國立臺灣大學生物資源暨農學院農藝學系 碩士論文
Department of Agronomy
College of Bio-Resources and Agriculture
National Taiwan University Master Thesis
蘭嶼姬蝴蝶蘭粗絲期染色體螢光原位雜交與核型分析 Fluorescence in situ hybridization and karyotyping analysis of
Phalaenopsis equestris using pachytene chromosomes
薛豪彥 Hao-Yen Hsueh
指導教授:常玉強 博士、張松彬 博士
Advisor: Yuh-Chyang Charng, Ph.D., Song-Bin Chang, Ph.D.
中華民國 101 年 7 月
July, 2012
誌謝
時間飛快地流逝,那徹夜趕論文、整理數據還有口試的點滴彷彿才在昨日 發生,如今我已即將要畢業了!!回想過去研究所的生活和時光,真的充滿許多 酸甜苦辣。說起來我非常幸運,碩二時跟著指導教授來到成大做研究,雖然為 了上課那學期每個禮拜要臺北、臺南兩邊跑,但碩士班三年中,我同時體驗了 北部與南部兩種生活環境,不僅在台大有知心的麻吉和同學,在成大也有認識 許多朋友和老師,不管在人際關係或學術研究上都受益良多。
很感謝張松彬老師這些年來對我生活上的關心與實驗上的細心指導,每每 在我心情沮喪或實驗失敗時,都能給予我許多建議以及無限的包容,也正因為 您的鼓勵和支持,我才能堅持到今天;也謝謝常玉強老師擔任我的指導老師,
從碩一修習您的功能基因體學課程時,就一直非常欣賞您的教學風格,能夠成 為您指導的學生我始終覺得自己很幸運並且與有榮焉;感謝陳文輝老師與吳文 鑾老師於我論文上的建議和細心斧正,讓我收穫很多,發現更多自己在論文寫 作與實驗上不足之處;我的論文得以順利完成,最重要的要感謝一直作我堅強 後盾的家人,我的爸媽以及哥哥、姊姊對我生活上的關懷與照顧以及提供經濟 來源,讓我得以無後顧之憂可以專心於研究,謝謝您們一路上的支持。很開心 認識台大的同學修巧、子儀、惠心還有鍾萍,我時常想起我們一起修課、吃飯、
一起談天說地分享生活點滴的回憶,還有筱薇、柏蓉和伊婷,妳們始終是我學 習的對象,要特別感謝在實驗上一起共度患難的伊婷,總是在我實驗受挫或生 活不如意時耐心聽我抱怨讓我有談心的對象,還有感謝成大實驗室的恒生學長 於生活上的諸多幫忙,還有兩位可愛的學妹筱琦和宛萱,有妳們和我一起分享、
交流實驗的點滴,常讓我茅塞頓開。還有軒妤、景安、儀慧以及凱翔和惠嵐,
實驗室有妳們總是充滿歡笑,謝謝大家對我的關心。最後要感謝琬婷不離不棄 的關心和照顧,有妳的鼓勵和支持,讓我在實驗上更有信心,這本論文才能順 利的完成,我相信有妳的陪伴,未來的人生道路上我一定會走得更加堅定。
中文摘要
蝴蝶蘭為國內重要的花卉產業,但蝴蝶蘭生長世代較長,遺傳圖譜尚未建 立,細胞遺傳研究也相當缺乏。蘭嶼姬蝴蝶蘭為台灣原生種且具代表性的蘭花,
基因組較小適合作為未來蝴蝶蘭基因組定序計畫的材料。蘭嶼姬蝴蝶蘭體細胞 中期染色體大小型態相似,以傳統染色法難以鑑別。本實驗分析減數分裂十九 對粗絲期染色體,由於收縮程度較低,解析力提升,有助於核型的建構。分析 染色體平均長度大小、染色質的分佈、中心節位置以及有無次級收縮區,可描 述單一染色體的型態。根據核型分析的結果顯示,姬蝴蝶蘭粗絲期染色體平均 長度為 21.95 µm,最大與最小相差 2.35 倍。就染色體型態而論,異染色質皆集 中在中心節兩側,但異染色質的大小與分佈位置在不同的染色體之間有很大的 差異,有些異染色質節清晰,可辨識不同的染色體。姬蝴蝶蘭多屬於中位中節 或次中位中節染色體,只有一對染色體為近端中節,該染色體的短臂末端可觀 察到次級收縮區。另外與有絲分裂中期染色體相較,粗絲期染色體提高 12 倍以 上的解析力,可提升染色體圖譜的應用性。進一步以螢光原位雜交技術將 5S rDNA、45S rDNA、阿拉伯芥型端粒序列與 SOC1 基因定位到染色體上,發現 5S rDNA 與 45S rDNA 分別位於第 17 對及第 13 對染色體的短臂末端,SOC1 位於第 1 對染色體的短臂上,端粒序列則分佈在每對染色體的端點。依據這些 序列在染色體上的分佈位置與訊號的差異,可結合分子標誌建立出更完整的核 型,辨別型態相近的染色體。
有別於蝴蝶蘭早期多以有絲分裂中期染色體為主的染色體研究,本實驗首 先建立了姬蝴蝶蘭粗絲期高解析核型,可成為模式植物供其他蘭花作比較分析,
以了解染色體在蝴蝶蘭屬植物演化過程中核型的演變與排列重組的可能演進。
關鍵詞:細胞遺傳、蘭嶼姬蝴蝶蘭、粗絲期染色體、核型分析、螢光原位雜交
Abstract
Phalaenopsis orchid is the important floral industry in Taiwan. It however has a
long period of life cycle and deficiency of genetic map as well as cytogenetic research. P. equestris is a native and representive orchid in Taiwan, and the kind of its relatively small genome is suitable for analysis in Phalaenopsis genome sequencing project in the future. The size and morphology among somatic metaphase chromosomes were close, and it is difficult to identify each with conventional staining. Ninteen pachytene chromosomes of meiotic stage were studied in this thesis showing their lower contration and high resolution that could contribute to construct a karyotype. Single chromosome morphology can be described by analyzing average length, distribution of chromatin, centromere position and presence of secondary constriction of chromosomes. Based on the results of karyotype, the average length of each pachytene chromosome was 21.95 µm and a 2.35-fold difference between the largest and smallest ones was found.
Regarding each chromosome morphology, all heterochromatin were restricted around centromere, but the amount and distribution had significant differences that could distinguish these chromosomes clearly with some heterochromatin knobs.
Almost all P. equestris chromosomes are metacentric or submetacentric while only one chromosome which includes a secondary constriction on the end of short arm belonging to subtelocentric. In comparison with mitotic metaphase, pachytene chromosomes possessed more than 12-fold higher resolution and thus they can facilitate the construction of chromosome maps. Furthermore, 5S rDNA,45S rDNA, Arabidopsis-type telomeric sequences and SOC1 gene were mapped to
chromosomes using fluorescence in situ hybridization. The results showed that chromosome 17 had a 5S rDNA locus and chromosome 13 had a 45S rDNA locus
on the ends of their short arms. Furthermore, SOC1 was located on the short arm of chromosome 1, and telo meric sequences located on both ends of every chro mo so mes. Based o n t he variat io n in t he spat ial and quant it at ive distribution of DNA sequences, a complete karyotype could be constructed combined wit h mo lecular markers to distinguish similar chro mosomes.
In this study we first developed P. equestris high-resolution karyotype better than previous studies which focus on mitotic metaphase chromosomes, and we hope it will become a model plant used for other orchids. In addition, in the future with the research of chromosome rearrangements of Phalaenopsis species, we may understand how Phalaenopsis chromosomes evolved.
Key words: cytogenetics, P. equestris, pachytene chromosomes, karyotyping, fluorescence in situ hybridization
目錄
口試委員會審定書... ... i
誌謝... ... ii
中文摘要 ... iii
Abstract ... iv
表目錄 ... viii
圖目錄 ... ix
一、前言 ...1
二、前人研究 ...3
(一) 蝴蝶蘭屬植物簡介與分類 ... 3
(二) 姬蝴蝶蘭型態及特徵描述 ... 4
(三) 蝴蝶蘭屬植物細胞遺傳之研究 ... 4
(四) 螢光原位雜交技術的發展用於核型分析 ... 5
(五) 利用核醣體基因與端粒序列探討核型 ... 7
(六) 分子基因庫的開發應用於基因定位與核型分析 ... 9
(七) 以高解析力粗絲期染色體建立核型 ... 11
(八) 姬蝴蝶蘭功能性基因 SOC1 之特性及定位 ... 12
三、材料與方法 ... 14
(一) 材料來源 ... 14
(二) 姬蝴蝶蘭粗絲期染色體的製備 ... 14
1.試驗材料的收集與保存 ... 14
2.花粉細胞粗絲期的鑑定 ... 14
3.以加熱板製備染色體(用於核型分析) ... 15
4.以液態氮製備染色體(用於核型分析與原位雜交反應) ... 15
(三) 螢光原位雜交反應 ... 16
1.質體 DNA 抽取 ... 16
2.探針的標定 ... 17
3.探針濃度之定量 ... 18
4.螢光原位雜交 ... 18
(四) 核型分析與雜交訊號影像處理 ... 20
四、結果 ... 21
(一) 姬蝴蝶蘭粗絲期染色體型態觀察 ... 21
(二) 姬蝴蝶蘭粗絲期染色體核型分析 ... 21
(三) 姬蝴蝶蘭螢光原位雜交與基因定位 ... 24
(四) 姬蝴蝶蘭結合分子標誌高解析核型 ... 25
五、討論 ... 28
(一) 姬蝴蝶蘭粗絲期染色體的製備與型態觀察 ... 28
(二) 姬蝴蝶蘭粗絲期染色體核型分析 ... 28
(三) 姬蝴蝶蘭螢光原位雜交與基因定位 ... 32
(四) 姬蝴蝶蘭粗絲體染色體的辨識與高解析核型 ... 34
(五) 核型統計數據的細微探討 ... 36
(六) 未來研究方向 ... 37
六、參考文獻 ... 38
七、附錄 ... 61
表目錄
表 1:螢光原位雜交反應使用的探針 ... 46
表 2:姬蝴蝶蘭粗絲期染色體核型分析統計數據 ... 47
表 3:姬蝴蝶蘭中期染色體長度統計分析... 48
表 4:姬蝴蝶蘭相異細胞中內含 45S rDNA 訊號的染色體細部分析 ... 49
附表 1:姬蝴蝶蘭粗絲期染色體模式核型圖(第一組)原始數據分析 ... 61
附表 2:姬蝴蝶蘭粗絲期染色體核型圖(第二組)原始數據分析 ... 62
附表 3:姬蝴蝶蘭粗絲期染色體核型圖(第三組)原始數據分析 ... 63
附表 4:姬蝴蝶蘭粗絲期染色體核型圖(第四組)原始數據分析 ... 64
附表 5:姬蝴蝶蘭粗絲期染色體核型圖(第五組)原始數據分析 ... 65
附表 6:五個供試細胞染色體編號比對的結果 ... 66
附表 7:五個供試細胞各染色體平均長度比對的結果 ... 67
附表 8:五個供試細胞異染色質長度比對的結果 ... 68
附表 9:五個供試細胞真染色質長度比對的結果 ... 69
附表 10:五個供試細胞染色體異染色質比率比對的結果... 70
圖目錄
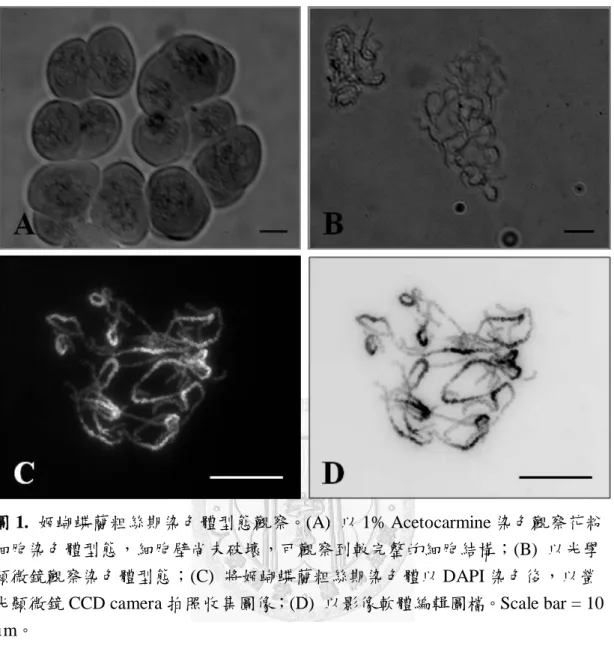
圖 1:姬蝴蝶蘭粗絲期染色體型態觀察 ... 50
圖 2:姬蝴蝶蘭粗絲期染色體模式核型圖(第一組核型) ... 51
圖 3:供作核型分析之細胞染色體圖 ... 52
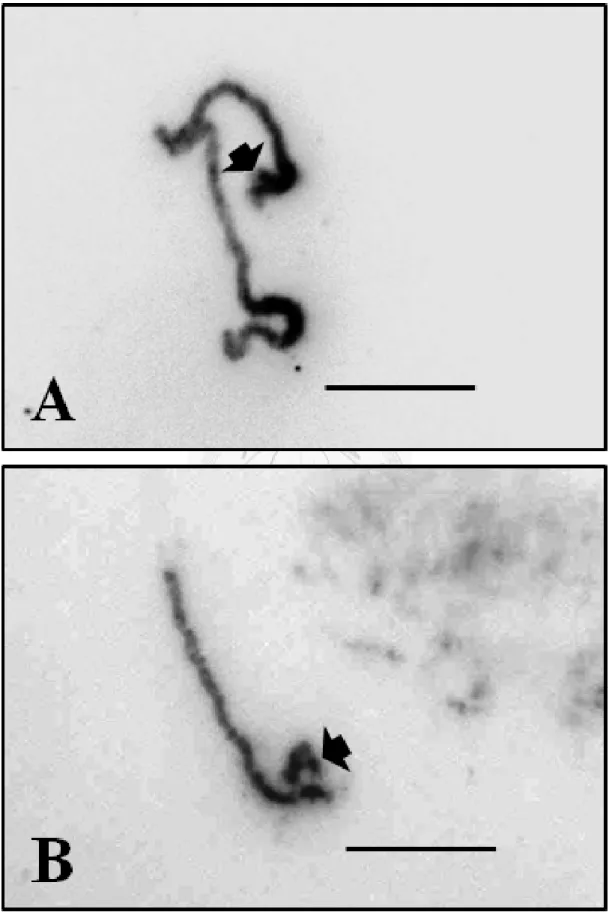
圖 4:觀察姬蝴蝶蘭單離染色體末端次級收縮區 ... 53
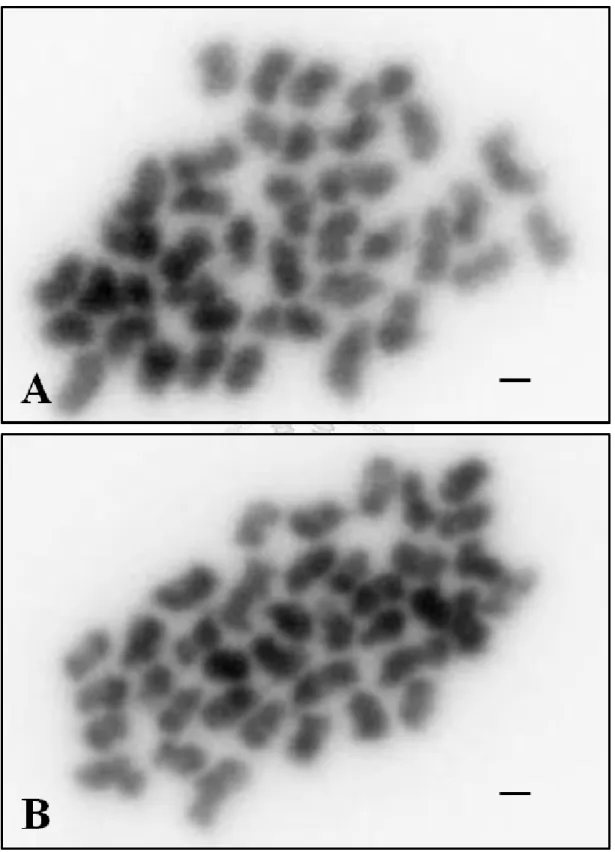
圖 5:觀察姬蝴蝶蘭中期染色體型態 ... 54
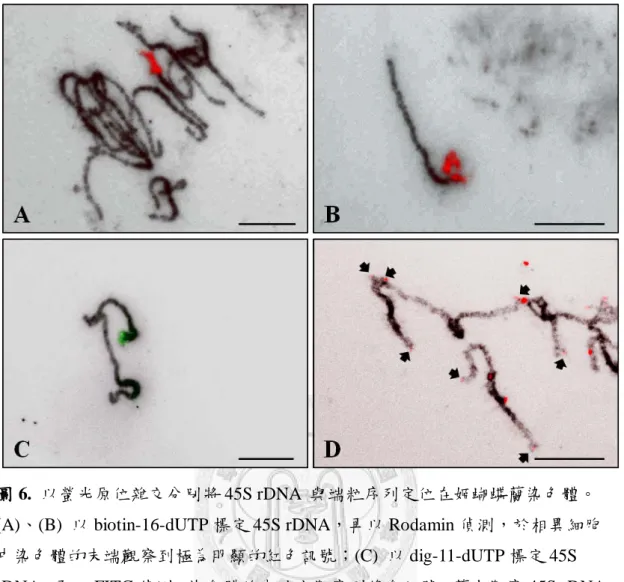
圖 6:以螢光原位雜交分別將 45S rDNA 與端粒序列定位在姬蝴蝶蘭染色體... 55
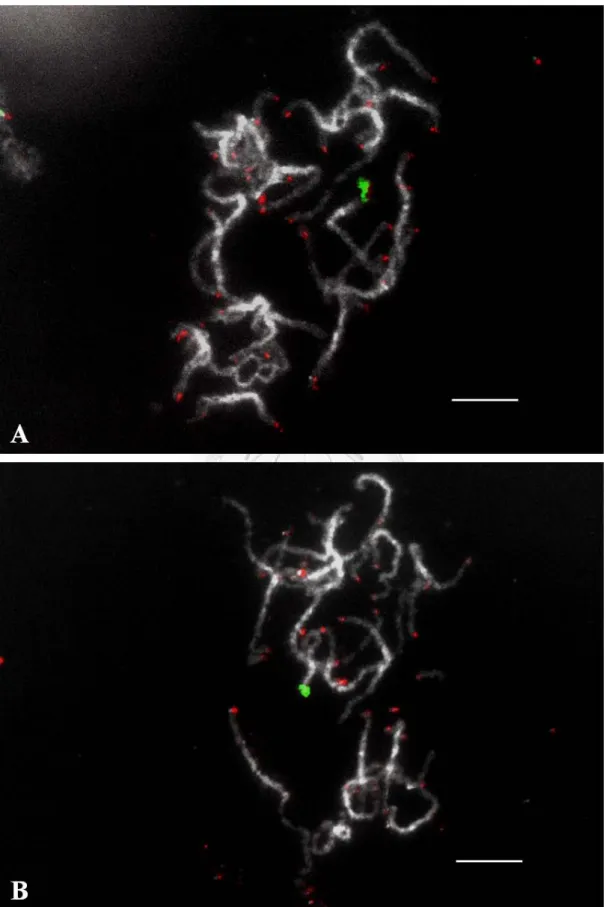
圖 7:以雙標的螢光原位雜交將兩種重複性序列定位在姬蝴蝶蘭粗絲期染色體 .. 56
圖 8:以螢光原位雜交將 5S rDNA 定位在姬蝴蝶蘭粗絲期染色體 ... 57
圖 9:獨特性序列 SOC1 基因利用螢光原位雜交定位在姬蝴蝶蘭粗絲期染色體 .. 58
圖 10:獨特性序列 SOC1 基因定位在姬蝴蝶蘭單離的粗絲期染色體上 ... 59
圖 11:姬蝴蝶蘭核型結構模式圖 ... 60
附圖 1:CMT 基因在染色體上的位置 ... 71
附圖 2:EFS 基因在染色體上的位置 ... 72
一、前言
蘭科植物 (Orchidaceae)分布廣泛,除了沙漠和兩極外無所不在。台灣地處 亞洲太平洋西緣,受太平洋暖流的影響終年四季如春,氣候環境非常適合蘭花的 生長,原產的蘭花約有八十屬四百種,其中蝴蝶蘭具有美麗的花形與豐富的顏色 特別受人喜愛 (陳,2002)。蝴蝶蘭屬 (Phalaenopsis)植物目前發現約有50個原 生種,其中6種可能已絕種,10多種不易尋獲,而台灣為常見的20多種之中其中 兩個種類:「台灣阿嬤蝴蝶蘭」(P. aphrodite subsp. formosana)與「蘭嶼姬蝴蝶 蘭」(P. equestris(Schauer)Rchb. f.)之原產地 (陳,2002)。由於蘭花種原歧異 度高,加上蘭科植物能夠經由人工雜交產生屬間的雜交種 (hybrid),目前蘭花品 種及市面上之蝴蝶蘭栽培種種類及數量繁多 (蔡與莊,2010),為國內最具市場 潛力的花卉產品。台灣於蝴蝶蘭的品種培育、種苗生產與商業栽培上,擁有純熟 的技術與悠久的歷史經驗,加上無菌播種與組織培養在蘭花繁殖上的應用,使得 台灣的蝴蝶蘭產業在國際間占有一席之地,每年有新品種育成 (林等,2008)。
然而過去雖已建立許多育種雜交譜系,常發現親本遺失或不明、子代株數不足,
有些品種雜交幾十個世代,染色體又有自然加倍的現象,現今有許多三倍體及四 倍體的植株,加上蝴蝶蘭分子生物與基因體相關研究缺乏的情況下,至今尚無完 整的遺傳圖譜,商業品種的遺傳背景相當複雜,對於未來分子育種與蝴蝶蘭品種 鑑定的應用上將造成侷限 (蔡與莊,2009)。
台灣不僅在蝴蝶蘭育種與產業上有深厚的基礎,學術研究上也有獨步全球的 成果,其生物科技經產學、官學與學術界共同努力下,已有許多製程與關鍵技術 獲得專利 (林等,2008)。然而蝴蝶蘭植株生長緩慢,需要2至3年方能完成一個 世代,對於建構遺傳圖譜尚須長時間的等待,在等待完整的遺傳圖譜建構完整前,
若能從染色體圖譜著手進行分析,能迅速提供現階段分子育種與蘭花生物技術之 研究和應用。未來在蝴蝶蘭基因組定序計畫中,應以基因組較小的種類為宜,因 此選用染色體較小且具臺灣代表性的蘭嶼姬蝴蝶蘭 (2n = 2x = 38)為材料。蘭嶼
蘭嶼姬蝴蝶蘭屬於小型的染色體,而過去的研究多以有絲分裂 (mitotic)之中期 (metaphase)染色體為主 (Kao et al., 2001),其收縮程度緊實、解析力低, 無法 鑑別單一染色體,不適合作為基因定位與核型分析 (karyotyping)理想的材料。
因此本研究中以減數分裂 (meiotic)粗絲期 (pachytene)染色體為對象建立高解 析核型,並定位染色體專一性DNA分子標誌。若此核型系統能完整建構,可提 供其他蝴蝶蘭或其他蘭科植物,甚至是同為單子葉的水稻作基因組排列 (synteny) 比較分析,並提升染色體圖譜的應用性,了解染色體在演化過程中序列重組現 象。
二、前人研究
(一) 蝴蝶蘭屬植物簡介與分類
蘭花為蘭科植物的統稱,為開花植物中最大、最具多樣性的一科,約有 17,000-35,000 種 (Dressler, 1993),除了南北兩極與沙漠外廣泛分布於世界各地,
尤以亞洲和南美洲熱帶地方最多。蝴蝶蘭於蘭科植物中最具觀賞價值,且為台 灣重要的出口觀賞花卉,屬名 Phalaenopsis 富含希臘文 phalaina (蛾蝶)與 opsis (形象)之含意,形容開花時花姿有如蝶蛾成群飛舞,英文稱之為 Moth Orchid (蛾蘭) (廖,1990),因花型優美、花色典雅與花期較長深受世人喜愛而有「蘭 中之后」的美稱。蝴蝶蘭屬植物約有 45~63 種 (Lin et al., 2005),原產於亞洲 熱帶及亞熱帶區域及太平洋島嶼,主要的分布範圍西起斯里蘭卡 (Sri Lanka)、
南印度 (south India)、喜馬拉雅山麓 (Himalayas)到亞洲東部的巴布亞新幾內 亞 (Papua New Guinea),北起中國南端 (southern China)、台灣 (Taiwan)、菲 律賓 (Philippines)、馬來西亞 (Malaysia)、印尼 (Indonesia)等居中的東南亞 各國延伸到澳洲北部 (northern Australia) (Christenson, 2001., Kao et al., 2007., Lin et al., 2005),台灣南部及蘭嶼可說是蝴蝶蘭天然分佈的北界。
蝴蝶蘭分類上屬於樹蘭亞科 (Epidendroideae)、萬代蘭族 (Tribe Vandeae)、
風蘭亞族 (subtribe Aeridinae)之蝴蝶蘭屬 (Phalaenopsis)植物 (Dressler, 1993)。
蝴蝶蘭分群最早由 Reichenbach (1860) 所提出,當時以唇瓣中裂片有無捲鬚的 特徵將其分為兩群;Bentham (1883) 除上述特徵,再依花瓣的寬度將十五種蝴 蝶蘭分成兩個節 (sections),而 Rolfe 於 1886 年以花瓣的特徵將蝴蝶蘭歸類為 四個節 (Sweet, 1980)。近代較廣為接受的分類系統,為 Sweet (1980) 依花瓣 與萼片的特徵將四十七種蝴蝶蘭分成九個節與四個亞節 (subsections),此系統 能有效分類原生種蝴蝶蘭。後來 Christenson (2001) 再依據花粉塊數目及花型與 葉片的型態擴大種類,將目前發現的六十三種蝴蝶蘭分成五個亞屬 (subgenus)
與八個節。
(二) 姬蝴蝶蘭型態及特徵描述
台灣地處北回歸線之熱帶及亞熱帶地區,氣候溼熱,雨量充足,氣溫條件 適合蘭苗生產,冬季則適合花芽分化。台灣有兩種蝴蝶蘭原生種,其中「蘭嶼 姬蝴蝶蘭(簡稱姬蝴蝶蘭)」最早是由 Schauer 於 1843 年所描述而稱為 Stauropsis equestris;Lindley 於 1848 年稱之為 Phalaenopsis rosea,形容其唇瓣顏色有如
粉紅色玫瑰般的豔麗;直到 1849 年 Reichenbach 正式再改命名為 Phalaenopsis equestris (Sweet, 1980),其種名“equestris”源自拉丁文的 “equester”,表「騎
士」之意,原產於菲律賓和台灣。姬蝴蝶蘭植株大小約 15~30 公分,為多年生 著生性蘭花,葉片大小及型態多變,呈現綠色至不同程度之紫紅色,無特殊斑 紋。花徑約 2~3 公分,花色變化大,有白色至不同程度之粉紅色及紅色,開花 期集中在春夏季。由於姬蝴蝶蘭同時具有紅花及白花種原,且梗上分枝多,每 株花數達數十朵,經常被用來與其他蝴蝶蘭雜交,是迷你多花性蝴蝶蘭育種的 主要親本,在蝴蝶蘭分子演化樹中,與同為 Stauroglottis 節的 P. lindenii 及 P.
celebensis 親緣較接近 (蔡與莊,2010)。姬蝴蝶蘭在型態和花的顏色上差異很 大而有 alba、aurea、rosea、leucaspis 與 leucotante 等不同的變種,受花青素和 類胡蘿蔔素的影響而有不同顏色的唇瓣 (Tang and Chen, 2007)。
(三) 蝴蝶蘭屬植物細胞遺傳之研究
蝴蝶蘭屬植物世代較長,植株生長緩慢,材料取得不易,細胞遺傳研究相 當缺乏。除了原生四倍體 P. buyssoniana 具有 76 條染色體 (2n = 4x = 76)外,所 有原生種蝴蝶蘭為具有 38 條染色體 (2n = 38)之二倍體植株 (Christenson, 2001)。
蝴蝶蘭屬植物雖然具有相同的染色體數目,但屬內種間之染色體大小差異很大。
Shindo 和 Kamemoto (1963) 觀察八種蝴蝶蘭及一種朵麗蘭 (Doritis)的核型,發 現染色體大小相差約 2.8 倍,依區域性分佈及染色體大小將此九種蘭花分成兩
群:一群以 P. amabilis 為首六種蝴蝶蘭主要分布於菲律賓,染色體較小核型較 對稱 (symmetry);另一群包括 P. mannii, P. violacea 及 D. pulcherrima 則分布於 菲律賓以外的地區,染色體較大核型較不對稱 (asymmetry),顯示染色體較大 的類群核型演化程度較高。Arends (1970) 進一步觀察這兩群蝴蝶蘭雜交種在第 一次減數分裂 (metaphaseI)時染色體配對情形,發現蝴蝶蘭染色體群內的同源 性 (homology)較高,而群間同源性較低,提供相異類群間蝴蝶蘭序列演化的證 明。隨著細胞定量分析的發展,Lin 等人 (2001) 利用細胞流測 (flow cytometry) 技術分析十八種原生種蝴蝶蘭及一種朵麗蘭基因組大小,發現 DNA 含量最小 的 P. sanderiana (2.74 pg/2C)與 DNA 含量最大的 P. parishii (16.61 pg/2C),兩者 相差 6.07 倍,其中 P. equestris 的 DNA 含量約為 3.37 pg/2C,於十八種蝴蝶蘭 中 DNA 含量僅高於 P. aphrodite、P. sanderiana 及 P. stuartiana。Kao 等人 (2001) 以九種原生種蝴蝶蘭與一種朵麗蘭為材料,使用 DNA-Feulgen 染色法觀察細胞 核及比較種間染色體核型之差異,發現基因組中異染色質含量與細胞核 DNA 含量 (DNA content)、總染色體體積 (total chromosome volume)及染色體大小呈 正相關,並且對照 P. aphrodite、P. venosa、P. mannii、P. violacea 及 D. pulcherrima 等五種蘭花核型,觀察到染色體較大之蝴蝶蘭含有較多的異染色質,反觀染色 體較小的蝴蝶蘭異染色質較少,而認為異染色質的累積是造成蝴蝶蘭種間核型 差異的重要因子。異染色質大多由重複性序列 (repetitive sequence)所組成,推 測蝴蝶蘭基因組的差異與重複性序列之多寡有關。為進一步探討蝴蝶蘭基因組 組成與親緣關係,Lin 等人 (2005) 分析七種原生種蝴蝶蘭雜交種,以基因組原 位雜交 (genomic in situ hybridization)技術檢視染色體上雜交訊號,發現具有較 大基因組的原生種蝴蝶蘭,無論在種類與量上均含有較多的重複性序列,然而 基因組較小的原生種亦含有本身獨特的序列。這些研究結果顯示異染色質與重 複序列,是影響蝴蝶蘭染色體核型變異與基因組演化重要的關鍵。
(四) 螢光原位雜交技術的發展用於核型分析
原位雜交技術 (in situ hybridization,ISH)是結合了分子生物學 (molecular biology)、免疫細胞化學 (immunocytochemistry)及細胞遺傳學 (cytogenetics)、
顯微鏡與電腦影像分析的技術 (Schwarzacher and Heslop-Harrison, 2000)。早 期原位雜交技術使用的探針為放射性標定,但放射線具危險性,且此法很難同 時滿足靈敏度與解析度這兩者原位雜交成功必需的條件。由於分子生物學的快 速發展,螢光原位雜交 (fluorescence in situ hybridization; 簡稱 FISH)為原有的 放射性原位雜交基礎上發展的新技術,利用螢光標定的外源核酸片段,與細胞 或染色體上待測 DNA 或 RNA 互補配對,依據染色體上雜交訊號的位點可提供 探針序列的定位信息。此技術的優點在於螢光試劑安全無放射線安全之疑慮、
探針穩定性高、實驗週期短且定位準確,能快速獲得結果,還可運用不同顏色 螢光素標記 DNA 探針,同時觀測相異的 DNA 雜交訊號,可定位重要的基因序 列 (Devi et al., 2005; Trask, 1991; Volpi and Bridger, 2008)。
核型分析是對生物細胞核內所有染色體型態及特徵進行分析,為區別物種 之基本遺傳學依據 (闫等,2008);一般可以依據染色體數目、長度、中心節 (centromere)位置或異染色質的分布,或以特定染色法如 Giemsa(G-banding)、
C-banding、R-banding 或觀察端粒 (telomere)之 T-banding 等條帶技術 (banding techniques)鑑定核型 (Miller et al., 2009)。例如應用 C-banding 技術觀察染色體 型態及異染色質的分布,在高粱 (Sorghum bicolor) (Yu et al., 1991)、西番蓮 屬 (Passiflora) (Viana and Souza, 2010)等植物都有相關報導。然許多物種染色 體大小型態相似,以傳統染色法無法加以鑑別;Hoshi 等人 (1998) 以 C-banding 染色觀察黃瓜 (Cucumis sativus )七對染色體,其中有五對染色體呈現明顯的條 帶,但無法觀察到次級收縮區 (secondary constriction)。現今 FISH 技術運用特 定的重複性序列片段作為探針,廣泛運用在核型分析;單一的 DNA 序列 (single copy sequence)片段可辨識特定染色體,而重複序列普遍存在於基因組中,其功 能至今尚未完全明瞭。許多研究顯示高等植物的基因組,重複性序列佔很大之
比例,片段長度從 2 bp 到 10,000 bp 以上,拷貝數 (copy numbers)更從幾百到 數百萬次 (Kubis et al., 1998)。番茄基因組中約有 30%為重複性序列 (Peterson et al., 1996),甜菜約有 63%的重複序列 ( Flavell et al., 1974),而玉米重複性序
列更高達基因組的 60 - 80% (Viotti et al ., 1985)。有些重複序列只在某一基因組 出現或聚集在特定染色體,能作為基因組或染色體專一性分子標記 (marker)。
這類專一性分子序列亦可用來辨識染色體,克服傳統染色法面臨的問題而提高 核型分析的效率。
(五) 利用核醣體基因與端粒序列探討核型
植物基因組之重複 DNA 主要分成兩類;一類為縱向重複性序列 (tandemly repeated DNAs),聚集於染色體特定區域,如衛星體 DNA (satellite DNA)、端粒 序列 (telomeric sequences)以及核醣體 DNA 基因 (ribosomal DNA genes,簡稱 為 rDNA)。另一類為散佈型重複序列 (dispersed repetitive DNA elements),分 散於整個基因組或特定染色體上,如 DNA 轉位子 (DNA transposable elements) 以及反轉位子 (retroelements) (Kubis et al., 1998)。許多研究顯示高等植物中 大多數重複性序列不具功能,但有些重複序列如 rDNA基因與蛋白質合成有關,
端粒序列能夠保護染色體及維持染色體結構之穩定,而基因組中某些反轉位子 與物種演化、基因調節 (gene regulation)與基因表現上扮演重要的角色 (Kubis et al., 1998)。其中 rDNA 及端粒序列於探討核型上應用最多。真核生物細胞含
大量的核醣體 rRNA,參與蛋白質的合成,包括 17~18S- 5.8S-25~28S rDNA (45S rDNA )與 5S rDNA,屬於保守性序列。45S rDNA 位於染色體的核仁形成區 ( nucleolar organizer regions; 簡稱 NORs ),是形成核仁之主要成分 (Sumner, 2003),且在細胞分裂時,可觀察到次級收縮區 (secondary constriction)。Fukui 等人 (1998) 以 45S rDNA 為探針分析三種十字花科 Brassica 屬間不同種的植 物,發現 45S rDNA 的基因座在 B. rapa (A genome)、B. nigra (B genome)及 B.
oleracea (C genome)等二倍體植株染色體上的數目分別為五對、三對及兩對,顯
示物種演化過程中會造成 45S rDNA 分佈上的差異。5S rDNA 在相異物種其長 度有所不同,轉錄區域約為 120 bp,具高度保守性,轉錄區域為基因間間隔區 (intergenic spacer;簡稱 IGS)所隔開,此區段變異較大。以小麥為例,其 5S rDNA 片段有兩種大小:其中一種長度為 500 bp,包含 120 bp 的轉錄區及 380 bp 的 基因間隔區;另一種長度只有 400 bp,包含轉錄區同樣為 120 bp 及 280 bp 的基 因間隔區 (Mukai et al., 1990);甜菜的 5S rDNA 片段長度也具有 349 bp 與 351 bp 兩種 (Schmidt et al., 1994),顯示其單位長度的差異乃由於 IGS 所造成,研究指 出 5S rDNA 與 45S rDNA 的基因座數量與分佈位置在相異物種間有極大的變化 (Heslop-Harrison and Schwarzacher, 2011)。
由 FISH 為基礎發展出的多重螢光原位雜交 (Multicolor fluorescence in situ hybridization,簡稱 mFISH),能夠檢測染色體上多個基因位置。此法能同時標 定兩個以上的螢光素,可一次在相同細胞內觀察數個探針訊號,擴大了 FISH 技術的運用 (Trask, 1991)。利用 mFISH 可將兩種 rDNA 序列同時定位在染色體 上,能夠辨識細胞中相異染色體或比較種間核型演化上之差異。植物基因組中 較小的阿拉伯芥 (Arabidopsis),具有五對大小型態相似的染色體,以 FISH 方 法僅將 5S rDNA 與 45S rDNA 兩種探針同時定位,即可辨識阿拉伯芥中所有染 色體 (Murata et al., 1997)。佘等 (2012) 觀察 DAPI 染色的顯帶 (pattern)和兩 種 rDNA 雜交信號,對落花生 (Arachis hypogaea)的染色體進行了準確排列和配 對,建立詳細的分子細胞遺傳核型;其他農藝作物如高粱 (Bustos et al., 1996;
Taketa et al., 1999)、玉米 (Li and Arumuganathan, 2001)或者園藝作物如野草莓 (Fragaria vesca) (Rho et al., 2012)以及豬屎豆屬(也稱野百合屬) (Crotalaria) (Morales et al., 2012)等綠肥作物都有相關的應用。由於此兩種 rDNA 序列保守 性皆高,基因座數量及位置於不同物種間有所差異,能探討種間親緣關係。李 (2007) 分析 5S 及 45S rDNA 基因座於七種蝴蝶蘭屬植物之分佈,發現到同為
Phalaenopsis 節的 P. sanderiana, P. stuartiana, P. schillerinan 及 P. philippinensis
於中期染色體上都具有一對 45S rDNA 基因座,但其中 P. sanderiana 有兩對 5S rDNA 基因座,其餘三種蝴蝶蘭則均具有三對的 5S rDNA 基因座,與先前研究 建立的親緣關係樹相比,將此四種蘭花歸為兩群之結果相吻合。此外 P. equestris, P. celebensis 與 P. lindenii 均具有一對 45S rDNA 基因座及 5S rDNA 基因座,也
符合傳統分類將此三種蘭花歸類為 Stauroglottis 節之結果。因此利用兩種 rDNA 基因實質定位可用於探討物種演化之親緣關係。除了蝴蝶蘭屬植物外,Lan 與 Albert (2011)也利用 rDNA 基因探討同為蘭科之仙履蘭屬 (Paphiopedilum)植物 中染色體之演變。由此可窺見 rDNA 於染色體鑑識及物種演化上確實提供了有 利的分子證明。
被子植物的端粒序列,以單體 (monomer)七個核苷酸 TTTAGGG 縱列重複 所組成,最早是由阿拉伯芥中分離出來 (Richards and Ausubel, 1988),許多高等 植物的端粒序列均屬於阿拉伯芥型,其拷貝數在不同物種間有明顯的差異。以 端粒序列為探針應用於 FISH,可觀察染色體末端有無發生斷裂,能夠量測染色 體雜交訊號與端點之間的距離,為研究染色體核型與細胞遺傳圖譜上重要的分 子標誌。
(六) 分子基因庫的開發應用於基因定位與核型分析
基因庫含有生物體基因之重組 DNA 隨機片段,可透過標記的探針分子進 行篩選,進一步的純化與擴增後可作為遺傳研究的材料;其中細菌人工染色體 (bacterial artificial chromosome,簡稱 BAC)能夠建構大片段選殖基因庫,其內含 RFLP (restriction fragment length polymorphism)分子或其他分子片段的 BAC 選 殖株可直接應用在 FISH 上,將特定基因或序列直接定位在染色體上,建立染 色體實質圖譜 (physical map)或細胞遺傳圖譜 (cytogenetic map),是目前分子遺 傳研究重要的工具。2000 年 Dong 等人利用相異 BACs 鑑別馬鈴薯十二對中期
染色體,這些 BACs 中即帶有馬鈴薯或番茄遺傳圖譜上特定的 RFLP 分子標誌。
此外由於重複性序列在物種間及種內相異染色體的分佈位置有所差別,內含不 同種類重複性片段的 BACs 也廣泛運用在建立核型、探討物種演化與染色體重 組現象。
2010 年 Talia 等人依據染色體長度、中節位置並利用 rDNA 及一種反轉移 子重複性序列在染色體上之顯帶分佈,鑑定出向日葵 (Helianthus annuus)十七 對染色體。目前尚無運用 BACs 建立蘭花核型的完整研究,但彭 (2007) 曾選 殖朵麗蘭重複性序列 Dpr67、微衛星序列 (DpGA1、DpGA2)以及兩種核醣體基 因與端粒序列,並定位到朵麗蘭中期染色體上,依據雜交訊號的差異與序列分 佈建立朵麗蘭的核型。
大量分子標誌的開發加速核型的發展,並從中得到許多遺傳與演化更精準 的信息,有別於早期主要依據型態與交配的能力探討物種親緣。Zhao 等人在 2011 年觀察栽培種黃瓜 C. sativus 與兩種野生種黃瓜 C. xishuangbannesis、C.
hardwickii 的核型,由雜交訊號發現 C. sativus 與 C. xishuangbannesis 間含有許 多保守序列 (conserved sequences),證明兩者親緣關係相接近。而利用跨物種 螢光原位雜交 (cross-species FISH)技術,能夠提供親緣相近物種間核型之應用。
黃瓜與香瓜親緣關係相近,Liu 等 (2010) 利用黃瓜基因庫中已建立的分子標誌 (fosmids)為探針進行 FISH,建立了香瓜 (C. melo)十二對染色體核型。此外這 些分子標誌亦能探討物種演化中染色體結構之演變。Xiong 等 (2010) 觀察大白 菜 (B. rapa)與甘藍 (B. oleracea)及雜交後代油菜 (B. napus)的核型,從這些分 子訊號結果進一步觀察到雜交種油菜染色體序列重組的位置。Findley 等 (2010) 使用包含中節序列在內的十五個 BACs 建構了大豆 (Glycine max) 二十對染色 體核型,並由雜交訊號的位置與野生種大豆 (G. soja)相較,發現第十一對及十 三對染色體的長臂有發生異位 (translocation)的現象,且進一步偵測到異位的 片段大小分別為 4.2 Mb 與 17.9 Mb。對照 Kao 等 (2001) 以 DNA-Feulgen 染色
研究蝴蝶蘭核型,可以發現分子標誌基因庫的建構與 FISH 等生物技術相結合,
可提供更完整的核型分析圖,供後續分子生物與染色體遺傳研究。
(七) 以高解析力粗絲期染色體建立核型
早期研究核型多以初生根的根尖為材料,根尖分生組織旺盛、容易培養且 不受季節因子影響,核型多以有絲分裂中期的染色體型態為主 (闫等,2008)。
中期染色體為高度凝縮,每條染色體具有可辨識的型態,染色後能觀察到著絲 點及深淺不等的亮帶。然而中期染色體不足之處便在於收縮緊實的特性,染色 體型態相似,兩個標記的螢光訊號須至少分隔 1 Mb 才能明確加以分辨,故中 期染色體儘管能運用 FISH 確認分子標記約略的位置,解析力仍不足以建構高 清晰的染色體圖譜。減數分裂粗絲期染色體收縮程度低,呈現染色質(chromatin) 特殊的型態,可鑑別單一染色體及提高定位的解析力,且染色體製備過程中花 粉母細胞 (pollen mother cells,簡稱 PMCs)之細胞壁較根尖易為酵素所破壞,加 上粗絲期包含兩條配對的同源染色體 (homologous chromosomes),與尚未配對 的體細胞染色體相較,有較強的訊號,有助於提升染色體圖譜與核型分析上之 應用 (Chen et al., 2000)。
Singh 與 Hymowitz 曾於 1988 年時分離出雜交種大豆 (G. max x G. soja)二 十對粗絲期染色體,並分析了染色體總長度、異染色質及長臂 (long arm)、短 臂 (short arm)等數據,但當時的研究尚未使用任何分子標誌。近代運用 FISH 分析粗絲期染色體,屬禾本科 (Poaceae)作物水稻 (Oryza sativa)、玉米與茄科 (Solanaceae)作物的番茄 (Solanum lycopersicum)、馬鈴薯(Solanum tuberosum) 等研究最多。水稻是重要的糧食作物,有完整的分子遺傳圖譜,也有已建構的 BAC library (O. sativa ssp. japonica var. Nipponbare),其十二對粗絲期染色體能 藉由中心節序列及包含長短臂在內相異的 BACs 加以鑑別 (Cheng et al., 2001;
Tang et al., 2007);禾本科另一重要作物玉米,Chen 等 (2000) 曾以 FISH 定位 中心節、rDNA、含異染色質結之 180-bp 及 TR-1 (350-bp)等重複性序列,建立
玉米十對高解析粗絲期染色體核型。而玉米就單一染色體型態而論,則以第九 對粗絲期染色體研究最多,許多分子標誌陸續被定位出來,其中即包含抗甘蔗 螟蟲 (sugarcane borer)等重要的基因。(Danilova and Birchler, 2008; Sadder et al., 2000; Wang et al., 2006)。
粗絲期染色體長度的拉大提升了定位解析度,在單一條染色體上能同時觀 察到多個分子標誌。馬鈴薯與番茄的第六對染色體型態相近,Iovene 等 (2008) 以 FISH 定位內含三十個馬鈴薯遺傳標誌之 BACs 於兩者第六對粗絲期染色體 上,發現兩者第六對染色體有序列的共線性 (colinearity),且發現短臂之真染色 質 (euchromatin)處有一染色體倒轉 (inversion)的區域,與 Achenbach 等 (2010) 的實驗對照,該研究中發現馬鈴薯和番茄第五對粗絲期染色體長臂也有一處倒 轉的區域。因此較以往以中期染色體為對象的遺傳研究,利用粗絲期高解析度 分析,更明確提供馬鈴薯和番茄基因組中染色體重新排列組合的證據。除了重 要的糧食作物外,黃瓜也有完整高解析粗絲期核型 (Koo et al., 2005),以及棉 花 (Wang et al., 2010)等都有相關的粗絲期染色體研究。
姬蝴蝶蘭屬於小型染色體,染色體大小型態相似難以區別,於核型研究上 較為困難,相較於中期染色體,以高解析的粗絲期染色體為對象進行分析,將 可提高染色體的辨識度而予以適當的編號。過去並無蘭花粗絲期染色體核型相 關方面的研究,若此核型能完整建立,將是蘭科植物中第一個建立的核型系統,
可成為模式植物,提供其他蘭花作比較分析。
(八) 姬蝴蝶蘭功能性基因 SOC1 之特性及定位
本研究用於基因定位的 BAC,為與開花時間相關的基因 SOC1。開花是植 物繁衍後代重要的階段,受遺傳因子所控制,在模式植物阿拉伯芥的研究中發 現,開花過程受光週期、激勃素、春化作用及自發性調控等四條路徑所調控,
而 SUPPRESSOR OF OVEREXPRESSION OF CONSTANS (SOC1)為這些路徑
下游的整合者 (He and Amasino, 2005),調控蝴蝶蘭開花的時間。李 (2006) 曾 選殖姬蝴蝶蘭 SOC1 基因並針對該序列作分析,在序列保守區設計簡併性引子 (degenerate primer)擴增 SOC1 片段並選殖定序,以 RACE (rapid amplification of cDNA ends)方式獲得 SOC1 基因 cDNA 全長 (命名 PeSOC1)為 1,227 bp,編碼 區大小有 666 bp,可轉譯出 221 個氨基酸,其氨基酸序列與水稻 OsSOC1 相似 度為 57%,與文心蘭相似度高達 97.8%;進一步以南方點墨法分析 PeSOC1 顯 示其為單一拷貝的基因,此獨特序列適合作為本實驗染色體定位的專一性分子 標誌。
本實驗以姬蝴蝶蘭減數分裂粗絲期染色體為材料,針對每一對染色體 的特徵作描述並予以編碼,將兩種核醣體 rDNA、阿拉伯芥型端粒序列及 SOC1 基因定位到染色體上,建立姬蝴蝶蘭高解析力核型,以提供未來染色體圖譜或 遺傳圖譜研究上有力的資源。
三、材料與方法
(一) 材料來源
本研究分析的材料為台灣原生種蝴蝶蘭之一的蘭嶼姬蝴蝶蘭 P. equestris (2n = 2x = 38),由伍圓蘭園提供,以水苔種植於成功大學生命科學系溫室。姬 蝴蝶蘭之花期為夏季,待開花後採適當大小花苞,將花苞內的花粉塊予以適當 固定保存供染色體分析用。
(二) 姬蝴蝶蘭粗絲期染色體的製備 1.試驗材料的收集與保存
粗絲期係位於細胞週期中減數分裂的前期 (prophase I),染色體時期可以 由尚未開花前之花苞長度作簡單的判定。姬蝴蝶蘭開花後,收集長度約 4.5-5 mm 大小之花苞,以聶子及刀片小心取出花苞內之黃色花粉塊,置於新鮮配置 的 carnoy 固定液 (100% acetic acid : 95% ethanol 為 1 : 3)保存在-20℃冰箱中,
待隔天花粉塊顏色由黃色轉變成白色後,將樣品置換到 70%酒精保存以提高固 定的效果。為了比較粗絲期與中期染色體收縮程度的差異,除了花粉外同時也 採集根尖細胞供後續的分析。待姬蝴蝶蘭新長出的幼根約 1-2 cm 左右時,以解 剖刀切取末端的根尖,將根尖垂直對半後浸置於 2 mM 8-hydroxyquinolino 溶液 中 4-5 小時,溫度保持在 15℃。之後取出根尖,在室溫下以蒸餾水洗淨根尖兩 次,每次時間約 5 分鐘,再將此根尖作 carnoy 固定液處理至少三小時或置於固 定液中,於-20℃環境下保存樣品,待根尖轉為白色再置換到 70%酒精保存。
2.花粉細胞粗絲期的鑑定
花粉細胞鑑定的方式,係依據成大生命科學系張松彬老師實驗室所建立的 鑑定方法。保存後的花粉細胞進行鑑定前,先置換到蒸餾水,隨後將花粉置於 載玻片上,以刀片小心切出少許分生組織後,剩餘的花粉塊細胞暫時置回蒸餾
水。用刀片將樣品搗碎後,加一滴 1% 醋酸洋紅 (acetocarmine)染劑,隨即覆 蓋蓋波片 (24 x 32 mm)。將玻片置於酒精燈上來回火烤 3-4 次,此步驟須注意 不要讓醋酸沸騰。接著讓載玻片正面朝下置於濾紙上,以食指及中指固定,再 以大拇指前端輕壓玻片將染色體壓散,於顯微鏡下觀察染色體的時期。姬蝴蝶 蘭粗絲期染色體的特徵為可見一條條明顯散開的絲狀物。若確認該花粉為粗絲 期,將蒸餾水中剩餘之花粉分生組織細胞繼續保存在 70%酒精供後續染色體製 備用。
3.以加熱板製備染色體(用於核型分析)
本實驗係參考台大農藝系劉麗飛老師與張松彬老師合編之植物細胞分子顯 微技術一書中所用的方法。將鑑定過的花粉塊從酒精取出時,先浸泡於蒸餾水 至少 30 分鐘洗去酒精,將花粉切成數小塊以利後續的酵素反應。把樣品先放到 1x citrate buffer (4 mM citric acid, 6 mM sodium citrate, pH 4.5)中,於室溫下靜置 5-10 分鐘,再換到配置的三合一酵素液 ( 1% (w/v) cellulose Onozuka RS (Yakult Honsha), 1% (w/v) pectolyase Y23, 1% (w/v) pectinase solution ),將樣品置於 37℃
烘箱中作用 30-40 分鐘。酵素作用完畢後,用滴管小心吸出樣品置換到蒸餾水,
減緩酵素作用。準備乾淨的載玻片,吸取一花粉置於玻片中央,以濾紙吸去多 餘水分。用酒精消毒過的別針針頭輕輕搗碎花粉後迅速加一滴 45% acetic acid,
用針頭使花粉與 acetic acid 溶液均勻混合。將載玻片放在 42℃加熱板上,靜置 約 5 秒讓已混合的溶液向中央聚集。以別針在不碰觸玻片情況下,來回牽動此 液體並沿著玻片周圍移動擴散,讓染色體能夠平均分散在整個玻片上。在加熱 板上時間約 2 分鐘或樣本液體未完全乾掉前將載玻片從加熱板上移開,用現配 的 carnoy 固定液沖洗此玻片,隨後浸泡於 95% ethanol 數秒,將載玻片置於架 上風乾,再移至 37℃烘箱中,隔天將樣品保存在 4℃冰箱作後續核型分析用。
4.以液態氮製備染色體(用於核型分析與原位雜交反應)
從冰箱取出花粉塊後,仍先將樣品置於蒸餾水至少 30 分鐘,再置換到三合 一酵素液,於 37℃烘箱進行酵素反應 30-40 分鐘,待作用完成將樣品小心置換 到蒸餾水以洗去殘餘的酵素液。準備乾淨的載玻片,吸取一花粉塊至玻片中央,
以濾紙吸去周遭多餘水分,此時加一滴 45% acetic acid 於樣品旁邊,用濾紙吸 去 2/3 的 acetic acid,以別針針頭搗碎樣品後,將剩餘的 acetic acid 與樣品均勻 混合,蓋上蓋玻片(18 x 18 mm)並將載玻片翻轉置於濾紙上,用鉛筆平面端以彈 跳的方式輕輕敲打蓋玻片中央及周圍四個角,讓染色體更容易散開。將載玻片 浸泡於液態氮中,待嘶嘶聲停止及不再冒出氣泡時將載玻片移出液態氮,迅速 以刀片將蓋玻片挑開並將玻片置於 42℃加熱板,待玻片上殘餘的液體尚未完全 烘乾前,以現配的 carnoy 固定液沖洗玻片,浸泡於 95% ethanol 數秒,將載玻 片置於架上風乾,晾乾後於 Nikon ECLIPSE 顯微鏡下觀察,挑選細胞多以及型 態完整、無細胞質且異染色質明顯的玻片,置於 37℃烘箱,隔天將樣品保存於 4℃冰箱作後續原位雜交實驗用。
(三) 螢光原位雜交反應 1.質體 DNA 的抽取
本實驗螢光原位雜交所使用的四種探針及標定方式列於(表 1)。45S rDNA 序列片段來源為小麥 (Triticum aestivum),單一拷貝數長度約 9 kb,5S rDNA 序 列來自玉米 (Zea mays),單一拷貝數長度約 700 bp,Telomeric repeat 片段則取 自阿拉伯芥。將含有質體之單一菌落置於 5 ml LB 培養液 (Luria-Bertani medium, 10 mg/ml ampicillin)試管,於 37℃震盪培養 16 小時。核醣體 rDNA 及 telomeric repeat plasmid 的抽取根據 EasyPure Plasmid DNA miniprep kit (Bioman, Scientific) 與 Molecular Biology Tools (BioKit)製備。
分子標誌 SOC1 基因選自姬蝴蝶蘭 BAC library,該基因庫為成功大學生命 科學系吳文鑾老師實驗室所建構,基因庫中的 BAC 選殖體可作為基因定址選
殖 (map-based cloning)與染色體圖譜中基因定位重要的材料。將含質體的菌落 置於 25 ml LB 培養液 (含 12.5 ug/ml CM, chloramphenicol macrolides),於 37℃
生長箱震盪培養 24 小時。隔天以 13,000 rpm,2 分鐘離心並去除上清液。加入 1 ml STE buffer (10mM Tris-HCl (pH 8.0), 1 mM EDTA (pH 8.0), 0.1 M NaCl)後 充分震盪讓菌塊懸浮,於常溫下以 13,000 rpm 離心 2 分鐘去除上清液,此步驟 重複 2 次。加入 200 µl MP I (25mM Tris-HCl (pH 8.0), 10 mM EDTA (pH 8.0), 50 mM glucose)及 2 µl RNase (10 mg/ml)混合後,在冰上靜置 1 小時,以促進 RNase 的作用。待反應完成,加入 400 µl 現配之 MPII (10N NaOH, 10% SDS, ddH2O),
翻轉一次且在溶液完全透明前加入 300 µl MPⅢ (3M Potassium acetate (KOAC), 28.5% v/v Acetic acid, ddH2O),輕輕翻轉數次使白色沉澱物位於溶液上方。在 4℃下以 12,000 rpm 離心 10 分鐘,取 650-700 µl 透明的澄清液至新的微量離心 管,再加入等體積之 isopropanol,搖晃均勻,在-20℃下靜置 20 分鐘。待 DNA 沉澱,以 13,000 rpm,4℃下離心 10 分鐘並去除上清液,以 500 µl 70% ethanol 清洗 DNA 沉澱物,在 4℃下以 13,000 rpm 離心 5 分鐘去除上清液,於室溫晾 乾後,以 20 µl 65℃無菌水回溶並置於 4℃保存。
2.探針的標定
將含有 5S rDNA、45S rDNA、telomeric repeat 及 SOC1 的 plasmid DNA , 採用 nick translation 的方式,以 biotin-16-dUTP 及 digoxigenin-11-dUTP 標定探 針,標定的方法參考 Roche (Nick Translation Mix for in situ probes)的實驗步驟進 行。取 1-2 µg template DNA ,加入無菌水使體積至 16 µl,接著再加入 4 µl 的 Digoxigenin-Nick translation mixture (Roche Molecular Biochemicals) 或 (Biotin- Nick translation mixture)使總體積為 20 µl。以 13,000 rpm 短暫離心 30 秒,於 15℃
反應 2 小時。待作用完成後加入 1 µl 0.5M EDTA (pH 8.0)於 65℃水浴槽加熱 10 分鐘以終止反應,再加入 2 µl 4M LiCl (1/10 倍體積)及 50 µl 預冷(-20℃) absolute ethanol (2.5 倍體積)並混合均勻,置於-20℃冰箱以沉澱 DNA。隔天在 4℃下以
13,000 rpm 離心 20 分鐘,倒掉上清液後直接晾乾或以 70% ethanol 再清洗一次,
最後再以 20 µl 預熱(65℃)之無菌水回溶,保存在-20℃冰箱備用。
3.探針濃度之定量
探針定量的標定方式,參考 (劉與張,2008) -植物細胞分子顯微技術中提 到的方法。準備兩個乾淨的塑膠培養皿,以酒精噴洗乾淨。剪取適當大小之轉 印用尼龍膜 (Hybond-N+ Membrane, Amersham Pharmacia Biotech),置於塑膠培 養皿內,以鉛筆輕輕在膜上邊緣作記。將 5 ng/µl 濃度之 control DNA 以無菌水 稀釋使之濃度為 1000 pg/µl,再依序稀釋 10 倍(1 : 10、1 : 100 及 1 : 1000),由 左而右、濃度由高到低依序取 1 µl 點漬到 membrane 上。將保存於-20℃中之探 針短暫 vortex 後離心,先取 1 µl 加入 9 µl 無菌水稀釋 10 倍(1:10),再連續稀釋 10 倍 (1 : 100、1 : 1000、1 : 10000),同樣由左而右、濃度高到低依序取 1 µl 點漬到 membrane,所有待測的探針均依此方式做連續稀釋並點漬於膜上。將 membrane 以 120,00 uJ/cm2進行 crosslink (UV stratalinker 1800)。以 buffer 1 (100 mM maleic acid, 150 mM NaCl, pH 7.5 )潤洗 membrane 數分鐘,再將 membrane 置換到 10 ml buffer 2 (1% Roche blocking reagent in buffer 1 ) 30 分鐘。接著加入 2 µl Anti-dig AP-conjugate (或 streptavidin AP-conjugate , 150 mU/ml 稀釋 1:5000) 到 buffer 2 作用 30 分鐘。待反應完成,將 membrane 置換到 wash buffer (10 ml buffer1+0.3% Tween20)清洗兩次,每次 15 分鐘。最終進行呈色反應,將 membrane 移至 10 ml buffer 3 (100 mM Tris-HCl,100 mM NaCl,50 mm MgCl2, pH 9.5 ),再 加入 100 µl NBT/BCIP,保持在黑暗環境並小心不要移動 membrane 以確保呈色 均勻,於 15 分鐘至 3 小時可見定量結果,觀察膜上圓點顏色深淺與 control DNA 作比較,檢查探針濃度及評估原位雜交反應所需加入的探針量。
4.螢光原位雜交
原位雜交係參考 (Schwarzacher and Heslop-Harrison, 2000)以及 (劉與張,
2008)所提到的方法。在進行雜交反應前,先將挑選過型態完整的染色體載玻片 置於 37℃烘箱至少 8 小時或於 65℃加熱板上加熱 30 分鐘。當染色體周圍細胞 質明顯時,將載玻片放入 0.01 M HCl 清洗 2 分鐘,加 100 µl 稀釋過的 pepsine 酵素溶液 (pepsine : 0.01M HCl 為 1 : 100)於蓋玻片(24 x 40 mm)上,將載玻片正 面朝下吸附蓋玻片,之後將玻片仍以正面朝下的方式倒置於染缸中,於 37℃反 應 5 - 10 分鐘。酵素作用完畢後以無菌水清洗 2 分鐘,再以 2x SSC 於振盪器中 清洗 2 次,每次 5 分鐘。以 70%、90%、100% ethanol 連續脫水各 3 分鐘,靜 待其自然乾燥即完成玻片之前處理。
配置 20 µl 雜交混合液 (4 µl 50% dextran sulphate, 10 µl 100% formamide, 2 µl 20 x SSC, 0.5 µl 10% SDS, 3.5 µl probe DNA),於滾燙的沸水煮熱 10 分鐘,冰浴 5 - 10 分鐘。每一載玻片滴上 20 µl 雜交混合液,蓋上蓋玻片(24 x 32 mm),隨即放 到 80℃加熱板上靜置 2 分 30 秒,再將載玻片以正面朝下方式置於染缸,將染 缸放在內含濕紙巾的保溫盒(潮濕容器)內於 37℃烘箱中進行雜合反應。
隔天將載玻片浸泡於 2x SSC,讓蓋玻片自然滑落,並於 2 x SSC 中震盪搖 動 2 次,每次 5 分鐘,再將玻片放入 42℃之 SF50 (50% formamide, 2 x SSC)溶 液中浸泡 3 次,每次 5 分鐘。接著以 2 x SSC 清洗 2 次,每次 5 分鐘,再置換 到 4T (4 x SSC, 0.5% Tween20 )溶液清洗 5 分鐘。加入 100 µl TNB (1% blocking reagent in 1 x TN, 10 x TN = 1 M Tris-HCl,1.5 M NaCl, pH 7.5 )溶液於蓋玻片(24 x 40 mm)上,將載玻片正面朝下吸附蓋玻片,倒放置於染缸內於 37℃潮濕容器 中作用至少 30 分鐘。後續的抗體抗原反應皆以此方式吸附蓋玻片。待作用完畢 浸泡 4T 溶液讓蓋玻片自然滑落,取出載玻片,滴上 100 µl 以 TNB 稀釋之 Avidin Texas-Red (Vector Laboratories)抗體溶液 (Texas-Red : TNB 為 1 : 800),此過程 須避光處理,置於潮濕容器中 37℃下處理 1 小時。取出載玻片,以 4T 溶液清 洗 5 分鐘並讓蓋玻片自然滑落,再以 TNT (1 x TN ,0.5% Tween20)溶液清洗載玻
片 2 次,每次時間約 5 分鐘。接著再滴上 100 µl 以 TNB 稀釋之 Biotinlyed Anti-avidine (Vector Laboratories)抗體溶液(Anti-avidine : TNB 為 1 : 100)或 Sheep anti-digoxigenin-FITC (fluorescein isothiocyanate ,Vector Laboratories)抗體溶液 (anti-dig : TNB 為 1 : 200),於 37℃潮濕容器中處理 1 小時。取出載玻片,以 TNT 溶液清洗 3 次並讓蓋玻片自然滑落,每次 5 分鐘。滴上 100 µl 以 TNB 稀釋之 Avidin Texas-Red 抗體溶液 (Texas-Red : TNB 為 1 : 800)或 Anti-sheep-FITC (Vector Laboratories)抗體溶液 (anti-sheep : TNB 為 1 : 800),於 37℃潮濕容器中 處理 1 小時。將載玻片以 2 x SSC 清洗 2 次並讓蓋玻片滑落,每次 5 分鐘,以 70%、90%、100% ethanol 依序脫水各 5 分鐘,置於架上晾乾。乾燥後滴上 12.5 µl DAPI 螢光染劑,蓋上 24 x 40 mm 蓋玻片,於 Nikon Ti 螢光顯微鏡下觀察實 驗結果。
(四) 核型分析與雜交訊號影像處理
將製備完成的染色體玻片以 DAPI ( 100 µg/ml DAPI : 抗衰退劑 Vetrashield 為 1 : 19)螢光染劑或 PI (propidium iodide, 5 mg/ml 稀釋)染色 (24 x 40 mm 蓋玻 片覆蓋),於 Nikon Ti ECLIPSE 螢光顯微鏡下觀察染色體分布型態,以 CCD (Cooled charged -coupled device) camera 拍照將影像輸入到電腦存檔。挑選解析 度 高 、 染 色 體 型 態 完 整 且 均 勻 散 開 的 染 色 體 圖 片 , 以 Image J software (http://rsb.info.nih.gov/ij)中「straighten」功能將每一條單離的粗絲期染色體追蹤 之後拉直,再利用 Adobe photoshop CS2 及 Microscope NIS-Elements Software 等影像處理軟體測量染色體長度、長短臂以及異染色質大小等數據,並推估中 心節的位置及觀察染色體型態,建立初步的核型圖。另外將螢光原位雜交實驗 後的染色體玻片,以 Rhodamine 或 FITC 等螢光光源激發觀察雜交訊號的位置,
以 CCD camera 收集影像,利用影像處理軟體 NIS-Elements Software 整合螢光 標定的結果,於相同細胞中觀察相異的螢光訊號。
四、結果
(一) 姬蝴蝶蘭粗絲期染色體型態觀察
本實驗分析之蘭花為台灣原生種蘭嶼姬蝴蝶蘭 (2n = 2x = 38),具有 19 對 染色體;收集適當長度的花苞,將花粉粒以配置的 1% Acetocarmine 染色,將 染色體壓散後於光學顯微鏡下觀察花粉細胞,可發現粗絲期染色體呈現長短不 等的絲狀體,均勻分散在細胞中 (圖 1A)。以酵素打破細胞壁破壞細胞的結構,
將染色體壓散後以光學顯微鏡 1000X 倍率(目鏡+物鏡)觀察粗絲期染色體的型 態 (圖 1B);進一步以螢光染劑染色,其中 DAPI 是利用紫外光波長光線激發,
與雙股 DNA 結合時,最大吸收波長為 358 nm,最大發射波長為 461 nm。DAPI 主要係染序列中 AT rich 之處,於螢光顯微鏡下可看到每條染色體皆有一段亮 帶 (圖 1C),為異染色質所在的區域,有些亮帶特別明顯,可見姬蝴蝶蘭異染 色質中富含 AT 的序列。以影像軟體 Photoshop 處理,染色體呈現黑色與灰色相 間的圖片,分別代表異染色質及真染色質的區塊 (圖 1D),染色質處對比明顯,
可觀察到粗絲期染色體明顯的異染色質區段。
(二) 姬蝴蝶蘭粗絲期染色體核型分析
禾本科模式作物水稻,由於前中期染色體較小,各型態特徵不明顯,目前 國際上傾向於依粗絲期染色體的長度進行編碼,而在水稻的細胞遺傳研究中,
染色體編號是依據核型分析結果中染色體長度由長到短的順序來進行(程等,
1999)。姬蝴蝶蘭中期染色體型態相似,相鄰染色體之間差異不大,在沒有相關 的遺傳圖譜及完整的細胞遺傳研究輔助下,欲對染色體進行編號實為困難。本 實驗採用水稻編碼的方式,以粗絲期染色體核型的結果,將長度由大至小予以 編號並進行後續的分析。從染色體製片中挑選一個解析度最佳、異染色質清晰 且染色體均勻分散的細胞作為模式核型,以染色體長度大小予以初編號 (圖 2),
再比對不同的細胞樣本數作統計分析,整理的核型分析結果如 (表 2)所示:
1. 染色體長度與相對長度
本研究從染色體製片中共挑選 5 個較為完整的粗絲期細胞測定數據,除了 模式核型的細胞外,另外供作分析的 4 個細胞詳見 (圖 3),測得相異細胞中染 色體總長以及相對長度(占總長度之百分率)的數值。依據資料 (表 2)的結果,
姬蝴蝶蘭粗絲期染色體平均總長為 417.14 µm,就單一染色體而論,平均長度 為 21.95 µm;染色體最長約 32.07 µm,最短約 13.65 µm,分別占染色體總長的 7.82% 及 3.34%,兩者相差 2.35 倍。根據染色體相對長度的大小,可將染色體 大致分成四組:第一組包含第 1、2、3 對屬於較長的染色體,這三對染色體占 了總長的 20%;第二組為第 4~8 對染色體,相對長度加總共占 30%左右,前 8 對染色體長度幾乎就占了總長的一半;第三組含括第 9~17 對染色體,就單一 染色體而論,其相對長度都介於 4%-6%之間,染色體相對長度的加總約占總長 40%左右,這兩組皆屬於中等長度的染色體。最後 2 對屬於較短的染色體,其 長度加總後不到總長的 10%。
2. 染色質的分佈
染色質的分佈是染色體重要的型態特徵,可作為辨識不同染色體的依據,
這也是粗絲期在核型分析上優於中期染色體的主要原因之一。姬蝴蝶蘭粗絲期 染色體上分佈許多染色較深的異染色質結,尤其在染色體分化良好、解析度佳 的細胞中尤為明顯。在 (圖 2)的模式核型圖之中,可以觀察到其中第 19 對染 色體具有五個清晰且染色較深的異染色質結以及另外兩個染色較淺、相對較不 明顯的異染色質結。
相異染色體間異染色質的大小有極大的差異,綜合 (表 2)的數據分析,可 發現第 1 對染色體有一段長度極大的異染色質區,此區域就占了該條染色體總 長的 1 / 3,而第 4、7、8、11、15、16 對染色體也都具有較大的異染色質,長
度介於 7~9 µm,但其中第 2、13、14 等三對染色體則僅含有極少的異染色質,
平均長度不到 5 µm。真染色質主要為基因表現的地方,姬蝴蝶蘭粗絲期染色體 中真染色質與異染色質的大小,皆沒有隨染色體總長的改變而呈現相對應的增 加或減少,且就單一的染色體型態而論,這兩種染色質相對大小最多相差 5.6 倍(第 2 對),最少只有相差 1.1 倍(第 19 對),顯示在姬蝴蝶蘭中異染色質的大 小在某些染色體間有極明顯的差異,有利於染色體的辨識。
3. 中心節的位置
中心節是染色體核型上重要的特徵之一,姬蝴蝶蘭在粗絲期染色體尚未收 縮很短時,中心節的位置較為清晰而易於判斷,在顯微鏡下可看到染色體的中 心節是一染色較淺而兩側分布著染色較深的區域,待中心節位置確定後便可測 定出各染色體長臂與短臂的臂比 (Arm ratio)。對照 (表 2)的統計資料,臂比最 大的第 13 對染色體為 4.03,最小的第 16 對染色體為 1.25,兩者相差 3.22 倍。
依中心節的位置不同可將染色體分成四種型式,判定的標準係根據 Rho 等人 (2012)所描述:臂比介於 1.00~1.67 間屬於中位中節 (metacentric)染色體,表 中心節在染色體正中央;臂比介於 1.67~3.00 間為次中位中節 (submetacentric),
中心節偏染色體中央;臂比介於 3.00~7.00 之間為近端中節 (subtelocentric),
中心節接近染色體的一端;另外臂比在 7.00 以上則是端點中節 (telocentric),
中心節在染色體端點。對照 (表 2)的中心節統計數據,可以發現姬蝴蝶蘭的 19 對染色體中,有 8 對染色體屬於中位中節,10 對染色體為次中位中節,只有第 13 對染色體為近端中節,中心節的位置接近染色體的短臂處。臂比在某些染色 體間存在很大的差異,亦可作為辨識不同染色體重要的指標。
4. 衛星染色體的觀察
有的染色體除了具有初級收縮區 (primary constriction),即中心節所在的位 置外,尚有另一段次級收縮區,此處附有一段染色體稱為衛星區 (satellite),具
有衛星區的染色體又稱為衛星染色體 (satellite chromosome),幾乎所有生物細 胞至少有一對衛星染色體,大小不等且通常附在染色體短臂之處。從 (圖 4 A、
B)箭頭所指的地方,可以觀察到姬蝴蝶蘭單離的染色體短臂上,末端有一處染 色較深,且尚未完全收縮進來的區域,此處即為次級收縮區所在。觀察該染色 體的型態,發現與細胞中第 13 對染色體相當接近。因此觀察次級收縮區(衛星 染色體)的有無,可作為染色體識別的重要標誌。
5. 染色體收縮程度的差異
為比較姬蝴蝶蘭中期與粗絲期染色體收縮程度的差異,同時觀察有絲分裂 中期染色體的型態 (圖 5 )並測量染色體長度 (表 3)。(圖 5 A)的中期染色體圖,
經測量後得到染色體平均長度為 1.78 µm,染色體最大與最小相差約 2 倍。而 (圖 5 B)的染色體平均長度是 1.76 µm,兩個樣本細胞中平均長度大小非常相近,
與粗絲期染色體 (21.95 µm)相較下,平均長度相差約 12 倍,兩者最大的染色 體間相差約 13 倍。因此以粗絲期染色體建立核型,將可提高至少 12 倍以上的 基因或序列定位解析力。
(三) 姬蝴蝶蘭螢光原位雜交與基因定位 1. 重複性序列的定位
核醣體 DNA 基因與端粒序列皆屬於縱向重複性序列,普遍分布在染色體 上特定的位置。將 45S rDNA 以 biotin-16-dUTP 及 digoxigenin-11-dUTP 標定探 針,以螢光原位雜交分別定位在姬蝴蝶蘭粗絲期染色體上 (圖 6),前者 (biotin) 以 Rodamin (紅色螢光)偵測,後者 (dig)以 FITC (綠色螢光)偵測。依據 (圖 6 ) 的結果,45S rDNA 於一對中等長度的染色體末端呈現明顯的訊號,而在 (圖 4)的單離染色體上曾觀察到次級收縮區,於 (圖 6 B、C)該染色體末端確實也 出現 45S rDNA 的訊號,顯示藉由 45S rDNA 的觀察,可判斷具有衛星體的染 色體。以阿拉伯芥型的端粒重複序列 (TTTAGGG)n 為探針進行螢光原位雜交,
觀察到單離染色體的端點上有微弱的訊號 (圖 6 D)。以雙標的螢光原位雜交同 時觀察這兩種序列的分佈 (圖 7 ),可發現到端粒序列的訊號大多位於染色體的 端點上,顯示姬蝴蝶蘭染色體的端粒是屬於阿拉伯芥型的端粒序列,而 45S rDNA 則出現在第 13 對染色體的末端。此外再將另一種核醣體基因 5S rDNA 定位在染色體上,螢光原位雜交的結果顯示,5S rDNA 於不同的細胞中皆出現 在一對長度較短的染色體上,且訊號皆分佈在染色體的末端 (圖 8 ),由 (圖 8 A) 內含 5S rDNA 染色體實際分析的數據指出,該染色體的平均長度將近 16 µm,
異染色質與真染色質分別約為 6.2 µm 以及 9.6 µm,以及依據染色體型態與異染 色質比率(40%)等型態與特徵皆與第 17 對染色體相似,比對 (圖 8 D)單離染色 體型態,共同之處皆在於,異染色質的分佈位置很靠近染色體的端點。
2.內含功能性基因獨特序列的定位
以螢光原位雜交技術將開花時間相關基因 SOC1 定位在染色體,發現於一 對染色體上出現明顯的訊號,該訊號的型式包含兩個明顯的點以及一個較微弱 的點,觀察細胞分裂中染色間期 (interphase)的細胞同樣也發現到一對訊號的分 佈 (圖 9 A)。另外在相異細胞中也觀察到其中一對染色體有出現 SOC1 基因的 訊號 (圖 9 B),對照 李(2006) 對 SOC1 基因進行的序列分析,本實驗證實了 SOC1 在姬蝴蝶蘭中確實為單一拷貝的基因序列;另外於相異細胞單離的染色 體上,同樣偵測到 SOC1 基因的訊號,而訊號的型式包含一個極為明顯的點以 及兩個相對較微弱的點 (圖 10 A),以影像軟體編輯加強染色質的對比 (圖 10 B) 並對染色體進行分析,發現該染色體長度約為 25.25 µm,異染色質與真染色質 大小分別為 8.58 µm 與 16.67 µm,其異染色質占了染色體總長的 34%,臂比約 為 1.4,SOC1 基因位於染色體短臂的真染色質上,距離短臂末端約 5.16 µm,
無論染色體型態與統計數據比對的結果發現最接近第 1 對染色體。
(四) 姬蝴蝶蘭結合分子標誌高解析核型
綜合不同細胞染色體型態的觀察、長度的測量、異染色質的大小與分佈位 置、臂比的計算以及整合螢光原位雜交訊號的結果(其中 CMT 與 EFS 基因的位 置係分別參考林 (2012) 以及周 (2012) 所分析的結果),手繪出姬蝴蝶蘭核型 之模式圖 (ideogram) (圖 11),建構姬蝴蝶蘭完整的核型分析。根據這些結果,
針對每一對染色體的特徵分別描述如下:
第 1 對染色體:屬於次中位中節染色體,平均長度為所有染色體中最大,中央 有一段極長的異染色質,約占染色體總長的 1 / 3,大多分佈在
長臂上。短臂真染色質上有一個 SOC1 的基因。
第 2 對染色體:屬於中位中節染色體,異染色質較少,均勻分佈在中心節的兩 側,真染色質與異染色質的相對比例為所有染色體中最大。
第 3 對染色體:屬於中位中節染色體,異染色質均勻分佈在中心節兩側,短臂 的長度為所有染色體中最大。
第 4 對染色體:屬於次中位中節染色體,異染色質較多,主要分佈在長臂。
第 5 對染色體:屬於中位中節染色體,異染色質均勻分佈在中心節兩側,長短 臂的比例較均勻。
第 6 對染色體:屬於次中位中節染色體,短臂上僅含有極少量的異染色質,長 短臂相差約 2 倍,靠近長臂的末端有一個 CMT 基因(附圖 1)。
第 7 對染色體:屬於次中位中節染色體,中心節兩側異染色質較大。
第 8 對染色體:屬於次中位中節染色體,有較多的異染色質均勻分佈在中心節 兩側,異染色質平均長度約 8.5 µm,含量僅次於第 1 對染色體。
第 9 對染色體:屬於次中位中節染色體,異染色質均勻分佈在中心節兩側,臂 比相對較大。
第 10 對染色體:屬於次中位中節染色體,異染色質集中在短臂處,長短臂相差 約 2 倍,靠近長臂的末端有一個 EFS 的基因(附圖 2)。
第 11 對染色體:屬於次中位中節染色體,有較多的異染色質,主要分佈在長臂。
第 12 對染色體:屬於中位中節染色體,長短臂的分佈較均勻,異染色質於短臂 上的分佈量略多於長臂。
第 13 對染色體:屬於近端中節染色體,具有最大的臂比,中心節靠近短臂的端 點。異染色質的大小與短臂皆為所有染色體中最小。其中異染 色質的分佈位置相當靠近短臂,且在短臂末端有一 45S rDNA
的基因,為染色體次級收縮區所在的地方。
第 14 對染色體:屬於次中位中節染色體,異染色質相對較少,於長臂上的分佈 量略多於短臂,長短臂相差約 2.5 倍。
第 15 對染色體:屬於中位中節染色體,有較多的異染色質,主要分佈在長臂。
第 16 對染色體:屬於中位中節染色體,異染色質均勻分佈在中心節兩側,長短 臂的分佈最均勻,臂比為所有染色體中最小。
第 17 對染色體:屬於次中位中節染色體,異染色質的分佈位置相當靠近短臂,
長短臂的分佈比較不均勻,臂比相對較大,短臂末端有一 5S rDNA 的基因。
第 18 對染色體:屬於中位中節染色體,異染色質較多,但主要集中在長臂,短 臂上僅含有極少量的異染色質。
第 19 對染色體:屬於中位中節染色體,平均長度最小。真染色質與長臂皆為所 有染色體中最小,長短臂相差約 1.5 倍,真染色質與異染色質 的相對比例亦為所有染色體中最小,僅相差 1.1 倍。
五、討 論
(一) 姬蝴蝶蘭粗絲期染色體的製備與型態觀察
蘭嶼姬蝴蝶蘭以花粉母細胞減數分裂中粗絲期染色體分析核型,在一定程 度上能夠克服體細胞染色體型態過小、解析力不足的缺點,不過以粗絲期染色 體進行核型分析時也存在不少的困難。由於粗絲期染色體長度過長,在製備的 過程中染色體容易纏繞,相互之間產生重疊而難以分散,無法對個別染色體型 態作分析;許多細胞除了相互纏繞外,還會有單一的染色體各自游離出去,常 無法收集到完整的 19 對染色體,如 (圖 3 C)與 (圖 3 D)的細胞中各缺少一對 染色體。另外也因為粗絲期染色體長度拉長的特性,染色體在製備過程中容易 有斷裂的情形,這在收縮較為緊實的中期染色體上較不明顯。雖然以新鮮採集 的花粉母細胞來製備可減少此情況的發生,但仍然無法完全避免,必須從製片 中盡量挑選出最為完整的細胞。此外各染色體上一些重要的型態特徵如中心節 的位置等,於粗絲期階段未必都能清楚加以辨識,加上細胞分裂與不斷的收縮 過程中,該特徵又會發生變化,間接造成核型分析上的誤差。儘管利用粗絲期 染色體分析核型有其難度,但仍有相對的優勢存在;粗絲期除了提高分析上的 解析力外,還有兩個重要的優點:其一即同源染色體聯會 (synapsis)可避免辨 別同源染色體的麻煩,另一方面粗絲期染色體上分佈許多染色後深淺不同的染 色質節,可作為識別特定染色體的重要標誌 (程等,1999)。
(二) 姬蝴蝶蘭粗絲期染色體核型分析
以禾本科模式作物水稻為例,Shastary 於 1960 年最早報導了水稻減數分裂 粗絲期核型分析結果,並以長度大小編碼染色體,但當時的染色體製片技術尚 未純熟,染色體不夠分散、染色質特徵不明顯,從而限制了粗絲期核型的應用。
Kurata 等人於 1981 年將體細胞染色體製片中酶解火焰乾燥法引進到粗絲期的 製備上,卢等 (1990) 即利用此方法,獲得了優良的粗絲期染色體製片,分析