國⽴台灣⼤學醫學院臨床醫學研究所 博⼠論⽂
Graduate Institute of Clinical Medicine College of Medicine
National Taiwan University Doctoral Dissertation
間葉幹細胞上類鐸受體 2 配體增強間葉幹細胞於氣喘模式⼩
⿏之療效
Toll-Like Receptor 2 Ligation Enhances Therapeutic Effects of Mesenchymal Stem Cells on Murine Model of Asthmatic Inflammation
俞惠潔
Hui-Chieh Yu
指導教授:江伯倫 教授
Advisor: Bor-Luen Chiang, M.D., Ph.D.
中華民國 107 年 4 ⽉
April, 2018
致謝
感謝指導教授以及⼝試委員們的指正與提醒,不僅幫助我完成這份論⽂,更
讓我學習到⽼師們做學問時求深且求精的態度。
感謝⼀路上有實驗室同仁們相互切磋,相互提攜,不僅幫助我完成學業,更
豐富我的⽣活經驗。
感謝我的家⼈及朋友們的⽀持與陪伴,讓我始終可以保持愉快的⼼情,積極
應戰。
在完成博⼠學位的路上,儘管碰撞,儘管孤獨,儘管像是⼀個被放逐到荒島
掙扎求⽣的歷程,因為有了師⾧與同仁的協助,有了家⼈與朋友的陪伴,我終於 有了享受在荒島上恣意舞蹈的勇氣。
Catalogue
中⽂摘要---8
Abstract---9
Chapter I: Introduction The facts and the unsolved problems of asthma---10
The attempts and the failures of treating asthma based on its molecular mechanisms--10
Mesenchymal stem cells in the treatment of immune disorders---11
The immunosuppressive mechanisms of mesenchymal stem cells---14
The immunoregulatory molecules of mesenchymal stem cells---16
Heterogenous mesenchymal stem cells---17
Mesenchymal stem cell polarization---19
CD4+CD25+Foxp3+ Treg---22
Chapter II: Materials and Methods Materials---25
Methods---27
Chapter III: Results Characterization of mouse BM-MSCs---35
Post-treatment as the more appropriate setting of BM-MSC treatment---35
Administering 5´105 of BM-MSCs for higher immunosuppressive activities in vivo---36
TLR mRNA expression of mouse BM-MSCs---36
Pam3CSK4 as the comparatively effective stimulant for enhancing regulatory activities in BM-MSCs---37
Deciding to treat BM-MSCs with 1 µg/ml Pam3CSK4 for 96 h---38
Enhanced immunosuppressive activities in Pam CSK -stimulated BM-MSCs---38
Pam3CSK4 changed the multipotent stem cell properties of BM-MSCs---40
STAT3 signals of the immunosuppression-prone BM-MSCs---40
Nitric oxide as the dominant immunoregulatory factor of Mpam---41
The mobility of BM-MSCs might be increased with Pam3CSK4 treatment---42
Unchanged immunoprivileged properties of the conditioned BM-MSCs---43
The molecules indicating apoptosis and anergy in CD4+ T cells were not induced by Mpam---43
Regulatory T cells induced by nitric oxide as the immunosuppressive operators---43
Increased CD4+CD25+Foxp3+ T cells induced by Mpam---44
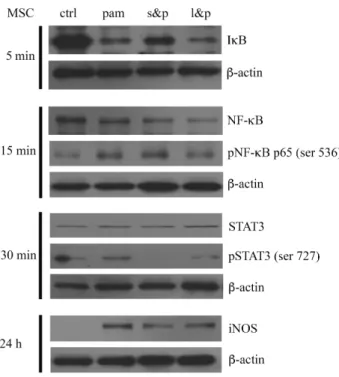
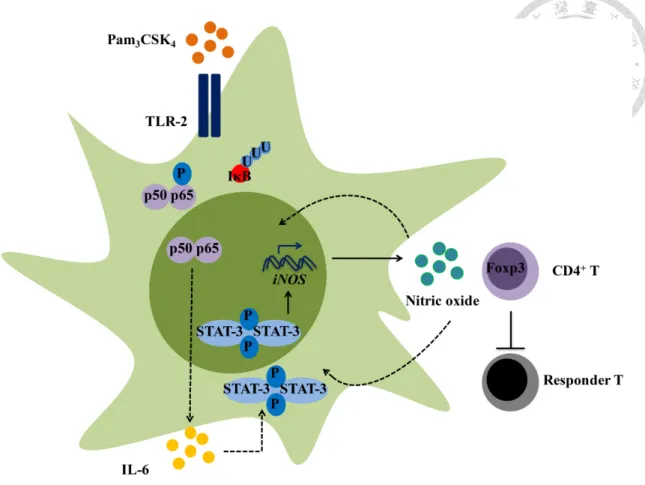
Sequentially activated NF-kB/STAT3/iNOS signaling in Mpam---45
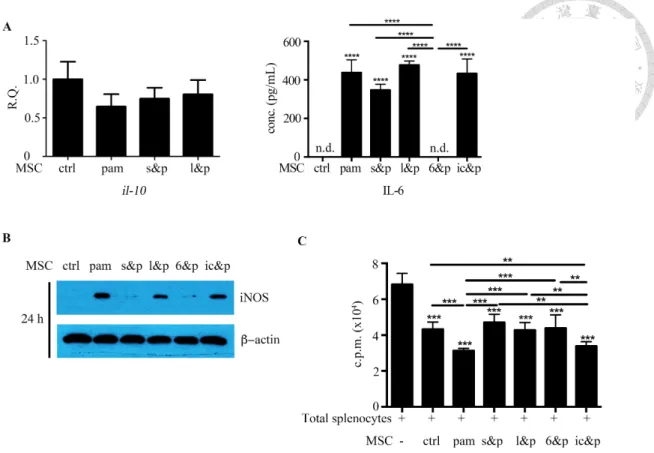
Mediation between NF-kB and STAT3 through IL-6---46
Chapter IV: Discussion This study established an appropriate priming protocol to enhance the immunosuppressive function of BM-MSCs---47
This study validated the Pam3CSK4-modified BM-MSCs with the understanding of their immunosuppressive mechanisms Increased NO secretion---51
More CD4+CD25+Foxp3+ Treg cells---55
Via TLR2/NF-kB/IL-6/STAT3 signaling pathway---60
Chapter V: Prospects---65
Chapter VI: References---67
Chapter VII: Figures Fig 1. Experimental designs---85
Fig 2. Characterization of BM-MSCs---87
Fig 3. Post-treatment as a more appropriate setting of BM-MSC treatment---89
Fig 4. Administering BM-MSCs in the amount of 5´105 cells resulted in
higher immunosuppressive activities in vivo---90 Fig 5. TLR expression in BM-MSCs---92 Fig 6. Pam3CSK4 as the comparatively effective TLR ligand for enhancing
regulatory activities in BM-MSCs---93 Fig 7. Pam3CSK4 as the comparatively effective stimulant for enhancing
regulatory activities in BM-MSCs---95 Fig 8. Pam3CSK4 at 1 µg/mL was chosen to modify BM-MSCs---97 Fig 9. Pam3CSK4 at 1 µg/mL was chosen to treat BM-MSCs for 96 h---99 Fig 10. Immunosuppressive activities of BM-MSCs were enhanced by
Pam3CSK4 stimulation---100 Fig 11. Mpam ameliorated systemic inflammation of asthmatic mice more
effectively than Mctrl---101 Fig 12. Mpam alleviated local symptoms of asthmatic mice more effectively than did Mctrl---102 Fig 13. Compared to Mctrl, Mpam was more likely to improve airway
remodeling in asthmatic mice---104 Fig 14. Pam3CSK4 changed the multipotent stem cell properties of
BM-MSCs---106 Fig 15. CD4+ T cells were suppressed through the TLR2/STAT3/iNOS
signaling pathway in Mpam in a cell-cell contact-dependent
manner---108 Fig 16. The mobility of BM-MSCs might be increased with Pam3CSK4
Treatment---110
Fig 18. The molecules indicating apoptosis and anergy in CD4+ T cells were not induced by Mpam---113 Fig 19. Mpam induced more regulatory T cells---114 Fig 20. More CD4+CD25+Foxp3+ T cells were detected after the Mpam
treatment in vitro and in vivo---116 Fig 21. Pam3CSK4 induced NF-κB/STAT3/iNOS signals in BM-MSCs---118 Fig 22. IL-6 Mediated NF-κB/STAT3 signaling in Mpam---119 Fig 23. Suggested mechanisms of the enhanced immunosuppressive
properties of Mpam---120 Chapter VIII: Appendix
Table 1. PCR primers used in this study---121 Fig 1. MSCs suppress immune cells directly and/or indirectly---123 Fig 2. MSCs suppress proinflammatory T cell subsets and natural killer cells while stimulating regulatory T cell subsets---124 Fig 3. MSCs polarize macrophages into regulatory macrophages, instead of proinflammatory macrophages---125 Fig 4. MSCs inhibit the differentiation, maturation, and proinflammatory
polarization processes of DCs---126 Fig 5. Structure and signaling pathway of TLR2---127 Chapter IX: Abbreviations---129
間葉幹細胞上類鐸受體 2 配體增強間葉幹細胞於氣喘模式⼩⿏之療效
摘要
現⾏的治療⽅法無法達成治癒氣喘的⽬標,因此發展新的氣喘療法仍是當務 之急。僅管間葉幹細胞近來已因其免疫抑制特性的優勢⽽有潛⼒成為新興氣喘療 法,其潛在的異質性將削弱其治療效益。在這篇論⽂中,我們進⾏了⼀個利⽤類 鐸受體 2 之配體刺激間葉幹細胞以增強其氣喘療效的實驗計畫。根據實驗結果,
在尾靜脈注射投予間葉幹細胞前,⽤⼀個類鐸受體 2 之配體(Pam3CSK4)刺激間
葉幹細胞,可增強其對已誘發氣喘的⼩⿏的療效。為確效此刺激⽅法,我們進⼀
步分析其作⽤機轉,並證實在間葉幹細胞中,Pam3CSK4 可透過轉錄因⼦ NF-κB
的途徑,刺激細胞激素 IL-6 的產⽣,進⽽活化轉錄因⼦ STAT3。轉錄因⼦
STAT3 磷酸化⽽活化後,隨即刺激激發型⼀氧化氮合成酶(iNOS)的表現量,
進⽽合成更多與間葉幹細胞免疫調控能⼒相關的關鍵因⼦⼀氧化氮(NO)。經
Pam3CSK4 刺激的間葉幹細胞最後便藉由誘發更多調控型 CD4+CD25+Foxp3+ T 細胞⽽達到其免疫抑制能⼒增強的效果。本篇研究所提出的間葉幹細胞改良⽅
法,除在細胞和⼩⿏氣喘模式上均可得到較為穩定的治療成效外,我們更進⼀步 藉由研究其免疫調控機轉⽽達到確效的⽬的。藉由對間葉幹細胞的改良,未來可 望能減少利⽤間葉幹細胞治療所產⽣的副作⽤,以利⾧期治療的規劃。
Abstract
Under current therapeutic strategies, the cure to asthma remains elusive; thus- novel approaches for treating asthma are desperately needed. Despite that mesenchymal stem cells (MSCs) have recently been established as potential candidates by virtue of their immunomodulatory properties, the underlying heterogeneity of MSCs diminishes their therapeutic efficacy. Here we evaluated a toll-like receptor (TLR) 2 ligation protocol of MSCs to augment their therapeutic benefits on asthma. We surmise that more effective MSCs were activated with a TLR2 ligand, Pam3CSK4, at low
concentration (1 µg/mL) for a longer period (96 h) induction prior to intravenous administration for ameliorating asthma in mice in a post-treatment manner. We further validated this regimen by demonstrating that Pam3CSK4 activated STAT3 in BM-MSCs through IL-6, which was likely stimulated with NF-κB signaling. Nitric oxide (NO), the key suppressive molecule of Pam3CSK4-primed BM-MSCs, was later highly increased through upregulation of iNOS, which was in the downstream of STAT3
phosphorylation. The intensified suppressive functions of BM-MSCs were then
executed by inducing CD4+CD25+Foxp3+ regulatory T cells in a post-treatment manner.
The consistent therapeutic outcomes and the valid suppressive mechanisms in the study might shed light on eliminating the proinflammation-prone uncertainties of MSCs, and enhancing the future feasibility of obtaining long-term effects with this regimen.
Key words: Mesenchymal stem cell; Heterogeneity; Toll-like receptor 2; Nitric oxide; Regulatory T cell
Chapter I: Introduction
The facts and the unsolved problems of asthma
Asthma is a chronic inflammatory disease, affecting an estimated 300 million individuals of all ages worldwide. Annual worldwide deaths from asthma have been estimated at 250,000 (1). The etiological complexity of asthma is attributed to the interaction between heritable and environmental components. Chronic airway
inflammation is induced to cause the chief complaints of asthma- recurring episodes of intermittent dyspnea, wheezing, and nighttime or early-morning coughing.
Subsequently, the recurrent inflammation leads to airway remodeling which causes smooth muscle spasm and airway narrowing.
Asthma is diagnosed based on medical and family histories, a physical
examination, and pulmonary function tests. Current pharmacotherapeutic strategies of asthma are categorized by the severity of the cases. According to the progression of the disease, short-term bronchodilators are prescribed for patients with mild asthma as needed, while a combination of long-term bronchodilators and steroids is prescribed for patients with severe cases (2). However, the standard strategies are still far from perfect.
Two problems remain unsolved: asthma can only be controlled with current protocols- even the patient with mild asthma despairs of being cured. Moreover, around 5% to 10% of the difficult-to-treat population is refractory to the most aggressive standard treatment (1, 3). Therefore, new approaches to treat asthma are desperately needed.
The attempts and the failures of treating asthma based on its molecular
Molecular therapies have recently been suggested as potential solutions (4). T helper (Th) 2 lymphocyte has been considered the most important part in the
pathogenesis of allergic asthma. Th2-type cytokines including interleukin (IL)-3, 4, 5, 9, 13, granulocyte-macrophage colony-stimulating factor (GM-CSF), thymic stromal lymphopoietin (TSLP) and their downstream immune responses contribute to varieties of pathological features. For example, IL-4 promotes differentiation and proliferation of Th2 cells which induces B cells into immunoglobulin (Ig)-E-producing plasma cells.
IL-5 signals are critical for the growth, maturation and activation of eosinophils. IL-5 associated eosinophil infiltration in the airway is related to the induction of airway hyper-responsiveness. IL-13 is secreted by activated Th2 cells. IL-13 stimulates airway hyperactivity and mucous hyperproduction through IgE production, eosinophil
recruitment and fibroblast proliferation in the lungs (5, 6). Hence, specific modulation of the Th2 profile has been suggested as a new strategy of asthmatic therapy. Except for Th2-derived cytokine antagonists or neutralizing antibodies, administration of several cytokines including IFN-γ, -α and -β and IL-12 have been trialed in attempts to suppress Th2 responses in asthma. However, the limited success with a particular target molecule is attributable to the complexity of asthmatic pathogenesis. In addition, no combination of different target molecules has achieved an advantageous benefit-risk ratio (6).
Moreover, neither non-selective standard therapeutic strategies nor these specific approaches lead to prolonged periods of drug-free remission (7). Therefore, novel treatments with homeostatic properties may achieve more useful results.
Mesenchymal stem cells in the treatment of immune disorders
Cell-based therapy is a follow-up to the small-molecule drugs and biologicals. In contrast to traditional drugs, cells are capable of sensing and responding to
environmental signals, not to mention that the longevity of cells provides long-term treatment with the potential to restore immunological imbalances (8).
Resident mesenchymal stem cells (MSCs) and MSC-like cells exist in most postnatal tissues. They can be isolated from bone marrow, muscle, fat, hair follicles, tooth root, placenta, brain periosteum, dermis, perichondrium, umbilical cord,
Wharton’s jelly, lung, liver and spleen. However, bone-marrow (BM)-derived MSCs are the most characterized MSCs displaying distinct transcriptomes, and can thus be considered distinct cell types in a therapeutic setting (8). BM-MSCs are identified as multipotent non-hematopoietic progenitor cells with three distinct qualities. First, they express mesenchymal lineage surface markers like CD73, CD105 and CD44, but not hematopoietic ones like CD34, B220 and CD11b. Second, they are capable of
differentiating into mesenchymal cell lineages including the adipogenic, osteogenic and chondrogenic lineages. And third, BM-MSCs perform their immunosuppressive
properties on a variety of cell types mediating both innate and adaptive immunity (8).
MSCs, identified initially as multipotent nonhematopoietic progenitor cells, are considered attractive candidates in regenerative medicine because of their self-renewal and tri-lineage differentiating features (9). However, despite the clear therapeutic benefits of MSC-based therapies reported in previous studies, less than 1% of infused MSCs have been found at the target sites, even though their homing to the injured tissues was tracked immediately after administration. Since MSCs disappear shortly after infusion in the injured tissues, the repairable microenvironment they build is instead suggested as a more powerful facility.
Consequently, the immunoprivileged properties and immunomodulatory molecular expression have been suggested as the main therapeutic mechanisms of MSCs, thus shedding light on the treatment of degenerative disorders, allograft rejection, and inflammatory and autoimmune diseases, even when cross-species applications are involved (10-12). In the treatment of multiple sclerosis (MS) and autoimmune type I diabetes, intravenous (i.v.) infused MSCs exhibit beneficial effects on experimental autoimmune encephalomyelitis (EAE) and non-obese diabetic (NOD) animal models, respectively. Allogenic MSCs protect fas-/- mice, a disease model of systemic lupus erythematosus (SLE), from autoantibody-induced renal dysfunction. In addition, even the xenogenic MSCs have been reported to significantly alleviate colon inflammation and body weight loss in inflammatory bowel disease (IBD) mice (13). Furthermore, despite that the precise MSC therapeutic mechanisms are unclear, many clinical trials are ongoing to evaluate its safety and efficacy (14). In SLE and Crohn’s disease patients, both autologous and allogenic MSCs are able to suppress inflammation and reduce damage to kidneys and intestines, supposedly by the induction of regulatory T cells in patients. BM-MSCs can also exert therapeutic effects on patients suffering from diabetes, MS, stroke and liver cirrhosis, likely through immediate immunomodulatory effects. One or two infusions of MSCs after BM transplantation greatly improve the survival rate of both mice and humans with graft-versus-host disease (GvHD). And most encouragingly, MSCs successfully reverse steroid-resistant GvHD in patients receiving BM transplantation (15).
More recent studies on the immunomodulatory effects of MSCs have also evidenced their efficacy in animal models of lung injuries, chronic obstructive pulmonary disease (COPD), asthma and pulmonary hypertension (16, 17). Using an acute ovalbumin (OVA)-induced asthma murine model, both autologous and allogenic
BM-MSCs ameliorate methacholine-induced airway hyper-reactivity. Th2-driven inflammatory signs such as circulating IgE and IgG1 levels, pulmonary eosinophil recruitment, and pulmonary IL-4 and IL-5 secretion are mitigated by allogenic BM- MSCs (18). Using a chronic OVA-induced asthma murine model, researchers have found that the airway remodeling can also be improved with infused BM-MSCs.
The immunosuppressive mechanisms of mesenchymal stem cells
Reports have revealed the correlation between the therapeutic effects of MSCs and their abilities to directly abrogate T cell, macrophage, dendritic cell (DC), neutrophil, and B cell proinflammatory functions (please see appendix for Fig. 1). The first evidence of the regulatory functions in MSCs in vivo came from models of GvHD.
These studies demonstrate that MSCs reduce allograft rejection partly by modulating T lymphocytes (19). In addition, the timing of MSC administration is critical for natural killer (NK) cell and cytotoxic T cell modulation. It seems that MSCs have to be present when NK cells are stimulated. Contrarily, MSCs are effective at modulating cytotoxic T cell proliferation at the beginning of the mixed lymphocyte reaction (20). Further, MSCs prevent the differentiation and expansion of T cell subtypes which possess proinflammatory behaviors, Th1 and Th17 (21, 22); while promoting T cell subtypes which possess immunomodulatory behaviors, Th3 and Treg (23). To date, several reports support that the ability of MSCs to promote the Treg subtype is dependent on cell-cell contact as well as specific MSC to T cell ratios. With a low MSC to T cell ratio, the T cell proliferation is reduced, however the percentage of Treg in the population is elevated. (24) (please see appendix for Fig. 2).
Next, MSCs are proved to maneuver macrophage plasticity towards regulatory macrophage (M2) behavior. In an experimental model of colitis, MSCs facilitate
therapeutic effects by directly acting on M2 induction (25) (please see appendix for Fig.
3).
Third, MSCs have been shown to affect DC differentiation, maturation, and function in numerous in vitro coculture systems. MSCs inhibit CD34+ and monocyte- derived DC through MSC-derived arrest of cell cycle in G0 (26). MSCs are as well observed to reduce major histocompatibility complex (MHC) class II and costimulatory molecules in DC, and consequently impair the stimulation of T cell proliferation (27).
Moreover, they are indicated to enhance regulatory mature DC (DC2) function, while decrease proinflammatory mature DC (DC1) function (28) (please see appendix for Fig.
4).
Fourth, in both the reduced neutrophil recruitment in mouse bleomycin-injured lungs as well as the lowered levels of granulocytic enzymes in septic mice, the effects of MSCs on neutrophil invasion are likely to be cell-contact independent (29).
Finally, MSCs have been found to modulate a B cell-implicated autoimmune disease, a model of SLE (30). Reduced antibody production in non-differentiated B cells, arrested cell cycle in stimulated B cells, and reduced responses of B cells to chemotactic ligands are also observed when B cells are cocultured with MSCs (31, 32).
Moreover, cell-cell contact is claimed to enhance MSC inhibitory effects on B cells.
This result is found to be related to the interaction between programmed death 1 (PD-1) receptors and PD-1 ligands (33) (please see appendix for Fig. 1).
The indirect immunomodulatory effects of MSCs on T cells and neutrophils are also found. MSCs can modulate monocyte and DC functions, which ultimately result in the suppression of T cell proliferation. It is also reported that certain subpopulations of
MSC produce IL-1 receptor antagonist (IL-1RA). IL-1RA may decrease neutrophil recruitment by blocking the production of proinflammatory cytokines from
macrophages, which consequently diminishes the expression of chemokine and adhesion molecules in endothelial cells (29) (please see appendix for Fig. 1).
The immunoregulatory molecules of mesenchymal stem cells
Several candidates are identified to be essential for MSCs to perform
immunosuppressive functions (34). MSC-secreted indoleamine 2, 3-dioxygenase (IDO) is a rate limiting enzyme in kynurenine-dependent catabolism of tryptophan, which halts T cell proliferation (22, 35). MSC secretion of IDO and nitric oxide (NO) respectively promotes T cell anergy, instead of T cell apoptosis (36). Enhanced IDO, IL-10, or prostaglandin E2 (PGE2) secretion of MSCs leads to regulatory T cell promotion and proinflammatory T cell inhibition (22). MSC secretion of IDO, PGE2, HLA-G5, and transforming growth factor-beta 1 (TGF-b1) have all been reported as the mediators which are responsible for modulating cytotoxic functions in NK cells (37, 38) (please see appendix for Fig. 2). PGE2, IL-6, and granulocyte-macrophage colony- stimulating factor (GM-CSF) secreted by MSCs may increase the mannose receptor CD206 presenting and IL-10 secreting macrophages (39) (please see appendix for Fig.
3). IL-6 secreted by MSCs is suggested to regulate DC maturation by reducing the expression of MHC class II and costimulatory factors and the secretion of IL-12 in DCs (40) (please see appendix for Fig. 4). MSC secretion of IL-6 additionally has anti- apoptotic and reactive oxidative species downregulating effects on neutrophils (41) (please see appendix for Fig. 1). PD-1 ligand and vascular cell adhesion molecule
(VCAM)-1 produced by MSCs can not only support cell-cell contact but also promote the immunomodulatory functions of MSCs (42).
Hence, the immunomodulatory effects of MSCs are likely mediated through soluble factors and cell contact-dependent mechanisms (42). The conditioned medium obtained from human palatine tonsil-derived MSCs inhibits fibrosis in the skeletal muscle cells through the counteraction of IL-1b by IL-1 receptor antagonist (IL-1RA) (43). On the other hand, several reports on culture systems have shown that cell-cell contact is a key factor involved in the immunomodulatory effects of MSCs. MSCs are found to decrease the survival and proliferation of T cells through cell contact-
dependent mechanisms (44). Moreover, MSC-T cell contact is required for Treg induction (45). PGE2 and NO, despite their seemingly soluble properties, contribute to MSC-mediated suppressive functions in a cell-cell contact-dependent manner. High level production of PGE2 is initially dependent on a cell contact-dependent cross talk between MSCs and their target cells (46). MSC-secreted NO is able to inhibit T cell responses at its high concentration. However, the concentration of the active form of NO drops sharply within 100 µm because of its quick diffusive property. Therefore, MSCs require proximity for the effective inhibition of T cells through NO (36).
Heterogeneous mesenchymal stem cells
Although the immunosuppressive properties of BM-MSCs are well documented, studies on the immunomodulatory effects of MSCs produce conflicting data. Some studies indicate that MSCs are effective in suppressing excessive immune responses of GvHD and SLE in both animal models and human diseases. Whereas several other studies find that MSCs are unable to prolong GvHD remission in vivo even when they
do suppress T lymphocyte proliferation in vitro. In addition, combined administration of MSCs and an immunosuppressant, cyclosporine A, accelerates graft rejection instead of the opposite. These studies reveal that MSCs are immunosuppressive, but not under all conditions.
Single cell-derived MSC studies create different clones with varied properties (47).
Scientists therefore suppose that the applications of BM-MSCs are limited by their underlying heterogeneity (48, 49). Tissue stem cells are defined as undifferentiated cells being able to self-renew their own population and to guarantee the long-term production of differentiated cells. A single cell is apparently unable to fulfill these demands
simultaneously. In this sense, it appears that the functional definition of stem cells refers to a population of heterogeneous cells. It is not known whether the immunoregulatory properties are common to all the different MSC subpopulations (50). It is also evident that contrary to the anti-inflammation and anti-scarring formation potential (51-54), BM-MSCs possess nurturing, chemotactic, proinflammatory, and wound-healing hazards (55, 56). MSCs support immune cells by secreting growth factors and home to damaged tissues by responding to chemokines. When the hosts are invaded by bacteria, MSC-like cells inhibit bacterial growth in two ways. They attack it by producing
bactericidal factors directly, as well as enhancing proinflammatory responses from hosts (57-59). For instance, in the E. coli pneumonia mouse model, human BM-MSCs are suggested to impede the growth of bacteria partially by secreting antimicrobial peptide LL-37 (58). Paradoxically, the survival rates of both bacteria and host cells from bacterial-caused sepsis increase with immunomodulatory BM-MSCs; the host cells and bacteria must be saved or destroyed together. Therefore, MSCs are a double-edged sword.
Mesenchymal stem cell polarization
Inspired by the special role of the hematopoietic niche for the maintenance of hematopoietic stem cells, the interactions between stem cells and the signaling context to which they are exposed allow for an adaptive control of the balance between self- renewal and differentiation, and a decision of which lineage they are committing to.
This concept assumes that stem cell development is directly influenced by each cell’s local microenvironment (50). Indeed, evidences do support that homogenously acting
BM-MSCs can be polarized through microenvironmental licensing (36, 60).
In support of the “licensing” theory, first, MSCs are probably most effective when administered after the onset of inflammatory diseases. Several reports have indicated that after disease stabilization, the therapeutic efficacy of MSCs is not as apparent in mouse GvHD and EAE models. This is probably because the levels of inflammatory mediators in recipients with stabilized disease may be insufficient to elicit the
immunomodulatory effect of MSCs. Second, pretreatment of MSCs with inflammatory factors has been reported to enhance the potential therapeutic efficacy of MSCs (36).
Being the licensing factors, interferon (IFN)-g, tumor necrosis factor (TNF)-a, and IL-1 a/b are recommended. Different single cell-derived MSC populations display varied immunosuppressive abilities. However, such varieties disappear when the monoclonal cell lines are pretreated with Th1-type cytokines, IFN-g and TNF-a. IFN-g and TNF-a treatment changes the proclivity of the poorly immunosuppression-acting clone. Its enhanced immunosuppressive function is then revealed by negatively affecting concanavalin A-induced T cell proliferation and alleviating OVA-induced delayed-type hypersensitivity response (61).
MSCs have also been demonstrated to be primed by TLR ligands, leading to modulation of their differentiation, migration, proliferation, survival, and
immunosuppressive capacities (35, 62). Cultured mouse BM-MSCs express TLR1 to TLR8. TLR2 has been shown to be involved in the cardioprotective effects of mouse BM-MSCs in an ischemia/reperfusion injury model despite that the immunosuppressive properties of mouse BM-MSCs in response to TLR2 activation remain elusive (63).
TLR3 and TLR4 are among the most well-examined TLR subtypes of MSCs despite that their positions toward polarizing MSCs are varied in different experimental settings. TLR3 and TLR4 binding antagonizes MSC immunosuppressive activity by interfering with the Jagged1-Notch1 loop, while the other reports support the role of either TLR3 or TLR4 in promoting immunosuppressive MSCs (47).
Apparently, the role of proinflammatory cytokine and TLR ligand licensing in priming MSCs is more complex. MSCs acquire different functional properties depending on the level of stimulation. MSCs can be antigen presenting cells with upregulated MHCII if exposed to low-level IFNg (10 U/mL), whereas high-level IFN- g (100 U/mL) or TGF-b suppresses MHCII expression in MSCs (47). Low or high levels of specific TLR ligand exposure may also explain the different immunogenic characteristics of MSCs. Exposure of human MSCs to a high-level TLR3 ligand, poly (I:C) (1 µg/mL), polarizes MSCs into a IDO, TGF-b1, and IL-6-secreting phenotype which suppresses T cell proliferation and induces Treg generation. By contrast,
exposure of human MSCs to a low-level TLR4 ligand, LPS (10 ng/mL), leads MSCs to secret chemokines, such as macrophage inflammatory protein-1a (MIP-1a), IFN-g- induced protein-10 (IP-10), chemokine (C-C motif) ligand 5 (CCL5), and chemokine (C-X-C motif) ligand 9 (CXCL9), and consequently activate T lymphocytes (64).
stimulation. For instance, while IFN-g alone induces IDO upregulation in MSCs, IFN-g activates COX2 induction in MSCs synergistically when in combination with TNF-a (7).
Hence, the enhanced immunosuppressive properties of mouse BM-MSCs may be manipulated through their functional plasticity with specific priming factors. However, the outcome of MSC activation is dependent on the types of stimulating factors as well as the timing and levels of MSC exposure (65). In this study, we eventually selected a TLR2 ligand, Pam3CSK4, to be the priming factor for its better effects on enhancing immunosuppressive function in mouse BM-MSCs, and started establishing a priming protocol to optimize the beneficial effects of MSC treatment. OVA-induced asthmatic mice were used in this study to evaluate whether BM-MSCs preactivated with
Pam3CSK4 (Mpam) could alleviate asthma symptoms more effectively than would control BM-MSCs (Mctrl).
Although MSCs are certainly functionally plastic, the unclear molecular
mechanisms underlying MSC polarization into two distinct directions discourage their applications. By investigating the immunoregulatory mechanisms of modified BM- MSCs, not only would we be allowed to enlarge their therapeutic index with the
understanding of properly administered conditions, but also, we would be able to select the validated BM-MSCs ahead of the administered time to prevent unwanted results.
Since STAT3 promotes the production of immunosuppressive factors in many cell types (66, 67), we first investigated the role of STAT3 signaling in augmenting the
immunomodulatory properties of BM-MSCs through TLR2 priming. Overall, the significance of this study is that we established a valid protocol to enhance the therapeutic effectiveness of BM-MSCs in asthma.
CD4+CD25+Foxp3+ Treg
The immune system is able to destroy exogenous and endogenous pathogens.
Because its responses are potent enough to impair the integrity of the host, regulatory mechanisms must be essential to prevent the inappropriate harmful effects. One of the regulatory mechanisms is the clonal deletion that occurs in the thymus. Other key mechanisms in the maintenance of tolerance are the activation-induced T cell death and T cell anergy. In addition, a distinct subset of CD4+ T cells, regulatory T cells, Treg in short, is now widely perceived as the mediator of permanent suppression of
autoimmune responses in the periphery (68).
Although Treg cells comprise only 3-5% of the peripheral blood T cell pool, they are important for regulating the activation of the immune system. The most extensively studied Treg is characterized by expression of CD4, CD25, and the forkhead/winged- helix transcription factor Foxp3.
A subpopulation of CD4+ T cells that expresses the a-chain of IL-2 receptors (IL- 2Ra, CD25) plays an important function in peripheral tolerance by suppressing autoimmune disease in mice and humans (69). CD25-deficient mice develop massive lymphocyte accumulation and autoimmune disease. On the other hand, with age, IL-2- deficient mice develop fatal inflammatory bowel disease and lymphocyte proliferation.
In addition, the peripheral T cell compartments of IL-2-deficient mice lack a well- defined population of CD25+, which never exceeds 1-2% of the CD4+ T cells.
Moreover, adoptive transfer of CD4+CD25+ T cells, instead of being
hyperproliferative in mouse chimeras reconstituted with BM from CD25-defecient mice, rescues these chimera mice from chaotic lymphocyte accumulation. And the
T cells in these CD25-deficient mice is also achieved with adoptive transfer of CD4+CD25+ T cells from IL-2-deficient mice. This additionally indicates that IL-
2/CD25 signal is required for the generation and peripheral expansion of a population of regulatory CD4+ T cells essential for peripheral CD4+ T cell homeostasis. Therefore, it is not the elusion from thymic negative selection, or the lack of sensitization to
apoptosis in periphery that causes the autoimmunity in IL-2/IL-2R-deficient mice.
Instead, it is the generation and maintenance of CD4+CD25+ Treg cells that are mediated through IL-2/IL-2R interaction (70).
Although most CD4+CD25+ T cells have regulatory properties in naïve mice, the isolation of a pure Treg population according to the expression of CD25 is not enough in diseased mice and for in vitro activated CD4+ T cells. Because CD25 is upregulated in activated CD4+ T cells, new additional molecules that allow a more precise definition of Treg are still needed.
Polyendocrinopathy enteropathy X linked syndrome (IPEX) is a severe
multisystemic autoimmune disease whose onset usually occurs in the neonatal period. A frameshift mutation-resulting truncation of Foxp3 causes IPEX and its equivalent murine model, scurfy mice (71). Further, Foxp3 is demonstrated to be required for the development and the function of Treg: mice with either foxp3- or foxp3+ reconstituted BM are used to demonstrate that CD4+CD25+ regulatory T cell development requires Foxp3. Moreover, retroviral-mediated ectopic expression of Foxp3 is sufficient to activate the suppressive function in peripheral CD4+CD25- T cells (72). Although CD4+Foxp3+ cells are also characterized by a particular expression pattern of other cell surface molecules including cytotoxic T lymphocyte antigen 4 (CTLA-4),
glucocorticoid-induced tumor necrosis factor receptor family-related gene (GITR), etc., they are at present best identified by their expression of CD4, very high level of CD25,
and Foxp3. Furthermore, several genes preferentially expressed by CD4+CD25+ Treg cells such as CD25, CTLA-4, and GITR are shown to be directly controlled by Foxp3 (73). Foxp3 therefore allows the most precise recognition of CD4+CD25+ Treg.
Chapter II: Materials and methods
Materials
Animals
BALB/cByJNarl, C57BL/6JNarl, B6.129-Tlr2tm1kir/J
Buffers
DPBS (Sigma), HBSS (Sigma), Golden lysis buffer [137 mM NaCl + 20 mM Tris- base + 5 mM EDTA + 1mM EGTA + 10% glycerol + 1% Triton X-100 + protease inhibitor cocktail (Roche) + phosphatase inhibitor cocktail (Roche)], ACK lysis buffer (150 mM NH4Cl + 10 mM KHCO3 + 0.1 mM Na2EDTA in distilled water)
Culture medium
Mouse MSC culture medium (MesenCultTM proliferation kit + 2 mM L-glutamine + 250 ng/mL amphotericin B), Adipogenesis kit (MesenCult), Osteogenesis kit (Gibco), Chondrogenesis kit (Gibco), Mouse CD4+ lymphocytes/total splenocytes and MSC coculture medium [RPMI 1640 medium (Hyclone) + 10% FBS (Gibco) + 2 mM L- glutamine (Biological Industries) + PSA (Biological Industries) + 15 mM pH7.0 HEPES (Sigma)]
Reagents
Polyinosinic-polycytidylic acid [poly (I:C)] (Sigma), Lipopolysaccharide [LPS]
(Sigma), Recombinant flagellin from S. typhimurium (Rec FLA-ST) (Invivogen), Recombinant TNF-a (PeproTech), Recombinant IFN-g (PeproTech), Pam3CSK4
(Invivogen), S3I-201 (Cayman), L-NMMA (Sigma), IL-6 neutralizing antibody (BD Pharmingen Cat. 554398), IgG1 k isotype control antibody (BD Pharmingen Cat.
554682), Anti-mouse CD16/32 (BD Pharmingen Cat. 553142), Foxp3/Transcription Factor Staining Buffer Set (eBioscience Cat. 005523), FITC anti-mouse CD25
(BioLegend Cat. 102005), FITC Rat anti-mouse CD34 (BD Pharmingen Cat. 560238), FITC anti-mouse CD80 (eBioscience Cat. 11-0801), FITC anti-mouse CD86 (BD Pharmingen Cat. 553691), FITC anti-mouse CD90.2 (eBioscience Cat. 11-0903), FITC anti-mouse Sca-1 (BioLegend Cat. 108105), PerCP-Cyanine5 anti-mouse CD4 (BD Pharmingen Cat. 550954), PE-Cyanine5 anti-human/mouse B220 (eBioscience Cat. 15- 0452), APC anti-mouse CD11c (eBioscience Cat. 17-0114), APC anti-mouse CTLA-4 (BioLegend Cat. 106310), APC anti-mouse/rat Foxp3 (eBioscience Cat. 17-5773), PE anti-mouse/rat CD29 (BioLegend Cat. 102208), PE anti-mouse CD44 (BD Pharmingen Cat. 553134), PE anti-mouse CD73 (eBioscience Cat. 12-0731), PE anti-mouse CD105 (BioLegend Cat. 120408), Recommended isotype control antibodies for flow cytometry (BD Pharmingen or eBioscience), Mouse CD4+ T cell isolation kit (EasySep), Anti- CD3ε (BioLegend Cat. 100314), Anti-CD28 (BioLegend Cat. 102112), Mitomycin c (Sigma), 3H-thymidine (PerkinElmer), DNA polymerase kit (GoTaq), SYBR Green system (Bio-genesis), Ovalbumin (Sigma), Alum (Thermo Scientific), Anti-IgE, IgG1, IgG2a antibody for ELISA (BD Pharmingen), Antibodies for cytokine levels (R&D), TMB substrate (Clinical), b-actin antibody (Thermo Scientific Cat. MA5-15739), IkBa antibody (Cell Signaling Cat. 9247), NF-kB p65 antibody (Cell Signaling Cat.
8242), Phospho-NF-kB p65 (Ser 536) antibody (Cell Signaling Cat. 13346), STAT3 antibody (Flarebio Cat. CSB-PA004174), Phospho-STAT3 (Ser727) antibody (Cell Signaling Cat. 9134), NOS2 antibody (eBioscience Cat. 14-5920), Acetyl-b-
Pharmingen Cat. 558642), Anti-PE enrichment set (BD IMag), Type II collagenase (Worthington), Ficoll solution (GE).
Methods
Passage numbers 6 to 13 of characterized mouse BM-MSCs were used in this study. A TLR2 ligand, Pam3CSK4, was selected to generate the conditioned BM-MSCs.
[3H]-thymidine incorporation assays and the acute OVA-induced asthma murine model were used to examine the immunosuppressive activity of the conditioned BM-MSCs in vitro and in vivo, respectively. To investigate the signaling pathways of the Pam3CSK4
induction in BM-MSCs, STAT3 inhibitor, S3I-201 (Cayman), iNOS inhibitor, L-NG- monomethyl arginine citrate (L-NMMA) (Sigma), IL-6 neutralizing antibody (BD Pharmingen), or IgG1 k isotype control antibody (BD Pharmingen) was added in addition to the Pam3CSK4 treatment. On the other hand, to investigate the suppressive mechanisms of Mpam, the conditioned CD4+CD25+ T cells were generated by
coculturing with the conditioned BM-MSCs. [3H]-thymidine incorporation assays were further performed to examine the immunomodulatory effects of the conditioned
CD4+CD25+ T cells. The induction of CD4+CD25+Foxp3+ cells through conditioned BM-MSCs were elucidated in vitro and further investigated in vivo.
Additional details are described as follow:
Isolation and culture of MSCs from mouse bone marrow
BM cells were obtained from femurs and tibias of 4-5 weeks old female Balb/c mice, flushed with mouse MSC culture medium (MesenCult). MSCs were obtained by
expansion of the BM plastic-adherent cell fraction. Passage numbers 6 to 13 of BM- MSCs were used in this study.
Flow cytometry
Cells were processed according to the manufacturer’s protocols. Non-antigen- specific bindings of primary antibodies to the FcγII and FcγIII, and possibly FcγI receptors of cells were blocked by anti-mouse CD16/32 (BD Pharmingen Cat. 553142).
Non-hematopoietic surface markers CD29, stem cell antigen (Sca)-1, CD73, CD44, and CD105, and hematopoietic surface markers CD11b, B220, CD34, and CD11c were used to identify the non-hematopoietic MSCs. We performed intracellular staining using Foxp3/Transcription Factor Staining Buffer Set (eBioscience Cat. 005523). The flow cytometric acquisition was performed on a BD FACSCalibur device. The cells were acquired using forward scatter and side scatter and gated to exclude debris. The
cytometer then acquired 10,000 gated cell events. The fluorescence analysis was further conducted using BD CellQuestTM Pro software.
Differentiation assays
The cells were differentiated into adipocytes, osteocytes, or chondrocytes with lineage induction medium (MesenCult, Gibco, and Gibco, respectively). The
manufacturer’s instructions were followed to stain the differentiated cells. Adipocytes were detected by Oil Red O (Merck Millipore), osteocytes by Alizarin Red S (2 % Alizarin Red S in distilled water, adjusted to pH4.1-4.3 with NH4OH), and
chondrocytes by Alcian Blue (1 % Alcian Blue in 0.1 N HCl) staining.
Spleens from 5-8 weeks old female Balb/c mice were dissected. Red blood cells in total splenocytes were lysed using Ammonium-Chloride-Potassium (ACK) lysing buffer. CD4+ T cells in splenocytes were isolated by mouse CD4+ T cell isolation kit (EasySep) according to the manufacturer’s protocol. Total splenocytes or CD4+ T cells were respectively preactivated with soluble or coated 2 µg/mL anti-CD3ε (BioLegend) and 2 µg/mL anti-CD28 (BioLegend) for 24 h before being cocultured with mitomycin c (Sigma)-induced mitotically inactive BM-MSCs for 48 h. The cell ratio of total splenocytes or CD4+ T cells to BM-MSCs was 20 to 1. 1 µCi/mL 3H-thymidine
(PerkinElmer) was added into each well and the plate was incubated for 18 h. The cells were harvested and the thymidine incorporation was detected by a β-counter (MatrixTM 96). The same process was applied with the transwell insertions in the transwell assays.
In the transwell system, CD4+ T cells were seeded in the bottom well, while BM-MSCs were placed in the upper insertions.
Semi-quantitative and quantitative reverse transcription-polymerase chain reaction (RT-PCR)
In vitro conditioned primary cells were collected after 24-h-conditioned treatment.
Total RNA was isolated from primary cultured cells and lung tissues using Trizol®
reagent. The RNA templates were reversed into first strand cDNA for PCR analysis (Fermentas). A DNA polymerase kit (GoTaq) and a SYBR Green system (Bio-genesis) were respectively used in semi-quantitative and real-time RT-PCR. In real-time PCR, gene expression levels were normalized to the housekeeping gene, gapdh, and
calculated using the 2^(-ΔΔCt) method. Please see appendix for the primer sequences used in this study.
Conditioned BM-MSCs
After being seeded for 24 h, BM-MSCs were either nontreated as the Mctrl or treated with a TLR2 ligand, Pam3CSK4 (InvivoGen) as the Mpam; a TLR3 ligand, polyinosinic-polycytidylic acid [poly (I:C)] (Sigma) as the M3; a TLR4 ligand, lipopolysaccharide (LPS) (Sigma) as the M4; a TLR5 ligand, recombinant flagellin from S. typhimurium (Rec FLA-ST) (Invivogen) as the M5; a recombinant TNF-a as the Ma (PeproTech); and a recombinant IFN-g as the Mg (PeproTech). To investigate the signaling pathway of immunosuppression-prone Mpam, a STAT3 inhibitor, S3I-201 (Cayman), an iNOS inhibitor, L-NMMA (Sigma), an IL-6 neutralizing antibody, or an IgG1 k isotype control antibody was added in addition to the Pam3CSK4 treatment to make Ms&p, Ml&p, M6&p, and Mic&p, respectively (Fig. 1A).
Acute OVA-induced asthma murine model
After acclimating to the Laboratory Animal Center for 1 week, 6-week-old female Balb/c mice were divided into four groups: the negative control (PBS), positive control (OVA), Mctrl treatment, and Mpam treatment groups. Each group originally had 9 mice in one experimental setting. Except for the negative control group, which was sensitized and challenged with PBS, they were intraperitoneally sensitized by 200 µl of 50 µg ova (Sigma)/2 mg Alum (Thermo Scientific) at day 0, 14, and 21, and then were intranasally challenged by 40 µl of 100 µg OVA at day 28, 29, and 30. After the third sensitization, OVA-sensitized mice with 0.5-0.7 ELISA units of serum OVA-specific IgE levels and negative control mice with less than 0.015 ELISA units of serum OVA-specific IgE levels were included in the following procedures. The OVA-sensitized groups were randomized into three groups based on their serum OVA-specific IgE levels. The
Mpam respectively, day 28 through day 30. The animals were either sacrificed or subjected to an airway resistance experiment on day 31 (Fig. 1B).
The composition of immune cells in lung
The bronchoalveolar lavage fluid (BALF) was collected from each sacrificed and tracheotomized mouse with 1 ml Hanks’ Balanced Salts (HBSS) buffer (Sigma). After the BALF was centrifuged at 1500 rpm for 5 min, the supernatant was used to examine the cytokine levels, while the nucleated cell pellet was resuspended with HBSS
containing 2% FBS to determine the cellular composition. The differential cell counts collected by Thermo ScientificTM CytospinTM 4 Cytocentrifuge were discriminated with Liu’s stain. The labels of the samples were covered by the other colleague during the counting processes, and a flow cytometric method was used to confirm the results (74).
Measurement of OVA-specific IgE levels in serum and OVA-specific cytokine levels in supernatants
The enzyme-linked immunosorbent assay (ELISA) protocol was followed to detect OVA-specific IgE (BD Pharmingen) and OVA-specific cytokine levels (R&D). The supernatants of the in vitro conditioned primary cells and the ex vivo restimulated splenocytes were collected after 72-h-conditioned treatment and 72-h-stimulation, respectively. The HRP-catalyzed oxidation of TMB substrate (Clinical) was finally conducted to detect the presence of the targets. And the optical density (O.D.)
differences between 540 nm and 450 nm wavelengths of the converted TMB were read on a standard electroimmunoassay plate reader (Molecular Devices, VersaMax).
Noninvasive measurement of airway hyperresponsiveness
The response of airway to aerosolized acetyl-b-methylcholine chloride (Sigma) in mice was detected using whole body plethysmography, and was analyzed using
FinePointTM system. The response of airway (Penh) to aerosolized acetyl-b- methylcholine chloride was then divided by the response of airway to aerosolized Dulbecco’s phosphate-buffered saline (DPBS) in each mouse, yielding Penh (%).
Invasive measurement of airway resistance
Airway function was measured by detecting changes in lung resistance (RL) in response to increasing doses of aerosolized acetyl-β-methylcholine chloride (Sigma) in anesthetized and tracheotomized mice. The calculation of RL was yielded by subtracting transpulmonary pressure (0.45 cmH2O.s.ml−1) from respiratory flow measurement. Data were presented as average RL in the ratio of average RL after PBS nebulization.
Histological evaluation
Mouse lung tissues were dissected and then fixed in a 10 % formaldehyde solution (Merck Millipore). Samples were embedded in paraffin blocks and the histological sections were stained by hematoxylin and eosin (H&E).
Conditioned CD4+CD25+ T cells
CD4+ T cells were prestimulated with coated 2 µg/ml anti-CD3ε and 2 µg/ml anti- CD28 for 24 h before they were cocultured with mitomycin c-treated conditioned BM- MSCs in a 20:1 ratio. CD25+ cells were then isolated from these CD4+ T cells by anti- CD25 PE antibodies (BD Pharmingen Cat. 558642) and an anti-PE enrichment set (BD IMag) to make the Tctrl, Tpam, Ts&p, and Tl&p (Fig. 1C).
Griess assay
Mitotically inactive BM-MSCs were seeded as 1×105 cells overnight. The medium was then replaced by 250 µl of culture medium with treatments and incubated for 96 hours. The supernatants were assayed for nitric oxide by mixing 100 µl supernatant with 100 µl Griess reagent (Sigma) for 15 min at room temperature and were read on an electroimmunoassay plate reader at 550 nm. The concentration of nitrite in the culture supernatant was quantified by a standard curve of 100-0.003 µΜ NaNO2 concentrations (Sigma).
Western blot analysis
Protein samples were collected using Golden lysis buffer (137 mM NaCl, 20 mM Tris, 10 mM NaF, 5 mM EDTA, 1 mM EGTA, 10 % (v/v) glycerol, 1 % Triton X-100, protease and phosphatase inhibitor cocktails, pH 7.9), separated on 10 % SDS-PAGE gels, and transferred onto NC membranes (Whatman). After the membranes were incubated with primary and secondary antibodies, protein bands were visualized using chemiluminescence reagent (BIOSCIENCES). IκBα (Cell Signaling Cat. 4814), NF-κB p65 (Cell Signaling Cat. 3034), p- NF-κB p65 (ser536) (Abcam Cat. ab76302), STAT3 (Flarebio Cat. CSB-PA004174), p-STAT3 (ser727) (Cell Signaling Cat. 9134), iNOS (eBioscience Cat. 14-5920), and b-actin (ThermoFisher Cat. MA5-15739) were used as primary antibodies.
Pulmonary mononuclear cell preparation
The protocol established by D.L. Wiesner et al. was referenced (75). Lungs were excised and minced to generate pieces in HBSS (Sigma) + 1.3 mM EDTA (Sigma) solution for 30 min at 37°C with agitation. The solution was transferred to RPMI 1640
medium (Hyclone) supplemented with 10% FBS (Gibco) and 150 U/ml type II
collagenase (Worthington), and incubated for 1 h at 37°C with agitation. The cells were then grounded through a 70 µm filter, pelleted and resuspended with DPBS (Sigma).
Ficoll solution (GE) was finally used to separate mononuclear cells from the pulmonary homogenate.
Statistical analysis
Data were analyzed using GraphPad Prism v5 and are presented as mean and individual data points. To determine the significant variances between groups, Student’s t test was used between two groups, while one-way ANONA was used for multiple comparisons and Newman-Keuls Multiple Comparison Test was used for post hoc analysis. The statistical significance was set at p < 0.05.
Chapter III: Results
Characterization of mouse BM-MSCs
According to the criteria of the International Society for Cellular Therapy, mouse BM-MSCs have been characterized by their mesenchymal lineage surface markers, tri- lineage differentiation abilities, and immunosuppressive properties. The positive expression of CD29, stem cell antigen (Sca)-1, CD73, CD44, and CD105 and the negative expression of CD11b, B220, CD34, and CD11c exhibited no contamination of the stromal cells with hematopoietic cells (Fig. 2A). To establish that the stromal cells contained MSCs, the cells were cultured under a variety of conditions to assess their capacity to differentiate into mesodermal lineages. The isolated, purified stromal cells were able to differentiate into adipocytes, osteocytes, and chondrocytes when they were incubated with a specific induction medium, suggesting that these isolated stromal cultures certainly contained MSCs (Fig. 2B). We further ascertained the
immunosuppressive functions of cultured plate-selected BM-MSCs through [3H]- thymidine incorporation assays. The proliferative response of CD4+ T cells was
diminished when these cells were cocultured with BM-MSCs (Fig. 2C). In addition, the suppressive function of BM-MSCs was neither enhanced with lower CD4+ T to BM- MSC ratio (2:1), nor diminished with higher CD4+ T to BM-MSC ratio (200:1) (Fig.
2D).
Post-treatment as the more appropriate setting of BM-MSC treatment
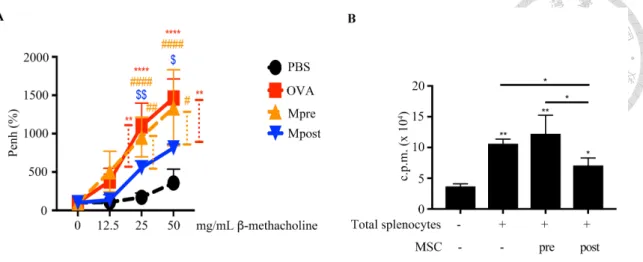
We next used an acute OVA-induced murine model to examine whether BM-MSCs would be a potential therapeutic strategy for asthma. First of all, we examined which time point we should intravenously administer BM-MSCs. Measured using whole body
plethysmography, the airway hyper-responsiveness of OVA-immunized mice was improved with BM-MSCs in the post-treatment setting (Mpost), but not in the pretreatment setting (Mpre) (Fig. 3A). In addition, less proliferative total splenocytes were observed in the Mpost group compared to those in the Mpre group from our ex vivo experiments (Fig. 3B).
Administering 5´105 of BM-MSCs for higher immunosuppressive activities in vivo After the immunosuppressive properties of BM-MSCs had been demonstrated in vivo, we subsequently examined with how many BM-MSCs would they become more effective. We intravenously administered 1´105 (Ma), 5´105 (Mb), or 1´106 (Mc)BM- MSCs, separately into OVA-challenged mice. Although BM-MSCs could improve airway hyper-responsiveness by the same extent no matter how many cells were
administered (Fig. 4A), different amounts of BM-MSCs decreased serum IgE level and Th2 cytokines in BALF by different degrees. Either 5´105 or 1´106 BM-MSCs would downregulate serum IgE level more effectively than that of 1´105 BM-MSCs (Fig. 4B).
Only 5´105 BM-MSCs however, decreased IL-4 and IL-5 secretion detected in BALF more effectively than that of 1´105 or 1´106 BM-MSCs (Fig. 4C). Hence, we decided that 5´105 BM-MSCs would be appropriate for this model.
TLR mRNA expression of mouse BM-MSCs
The TLR mRNA expression of BM-MSCs was examined using semi-quantitative RT-PCR to reveal the potential TLR ligand candidates. Specific primers were used to amplify sequences from TLR1 to TLR9, and the RAW264.7 cell line was used as a positive control (Please see appendix for Table 1). TLR3 and TLR4 are the most studied
MSC-priming TLR ligands, while the expression of TLR2, TLR4, and TLR5 was particularly high in the mouse BM-MSCs under our culture system (Fig. 5).
Pam3CSK4 as the comparatively effective stimulant for enhancing regulatory activities in BM-MSCs
TLR ligands can polarize MSCs according to other reports. We thus examined TLR2, 3, 4, 5 ligands for their possibilities to polarize MCSs or for their abundance in MSCs under our system. According to our results from [3H]-thymidine incorporation assays, Pam3CSK4 at 1 µg/mL for 72 h further enhanced the suppressive function of BM-MSCs whereas a TLR3 ligand, poly (I:C) at 10 µg/mL; a TLR4 ligand, LPS at 10 µg/mL; and a TLR5 ligand, Rec-FLA-ST at 100 ng/mL for 72 h did not (Fig 6A). After we analyzed the regulatory factors expressed in BM-MSCs with different stimulation, we observed using qPCR that 1 µg/mL Pam3CSK4 and 10 µg/mL LPS substantially enhanced inducible nitric oxide synthase (iNOS) in BM-MSCs (Fig. 6B). The
expression of cox-2, il-1ra, il-10, tsg-6, hgf, ido, and tgf-b in BM-MSCs, on the other hand, was affected through neither TLR ligands we examined (data not shown). At the protein level, Pam3CSK4 at 1 µg/mL and LPS at 10 µg/mL for 96 h enhanced NO secretion in BM-MSCs, in parallel with the results at the mRNA level (Fig. 6C).
In addition to TLR ligands, proinflammatory cytokines, such as TNF-a and IFN-g, are well-known microenvironmental factors to stimulate MSCs. According to our results from [3H]-thymidine incorporation assays, Pam3CSK4-activated BM-MSCs performed the most effective immunomodulatory function among the BM-MSCs activated with different stimulants (Fig. 7A). After we analyzed the regulatory factors expressed in BM-MSCs with different stimulation, we observed using qPCR that iNOS could be substantially enhanced through Pam3CSK4 in BM-MSCs, and ido could be
enhanced through 200 ng/mL IFN-g with or without 10 ng/mL TNF-a (Fig. 7B). In addition, IL-1RA expression detected by ELISA assays could not be induced through IFN-g without TNF-a in BM-MSCs (Fig. 7C).
Deciding to treat BM-MSCs with 1 µg/mL Pam3CSK4 for 96 h
We subsequently examined the ideal Pam3CSK4 stimulation protocol for BM- MSCs. In OVA-induced asthma model, compared to 5 µg/mL (Mpam5), 1 µg/mL Pam3CSK4-treated BM-MSCs (Mpam1) alleviated airway hyper-responsiveness (Fig.
8A) and diminished IL-5 secretion in BALF (Fig. 8C) more effectively. Further, compared to 1 µg/mL Pam3CSK4 treated for 24 h, treating BM-MSCs with 1 µg/mL Pam3CSK4 for 96 h was a more effective regimen to enhance the immunosuppressive function of BM-MSCs (Fig. 9).
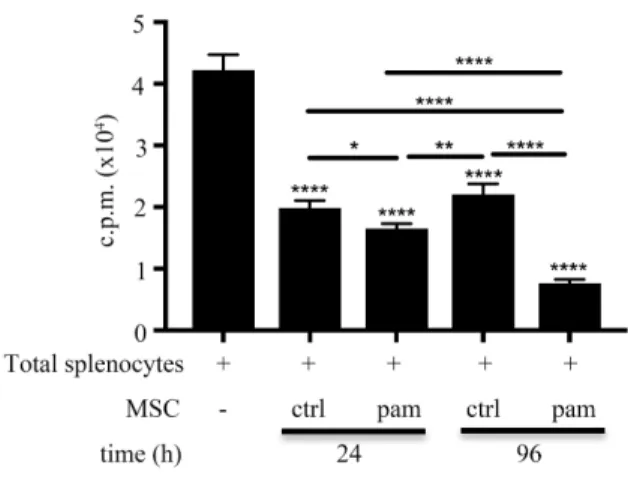
Enhanced immunosuppressive activities in Pam3CSK4-stimulated BM-MSCs To examine whether Pam3CSK4 could enhance the immunoregulatory function of BM-MSCs, we performed the [3H]-thymidine incorporation assays in vitro. The incorporated radioactivity increases proportionally with CD4+ T cell growth because CD4+ T cells use [3H]-thymidine as a nucleoside resource during mitosis. Consequently, CD4+ T cell proliferation was further inhibited significantly when they were cocultured with Mpam compared with when they were cocultured with Mctrl (Fig. 10).
Mpam as a more effective treatment in the asthma murine model
We further performed more experiments to compare the therapeutic effects of Mctrl and Mpam on OVA-induced allergic asthma murine model. The observations of
cytokine profiles on the BALF, eosinophil infiltration in BALF, airway resistance, and pathological changes in lung tissues are the main criteria for studying OVA-induced asthmatic mice. Based on our results, serum IgE level was significantly decreased by BM-MSCs, irrespective of the Pam3CSK4 treatment; however, serum IgG1/IgG2a level was further decreased in the Mpam-treated group, compared to that in the Mctrl-treated group (Fig. 11A). Th2-related IgG1 and Th1-related IgG2a represented whether
immune homeostasis was achieved in our asthma murine model. In our results, the Mpam treatment decreased the serum IgG1/IgG2a levels more effectively than those did the Mctrl treatment, suggesting that the imbalanced phenomenon in asthmatic mice was corrected more effectively with the Mpam than with that of the Mctrl treatment. IL-13 secretion of the OVA restimulated splenocytes was significantly downregulated only by Mpam treatment (Fig. 11B). The decreased serum IgG1/IgG2a ratio and IL-13 secretion of the restimulated splenocytes might suggest the advanced systematic therapeutic effects of Mpam on asthmatic mice. For the cytokine profiles and the infiltrated cell compositions from BALF, less Th2 cytokines (such as IL-4 and IL-5) were secreted (Fig. 12A) and less eosinophil was accumulated (Fig. 12B) in the Mpam-treated group compared with those in the Mctrl-treated group. In addition, the cell compositions in BALF were confirmed through flow cytometry (Fig. 12C). The results supported that the pulmonary inflammation attributed to OVA sensitization was alleviated more
effectively by Mpam than that by Mctrl. Evidenced by a plethysmograph with paralyzed and tracheotomized mice, the increased airway resistance in OVA-sensitized mice revealed that the recurrent inflammation made their airways susceptible to stimulants.
Our results demonstrated that the airway resistance of the OVA-induced asthmatic mice could be ameliorated further with the Mpam treatment than with the Mctrl treatment (Fig. 13A). The susceptible airways of asthmatic mice aggravated the pathological
effects of the recurrent inflammation, accordingly leading to the remodeled
parenchymal lung tissues as the disease progressed. Among the lung histopathological sections of all groups, both Mctrl and Mpam apparently diminished the inflammatory cell infiltration and the bronchial epithelial thickness (Fig. 13B). Moreover, mucin 5AC, which is one of the major components of the mucus secreted by goblet cells in the respiratory tracts, was also measured (76). Further downregulated muc5ac expression in the lung tissues was observed in the Mpam-treated group, compared to that in the Mctrl- treated group. This result implied that the Mpam treatment might be able to abolish mucin hypersecretion more effectively than did Mctrl treatment (Fig. 13C). In summary, the therapeutic effectiveness of BM-MSCs on asthmatic mice can be enhanced with Pam3CSK4 preactivation.
Pam3CSK4 changed the multipotent stem cell properties of BM-MSCs Mpam was found to form more colonies than Mctrl according to our colony formation assays (Fig 14A). In other words, Pam3CSK4 might help enhancing the self- renewal properties of BM-MSCs. In addition, the expression of the stem cell factor, Sca-1, was slightly enhanced in Mpam, compared to that in Mctrl (Fig 14B). Further, under the adipogenic condition, while Pam3CSK4 seemed to impair the adipogenic ability of normal mice-derived BM-MSCs, BM-MSCs isolated from TLR2 knockout mice were more prone to differentiate into adipocytes (Fig 14C).
STAT3 signals of the immunosuppression-prone BM-MSCs
To determine whether the STAT3 signaling pathway was involved in the Pam3CSK4-enhanced immunoregulatory function of BM-MSCs, we used S3I-201, a
assays revealed that the CD4+ T cell proliferative reduction affected by Mpam was reversed when the STAT3 activating process of Mpam was disturbed. This result supported that the STAT3 signaling pathway was involved in the enhanced regulatory function of BM-MSCs induced by Pam3CSK4. However, neither Pam3CSK4 nor S3I- 201 affected the immunosuppressive function of BM-MSCs in the transwell system (Fig. 15A). It additionally suggested that the enhanced immunosuppressive function of Mpam through STAT3 signaling was cell-cell contact-dependent.
The role of STAT3 activation in enhancing the regulatory function of Mpam was also suggested in vivo. Compared to the Mctrl-treated group, less eosinophils infiltrated to lungs in the Mpam-treated asthmatic mice. However, such diminished eosinophil infiltration in the Mpam-treated group was reversed in the Ms&p-treated group. It might imply that the enhanced ameliorating effect of Mpam in asthma murine model was abolished by STAT3 inhibition (Fig 15B).
Nitric oxide as the dominant immunoregulatory factor of Mpam To evaluate which regulatory molecules determined the enhanced
immunosuppressive effects of Mpam, we screened the well-known candidates through qPCR. The mRNA upregulation of iNOS, interleukin-1 receptor antagonist (il-1ra), tumor necrosis factor-stimulated gene sequence (tsg)-6, and hepatocyte growth factor (hgf) observed in the TLR2 ligand-activated BM-MSCs was downregulated with STAT3 inhibition (Fig. 15C). By contrast, the mRNA expression of cyclooxygenase (cox)-2, indoleamine 2,3-dioxygenase (ido), and transforming growth factor (tgf)-β in BM-MSCs was not influenced by TLR2/STAT3 signaling (data not shown). The high expression of iNOS and il-1ra in Mpam was further confirmed at the protein level. In coherence with the patterns at the mRNA level, the increased NO secretion detected
using Griess assays and the increased IL-1RA secretion detected using ELISA in Mpam were diminished in Ms&p (Fig. 15D, E). Therefore, because the NO and IL-1RA production in the Pam3CSK4-activated BM-MSCs were upregulated without STAT3 inhibition but were downregulated with STAT3 inhibition, STAT3 signaling pathway was involved in Pam3CSK4-induced NO and IL-1RA production. Although both iNOS and il-1ra were overexpressed in BM-MSCs through the TLR2/STAT3 signaling pathway, NO worked in a cell-cell contact-dependent manner, whereas IL-1RA worked as a soluble factor. The Pam3CSK4-enhanced immunosuppressive function in BM- MSCs and the diminished effects on Mpam through S3I-201 inhibition were observed using [3H]-thymidine incorporation assays without transwell inserts; however, neither TLR2 activation nor STAT3 inhibition induced any differences in the
immunosuppressive properties of BM-MSCs in the transwell system (Fig. 15A). The results suggested that cell contact dominated the enhanced regulatory effects of Mpam through the STAT3 pathway. Therefore, NO, rather than IL-1RA was the plausible Pam3CSK4-induced regulatory factor in BM-MSCs.
The mobility of BM-MSCs might be increased with Pam3CSK4 treatment
It is easy for MSCs to recognize and then migrate to inflammatory sites in hosts. It is therefore believed that MSCs are able to exert their proinflammatory/regulatory function because of such property. We thus examined whether Pam3CSK4 increased any chemokines and/or adhesion molecules in BM-MSCs. Among the chemoattractant which is induced in MSCs by TLR ligation, we found that the expression of ccl7 (detected by qPCR), mcp-1 (detected by qPCR), CCL3 (detected by ELISA), CCL5 (detected by ELISA), and VCAM-1 (detected by flow cytometry) was upregulated in