 46:205-221.
Rengefors K, Ruttenberg KC, Haupert CL, Taylor C, Howes BL, Anderson DM (2003) Experimental investigation of taxon-specific response of alkaline phosphatase activity in natural freshwater phytoplankton. Limnol. Oceanogr. 48:1167-1175.
Rose C, Axler RP (1998) Uses of alkaline phosphatase activity in evaluating phytoplankton community phosphorus deficiency. Hydrobiologia 361:145-156.
Scanlan DJ, Wilson WH (1999) Application of molecular techniques to addressing the role of P as a key effector in marine ecosystems. Hydrobiologia 401:149-175.
Schindler DW (1977) Evolution of phosphorus limitation in lakes Science 195:260-262.
Sebastian M, Aristegui J, Montero MF, Escanez J, Niell FX (2004) Alkaline phosphatase activity and its relationship to inorganic phosphorus in the transition zone of the North-western African upwelling system. Progress in Oceanography 62:131-150.
Sebastian M, Niell FX (2004) Alkaline phosphatase activity in marine oligotrophic environments: implications of single-substrate addition assays for potential activity estimations. Mar. Ecol.-Prog. Ser. 277:285-290.
Solorzano L, Sharp JH (1980) Determination of total dissolved phosphorus and particulate phosphorus in nature waters. Limnol. Oceanogr. 25:754-757.
Strojsova A, Vrba J, Nedoma N, Komarkova J, Znachor P (2003) Seasonal study of extracellular phosphatase expression in the phytoplankton of a eutrophic reservoir.
European Journal of Phycology 38:295-306.
Vrede K, Heldal M, Norland S, Bratbak G (2002) Elemental composition (C, N, P) and cell volume of exponentially growing and nutrient-limited bacterioplankton. Appl.
Environ. Microbiol. 68:2965-2971.
- 12 -
Fig. 1.1. Brief diagram shows the ecological importance of alkaline phosphatase (APase) in aquatic ecosystems. In phosphorus (P)-limited conditions, plankton (including bacterioplankton, phytoplankton, and zooplankton) could produce extracellular APase to hydrolyze dissolved organic phosphorus (DOP) and get additional phosphate (PO43-
) source in compensating for their P-deficiency. The figure was modified from the book “Enzymes in the Environment: Activity, Ecology, and Applications” (Chrost & Siuda 2002).
Chapter 2
Temporal Variations of Alkaline Phosphatase Activity in a Subtropical Reservoir
Abstract
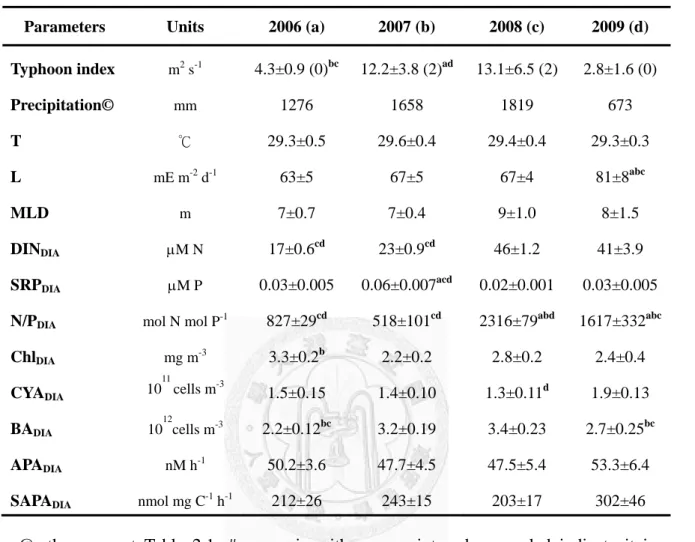
Weekly to bi-weekly samplings of alkaline phosphate activity (APA) as well as related environmental variables were investigated in a deep (100 m depth) subtropical reservoir during the period of 2006~2009. Within the epilimnion (depth <20 m), integrated-averaged bulk APA (1.6~95.2 nM h-1) and biomass normalized specific APA (124~1,253 nmol mgC-1 h-1) varied obviously during the investigation period. Multiple linear regression analysis indicated that in the order of importance, mixed layer depth (MLD, 3~90 m, an index of phosphate availability), picocyanobacteria abundance (0.3~3.7×1011 cells m-3), light intensity (0~102 mE m-2 d-1), and soluble reactive phosphorus concentrations (<0.02~0.15 μM P) were the four major factors that accounted for 65% of the variation of bulk APA. As to specific APA, light intensity, MLD, and temperature (17.5~32.1℃) explained 66% of its variation. Further analysis depicted that the strength of summer typhoon was the factor responsible for the inter-annual variability of bulk and specific APA. The temperature responses of bulk and specific APA in the strong-typhoon-years (2007 & 2008) were significant, while those of the weak-typhoon-years (2006 & 2009) became either lower or insignificant.
This highlighted the importance of episodic events (e.g. strong typhoon and extreme precipitation) in affecting the seasonal cycles of plankton APA in sub-tropical to tropical aquatic ecosystems.
2.1 Introduction
In phosphorus (P)-limited aquatic ecosystems, phosphate is usually in a concentration of nano molar, which is far lower than the detection limit of chemical (spectro-photometry) method (~20 nM; Parsons et al. 1984). Alternatively, extracellular enzyme activity has been adopted as an indicator for the responses of plankton communities to P-deficiency. In aquatic systems, plankton can utilize dissolved organic phosphate (DOP) as an alternative P-source to sustain their growth by producing extracellular alkaline phosphatase (i.e. APase) under P-limited conditions (Chapter 1, Fig. 1.1). Numerous researches have demonstrated that APase activity (i.e. APA) changed proportionally with the status of P-deficiency (Berman 1970, Jones 1972, Jansson 1976, Jansson et al. 1988b, Newman et al. 1994, Cao et al. 2010). This phenomenon has been named as the “induction-repression” mechanism (Jansson et al.
1988a). Accordingly, APA has been suggested as a good indicator for P-status in plankton communities (Healey & Hendzel 1980, Pettersson 1980, Istvanovics et al. 1992, Rose &
Axler 1998, Ammerman & Glover 2000, Kahlert et al. 2002, Cao et al. 2005, Guildford et
al. 2005, Gouvea et al. 2006, Strojsova & Vrba 2009).
The purposes of this study were: (1) to investigate seasonal variations of APA and its ecological relationships (controlling mechanisms) with other environmental factors, and (2) to examine summer typhoon impact on the inter-annual variations of APA, so that the regulation mechanisms of APA at seasonal and inter-annual scales could be explored. The physical measurements included temperature, light intensity, mixed layer depth; the chemical measurements included dissolved inorganic nitrogen and soluble reactive phosphorus concentrations; and the biological measurements included chlorophyll a concentrations and the abundances of picocyanobacteria and heterotrophic bacteria.
2.2 Materials and Methods 2.2.1 Study site and sampling
The study site (Feitsui Reservoir) locates in northern Taiwan (24°55’N, 121°35’E;
Fig. 2.1) with an averaged surface area of 10.24 km2 and a mean depth of 40 m. It is an artificially-build reservoir, and has served as the major source of drinking water for mega Taipei city. This reservoir (and its tributaries) has been well-protected from anthropogenic activities since 1976.
Weekly to biweekly sampling was conducted at the dam-site (depth ~100 m) from Jan 2006 to Dec 2009. The 5-Liter Go-Flo bottles were used for water sampling. The vertical sampling was conducted at 10 depths (0, 2, 5, 10, 15, 20, 30, 50, 70, and 90 m) from the surface to the near bottom manually. Conductivity-temperature-depth (CTD) and the sensors attached onto it were used to record the vertical structures of the measurements, which included temperature, photosynthetic available radiance (PAR), and chlorophyll fluorescence. Water samples stored in 20 L polycarbonate bottles were transported back to the laboratory within 2 hrs for the measurements listed below. Daily hydrographic data (reservoir water levels, water discharges, and precipitation data) were obtained from the web-site (www.feitsui.gov.tw) of Taipei Feitsui Reservoir Administration Bureau. The mixed layer depth (MLD) is defined as the depth at which its temperature is 0.5℃ lower than the surface (Levitus et al. 1982). Typhoon impact index was calculated as the product of daily maximum wind speed and daily maximum precipitation for each typhoon event.
2.2.2 Inorganic Nutrients
Water samples for nutrient analysis were filtered through 500℃ pre-combusted 47-mm GF/F filters under low (<100 mmHg) pressure. The filtrates were used for
nutrients analysis immediately. Nitrate, nitrite, and soluble reactive phosphorus (SRP) concentrations were determined following the methods of Parsons et al (1984) with a spectrophotometer (Shimadzu, UV-1201). Dissolved inorganic nitrogen (DIN) was the sum of nitrate and nitrite. In calculating dissolved N/P ratio, the SRP data below the detection limit (0.02 μM P) were not included. Total phosphorus (TP) concentrations of the surface water were obtained from Taipei Feitsui Reservoir Administration Bureau.
2.2.3 Chlorophyll a (Chl a)
Chl a concentrations were determined by the non-acidification fluorometric procedure of Welschmewer (1994). Water samples were filtered through 47-mm GF/F filters, the filters were extracted with 100% v/v acetone in the dark at -20℃ for 12~16 hrs. Fluorescence was measured using a fluorometer (Turner Designs, TD-700). Algal biomass in carbon (C) unit was determined with a C: Chl a factor of 50 gC gChl a-1 (Antia et al. 1963).
2.2.4 Abundance of picocyanobacteria (CYA) and heterotrophic bacteria (BA)
CYA and BA (<3 μm size fraction) were enumerated by flow cytometry (Partec CyFlow) quipped with a 15 mW, 488 nm argon laser, and the FloMax analysis software.CYA signals were identified by their signatures in a plot of red fluorescence versus orange fluorescence. For BA, samples were pre-diluted 10 times and then stained with SYBR Green (Molecular Probes; final concs., 2.5 μM) for 15 mins. BA signals were identified by their signatures in a plot of side scatter vs. green fluorescence. A solution of yellow-green 1 μm latex beads (~103 beads mL-1; Polysciences) was used as the size indicator. Samples were run at speeds of 800~1200 particles s-1 until ~30000 counts were made. Bacteria biomass in C unit was converted by a conversion factor of 20 fg C cell-1 (Lancelot & Billen 1984).
2.2.5 Alkaline phosphatase activity (APA)
Bulk APA was derived by a fluorometric assay using 3-0-methylfluorescein phosphate (3-0-MFP; Sigma) as the substrate (Perry 1972). Water samples were pre-filtered through a 100 μm nylon sieve to remove large zooplankton. Triplicate 6 mL subsamples were incubated with 750 μL of 3-0-MFP (final concs., 200 nM) in the dark at 25℃ for 1 hr. The fluorescence produced by the 3-0-methylfluorescein (3-0-MF;
excitation, 435 nm; emission, 520 nm) was measured with a fluorometer (Turner Designs, TD-700). Calibration was performed with 3-0-MF standard solutions (Sigma) in the range 20~200 nM. Specific APA (nmol mg C-1 h-1) was derived from the division of APA by the sum of the biomass (in C unit) of Chl a and bacteria. This was based on an assumption that phytoplankton (eukaryotic algae and cyanobacteria) and heterotrophic bacteria constituted the majority of plankton biomass.
2.2.6 Statistical analysis
The depth-integrated averages within epilimnion (upper 20 m) were acquired using trapezoidal method. This is because the signals of APA and many other measurements appeared mostly at a depth <20 m (see the Result section). Statistical analyses including linear correlation analysis, multiple linear regression analysis, one-way ANOVA, and ANCOVA were performed using the statistical software SPSS 12.0TM.
2.3 Results
2.3.1 Physical environment
The depth contour of water temperature (15.9~32.1℃; Fig. 2.2A) revealed that the water column was well-mixed during winter (Dec~Feb of the next year). Stratification occurred during the period of early Apr to late Oct. Surface water temperature (17.5~32.1℃; Fig. 2.3A) varied seasonally with the coldest and warmest temperature recorded in Feb and Aug, respectively. Values of the mixing layer depth (MLD) ranged 3~90 m, with the shallowest and the deepest MLD in summer and winter, respectively (Fig. 2.3B). Weekly light intensity (0~102 mE m-2 d-1; Fig. 2.3A) showed apparent seasonality but seemed varied more than that of surface temperature. Daily precipitation ranged from 0~406 mm (Fig. 2.3C) with lower values recorded in the dry seasons, which covered the period from Nov to Feb of the next year. Higher precipitations came from two sources, the summer evening thunder-showers and the rainfalls induced by typhoon (Fig. 2.3C).
2.3.2 Chemical variables
In the epiliminion, most of the individual SRP (<0.02~0.25 μM P; Fig. 2.2B) and depth-integrated averaged SRP (SRPDIA; <0.02~0.15 μM P; Fig. 2.4B) concentrations were under detection limit (<0.02 μM P) during the stratified seasons except several spikes recorded during post-typhoon periods (Fig. 2.3C). Higher individual SRP and SRPDIA concentrations occurred in the mixing seasons, especially in the winter. The temporal changes of the SRPDIA concentrations in the hypolimnion (20~90 m) were higher than those recorded in the epilimnion (0~20 m) (Fig. 2.4B), but with similar trends (r = +0.60, p<0.01, n=126). Strong inter-annual variation of SRPDIA in both stratified and mixing seasons was noted (Fig. 2.4B). For the stratified seasons, SRPDIA
concentrations in 2007 were very high, even higher than the winter-spring (Dec~May of
the next year) values recorded in 2008. SRPDIA concentrations in the mixing season of 2007 were most undetectable.
During the investigation period, individual dissolved inorganic nitrogen (DIN, = nitrate + nitrite; 14~87 μM N; data not shown) concentrations were always detectable, even in the epilimnion during summer (Fig. 2.4A). In term of stoichiometry, DIN seemed to be much surplus to SRP. The molar ratios of N/P ranged from 230 to 2,995 molN molP-1 with an average of 1,432±685 mol N mol P-1 (Fig. 2.4C), which was about 100-fold greater than that of the Redfield ratio (N/P = 16; Redfield 1958). Total phosphorous (TP, = SRP + DOP) in the surface water ranged from 0.16 to 2.06 μM P with an average of 0.47±0.38 μM P (Fig. 2.5). SRP constituted ca. 8% of the TP.
2.3.3 Biological measurements
Vertical contours of Chl a (data not shown) indicated that phytoplankton biomass was restricted in the upper 20 m. Epilimnic depth-integrated averaged Chl a concentrations (ChlDIA; Fig. 2.6A; range, 0.5~9.7 μg L-1; mean, 2.4±1.2 μg L-1) varied seasonally, and basically followed the trend of temperature (Table 2.1). In this system, the scale of spring bloom was less significant when compared with that of autumn, as evident by the very high ChlDIA (>9 μg L-1) recorded in Oct 2006. Vertical contour of picocyanobacteria (data not shown) indicated that they distributed mostly in the upper 20 m, and the abundance was higher in the surface water and then decreased with depth.
Epilimnic depth-integrated average of picocyanobacteria abundance (CYADIA) ranged from 0.3 to 3.7×1011 cells m-3, with an average of 1.5±0.6×1011 cells m-3 (Fig. 2.6B). The values of CYADIA were positively correlated with ChlDIA (Table 2.1), and higher CYADIA
values were generally recorded in late autumn. CYA were the most abundant species of the phytoplankton community in Feitsui Reservoir. During 2006 and 2007, CYA on
average constituted 87±12% of the total algal cell counts (Fig. 2.7). Heterotrophic bacteria abundance (BA) was high in the upper 20 m, and then dwindled with depth (data not shown). In term of seasonal variation, depth-integrated average of BA (BADIA; Fig.
2.6B; range, 0.8~4.6×1012 cells m-3; mean, 2.3±0.8×1012 cells m-3) changed positively with temperature and CYADIA (Table 2.1). A negative correlation was observed for BADIA vs. MLD.
2.3.4 Seasonal and inter-annual analyses of bulk APA
Signals of bulk alkaline phosphatase activity (APA) were only observed in the upper 20 m, and then decreased significantly with depth (Fig. 2.2C). Epilimnic depth-averaged bulk APA (APADIA; Fig. 2.6C) varied ~100X with a range of 1.6~95.2 nM h-1, and a mean of 40.4±21.5 nM h-1. In the epilimnion, values of the biomass normalized bulk APA (i.e. specific APADIA; Fig. 2.6C) varied ~10X with a range of 124~1,253 nmol mgC-1 h-1, and a mean of 391±207 nmol mgC-1 h-1.
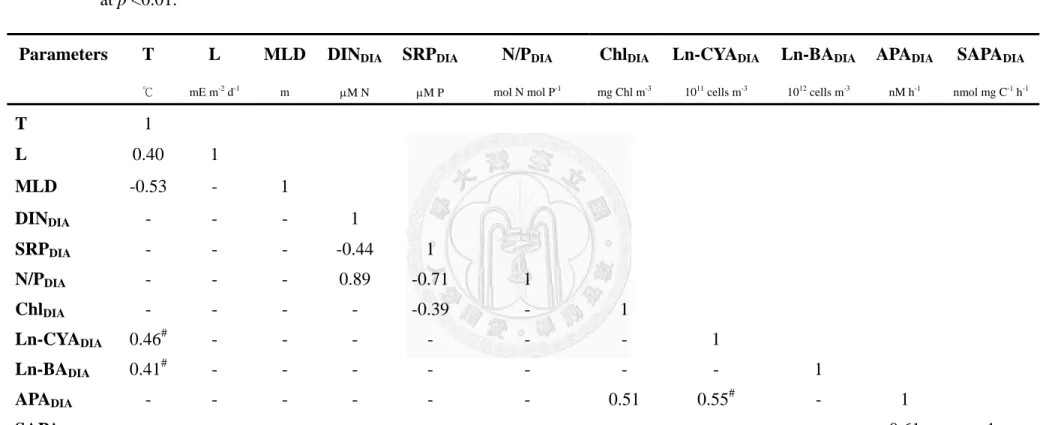
In term of seasonal variation, values of bulk APADIA were positively correlated with of the changes of temperature, light intensity, ChlDIA, CYADIA, and BADIA; values of bulk APADIA were also negatively correlated with MLD, DINDIA, and SRPDIA
concentrations (Table 2.1). A closer examination indicated that the temperature response of bulk APADIA of each year were different (Fig. 2.8). The slope of bulk APADIA vs.
temperature of 2009 was insignificant, while the slopes of the other 3 years were significant with values ranged 0.08~0.14. The slope of 2008 was significantly different from those of 2006 and 2007 (ANCOVA, p<0.05), while the latter two were not different from each other (ANCOVA, p>0.05). The relationships of specific APADIA to other variables were the same as those of bulk APADIA (Table 2.1).
Results of the multiple linear regression analysis indicated that 65% of the bulk
APADIA variability could be explained by the combination of light intensity, MLD, DINDIA, SRPDIA, and CYADIA (Table 2.2). The relative importance (standardized regression coefficient, i.e. Beta weight) of these independent variables on bulk APADIA
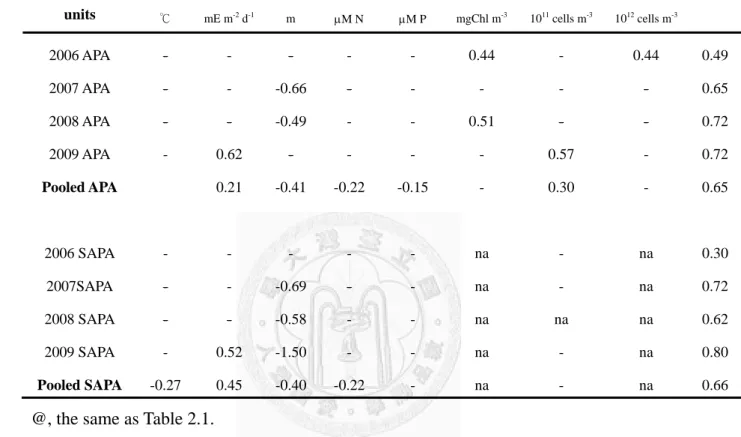
in order was -0.40 for MLD, 0.30 for CYADIA, -0.22 for DINDIA, 0.21 for light intensity, and -0.15 for SRPDIA. The same procedure was performed on the data of each single year. The results showed apparent inter-annual variability of the independent variables in explaining the variation of bulk APADIA. In 2006, ChlDIA and BADIA were the best factors for the changes of bulk APADIA. In 2007, the best combination switched to MLD only. In 2008, MLD and ChlDIA were responsible for 72% of the variation. In 2009, light intensity and CYADIA explained 72% of the variation. Basically the analysis results of specific APADIA were the same as those of bulk APADIA. For the pooled data set, light (Beta = 0.45) and MLD (Beta = -0.41) were the two factors affecting the variability of specific APADIA most.
2.3.5 Typhoon impact
During the investigation period, a total of 18 typhoons had swept through the study site during summer periods (Fig. 2.3C). To test typhoon effects on the behaviors of bulk APADIA and specific APADIA, the data collected during the period of Jul~Sep were analyzed. Table 2.3 indicated that 2007 and 2008 could be categorized as the strong-typhoon years with typhoon impact indices of 12.2±9.2 and 13.1±6 m2 S-1, respectively. On the other hand, typhoon impact indices of 2006 (4.3±2.4 m2 S-1) and 2009 (2.8±3.3 m2 S-1) were ~one-third of those in 2007 and 2008. Accordingly, 2006 and 2009 were considered as the weak-typhoon years. Physical parameters (i.e.
temperature, light intensity, and MLD) showed no difference between strong- and weak-typhoon periods. SRPDIA concentration (0.06±0.03 μM P) in 2007 was ~2-fold higher than those of the other years (p <0.01). CYADIA value (1.3±0.4 ×1011 cells m-3)
recorded in 2008 (one of the strong-typhoon years) was significantly lower than the weakest typhoon year (2009; 1.9±0.3×1011 cells m-3). BADIA values of the strong-typhoon years (3.2~3.4×1012 cells m-3) were higher than those of the weak-typhoon years (2.2~2.7×1012 cells m-3). Bulk APADIA and specific APADIA showed no difference between the strong- and the weak-typhoon years. Correlation analysis of the pooled data set showed that bulk APADIA was positively correlated with ChlDIA and CYADIA, and specific APADIA correlated positively with bulk APADIA (Table 2.4).
2.4 Discussion
In terms of physical structures, the study site is characterized with strong seasonality. The deepening of MLD (Fig. 2.3B) in the cold seasons apparently served as a regular source of inorganic nutrients for plankton growth in the epilimnion. The extremely high N/P ratios (1,432±685 molN molP-1; Fig. 2.4C) recorded in the epilimnion indicted that many plankton (phytoplankton and bacterioplankton) were subjected to P-deficiency, especially during the warm and stratified seasons, as revealed by higher readings of bulk APADIA and specific APADIA (Fig. 2.6C). However, both enzymatic readings fluctuated greatly during the stratified seasons, implying that episodic event (i.e. typhoon) might also affect the SRP concentrations at the dam-site (Fig. 2B), and thus the behaviors of bulk APADIA and specific APADIA (more discussion below).
Bulk APA is an enzymatic reaction. Its expression in the field is subjected to physical, chemical (i.e. substrate availability), and biological regulations. Table 2.1 indicated that the values of bulk APA in this system as a whole could be affected by the changes of physical (temperature and light intensity), chemical (SRP concentrations and availability, i.e. MLD), and biological (Chl a, the abundances of picocyanobacteria and bacteria) parameters. Multiple linear regression analysis (Table 2.2) indicated that MLD (Beta = -0.41) seemed affected the total variations (seasonal and inter-annual) of bulk APADIA more than the biomass of CYA (Beta = +0.30). Overall, it suggested that the changes of phosphate availability (as inferred from MLD) and pico-phytoplankton biomass were the two important factors in determining the seasonal and inter-annual variability of bulk APA in the epilimnion. After biomass normalization, MLD still was one of the essential factors responsible for the variation of specific APADIA.
Based on the negative correlation between phosphate concentrations and APA, many field and enclosure studies have concluded that phosphate supply could be one of the most important factors regulating APA (Chrost & Overbeck 1987, Siuda & Chrost 1987, Istvanovics et al. 1992, Zhou & Zhou 1997, Nausch 1998, Labry et al. 2005). High phosphate concentrations often repressed the synthesis rate of APase (Perry & Eppley 1981, Jamet et al. 1997, Kruskopf & Du Plessis 2004, Labry et al. 2005, Kim et al. 2007, Cao et al. 2010). An “induction-repression” mechanism of phosphate availability on APA has been proposed by Jansson et al (1988a). This study verified the negative relationship of SRPDIA (consider as phosphate) concentration on bulk APADIA and specific APADIA (Table 2.1). However, it is further identified that the “availability” of phosphate that is the changes of the mixed layer depth, is more appropriate and representative than “concentration” itself in explaining the variations of bulk and specific APA (Table 2.2). Several studies suggested that the stoichiometry of inorganic nutrient (i.e. N/P ratios) might affect the expression of APA in the field. For instance, Petterson (1985) found that specific APA in oligotrophic Lake Erken (with dissolved inorganic N/P ratio varied from >1,200:1 in Apr to 8:1 in Sep) increased 10 fold during P-limited period (May~Jun) but decreased to undetectable during N-limited season (Sep). A identical phenomenon was also be found in Chesapeake Bay (Fisher et al.
1992). However, the results of analyses of correlation (Table 2.1) and multiple linear regression analysis (Table 2.2) indicated that this is not the case for the study site.
Potential reason might be that DIN concentrations (14~87 μMN) and N/P ratios (1,432±685 mol N mol P-1) were too high in this system, so that N-limitation could never occurred.
In addition to physical mixing processes and limiting-mineral availability, light intensity, through its effects on autotrophs, could be also important in regulating bulk
and specific APA. In fact, surface light intensity ranked 4th among the five most suitable variables for bulk APA, and ranked 1st among the four most suitable variables for specific APA (Table 2.2). Intuitively, light may enhance autotrophs’ C-fixation rate and results in a higher demand of non-carbon materials (e.g. phosphate) simultaneously. An elevation of APA under higher light intensities eventually would be expected. It is well known that the physiological responses of heterotrophs including bacteria, are light-independent. Light might still affect bacterial APA indirectly because of the mineral-competition between picocyanobacteria and heterotrophic bacteria (i.e.
osmotrophs) in many mineral-limited environments (Thingstad et al. 1993). In another word, it is suspected that light might have an additive (or even multiplicative) effect on either bulk or specific APA. Light effect on osmotrophs’ APA behaviors will be specifically examined in Chapter 5.
Typhoon is a summer-to-autumn episodic event in the northern Hemisphere. On average, more than 20 typhoons were formed in the tropical Pacific Ocean each year, and 6~7 of them passed through Taiwan (data source, Taiwan Central Weather Bureau, www.tcwb.gov.tw). During typhoon events, free phosphate and particle-attached phosphate would be transported from up-stream and the tributaries down to the study site by hyper-pycnal flow formed at the depths of 40~80 m (Chen et al. 2006). The magnitude of the sub-surface SRP maximum (Fig. 2.2B) formed in the mid-waters reflected the strength of typhoon, and served important phosphate source for plankton grown in the epilimnion (Tseng et al. 2010). The strength of summer typhoons and thus phosphate supply could affect the seasonal trend of bulk APA. This was justified by the results of Fig. 2.8 indicating that the temperature responses (i.e. the slopes) of bulk APADIA were quite different among the four sampling years. This implies that the potential impact of episodic events (typhoon and extreme precipitation) can’t be ignored
especially for systems located at typhoon prevailing areas. Recent studies indicated that the intensity (and frequency) of strong typhoon (Chan & Liu 2004, Webster et al. 2005, Wu et al. 2005) and extreme precipitation (Alexander et al. 2006, Kwon et al. 2007) might be enhanced under warming climate. Based on this line of reasoning, many mineral-limiting freshwater ecosystems in sub-tropical to tropical areas might become less P-deficit for plankton growth in summer.
In this system, DOP contributed >90% of total phosphate (TP = SRP +DOP; Fig.
2.5). DOP could serve as an additional source of P for plankton growth. The turn-over times of TP (=TP inventories/APA) estimated to be in the range of 0.2~11.8 d-1, which were within the range (12~24 d-1) reported by Labry et al (2005). The reported values of bulk APA (1~95 nM h-1) of this study were comparable to the oligotrophic ecosystems (Table 2.5), such as the Red Sea (40~150 nM h-1) and the Baltic Sea (40~160 nM h-1).
Since bulk APA is a function of living biomass and specific APA (Bulk APA = biomass x specific APA), specific APA has been recognized as a better indicator for P-deficiency of plankton. In this study, specific APA was derived from the normalization of bulk APA by the biomasses of phytoplankton and bacteria, despite of the fact that all plankton groups or cells respond equally to P-stress (Rengefors et al. 2003, Lomas et al.
2004). Healey & Hendzel (1979), Pettersson (1980, 1985) and Gage & Gorham (1985) defined that a system would be in a status of “critical” and “severe” P-deficiency when the observed specific APA were in the range of 40~250 nmol mgC-1 h-1 and >250 nmol mgC-1 h-1, respectively (Table 2.6). In this system, average values of the specific APA during the cold-mixing seasons were 158±138 nmol mgC-1 h-1, implied that plankton were critically P-deficient. In warm-stratified seasons, plankton were facing severe P-deficiency since the averaged specific APA reached 391±207 nmol mgC-1 h-1.
2.5 Conclusion
The study system was an oligotrophic environment where plankton were subjected to conditions of “critically phosphate-deficiency” and “severely phosphate-deficiency”
during cold-mixing and warm-stratified seasons, respectively. Seasonal variations of bulk APA and biomass normalized APA were mainly controlled by the changes of phosphate availability (i.e. mixed layer depth) and light intensity. Typhoon strength in the summers accounted for the inter-annual variations of bulk and specific APA.
Picocyanobacteria and heterotrophic bacteria were the two most abundant plankton in this system, their relative contributions to bulk APA is one of the important issues to be identified.
References
Alexander LV, Zhang X, Peterson TC, Caesar J, Gleason B, Tank A, Haylock M, Collins D, Trewin B, Rahimzadeh F, Tagipour A, Kumar KR, Revadekar J, Griffiths G, Vincent L, Stephenson DB, Burn J, Aguilar E, Brunet M, Taylor M, New M, Zhai P, Rusticucci M, Vazquez-Aguirre JL (2006) Global observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res.-Atmos.
111:22.
Ammerman JW, Glover WB (2000) Continuous underway measurement of microbial ectoenzyme activities in aquatic ecosystems. Mar. Ecol.-Prog. Ser. 201:1-12.
Antia NJ, McAllister CD, Parsons TR, Stephens K, Strickland JDH (1963) Further measurements of primary production using a large-volume plastic sphere. Limnol.
Oceanogr. 8:166-183.
Berman T (1970) Alkaline phosphatase and phosphorus availability in Lake Kinneret Limnol. Oceanogr. 15:663-674.
Cao XY, Song CL, Zhou YY (2010) Limitations of using extracellular alkaline phosphatase activities as a general indicator for describing P deficiency of phytoplankton in Chinese shallow lakes. J. Appl. Phycol. 22:33-41.
Cao XY, Strojsova A, Znachor P, Zapomelova E, Liu GX, Vrba J, Zhou YY (2005) Detection of extracellular phosphatases in natural spring phytoplankton of a shallow eutrophic lake (Donghu, China). European Journal of Phycology 40:251-258.
Chan JCL, Liu KS (2004) Global warming and western North Pacific typhoon activity from an observational perspective. J. Clim. 17:4590-4602.
Chen YJC, Wu SC, Lee BS, Hung CC (2006) Behavior of storm-induced suspension interflow in subtropical Feitsui Reservoir, Taiwan. Limnol. Oceanogr.
51:1125-1133.
Chrost RJ, Overbeck J (1987) Kinetics of alkaline phosphatase activity and phosphorus availability for phytoplankton and bacterioplankton in Lake Plusssee (North-German Eutrophic Lake). Microb. Ecol. 13:229-248.
Fisher TR, Peele ER, Ammerman JW, Harding LW (1992) Nutrient limitation of phytoplankton in Chesapeake Bay. Mar. Ecol.-Prog. Ser. 82:51-63.
Gage MA, Gorham E (1985) Alkaline phosphatase activity and cellular phosphorus as an index of the phosphorus status of phytoplankton in Minnesota lakes. Freshwater Biology 15:227-233.
Gouvea SP, Melendez C, Carberry MJ, Bullerjahn GS, Wilhelm SW, Langen TA, Twiss MR (2006) Assessment of phosphorus-microbe interactions in Lake Ontario by multiple techniques. J. Gt. Lakes Res. 32:455-470.
Guildford SJ, Hecky RE, Smith REH, Taylor WD, Charlton MN, Barlow-Busch L, North RL (2005) Phytoplankton nutrient status in Lake Erie in 1997. Journal of Great Lakes Research 31:72-88.
Healey FP, Hendzel LL (1979) Fluorometric measurement of alkaline phosphatase activity in algae. Freshwater Biology 9:429-439.
Healey FP, Hendzel LL (1980) Physiological indicators of nutrient deficiency in lake phytoplankton. Can. J. Fish. Aquat. Sci. 37:442-453.
Istvanovics V, Pettersson K, Pierson D, Bell R (1992) Evaluation of phosphorus deficiency indicators for summer phytoplankton in Lake Erken. Limnol.
Oceanogr. 37:890-900.
Jamet D, Amblard C, Devaux J (1997) Seasonal changes in alkaline phosphatase activity of bacteria and microalgae in Lake Pavin (Massif Central, France). Hydrobiologia 347:185-195.
Jansson M (1976) Phosphatases in lake water: characterization of enzymes from phytoplankton and zooplankton by gel-filtration. Science 194:320-321.
Jansson M, Olsson H, Pettersson K (1988) Phosphatase: origin, characteristics, and functions in lakes. Hydrobiologia 170:157-175.
Jones J (1972) Studies on freshwater bacteria: association with algae and alkaline phosphatase activity. Ecol 60:59-75.
Kahlert M, Hasselrot AT, Hillebrand H, Pettersson K (2002) Spatial and temporal variation in the biomass and nutrient status of epilithic algae in Lake Erken, Sweden. Freshwater Biology 47:1191-1215.
Kim C, Nishimura Y, Nagata T (2007) High potential activity of alkaline phosphatase in the benthic nepheloid layer of a large mesotrophic lake: implications for phosphorus regeneration in oxygenated hypolimnion. Aquat. Microb. Ecol.
49:303-311.
Kruskopf MM, Du Plessis S (2004) Induction of both acid and alkaline phosphatase activity in two green-algae (chlorophyceae) in low N and P concentrations.
Hydrobiologia 513:59-70.
Kwon M, Jhun JG, Ha KJ (2007) Decadal change in east Asian summer monsoon circulation in the mid-1990s. Geophys. Res. Lett. 34:6.
Labry C, Delmas D, Herbland A (2005) Phytoplankton and bacterial alkaline phosphatase activities in relation to phosphate and DOP availability within the Gironde plume waters (Bay of Biscay). Journal of Experimental Marine Biology and Ecology 318:213-225.
Lancelot C, Billen G (1984) Activity of heterotrophic bacteria and its coupling to primary production during the spring phytoplankton bloom in the southern bight of the North Sea. Limnol. Oceanogr. 29:721-730.
Levitus S, United States. National O, Atmospheric A (1982) Climatological atlas of the world oceanedn. U.S. Dept. of Commerce, National Oceanic and Atmospheric Administration, Rockville, Md.
Lomas MW, Swain A, Shelton R, Ammerman JW (2004) Taxonomic variability of phosphorus stress in Sargasso Sea phytoplankton. Limnol. Oceanogr.
49:2303-2310.
Nausch M (1998) Alkaline phosphatase activities and the relationship to inorganic phosphate in the Pomeranian Bight (southern Baltic Sea). Aquat. Microb. Ecol.
16:87-94.
Newman S, Aldridge FJ, Phlips EJ, Reddy KR (1994) Assessment of phosphorus availability for natural phytoplankton populations from a hypereutrophic lake.
Archiv Fur Hydrobiologie 130:409-427.
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis., 1st edn. Oxford, New York.
Perry MJ (1972) Alkaline phosphatase activity in subtropical Central North Pacific waters using a sensitive fluorometric method. Marine Biology 15:113-119.
Perry MJ, Eppley RW (1981) Phosphate-uptake by phytoplankton in the Central North Pacific-Ocean. Deep-Sea Research Part a-Oceanographic Research Papers 28:39-49.
Pettersson K (1980) Alkaline phosphatase activity and algal surplus phosphorus as phosphorus deficiency indicators in lake Erken. Archiv Fur Hydrobiologie 89:54-87.
Pettersson K (1985) The availability of phosphorus and the species composition of the spring phytoplankton in Lake Erken. Internationale Revue Der Gesamten Hydrobiologie 70:527-546.
Redfield AC (1958) The biological control of chemical factors in the environment American Scientist 46:205-221.
Rengefors K, Ruttenberg KC, Haupert CL, Taylor C, Howes BL, Anderson DM (2003) Experimental investigation of taxon-specific response of alkaline phosphatase activity in natural freshwater phytoplankton. Limnol. Oceanogr. 48:1167-1175.
Rose C, Axler RP (1998) Uses of alkaline phosphatase activity in evaluating phytoplankton community phosphorus deficiency. Hydrobiologia 361:145-156.
Siuda W, Chrost RJ (1987) The relationship between alkaline phosphatase (APA) activity and phosphate availability for phytoplankton and bacteria in eutrophic lakes. Acta Microbiologica Polonica 36:247-257.
Strojsova A, Vrba J (2009) Short-term variation in extracellular phosphatase activity:
possible limitations for diagnosis of nutrient status in particular algal populations.
Aquatic Ecology 43:19-25.
Thingstad TF, Skjoldal EF, Bohne RA (1993) Phosphorus cycling and algal-bacterial competition in Sandsfjord, Western Norway. Mar. Ecol.-Prog. Ser. 99:239-259.
Tseng YF, Hsu TC, Chen YL, Kao SJ, Wu JT, Lu JC, Lai CC, Kuo HY, Lin CH, Yamamoto Y, Xiao TA, Shiah FK (2010) Typhoon effects on DOC dynamics in a phosphate-limited reservoir. Aquat. Microb. Ecol. 60:247-260.
Webster PJ, Holland GJ, Curry JA, Chang HR (2005) Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 309:1844-1846.
Welschmeyer NA (1994) Fluorometric analysis of chlorophyll-a in the presence of chlorophyll-b and pheopigments. Limnol. Oceanogr. 39:1985-1992.
Wu LG, Wang B, Geng SQ (2005) Growing typhoon influence on east Asia. Geophys.
Res. Lett. 32:4.
Zhou YY, Zhou XY (1997) Seasonal variation in kinetic parameters of alkaline phosphatase activity in a shallow Chinese freshwater lake (Donghu Lake). Water Research 31:1232-1235.
- 34 -
depth-integrated averages (
DIA)
@collected from the study site during the period of 2006~2009. All are significant at p <0.01.
@, T, L, MLD, DIN
DIA, SRP
DIA, N/P
DIA, Chl
DIA, CYA
DIA, BA
DIA, APA
DIA, and SAPA
DIAindicated surface temperature, weekly-averaged light intensity, and epilimnic depth-integrated averages of mixed layer depth, dissolved inorganic nitrogen (nitrate + nitrite), soluble reactive phosphorus, ratio of DIN to SRP, chlorophyll a, picocyanobacteria abundance, bacteria abundance, alkaline phosphatase activity, and specific phosphatase activity, respectively. Ln-, natural-log transformed; #, power function fit. -, zero correlation.
Parameters T L MLD DIN
DIASRP
DIAN/P
DIAChl
DIALn-CYA
DIALn-BA
DIAAPA
DIASAPA
DIAUnits
℃ mE m-2 d-1 m μM N μM P mol N mol P-1 mg Chl m-3 1011 cells m-3 1012 cells m-3 nM h-1 nmol mg C-1 h-1T 1
L 0.36 1
MLD -0.65 -0.44 1
DIN
DIA-0.32 -0.24 0.30 1
SRP
DIA- - - 0.25 1
N/P
DIA- - - 0.44 -0.64 1
Chl
DIA0.23 - - - - - 1
Ln-CYA
DIA- - -0.28
#- - - 0.39
#1
Ln-BA
DIA0.58
#- -0.53
#- - - - 0.41
#1
APA
DIA0.48 0.47 -0.68 -0.42 -0.25 - 0.27 0.47
#0.44
#1
SAPA
DIA0.24 0.61 -0.55 -0.32 -0.22 - na 0.28
#na 0.77 1
- 35 -
Table 2.2. Multiple linear regression analysis of year-to-year and pooled APA and specific APA (SAPA) over other environmental factors@. Numerical indicated the standardized regression coefficient (Beta weight). R2, coefficient of determination. na, not analyzed.
@, the same as Table 2.1.
Year T L MLD DINDIA SRPDIA ChlDIA CYADIA BADIA R2 units ℃ mE m-2 d-1 m μM N μM P mgChl m-3 1011 cells m-3 1012 cells m-3
2006 APA - - - - - 0.44 - 0.44 0.49
2007 APA - - -0.66 - - - - - 0.65
2008 APA - - -0.49 - - 0.51 - - 0.72
2009 APA - 0.62 - - - - 0.57 - 0.72
Pooled APA 0.21 -0.41 -0.22 -0.15 - 0.30 - 0.65
2006 SAPA - - - na - na 0.30
2007SAPA - - -0.69 - - na - na 0.72
2008 SAPA - - -0.58 - - na na na 0.62
2009 SAPA - 0.52 -1.50 - - na - na 0.80
Pooled SAPA -0.27 0.45 -0.40 -0.22 - na - na 0.66