國立台東大學生命科學研究所 碩士論文
指導教授:彭仁君 博士
台東果園內之烏頭翁繁殖季生活史特性
Life History Traits of Taiwan Bulbul (Pycnonotus taivanus Styan) in Breeding Season in Taitung Orchard
研究生:蘇俊榮 撰
中華民國九十六年一月
致謝
終於畢業啦!真的要特別感謝我的指導老師 彭仁君博士,兩年多的日子以 來,從擬論文計畫到撰寫、審閱,著實在駑鈍的我身上浪費了不少寶貴的生命,
也被我折磨得快掛了!大恩大德,沒齒難忘!謹致上無限的感激與謝意!
此外,非常感謝國立中山大學生命科學系 張學文博士以及國立屏東科技大學 野生動物保育研究所 孫元勳博士兩位口試委員遠道而來,並給予寶貴的指教與建 議,讓我受益匪淺。
野外調查期間感謝官建維和龍田李萬枝先生的經驗指教、台東鳥會王克孝老 師的鼓勵、志奇測量器材的支援以及莊見凰、曾俊榮、洪文照、林盈助等各果園 主人的協助與配合,讓我能順利收集資料。感謝尚澄統計分析的協助和錦尚的製 圖。也感謝協助口試各項事宜的淳信、俊偉、惠嵐、育峰、旭昇、雅靖以及郁嫻。
感謝蔡見明校長的支持與包容,也感謝東宏、佳蓉、君軒的行政協助,振瑋 和寰文的程式協助,翟阿姨、瓊儀、淑莉以及其他族繁不及備載之同仁們的鼓勵。
感謝父母給我的支持與鼓勵,而岳母協助照料全家,讓我無後顧之憂,真是 感激不盡。大兒子洋洋除了偶爾陪我去調查外,還不時為我加油打氣,真是窩心。
最後,要感謝我的最愛,在這段時間除了給我適時的打氣與安慰外,還要承 受我陰晴不定的情緒,真的要謝謝妳的體諒與包容,尤其在小旗魚來報到時,還 丟下妳母子倆去調查。謹將這充滿血淚的論文獻給妳~我的老婆!
台東果園內之烏頭翁繁殖季生活史特性
蘇俊榮
國立台東大學生命科學研究所
中文摘要
烏頭翁(Pycnonotus taivanus Styan)屬於台灣特有之保育物種。本論 文探討台東縣烏頭翁在農業果園環境下繁殖期之生活史特性,自 2005 年 3 月至 2006 年 8 月在台東地區共調查 177 個巢的繁殖表現。
繁殖季從 3 月上旬到 8 月中旬。窩卵數在 2~4 個蛋之間,平均值 為 3.0 ± 0.5 個(n=177),其在不同果園間無顯著差異(X
2=3.97, P=0.137) 。繁殖季不同時期間蛋重平均為 2.79 ± 0.25 g,蛋長為 21.44 ± 1.02 mm,蛋寬為 15.82 ± 0.47 mm,三者都以繁殖季早期顯著大於晚 期。蛋之平均孵化率為 67.9%,平均每巢雛鳥數為 2.0 ± 1.3 隻
(n=177),平均離巢率為 31.0%,平均每巢離巢幼鳥數為 0.8 ± 1.2 隻 (n=135)。從蛋期至幼鳥成功離巢之繁殖成功率僅為 21.1%。平均孵卵 期為 10.6 ± 0.9 天 (n=60),平均育雛期為 10.2 ± 1.1 天(n=48)。
巢的構造特徵(巢內口徑、巢外口徑、巢高及平均巢深)在果園
間無顯著差異。但是巢位特徵中之巢位高度、樹高與巢位高之差、樹
冠位置指標、巢樹的分叉數、巢樹的截面積及在巢高度的植被密度這
六項則分別在不同果園間都有顯著差異,而其他七項巢位特徵在不同 果園間則無顯著差異。相對而言,龍田果園的果樹較為低矮、分叉數 多、主幹較細小且果樹密度也較小,烏頭翁在此園的巢位較靠近巢樹 主幹,巢位高度也較低。
本研究結果發現,平均孵化率在不同年間(X
2=3.02, P=0.082) 、不 同果園間(X
2=0.60, P=0.437)及繁殖季不同時期間(X
2=0.90, P=0.637)
都無顯著差異。然而,離巢率則在不同果園間(X
2=31.41, P<0.0001)
及繁殖季不同時期間(X
2=12.98, P=0.002)都有顯著差異,而在不同 年間雖無顯著差異,但都有隨著巢數的增加而降低的趨勢。
此外,每巢平均離巢幼鳥數與巢位高度之間有明顯且一致的正相 關變化趨勢,而因巢位高度與天敵掠食有重要關係。顯示巢位高度對 於繁殖成功率具有相當程度的影響。
台東縣烏頭翁在果園環境下之繁殖成功率僅約 20%,與其平均窩 卵數為 3 相比,僅不到 1 隻雛鳥可繁殖成功,此約為墾丁同種表現之 ㄧ半而已,其繁殖獲利與投資比是相當偏低的。果園雖然能讓較多烏 頭翁築巢與產卵順利完成,但在雛鳥期卻面臨更大之掠食與食物競爭 危機,其長期對烏頭翁族群動態之影響,是值得加以重視之課題。
關鍵字:烏頭翁、繁殖生態學、繁殖成功、巢位特性、生活史特性
Life History Traits of Taiwan Bulbul (Pycnonotus taivanus Styan) in Breeding Season in Taitung Orchard
Chun-Jung Su
Abstract
Taiwan bulbul (Pycnonotus taivanus Styan) is an endemic and conserved species in Taiwan. This study was conducted to investigate their life history traits during breeding season in Taitung orchard environment.
A total of 177 nests were monitored in the breeding seasons of 2005 and 2006. The breeding season stretched from March to mid-August. The average clutch size was 3.0 ± 0.5 eggs (range: 2-4 eggs), and they had no significant difference between orchards (P=0.137). The mean egg mass was 2.79 ± 0.25 g, the mean egg length was 21.44 ± 1.02 mm, and egg width was 15.82 ± 0.47 mm. Both egg mass and egg size in early-period were significantly higher than those in later-period of breeding seaeon.
The hatching success rate was 67.9%, and the average brood size per nest
was 2.0 ± 1.3 (n=177). The average number of young fledged per nest
was 0.8 ± 1.2 (n=135), and nestling success was 31.0%. Breeding success
was 21.1%. Incubation period was 10.6 ± 0.9 days (n=60), and nestling period stage was 10.2 ± 1.1 days (n=48). The characteristics of nest structure had no significant difference between orchards (P>0.05). Six of 13 nest-site characteristics were significantly different between orchards (P<0.05). The hatching success had no significant difference between years (P=0.082), between orchards (P=0.437), and between different periods of breeding season (P=0.637). However, the nestling success was decreasing with increasing nest number in different years, different orchards and different periods of breeding season. Besides, there was positive correlationship between average number of young fledged per nest and nest height above ground. We suggest that the main factors determining the reproductive success of Taiwan bulbul including food competitions and predation.
Keywords:Pycnonotus taivanus, Breeding ecology, Reproductive
success, Nest characteristic, Life history trait.
目 錄
中文摘要………..………i
英文摘要………iii
目錄………..…v
表目錄………...…………vii
圖目錄………..………viii
壹、前言………1
貳、材料及方法……….………...……4
一、研究樣區描述………..………...…4
二、研究方法……….…7
三、資料分析………...…10
参、結果………..11
一、繁殖季不同時期之巢數頻度分布………...…11
二、窩卵數…………...………12
三、鳥蛋重量與大小…...………12
四、孵化率、孵化雛鳥數、離巢率、離巢幼鳥數、繁殖成功率…..….14
五、發育期與其發育速率………...………18
六、造成孵卵失敗及雛鳥死亡的原因………...………20
七、巢構造………...………20
八、巢位特徵………...………21
肆、討論………..………22
一、烏頭翁對果樹之築巢利用………...……22
二、繁殖季………...…22
三、窩卵數………...………23
四、鳥蛋重量與大小………...……24
五、孵化率、離巢率、繁殖成功率與離巢成功幼鳥數…...……25
六、孵卵期與育雛期之發育速率………...…29
伍、結論………..………..………..30
陸、引用文獻………..……31
表目錄
表 1. 烏頭翁在不同年間、不同果園間、繁殖季不同時期及不同窩卵數 間之繁殖特性差異顯著性比較………52 表 2. 烏頭翁在兩不同年間、兩不同果園、繁殖季不同時期間及不同窩
卵數間的窩卵數、孵化數與離巢幼鳥數………...……...…53
表 3. 2005-2006 年烏頭翁蛋及雛鳥致死原因……….…54
表 4. 2005-2006 年烏頭翁在不同果園間的巢構造特徵表……….……55
表 5. 2005-2006 烏頭翁築巢的巢樹種類及數量統計表………….……55
表 6. 2005-2006 年烏頭翁在不同果園間的巢位特徵表………….……56
圖目錄
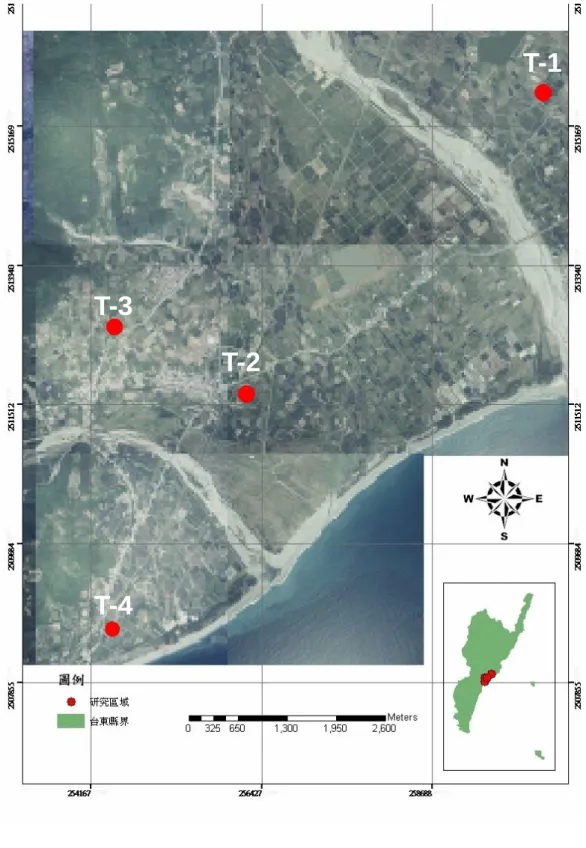
圖 1. 研究樣區地圖—台東市果園………...………37
圖 2. 研究樣區地圖—龍田果園………...………38
圖 3. 烏頭翁巢構造各部份示意圖………….………..………39
圖 4. 巢位特徵之樹冠位置指標示意圖……….……..………39
圖 5. 2005-2006 年台東縣烏頭翁繁殖季不同時期之孵卵期巢數頻度分 布……….40
圖 6. 2005-2006 年台東縣烏頭翁繁殖季不同時期之育雛期巢數頻度分 布……….40
圖 7. 2005-2006 年烏頭翁在不同年間之窩卵數巢數頻度分布……...…41
圖 8. 2005-2006 年烏頭翁在不同果園間之窩卵數巢數頻度分布…..…41
圖 9. 2005-2006 年烏頭翁在繁殖季不同時期間之窩卵數巢數頻度分 布……….42
圖 10. 2005-2006 年烏頭翁在不同年間蛋重、蛋長及蛋寬比較……...…43
圖 11. 2005-2006 年烏頭翁在不同果園間蛋重、蛋長及蛋寬比較…...…43
圖 12. 2005-2006 年烏頭翁在繁殖季不同時期蛋重、蛋長及蛋寬比較...44
圖 13. 2005-2006 年烏頭翁在不同窩卵數間蛋重、蛋長及蛋寬比較…...44
圖 14. 烏頭翁在不同年間的巢數頻度分布、孵化率、離巢率與繁殖成
功率變動趨勢比較……….……45 圖 15. 2005-2006 年烏頭翁在不同果園間的巢數頻度分布、孵化率、離 巢率與繁殖成功率變動趨勢比較……….……45 圖 16. 2005-2006 年烏頭翁在繁殖季不同時期的巢數頻度分布、孵化 率、離巢率與繁殖成功率變動趨勢比較………...……46 圖 17. 2005-2006 年不同窩卵數的巢數頻度分布、孵化率、離巢率與繁
殖成功率變動趨勢比較……….……46
圖 18. 2005-2006 年烏頭翁卵期與雛鳥期之發育速率在不同年間的比
較……….47
圖 19. 2005-2006 年烏頭翁卵期與雛鳥期之發育速率在不同果園間的比
較……….……47
圖 20. 2005-2006 年烏頭翁卵期與雛鳥期之發育速率在繁殖季不同時期
的比較……….……48
圖 21. 2005-2006 年烏頭翁卵期與雛鳥期之發育速率在不同窩卵數間的
比較……….…48
圖 22. 2005-2006 年 巢 中 不 同 雛 鳥 數 的 巢 數 頻 度 分 布 與 離 巢
率……….……49
圖 23. 2005-2006 年台東市果園烏頭翁育雛期雛鳥致死原因分析圖.…50
圖 24. 2005-2006 年龍田果園烏頭翁育雛期雛鳥致死原因分析圖….…50 圖 25. 2005-2006 年台東縣烏頭翁與各研究間平均蛋數、離巢數及巢位
高之比較……….……51
壹、前言
每一生物在自然界中都擁有其特定的生態棲位(niche),但隨著棲地環境條件 在時間與空間上的變化,鳥類對棲地的選擇策略也有所調整改變(Steele 1993)。
尤其是正值繁殖期的鳥類,會選擇有利於其繁殖成功的特定棲地,這可能是其對 棲地利用後,再經由演化過程作用所形成的結果(Hoover and Brittingham 1998)。
因為所選擇的棲地必須能提供維生和繁殖所必要的資源,這些資源包括食物 (Martin 1987)、覓食地點(Martin 1992)、巢位(Martin 1995 ; Maicas and Haeger 2004)、適宜的微氣候(Marshall 1959)與能避免天敵(Martin et al. 1993)、寄生 蟲(Moller 1990)及種內、外競爭的處所等(Hilden 1965)。若巢位的品質要求 取決於棲地的選擇,則有效、合適的巢位會是族群發展的重要因素之ㄧ(Steele 1993)。
生活史特性(life history traits)為演化天擇之結果,適當的生活史特性使物種 之發展延續得以維持(Stearns 1977 ; Linden and Moeller 1989)。鳥類繁殖期之生活 史特性包含產卵數、孵化率、離巢率、蛋大小與發育時間等,皆為瞭解鳥類繁殖 生態之重要測量變數,乃研究其族群變動及繁殖潛能的重要族群統計介量(Ricklefs 1969a ; Martin and Pingjun 1992 ; Robertson 1995 ; Sedinger and Flint 1995)。
天敵掠食是造成鳥類繁殖失敗主要的原因之ㄧ(Skutch 1949 ; Ricklefs 1969a ; Loiselle and Hoppes 1983 ; Martin 1992),而生殖巢位的高度(Martin 1993 ; Wilson and Cooper 1998 ; Reale and Blair 2005)、遮蔽度(Holway 1991)及環境條件
(Sockman 1997 ; Hilden 1965)等因素都會影響繁殖的成敗,因此生殖巢位的選擇
2003)。
築巢繁殖的鳥種數量在不同的地景植被型態間會有所差異(Best et al 1995 ; Smith and Bruun 2002)。在人為管理經營不同作物的農業耕地間,或在有人為管理 的耕地和無人管理的放牧地間,在其中進行繁殖的鳥種,其繁殖表現均有差異
(Rodenhouse and Best 1983 ; Galbraith 1988)。
烏頭翁(Pycnonotus taivanus Styan) 在分類上屬於燕雀目(Passeriformes)、鵯科
(Pycnonotidae)(Rand and Deignan 1960)。原被定為台灣特有種,乃全球受脅鳥 種之ㄧ,被國際自然保育聯盟紅皮書劃分為受脅 9 個等級中的易危等級,受脅的 主要理由為與白頭翁(Pycnonotus sinensis Hartert)的雜交問題(方 2005)。其外 部型態頭上至後頸黑色,頰和耳羽白色,下嘴基部有一橙黃色小斑點,有黑色粗 顎線。分布範圍為台灣東部花蓮縣天祥以南、台東縣和屏東縣的楓港以南地區
(Severinghaus and Blackshaw 1976;王等 1991;劉 1990、1991、2003、2005)。
其活動棲地很廣,從岩壁、次生林、農田以至庭園都可能出現(方 2005)。以往 之研究分別探討烏頭翁與白頭翁之間的形態、棲地利用、領域、群聚行為、生殖 行為(Hsu and Lin 1993;徐 1984;姚 1984)、鳴唱行為(張 1990;林 1992)及 演化親緣關係(許 1999)上的差異,但這些研究結果共同顯示兩者間似乎沒有明 顯的差異。對於烏頭翁與白頭翁兩者之雜交帶分布調查和雜交子代的生存與擴散
(劉 1990、1991、2005)之研究,則建立烏、白頭翁雜交與擴散現象的基本資料,
可提供擬定烏頭翁的保育措施時之參考。
台東縣位於台灣本島之東南部,其農業耕作面積 47,681 公頃約佔全縣面積的 13.6%,其中果樹種植面積為 13,267 公頃,約佔全縣農業耕作面積的 27.8%(台東
業環境的植被分布可算是普遍。
有關烏頭翁繁殖期生活史特性研究的報導,過去之研究區域只侷限在屏東恆 春半島地區的次生林或放牧地(徐 1984;Hsu and Lin 1993 ; Hsu and Lin 1997;劉 2005),台東縣烏頭翁多年來被觀察到在釋迦 (Annona squamosa Linn.) 園與荔枝 (Litchi chinensis Sonn.) 園中覓食並進行繁殖(李萬枝 個人通訊),但它們在果園 環境下之繁殖特性與族群增長潛力的表現,則尚無深入研究。
因此本研究以台東縣農業果園環境為研究區域,調查記錄烏頭翁繁殖期的窩 卵數、鳥蛋重量與大小、孵化率、孵化雛鳥數、離巢率及離巢幼鳥數等生活史特 性,並與前人在其它地區的研究結果進行比較,以探討在農業果園環境下影響烏 頭翁在繁殖期的生活史特性之可能原因。
貳、材料與方法
一、研究樣區描述
本研究依地區與集散程度,將 2005 年及 2006 年兩年調查所有樣區,分為台 東市果園及龍田果園,以下分別敘述各果園內不同樣區之環境條件。
(一)台東市果園
包含台東市行政區域內的康樂里、知本里和鄰近台東市的太麻里鄉三和 村(圖 1),各樣區描述如下:
T-1—台東市康樂里荔枝園,此樣區長 83 m、寬 54 m,面積約 4,482 平方 公尺的長方形。園內約有 322 棵荔枝樹,樹高約 2 m。此區塊東邊為番茄 (Lycopersicon esculentum Mill) 園,西邊為荖葉 (Piper betle L.) 園,南邊為釋 迦園,北邊為荖葉園。
T-2—台東市知本里兩個青脆枝 (Nothapodytes nimmoniana Mabb.) 園,為 兩個不規則形之園區,被產業道路及荒地相隔之,兩者相距約 30 公尺。一園 之邊長分別為 34 m、11 m、9 m、43 m、76 m 及 65 m,另一園為 112 m、43 m、
12 m、54 m、15 m、38 m 及 13 m,二果園之面積共約 5,931 平方公尺。兩園 內共約有 5,189 棵青脆枝樹,樹高約 5 m。此區塊東邊為荖葉園、芭樂 (Psidium guajava L.)、釋迦園,西邊為蔥 (Allium fistulosum L.) 園、鐵道、橋樑及住家,
南邊為荒地及荖葉園,北邊為玉米 (Zea mays Linn.) 田。
T-3—台東市知本里荔枝園,此梯形樣區的邊長分別為 29 m、84 m、119
m、131 m,面積約 5,624 平方公尺。園內約有 214 棵荔枝樹,樹高約 2 m。此 區塊東邊為香蕉 (Musa sapientum Linn.) 園、芭樂園及荒地,西邊為釋迦園及 荖葉園,南邊為荔枝園,北邊為釋迦園及酪梨 (Persea Americana Mill.) 園。
T-4—太麻里鄉三和村酪梨園,此長方形園區長約 102 m、寬約 26 m,面 積約 2,652 平方公尺。園內約有 61 棵酪梨樹,樹高約 4 m。此區塊東邊為荖 葉 園 , 西 邊 為 台 九 號 省 道 及 住 家 , 南 邊 為 荖 葉 園 及 扇 葉 蒲 葵 (Livistona chinensis R. Br. var.) 苗圃,北邊為農耕開墾地,雜植木瓜 (Carica papaya
Linn.)、蔥、芋頭 (Colocasia esculenta L. schott)、香蕉、小葉桑 (Morus australis Poir) 及椰子 (Cocos nucifera L.) 等。
在 2005 年及 2006 年 3 月至 8 月研究期間,據中央氣象局台東氣象站 2005 年及 2006 年的氣象資料顯示,月均溫在 19.5℃至 29.1℃之間,月累積降雨量 在 12.3 mm 至 360 mm 之間,相對溼度在 70.0%至 77.3%之間,日照累積時數 在 77.9 至 270.4 小時間,平均風速在 1.5 m/s 至 2.0 m/s 間。
(二)龍田果園
位台東縣鹿野鄉龍田村(圖 2),龍田村(22。51'N,121。07'E)北鄰 鹿野山,西、南臨鹿野溪,東靠卑南溪,海拔高度約 175 公尺,長約 3.7 公里,
寬約 2 公里,面積約 7.4 平方公里,主要為荔枝、釋迦、枇杷 (Eriobotrya japonica Lindl.)、鳳梨 (Ananas comosus Merr.)、香蕉及茶樹 (Camellia sinensis L.) 等作 物及荒地。
本樣區共有三個區塊,分別描述如下:
L-1—為釋迦、荔枝及枇杷園,此三果園緊鄰在一起,形成兩個長方形。
兩區分別為長 81 m、寬 63 m 及長 166 m、寬 160 m 面積共約 36,523 平方公尺。
釋迦園內約有 1,095 棵釋迦樹,樹高約 3 m;荔枝園內約有 105 棵荔枝樹,樹 高約 2 m;枇杷園內約有 266 棵枇杷樹,樹高約 2 m。此區塊東邊為茶園,西 邊為鳳梨園及香蕉園,南邊為荒地、鳳梨園及釋迦園,北邊為鳳梨園、枇杷 園、荒地及住家。
L-2—為釋迦園,為長方形之園區,其長約 92 m、寬約 55 m,面積約 5,060 平方公尺。釋迦園內約有 220 棵釋迦樹,樹高約 3 m。此區塊東邊為茶園,西 邊為桃子 (Prunus persica L.) 園及茶葉園,南邊為茶園及鳳梨園,北邊為鳳梨 園及茶園。
L-3—為住家旁果園,區內果樹混植,主要為荔枝、釋迦、香蕉、枇杷,
其他如檸檬 (Citrus limonum Burm.)、芭樂及柚子 (Citrus grandis Osbeck) 等數 棵。其長方形園區長約 93 m、寬約 76 m,面積約 5,375 平方公尺。園內約有 109 棵釋迦樹,樹高約 3 m;62 棵荔枝樹,樹高約 2 m;121 棵香蕉樹,樹高 約 4 m;20 棵枇杷樹,樹高約 2 m。此區塊東邊為住家,西邊為菠蘿蜜 (Artocarpus heterophyllus Lam.) 園、釋迦園及鳳梨園,南邊為荒地及鳳梨園,
北邊為香蕉園及荖葉園。
據行政院農業委員會茶葉改良場台東分場農業氣象站的資料提供,本樣 區在 2005 年及 2006 年 3 月至 8 月研究期間,月均溫在 17.5℃至 28.3℃之間,
月累積降雨量在 29 mm 至 435 mm 之間,相對溼度在 79.0%至 85.0%之間,日 照累積時數在 57.8 至 225.9 小時間,平均風速在 0.9 m/s 至 1.4 m/s 間。
二、研究方法
(一)繁殖季、窩卵數、雛鳥數及離巢幼鳥數
繁殖季的定義為一年中,開始於發現的第一個巢產下第一顆卵,至最後 一個巢的所有幼鳥離巢為止 (Cambell and Lack 1985)。若不確知實際產卵日 期,則透過持續觀察記錄所得確實的孵化日、離巢日再回推之 (李壽先 1989)。
本研究於 2005 年 3 月 26 日至 8 月 9 日及 2006 年 3 月 1 日至 8 月 13 日,
在週一至週五的下午 4 時 30 分至日落期間,週六、週日則從上午 8 時至日落 期間,約每間隔 1~2 日,在各研究樣區內,先進行尋巢,再進行探巢等調查 記錄。尋巢方式是先觀察烏頭翁是否有啣巢材進出樣區,或有無成對烏頭翁 在樣區中活動,再進入樣區中以密集徒步方式對每一棵樹搜尋。找到巢後先 對每一鳥巢予以編號,之後每間隔 1~2 日檢查一次,以確認巢內狀況,並記 錄其生殖階段為孵卵期或育雛期,以及各巢窩卵數、雛鳥數、離巢幼鳥數等 資料,直到幼鳥離巢或生殖失敗為止。為減少探巢時的干擾,探巢時間以不 超過 10 分鐘為原則。
(二)蛋長、蛋寬及蛋重
在發現鳥巢中的鳥蛋時即以日製 Mitutoyo 游標尺(準確至 0.01 mm)測 量蛋長及蛋寬,並以瑞士製 Jadever 電動秤(準確至 0.005 g)測量蛋重,並以 油性簽字筆分別標示之。
(三)繁殖成功率
繁殖成功(breeding success)分為卵的孵化及幼雛的離巢兩個階段。各巢 的孵化率(hatching rate)為順利孵化的雛鳥數/窩卵數;離巢率(nest success) 為成功離巢的幼鳥數/孵化時的雛鳥數。繁殖成功率(breeding success rate)為成 功離巢的幼鳥數/蛋數。
(四)巢的構造特徵及巢位特徵的測量
在幼鳥離巢後,再使用捲尺及皮尺(50 m),進行巢的構造特徵及巢位特 徵項目的測量工作,以減低對巢的干擾。測量項目包括:
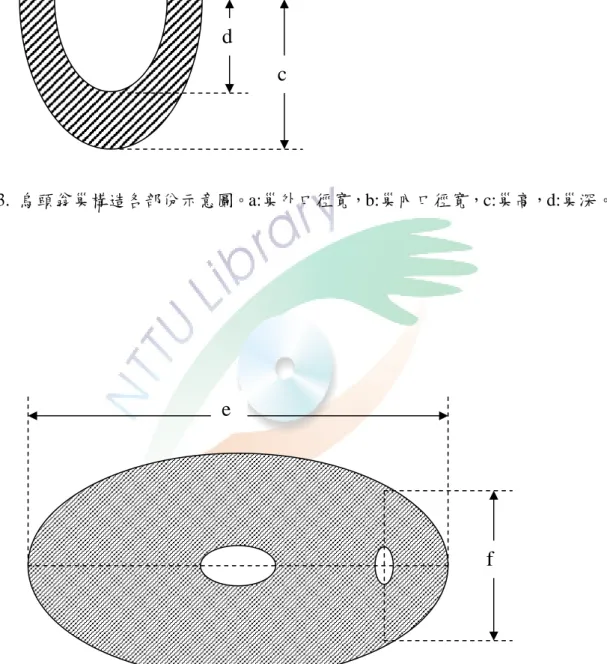
1. 巢的構造特徵(圖 3)
(1) 巢的外口徑寬:巢口外部最大直徑距離。
(2) 巢的內口徑寬:巢口內部最大直徑距離。
(3) 巢高:巢口至巢外底部之垂直距離。
(4) 巢深:巢口到巢內底部之垂直距離。
2. 巢位特徵
(1) 巢位高(Nest height above ground):巢外底部距地之垂直距離。
(2) 樹頂高度與巢位高度之差(Tree-nest difference):巢樹高度減去 巢位高,亦即巢底部至巢樹頂部之距離。
(3) 巢至最近葉之距離(Distance to nearest leaf):巢口到其上覆蓋葉 之最近距離。
(4) 樹冠位置指標(Nest position index to canopy):在樹冠的橫剖面 中,取通過巢與樹心的軸為長軸,通過巢與長軸垂直的軸為短 軸,短軸與長軸的比值即為指標值(圖 4)。
(5) 巢距主幹(Distance to main trunk):巢距主幹的水平距離。
(6) 巢距最近空間(Distance to nearest open space):巢距樹冠緣的水 平最近距離。
(7) 支持幹間距離(Distance between supporting branches):巢支持幹 的平均距離。計算方法為:巢口外周長減去支持幹直徑總和再 除以支持幹的數目。
(8) 支持巢的枝幹數目(Number of supporting branches)。 (9) 支持巢枝幹的截面積和(Supporting branches basal area)。 (10) 巢樹的截面積(Nest tree basal area):量取 130 公分高處巢樹的 直徑(胸徑),計算橫截面積。
(11) 巢樹的分叉數(Number of nest tree trunks):計算距地 130 公分 高處的巢樹分叉數。
(12) 在巢高度的植被密度(Tree density at nest height):樣區內的樹 木總數(棵)/樣區面積(公頃)。
(13) 距路或房舍之距離(Distance to road or building):量測巢與最 近道路或房舍之距離。
三、資料分析
資料分析以 SAS(SAS 1989)統計軟體進行。
(一)以卡方檢定(Chi-square Test)分別檢驗:
1. 繁殖季不同時期間及不同窩卵數間的巢數頻度分布之差異。
2. 兩不同年間、兩不同果園間及繁殖季不同時期間的窩卵數之差異。
3. 兩不同年間、兩不同果園間、繁殖季不同時期間及不同窩卵數間的孵 化率、離巢率與繁殖成功率之差異。
(二)以 Wilcoxon rank-sum Test 分別檢驗:
1. 兩不同年間及兩不同果園間的離巢幼鳥數、孵卵期與育雛期之差異。
(三)以 Kruskal-Wallis Test 分別檢驗:
1. 繁殖季不同時期間及不同窩卵數間的離巢幼鳥數、孵卵期與育雛期之 差異。
(四)以 t-test 分別檢驗:
1. 兩不同年間及兩不同果園間的蛋長、蛋寬與蛋重之差異。
2. 兩不同果園間巢的構造特徵及巢位特徵之差異。
(五)以單因子變異數分析(One-way ANOVA)分別檢驗:
参、結果
在 2005 年及 2006 年繁殖季分別共發現 54 及 184 個巢,若發現時巢中已是雛 鳥或雛鳥與蛋共存的狀態,以及在未確定窩卵數前鳥蛋因為不明原因而不見,這 些資料都扣除不納入分析。因此,使用做為資料分析的巢數,在 2005 年有 37 巢,
2006 年有 140 巢,合計 177 巢。各果園在兩年繁殖季所觀察的巢數分別為:台東 市果園 12 巢及 50 巢,合計 62 巢;龍田果園 25 巢及 90 巢,合計 115 巢。
一、繁殖季不同時期之巢數頻度分布
烏頭翁的繁殖季從 3 月上旬到 8 月中旬約半年。本研究將繁殖季分為 3 個時 期,分別為繁殖季前期為 3-4 月,繁殖季中期為 5-6 月,繁殖季晚期為 7-8 月。
2005 年孵卵期的巢數,以在繁殖季中期之 21 巢最多,佔 56.8%(圖 5);同年 育雛期的巢數,也以在繁殖季中期 19 巢最多,佔 67.9%(圖 6)。
2006 年孵卵期的巢數,以在繁殖季中期之 95 巢最多,佔 67.9%(圖 5);同年 育雛期的巢數也以在繁殖季中期 68 巢最多,佔 65.4%(圖 6)。
將兩年資料合併,孵卵期的巢數頻度在繁殖季各時期間有顯著差異(Chi-square Test X2=82.81, d.f.=2, P<0.0001)(表 1),以繁殖季中期的 116 巢最多,其次為早期 的 33 巢和晚期的 28 巢。
二、窩卵數
在兩年繁殖季 177 巢中,共發現 527 顆蛋。全部窩卵數在 2~4 個蛋之間。其 中,窩卵數為 2 個卵的有 20 巢,佔 11.3%;3 個卵的有 141 巢,佔 79.7%;4 個卵 的有 16 巢,佔 9.0%,平均窩卵數為 3.0 ± 0.5 個。
窩卵數在兩年間無顯著差異(Chi-square Test X2=0.57, d.f.=1, P=0.753)(表 1),
兩年平均窩卵數都為 3.0 個(表 2)。
窩卵數在兩果園間亦無顯著差異(Chi-square Test X2=3.97, d.f.=1, P=0.137)(表 1),但以龍田果園每窩平均 3.0 ± 0.5 個(n=115) 稍高於台東市果園之 2.9 ± 0.4 個(n=62) (表 2)。
窩卵數在繁殖季不同時期之間具有顯著差異(Chi-square Test X2=14.47, d.f.=2, P=0.006)(表 1),以繁殖季晚期平均窩卵數 3.0 ± 0.3 個(n=28)顯著高於早期的 2.7 ± 0.5 個(n=33) (表 2)。
在不同年間(圖 7)、不同果園間(圖 8)及繁殖季不同時期間(圖 9)窩卵數 的巢數頻度分布均以每窩 3 卵的巢數較多。
三、鳥蛋重量與大小
在兩年繁殖季所發現的 524 顆蛋,平均蛋重為 2.79 ± 0.25 g,平均蛋長為 21.44
± 1.02 mm,平均蛋寬為 15.82 ± 0.47 mm。
(一)蛋重
蛋重在不同年間有顯著差異(t-test t=-3.21,P=0.001)(表 1),以 2006 年 的 2.81 ± 0.25 g (n=413)較重;2005 年的 2.72 ± 0.25 g(n=111)較輕(圖 10)。
蛋重在不同果園間亦有顯著差異(t-test t=-2.21,P=0.027)(表 1),以台東 市果園的 2.82 ± 0.24 g(n=181)較重;龍田果園組的 2.77 ± 0.26 g(n=343)
較輕(圖 11)。
蛋重在繁殖季不同時期間也有顯著差異(One-way ANOVA F2,521=14.85,
P<0.0001)(表 1),以繁殖季早期的 2.86 ± 0.21 g (n=88)較重;繁殖季晚期 的 2.67 ± 0.22 g(n=85)較輕(圖 12)。
蛋重在不同窩卵數間則無顯著差異(One-way ANOVA F2,521=0.79 , P=0.456)(表 1),但以窩卵數 1 窩 2 卵的 2.84 ± 0.29 g(n=40)較重;1 窩 3 卵的 2.78 ± 0.25 g(n=420)和 1 窩 4 卵的 2.79 ± 0.23 g(n=64)較輕(圖 13)。
(二)蛋長
蛋長在不同年間無顯著差異(t-test t=-0.67,P=0.501)(表 1),2006 年為 21.45 ± 1.03 mm(n=413);2005 年為 21.38 ± 0.96 mm(n=111)(圖 10)。
蛋長在不同果園間無顯著差異(t-test t=-1.73,P=0.085)(表 1),台東市 果園為 21.54 ± 0.97 mm(n=181);龍田果園為 21.38 ± 1.03 mm(n=343)(圖 11)。
蛋長在繁殖季不同時期間有顯著差異(One-way ANOVA F2,521=19.6,
P<0.0001)(表 1),以繁殖季早期的 21.91 ± 0.95 mm (n=88)較長;繁殖季晚
期的 20.98 ± 0.86 mm(n=85)較短(圖 12)。
蛋 長 在 不 同 窩 卵 數 間 有 顯 著 差 異 ( One-way ANOVA F2,521=11.11 , P<0.0001)(表 1),以窩卵數 1 窩 2 卵的 22.14 ± 1.29 mm(n=40)較長;1 窩 4 卵的 21.28 ± 0.89 mm(n=64)較短(圖 13)。
(三)蛋寬
蛋寬在不同年間無顯著差異(t-test t=0.88,P=0.382)(表 1),2005 年為 15.85 ± 0.45 mm(n=111);2006 年為 15.81 ± 0.48 mm(n=413)(圖 10)。
蛋寬在不同果園間亦無顯著差異(t-test t=-1.22,P=0.224)(表 1),台東 市果園為 15.85 ± 0.44 mm(n=181);龍田果園為 15.80 ± 0.49 mm(n=343)(圖 11)。
蛋寬在繁殖季不同時期間則有顯著差異(One-way ANOVA F2,521=3.66,
P=0.027)(表 1),以繁殖季早期的 15.89 ± 0.39 mmg(n=88)較長;繁殖季晚 期的 15.70 ± 0.46 mm(n=85)較短(圖 12)。
蛋寬在不同窩卵數間無顯著差異(One-way ANOVA F2,521=0.77,P=0.465)
(表 1),窩卵數 1 窩 4 卵為 15.86 ± 0.48 mm(n=64);1 窩 2 卵為 15.75 ± 0.46 mm(n=40)(圖 13)。
四、孵化率、孵化雛鳥數、離巢率、離巢幼鳥數及繁殖成功率
(一)孵化率
在兩年繁殖季共記錄有 527 顆蛋,其中,成功孵出 358 隻雛鳥,孵化率 為 67.9%。
孵化率在不同年間無顯著差異(Chi-square Test X2=3.02, d.f.=1, P=0.082)
(表 1),以 2005 年的 74.8%(n=37)稍高於 2006 年的 66.1%(n=140)(圖 14)。 孵化率在不同果園間也無顯著差異(Chi-square Test X2=0.60, d.f.=1, P=0.437)(表 1),以龍田果園的 69.1%(n=115)較高;台東市果園的 65.8%
(n=62)較低(圖 15)。
孵化率在繁殖季不同時期之間也無顯著差異(Chi-square Test X2=0.90, d.f.=2, P=0.637)(表 1),以繁殖季早期的 68.9%(n=33)稍高於繁殖季晚期的 63.5%(n=28)(圖 16)。
孵化率在不同窩卵數間則有顯著差異(Chi-square Test X2=11.36, d.f.=2, P=0.003)(表 1),以每窩 4 卵的 75.0%(n=64)較高;每窩 2 卵的 45.0%(n=40)
較低(圖 17)。
(二)雛鳥數
在兩年繁殖季 177 巢中共計有 358 隻雛鳥完成發育,平均每巢雛鳥數為 2.0 ± 1.3 隻。
平均每巢雛鳥數在不同年間以 2005 年 2.2 ± 1.3 隻(n=37)較多,2006 年為 2.0 ± 1.3 隻(n=140)較少(表 2)。
平均每巢雛鳥數在不同果園間以龍田果園的 2.1 ± 1.3 隻(n=115)稍多於 台東市果園組的 1.9 ± 1.3 隻(n=62)(表 2)。
平均每巢雛鳥數在繁殖季不同時期以繁殖季中期的 2.1 ± 1.3 隻(n=116)
較多於繁殖季早期的 1.9 ± 1.2 隻(n=33)(表 2)。
平均每巢雛鳥數在不同窩卵數間以 1 窩 4 卵的 3.0 ± 1.6 隻(n=16)較多;
1 窩 2 卵的 0.9 ± 1.0 隻(n=20)較少(表 2)。
(三)離巢率
在兩年繁殖季合計 358 隻雛鳥,成功離巢幼鳥數為 111 隻,總離巢率為 31.0%。
平均離巢率在不同年間無顯著差異(Chi-square Test X2=2.03, d.f.=1, P=0.154)(表 1),但以 2005 年的 37.4%(n=83)較高於 2006 年的 29.1%(n=275)
(圖 14)。
平均離巢率在不同果園間有顯著差異(Chi-square Test X2=31.41, d.f.=1, P<0.0001)(表 1),以台東市果園的 50.4% (n=119)較高;龍田果園組的 21.3%
(n=239)較低(圖 15)。
平均離巢率在繁殖季不同時期亦具有顯著差異(Chi-square Test X2=12.98, d.f.=2, P=0.002)(表 1),繁殖季早期的 40.3%(n=62)高於繁殖季晚期的 11.1%
(n=54)(圖 16)。
平均離巢率在不同窩卵數間則無顯著差異(Chi-square Test X2=3.89, d.f.=2, P=0.143)(表 1),以 1 窩 2 卵的 33.3%(n=18)高於 1 窩 4 卵的 18.8%(n=48)
(圖 17)。
(四)離巢幼鳥數
在兩年繁殖季 135 巢中共 111 隻幼鳥成功離巢,其平均每巢離巢幼鳥數為 0.8 ± 1.2 隻。
平均每巢離巢幼鳥數在不同年間無顯著差異(Wilcoxon rank-sum Test Z=0.91, d.f.=1, P=0.361)(表 1),以 2005 年的 1.0 ± 1.4 隻(n=30)稍高於 2006 年的 0.8 ± 1.2 隻(n=105)(表 2)。
平均每巢離巢幼鳥數在不同果園間有顯著差異(Wilcoxon rank-sum Test Z=3.78, d.f.=1, P=0.0002)(表 1),以台東市果園的 1.4 ± 1.3 隻(n=44)高於 龍田果園組的 0.6 ± 1.1 隻(n=91)(表 2)。
平均每巢離巢幼鳥數在繁殖季不同時期無顯著差異(Kruskal-Wallis Test X2=5.66, d.f.=2, P=0.059)(表 1),但以繁殖季早期的 1.0 ± 1.3 隻(n=25)高於 繁殖季晚期的 0.3 ± 0.9 隻(n=20)較少(表 2)。
平均每巢離巢幼鳥數在不同窩卵數間亦無顯著差異(Kruskal-Wallis Test X2=0.41, d.f.=2, P=0.817)(表 1),以 1 窩 3 卵的 0.9 ± 1.2 隻(n=111)稍高於 1 窩 2 卵的 0.6 ± 0.8 隻(n=11)(表 2)。
(五)繁殖成功率
在兩年繁殖季合計 527 顆蛋,成功離巢幼鳥數為 111 隻,繁殖成功率(總 離巢幼鳥數/總蛋數)為 21.1%。
繁殖成功率在不同年間則無顯著差異(Chi-square Test X2=0.67, d.f.=1, P=0.414)(表 1),但以 2005 年的 34.4%(n=29)較高於 2006 年的 25.4%(n=105)
(圖 14)。
繁殖成功率在不同果園間有顯著差異(Chi-square Test X2=10.60, d.f.=1, P=0.001)(表 1),以台東市果園的 45.5%(n=44)較高於龍田果園組的 18.7%
(n=90)(圖 15)。
繁殖成功率在繁殖季不同時期亦有顯著差異(Chi-square Test X2=6.85, d.f.=2, P=0.033)(表 1),以繁殖季早期的 36.8%(n=25)較高,繁殖季晚期的 9.8%(n=20)較低(圖 16)。
繁殖成功率在不同窩卵數間則無顯著差異(Chi-square Test X2=0.66, d.f.=2, P=0.719)(表 1),以 1 窩 3 卵的 28.8%(n=111)較高,1 窩 4 卵的 17.3%(n=13)
較低(圖 17)。
五、發育期與其發育速率
(一)孵卵期
在兩年繁殖季的孵卵期約需 9~13 天,平均為 10.6 ± 0.9 天(n=60)。
平均孵卵期在不同年間無顯著差異(Wilcoxon rank-sum Test Z=1.49, d.f.=1, P=0.137)(表 1),以 2005 年的 11.3 ± 1.2 天(n=4)稍長於 2006 年的 10.6 ± 0.9 天(n=56)(圖 18)。
平均孵卵期在不同果園間無顯著差異(Wilcoxon rank-sum Test Z=-0.92, d.f.=1, P=0.360)(表 1),龍田果園為 10.7 ± 0.9 天(n=36);台東市果園為 10.5
± 0.9 天(n=24)(圖 19)。
平 均 孵 卵 期 在 繁 殖 季 不 同 時 期 間 則 有 顯 著 差 異 (Kruskal-Wallis Test X2=7.34, d.f.=2, P=0.025)(表 1),以繁殖季早期的 11.3 ± 0.8 天(n=12)長於 繁殖季中期的 10.5 ± 0.8 天(n=39)(圖 20)。
平均孵卵期在不同窩卵數間亦有顯著差異(Kruskal-Wallis Test X2=1.00, d.f.=2, P=0.0002)(表 1),以窩卵數 1 窩 2 卵的 11.9 ± 0.4 天(n=6)稍長於 1 窩 4 卵的 9.8 ± 0.3 天(n=5)(圖 21)。
(二)育雛期
在兩年繁殖季的育雛期約需 8~12 天,平均為 10.2 ± 1.1 天(n=48)。
平均育雛期在不同年間無顯著差異(Wilcoxon rank-sum Test Z=-0.52, d.f.=1, P=0.601)(表 1),2006 年為 10.3 ± 1.1 天(n=36);2005 年為 10.0 ± 1.2 天(n=12)(圖 18)。
平均育雛期在不同果園間無顯著差異(Wilcoxon rank-sum Test Z=-0.59, d.f.=1, P=0.557)(表 1),台東市果園為 10.4 ± 1.1 天(n=26);龍田果園為 10.1
± 1.1 天(n=22)(圖 19)。
平 均 育 雛 期 在 繁 殖 季 不 同 時 期 間 亦 無 顯 著 差 異 (Kruskal-Wallis Test X2=1.64, d.f.=2, P=0.440)(表 1),以繁殖季晚期的 11.0 ± 1.4 天(n=2)稍長於 繁殖季中期的 10.1 ± 1.1 天(n=35)(圖 20)。
平均育雛期在不同窩卵數間也無顯著差異(Kruskal-Wallis Test X2=3.71, d.f.=2, P=0.156)(表 1),以窩卵數 1 窩 4 卵的 11.1 ± 1.2 天(n=4)稍長於窩 卵數 1 窩 2 卵的 9.5 ± 1.0 天(n=4)(圖 21)。
六、造成孵卵失敗及雛鳥死亡的原因
在兩年繁殖季所觀察的烏頭翁巢中,合計共有 527 顆蛋,其中有 169 顆蛋在 孵卵過程中死亡或不明原因不見,佔總產卵數的 32.1%;而孵出的雛鳥總數為 358 隻,其中有 247 隻在離巢前死亡或不明原因不見,佔所孵化雛鳥數的 69.0%。
其中,在 2005 年有 28 個蛋在孵卵過程中死亡或不明原因不見,佔該年總產 卵數(111 個)的 25.2%;有 52 隻雛鳥在離巢前死亡或不明原因不見,佔該年所 孵化雛鳥數(83 隻)的 62.7%。2006 年有 141 個蛋在孵卵過程中死亡或不明原因 不見,佔該年總產卵數(416 個)的 33.9%;有 195 隻雛鳥在離巢前死亡或不明原 因不見,佔該年所孵化雛鳥數的 70.9%。2005 年及 2006 年二個繁殖季烏頭翁孵卵 失敗和雛鳥死亡及不明原因不見數合計 416 例,不明原因不見佔 79.8%、未孵化或 在巢中死亡佔 8.7%、人為因素造成佔 7.2%、天候因素造成佔 3.1%及被掠食佔 1.2%
(表 3)。
七、巢構造
烏頭翁的巢約為碗形,在巢的構造測量項目方面,巢內口徑 6.9 ± 0.8 ㎝
(n=137)、巢外口徑 11.9 ± 1.2 ㎝(n=137)、巢高 10.0 ± 1.8 ㎝(n=137)及巢深 5.0 ± 0.7 ㎝(n=137)。巢構造各測量項目以 t-test 進行檢驗,在果園間均無顯著差 異(表 4),以台東市果園的巢內口徑 7.0 ± 0.8 ㎝(n=44)、巢外口徑 12.2 ± 1.5 ㎝
(n=44)及巢高 10.1 ± 1.6 ㎝(n=44)大於龍田果園,巢深以龍田果園的 5.1 ± 0.7
八、巢位特徵
在兩年繁殖季烏頭翁的 177 個巢中有 67 巢在荔枝樹上發現,40 巢在釋迦樹 上,26 巢在酪梨樹上,23 巢在枇杷樹上,19 巢在青脆枝上,在龍船花 (Clerodendrum paniculatum L.) 及蒲桃 (Syzygium jambos (L.) Alston) 上則各僅 1 巢,其中荔枝、
酪梨、釋迦、枇杷等果樹佔 88.2%(表 5)。在 135 個巢中巢樹最高為 4.7 公尺,
最低為 1.6 公尺,平均樹高為 2.7 ± 0.7 公尺(n=135)。平均巢位高度為 170.3 ± 53.7 cm(n=135),樹高與巢位高之差為 102.5 ± 38.0 cm (n=135),巢至最近葉之距 離 1.9 ± 2.6 cm(n=135),樹冠位置指標 77.7 ± 24.5(n=135),巢距主幹 91.1 ± 68.1 cm(n=135),巢距最近空間 50.1 ± 23.7 cm(n=135),支持幹間距離 12.3 ± 4.9 cm
(n=135),支持巢的枝幹數目 3.0 ± 0.8 根(n=135),支持巢枝幹的截面積和 4.1
± 3.3 cm2(n=135),巢樹的分叉數 13.1 ± 11.5 根(n=135),巢樹的截面積 30.9 ± 51.4 cm2(n=135),在巢高度的植被密度 1036.7 ± 2193.5 棵/公頃(n=135),距 路或房舍之距離 41.6 ± 24.8 m(n=135)(表 6)。
對巢位特徵的 13 個測量項目進行 t-test 檢驗,巢位高度、樹高與巢位高之差、
樹冠位置指標、巢樹的分叉數、巢樹的截面積及在巢高度的植被密度這六項在果 園間有顯著差異,其他七項則無顯著差異(表 6)。
肆、討論
一、烏頭翁對果樹之築巢利用
Hsu and Lin(1997)在屏東墾丁青蛙石海岸次生林進行烏頭翁繁殖研究,其 主要植被為木麻黃 (Casuarina equisetifolia L.)、瓊崖海棠 (Calophylium inophyllum L.) 、 相 思 樹 (Acacia confuse Merr.) 、 馬 纓 丹 (Lantana camara L.) 和 長 穗 木 (Stachytarpheta jamaicensis (L.) Vahl.) 等,所調查記錄的 13 種烏頭翁巢樹中,有高 達 75%都為瓊崖海棠;但劉(2005)在墾丁國家公園海岸林帶的植物群落所調查 的記錄則有 26 種以上的巢樹中,其中芒果樹 (Mangifera indica L.)、銀葉樹 (Heritiera littoralis Dryand.)、福木 (Garcinia suubelliptica Merr.)、荔枝樹、榕樹 (Ficus microcarpa L.f.)、瓊崖海棠、枯里珍 (Antidesma pentandrum Merr.)、黃槿 (Hibiscus tiliaceus L.)及大戟科鐵色 (Drypetes littoralis (C.B.Rob.) Merr.)共佔了 79.8%,顯示烏頭翁選擇築巢的巢樹種類亦相當廣泛,並未必限定在特定樹種上。
本研究在台東農業果園環境中,發現烏頭翁親鳥共利用 7 種植物來築巢繁殖,其 中荔枝、酪梨、釋迦、枇杷等果樹佔 88.2%,因此烏頭翁可利用不同果樹築巢,
其對果園環境應具有相當程度的繁殖適應力。
二、繁殖季
烏頭翁在屏東墾丁青蛙石區域從 3 月下旬開始築巢、生蛋, 4 月為築巢期與 孵卵期的巔峰期,之後下降到 7 月上旬止(Hsu and Lin 1997)。在台東縣果園環境,
烏頭翁的繁殖季從 3 月上旬即開始築巢、下蛋,孵卵階段的巢數在 5、6 月達到最 多,再下降至 8 月中旬結束,烏頭翁在本研究中的繁殖季比屏東墾丁早約半個月,
延後約 1 個月。
徐(1984)提到烏頭翁的生殖腺在 3~5 月有明顯增重,在 7~8 月逐漸退化 的情形;烏頭翁的睪丸和卵巢的重量均於 4 月最重,再從其卵巢重量和孵卵斑的 出現推測雌鳥在 4~6 月參與生殖來看,徐(1984)與本研究的結果顯示 4~6 月 為烏頭翁繁殖季合理的高峰期。
徐(1984)提到烏頭翁在乾季長達 9 個月的墾丁地區,雨量和食物可能是影 響繁殖季開始時間不同的兩個因素。台東地區的平均降雨量較墾丁地區豐沛,食 物資源可能較豐富,而使得烏頭翁能夠較早進入繁殖季。
三、窩卵數
(一)最適窩卵數
在 Hsu and Lin (1997)屏東墾丁烏頭翁之 34 個鳥巢調查中,其窩卵數 在 3~4 個之間,其中每窩 3 卵的巢數佔 66%,每窩 4 卵的巢數佔 23%,平均 窩卵數為 3.3 個。本研究在烏頭翁 177 個鳥巢的窩卵數在 2~4 個之間,其中 每窩 2 卵的巢數佔 11%,每窩 3 卵的巢數佔 80%,每窩 4 卵的巢數佔 9%,平 均窩卵數為 3.0 個。
鳥類會依所能供給雛鳥充分食物的前提下,趨向產下最大的窩卵數(Lack
,並再經由天擇之作用,以存活最多幼鳥數的窩卵數,做為其最適窩卵
數大小(Lack 1948)。在屏東及台東兩地區烏頭翁的窩卵數均以每窩 3 卵最 多,平均窩卵數也大約為 3,本研究中以每巢 3 隻雛鳥的巢數佔最多,其離巢 率也最高(圖 22),因此,烏頭翁的最適窩卵數應該是每窩 3 卵。
(二)巢數對窩卵數之影響
但是,本研究之資料顯示:當巢數在不同年間、繁殖季不同時期間與不 同果園間明顯增加時,其成功存活離巢之幼鳥數卻都一致下降。而且對照本 研究烏頭翁在果園繁殖窩卵數低於 Hsu and Lin (1997)在墾丁次生林之表 現,這些顯示巢數多可能引起種內競爭上升,其影響首先反映於母鳥產下之 窩卵數較低,並又進一步反映在母鳥育雛成功存活幼鳥數也相對較低。
四、鳥蛋重量與大小
(一)窩卵數之影響
然而,本研究之烏頭翁蛋長在不同窩卵數間有顯著差異,其蛋長隨著窩 卵數的增加而減少,以每窩 2 卵的蛋長最長,每窩 4 卵的蛋長最短;屏東墾 丁烏頭翁窩卵數每窩 3 卵的蛋長也較每窩 4 卵為長 Hsu and Lin(1997)。
Williams(2001)提及在鳥類生殖資源提供有限的情形下,其窩卵數高低和蛋 尺寸大小之間具有取捨(trade-off)表現,亦即在兩者之間會形成一種負相關 的變化關係。烏頭翁之蛋長與窩卵數間似有相同之取捨表現。
(二)繁殖季不同時期之影響
在本研究中,烏頭翁之鳥蛋重量與其大小都以繁殖季早期顯著高於中、
晚期。而蛋重量與蛋大小也都以在台東市果園高於龍田果園。造成蛋重量與 蛋大小在不同繁殖季時期或不同果園環境之間會形成差異的原因,本研究推 論主要可能與食物資源競爭有關。亦即由於在繁殖季早期和台東市果園的繁 殖巢數都相對明顯較少,負責產卵的母鳥由於在種內個體間競爭低的條件 下,每一母鳥相對可獲得較高的食物量,因此使其得以產下較重又較大的鳥 蛋;相對的,在繁殖季中期或龍田果園中之繁殖巢數則都顯著較高,在此兩 環境中,種內個體間對食物資源的競爭將大增,每一母鳥平均可得之食物較 不足,可能是造成蛋重與蛋大小明顯較低的原因。
五、孵化率、離巢率、繁殖成功率與離巢成功幼鳥數
烏頭翁之孵化率在不同年間(圖 14)、不同果園間(圖 15)及不同繁殖季時 期間(圖 16)都無顯著差異,但離巢率與繁殖成功率在以上三種不同情形下,卻 都有隨著巢數的增加而降低之一致的變化趨勢。孵化率都無差異,但是離巢率卻 都一致偏低,顯示有相當大比例的孵化雛鳥數無法順利發育至成功離巢。
在窩卵數、孵化雛鳥數及成功離巢幼鳥數中,只有成功離巢幼鳥數才能真正 加入烏頭翁之成鳥族群中,並進行配對繁殖,產生後代以維持族群之發展。而成 功離巢幼鳥數在不同年度間與繁殖季不同時期而改變,在龍田果園的成功離巢幼 鳥數則顯著低於台東市果園,加上整體繁殖成功率只有 20%左右,和屏東墾丁的
烏頭翁 43.8%(徐 1984)、小雲雀 (Alauda gulgula) 40.6%(袁 1986)、麻雀 (Passer montanus) 75%(陳 1986)、繡眼畫眉 (Alcippe morrisonia) 63.8%(林 1996)、金 翼 白 眉 (Garrulax morrisonianus) 81.3% ( 游 1998 )、 菲 律 賓 輝 椋 鳥 (Aplonis panayensis) 51.6%(林 2001)及家燕 (Hirundo rustica) 63.4%(賴 2004)相較之
下明顯偏低,顯示在雛鳥發育期面臨相當嚴苛的生存威脅,推測主要可能包括雛 鳥間對食物的競爭、母鳥取得食物難易、天敵危害及果園農作管理等因素。以下 針對這些因素分別加以探討:
(一)烏頭翁雛鳥食物資源的競爭
Martin(1987)整理鳥蛋的大小和蛋的品質之相關研究指出,較大的蛋其孵 化率、雛鳥重量、幼雛成長速率及離巢率等表現均優於比較小的蛋。Amundsen
(1995)提到雪鸌(Pagodroma nivea)的蛋大小與剛孵化 2~4 天的雛鳥體重 之間存有正相關的變化關係。Perring(1996)也報導母鳥在產卵前除了需要 獲取維持生命所需的營養外,還需攝取額外的食物量以供產卵所需,母鳥產 下的蛋大小與所需的食物營養呈正相關,而大山雀(Parus major)所產下較 大的蛋其孵化率和離巢率也較高。烏頭翁的鳥蛋大小在台東市果園比龍田果 園大,在繁殖季早期也比中、晚期大,而其孵化率、離巢率和繁殖成功率也 均較高。蛋大小影響孵化率、離巢率的情形與國外的研究結果一致。種內個 體間對食物資源的競爭可能是影響烏頭翁蛋大小的因素,也可能間接影響到 孵化率、離巢率和繁殖成功率的表現。
Martin(1987)也提到食物豐富度會隨著繁殖季急速減少,在多數幼鳥離
巢時期達到最低量,因此在繁殖季早期的成功離巢幼鳥數會比繁殖季晚期 多。符合本研究烏頭翁在不同繁殖季時期的離巢率、離巢幼鳥數和繁殖成功 率的表現。而在 2006 年及龍田果園進行繁殖的巢數明顯較多,雛鳥的食物需 求量也最多,在有限的繁殖棲地環境下所能提供的食物量也有限,因此可能 也是因為食物資源的不足,親鳥能爭奪到的食物量不足以餵養已孵化的雛 鳥,而導致離巢幼鳥數偏低。
(二)烏頭翁雛鳥致死的原因探討
劉(2005)在屏東墾丁的研究指出影響烏頭翁繁殖成功的主要原因有親 鳥棄巢、持續惡劣天候、天敵掠食、棲地變化及人為干擾(例如割草或果農 噴灑農藥等)。本研究依實際觀察結果將烏頭翁雛鳥致死的原因分為在巢中死 亡、不明原因不見、被掠食、人為因素及天候因素五個原因,對照兩果園烏 頭翁雛鳥致死原因(圖 23)、(圖 24),台東市果園除了人為因素佔 5%外,其 他均為不明原因不見佔有 95%;而龍田果園除了人為因素相同佔 5%之外,還 有在巢中死亡佔 4%和被掠食佔 3%,其餘均為不明原因不見佔 88%。由以上 可知兩果園烏頭翁雛鳥致死原因主要為不明原因不見。Sockman(1997)將前 次探巢時還存有未離巢的蛋或雛鳥,而在下一次探巢時不見的蛋及雛鳥判定 為被掠食,本研究若依 Sockman 的方法將不明原因不見的雛鳥判定為被掠 食,再加上原有確定被掠食的雛鳥比例,則在兩果園環境中,所有雛鳥致死 的原因以被掠食的比例最高分別為 95%和 91%。不過,親鳥也有會將病死之 幼鳥叼出鳥巢之情形,因此,確實之原因,仍有待進一步之研究加以釐清。
(三)巢構造與巢位選擇
由於巢構造的所有測量項目在兩果園間都無顯著差異,因此烏頭翁的巢 位選擇或為影響繁殖成功與否的重要因素。台東市果園的平均巢位高度為 213 cm,龍田果園的平均巢位高度為 149.6 cm,兩者有明顯差異,再進一步將兩 果園與 Hsu and Lin(1997)和劉(2005)的巢位高度、平均蛋數和平均離巢 數共同比較,發現平均每巢離巢幼鳥數與巢位高度具有明顯一致之正相關變 化趨勢(圖 25)。
對於陸鳥來說,天敵掠食是造成繁殖失敗主要的原因(Ricklefs 1969a),
Wilson and Cooper(1998)報導中美綠蚊霸鶲 (Empidonax virescens) 的巢位愈 低,被天敵掠食的機率也愈高,其繁殖成功率也愈低。Sockman(1997)將鳥的 巢樹分為上、中、下三等高度檢驗,發現築巢位置於下方所受到天敵掠食的 比率高於築巢位置在巢樹的中、上方。由於囓齒目動物對於雀形目鳥類築於 地面或接近地面的巢都有一定程度的掠食威脅(Ettel et al 1998 ; Hannon and Cotterill 1998),而雛鳥發出求食聲的次數愈多、音量愈大,被來自地面或天 空的天敵掠食的機率就愈大(Dearborn 1999)。Hsu and Lin(1997)報導在屏 東墾丁地區威脅烏頭翁最大的天敵可能就是在當地數量眾多的野貓 (felis catus) 和赤腹松鼠 (Callosciurus erythraeus),此兩物種在龍田果園也有一定的
族群數量,另外,本研究曾記錄到紅尾伯勞 (Lanius cristatus) 啄食並叼走烏 頭翁尚未離巢幼鳥之情形。綜合以上所述,導致烏頭翁雛鳥死亡的因素主要 可能為被天敵所掠食。
綜合以上我們推論:當進行繁殖的巢數增加,將造成烏頭翁的種內食物
競爭上升,使食物資源量無法充分滿足雛鳥需求;而巢位高度低矮,會導致 地面天敵掠食率偏高,應為影響繁殖成功率顯著下降的主要原因。
六、孵卵期與育雛期之發育速率
屏東墾丁烏頭翁的孵卵期為 11~12 天,平均孵卵期為 11.4 天;育雛期為 9~
10 天(Hsu and Lin 1997)。本研究烏頭翁的孵卵期為 9~13 天,平均孵卵期為 10.6 天;育雛期 8~12 天。
Martin(1987)報導若孵卵親鳥擁有健康與足夠體重,將可減少覓食次數,進 行長時間的孵卵工作,以維持巢內蛋的溫度,縮短孵卵期;Sanz(1996)關於班 鶲(Ficedula hypoleuca)在不同食物供給量下,母鳥體重與孵卵期之間呈現負相關 的變化趨勢,即母鳥重量愈重,孵卵期愈短。
Ricklefs(1969b) 提出在育雛階段親鳥若能提供雛鳥足夠的食物量,則雛鳥便 能以最快的速率成長,縮短育雛期。Williams(1980)報導大賊鷗(Stercorarrus skua lonnbergi)從較重的鳥蛋所孵化出來的雛鳥,其生長會較為快速,亦即會縮短育雛
期。本研究烏頭翁在不同窩卵數間以窩卵數較低且蛋重較重的育雛期比窩卵數較 高且蛋重較輕的育雛期來得短;而在繁殖季早期蛋重較重的育雛期也比繁殖季晚 期蛋重較輕的育雛期短。
綜合影響孵卵期和育雛期的因素,負責產蛋、孵卵的親鳥體重佔有很重要的 角色,而本研究為避免對親鳥育雛之干擾,並未針對親鳥體型進行測量,因此在 孵卵期和育雛期的差異原因方面有待後續研究進一步探討。
伍、結論
從本研究顯示,於原始林邊緣地帶生長的烏頭翁族群,在台東縣境內果園環 境,普遍地進行築巢繁殖,但是可能因為同時進行繁殖的烏頭翁數量多,食物資 源的激烈競爭和不足,讓母鳥無法產下品質較佳的鳥蛋,致使鳥蛋較輕、較小;
也因育雛的食物不足,使得成功發育至離巢的幼鳥數也偏低;而果園管理經營所 需,導致巢樹低矮,相對地也影響到烏頭翁築巢的高度不高,而招致來自地面天 敵掠食率偏高,進而影響其族群增長的繁殖成功率介量。在本研究龍田果園環境 中,極可能就是因為同鳥種間的食物資源的激烈競爭,造成食物不足,以及巢位 高度低,招致天敵掠食率偏高,而導致繁殖成功率明顯偏低。
需要注意的是,類似龍田果園的環境條件,在台東地區相當普遍,依據台東 縣政府 2006 年農業統計分析:台東縣的果樹種植面積為 13,267 公頃,而釋迦(番 荔枝)種植面積有 5,041 公頃、枇杷 141 公頃及荔枝 113 公頃,共佔全縣果樹總種 植面積的 39.9%(台東縣政府 2006)。烏頭翁在果園大量築巢但離巢成功率卻偏 低,此對於其族群發展之影響,將是一個值得研究探討的方向。
陸、引用文獻
王嘉雄、吳森雄、黃光瀛、楊秀英、蔡仲晃、蔡牧起、蕭慶亮。1991。台灣野鳥 圖鑑。亞舍圖書有限公司,台北。
方偉宏。2005。台灣受脅鳥種圖鑑。貓頭鷹出版社,台北。
李壽先。1989。綠繡眼的生殖及非生殖季成群之研究。東海大學生物研究所碩士 論文。
林宗鍵。2001。高雄市菲律賓輝椋鳥食性適應與生殖適應之研究。國立高雄師範 大學生物科學研究所碩士論文。
林華慶。1992。白頭翁與烏頭翁之鳴唱聲比較。國立台灣大學動物學研究所碩士 論文。
林瑞興。1996。繡眼畫眉生殖及鳥群生態之研究。國立台灣大學動物學研究所碩 士論文。
姚姿君。1984。白頭翁烏頭翁在形態上以判別分析法區分雌雄之探討。國立台灣 大學農藝所碩士論文。
徐芝敏。1984。烏頭翁與白頭翁之生物學研究。國立台灣大學動物學研究所碩士 論文。
袁孝維。1986。墾丁牧場小雲雀之行為及生態學研究。國立台灣大學動物學研究 所碩士論文。
許譽騰。1999。由族群遺傳結構探討白頭翁與烏頭翁之演化關係。國立台灣大學 動物學研究所碩士論文。
所碩士論文。
張瓊文。1990。白頭翁與烏頭翁鳴聲的分析與比較。國立台灣大學動物學研究所 碩士論文。
游淑鈞。1988。塔塔加地區金翼白眉生殖及覓食生態之研究。國立臺灣大學森林 學研究所碩士論文
賴秀花。2004。新竹市及鄰近地區家燕生殖生態調查研究。進修部數理教育碩士 班(自然組)碩士論文。
劉小如。1990。太魯閣國家公園烏頭翁及白頭翁分布調查。花蓮:太魯閣國家公 園管理處。
劉小如。1991。太魯閣國家公園烏頭翁及白頭翁生態及行為研究。花蓮:太魯閣 國家公園管理處。
劉小如。2003。墾丁國家公園及鄰近地區烏頭翁與白頭翁雜交狀況調查。屏東:
墾丁國家公園管理處。
劉小如。2005。墾丁國家公園烏、白頭翁擴散暨烏頭翁保護區劃設之可行性研究
(一)。屏東:墾丁國家公園管理處。
台 東 縣 政 府 農 業 統 計 分 析 。 2006 年 12 月 30 日 , 取 自 : http://www.taitung.gov.tw/department/p4/p4b12.htm/果品。
Amundsen, T. 1995. Egg size and early nestling growth in the Snow Petrel. Condor 97:
345-351.
Best, L. B., K. E. Freemark, J. J. Dinsmore, and M. Camp. 1995. A review and synthesis of habitat use by breeding birds in agricultural landscapes of Iowa. American Midland Naturalist 134: 1-29.
Campbell, B. and E. Lack. (eds) 1985. A Dictionary of Birds. Calton (Poyser) and Buteo Books, Vermillion.
Dearborn, D. C. 1999. Brown-headed Cowbird nestling vocalizations and risk of nest predation. Auk 116: 448-457.
Ettel, T. L., D. A. Buehler, and A. E. Houston. 1998. Egg size and cotton rat predation.
Wilson Bulletin 110: 575-578.
Filliater, T. S., R. Breitwisch, and P. M. Nealen. 1994. Predation on northern cardinal nests: does choice of nest site matter? Condor 96: 761–768.
Galbraith, H. 1988. Effects of agriculture on the breeding ecology of Lapwings Vanellus vanellus. Journal of Applied Ecology 25: 487-503.
Gregoire, A., S. Garnier, N. Dreano, and B. Faivre. 2003. Nest predation in Blackbirds (Turdus merula) and the influence of nest characteristics. Ornis Fennica 80: 1–10.
Hannon, S. J., and S. E. Cotterill. 1998. Nest predation in aspen woodlots in an agricultural area in Alberta: the enemy from within. Auk 115: 16-25.
Hilden, O. 1965. Habitat selection in birds. Annales Zoologici Fennici 2: 53-75.
Holway, D. A. 1991. Nest-site selection and the importance of nest concealment in the black-throated blue warbler. Condor 93: 575–581.
Hoover, J. P., and M. C. Brittingham. 1998. Nest-site selection and nesting success of wood thrushes. Wilson Bulletin 110: 375-383.
Hsu, M. J., and Y. S. Lin. 1993. Morphology, sexual dimorphism and hybridization in Styan’s bulbuls Pycnonotus taivanus and Chinese bulbuls P. sinensis formosae.
Acta Zoologica Taiwanica 4: 103-111.
Hsu, M. J., and Y. S. Lin. 1997. Breeding ecology of Styan’s Bulbul Pycnonotus
Lack, D. 1947. The significance of clutch size. Ibis 89: 302-352.
Lack, D. 1948. The significance of clutch size III. Ibis 90: 25-45.
Linden, M., and A. P. Moeller. 1989. Cost of reproduction and covariation of life history traits in birds. Trends in Ecology & Evolution 4: 367-371.
Loiselle, B. A., and W. G. Hoppes. 1983. Nest predation in insular and mainland lowland rainforest in Panama. Condor 85: 93-95.
Maicas, R., and J. F. Haeger. 2004. Pine plantations as a breeding habitat for a hole-nesting bird species crested tit (Parus cristatus) in southern Spain. Forest Ecology and Management 195: 267-278.
Marshall, A. J. 1959. Internal and environmental control of breeding. Ibis 101: 456-478.
Martin, T. E. 1987. Food as a limit on breeding birds: A life-history perspective. Annual Review of Ecology and Systematics 18: 453-487.
Martin, T. E. 1992. Interaction of nest predation and food limitation in reproductive strategies. Current Ornithol. 9: 163-197.
Martin, T. E. 1993. Nest predation among vegetation layers and habitat types: revising the dogmas. American Naturalist 141: 897-913.
Martin, T. E. 1995. Avian life history evolution in relation to nest sites,nest predation, and food. Ecological Monographs 65: 101-127.
Martin, T. E., and J. J. Roper. 1988. Nest predation and nest-site selection of a western population of the Hermit Thrush. Condor 90: 51-57.
Martin, T. E., and Li. Pingjun. 1992. Life history traits of open- vs. cavity- nesting birds.
Ecology 73: 579-592.
Martin, T. E., M. L. Morton, K. W. Sockman, and L. E. Peterson. 1993. Nest Predation in the Mountain White-Crowned Sparrow. Condor 95: 72-82.
Moller, A. P. 1990. Effect of parasitism by a heamatophagous mite on reproductive in the barn swallow. Ecology 71: 2345-2357.
Nice, M. M. 1941. The role of territory in bird life. American Midland Naturalist 26:
441-487.
Perrins, C. M. 1996. Eggs, egg formation and the timing of breeding. Ibis 138: 2-15.
Rand, A. L., and H. G. Deignan. 1960. Family Pycnonotidae. Jr. pp.221-300 In Check-List of Birds of the World. Vol IX. ed. E. Mayr, and J. C. Greenway.
Museum of Comparative Zoology, Cambridge, Mass.
Reale, J. A., and R. B. Blair. 2005. Nesting success and life-history attributes of bird communities along an urbanization gradient. Urban Habitats 3: 1-24.
Ricklefs, R. E. 1969a. Analysis of nesting mortality in birds. Smithsonian Contributions to Zoology 9: 1-48.
Ricklefs, R. E. 1969b. Preliminary models for growth rates in altricial birds. Ecology 50:
1031-1039.
Robertson, G. J. 1995. Factors affecting nest site selection and nesting success in the common eider Somateria mollissima. Ibis 137: 109-115.
Rodenhouse, N. L., and L. B. Best. 1983. Breeding ecology of Vesper Sparrows in corn and soybean fields. American Midland Naturalist 110: 265-275.
Sanz, J. J. 1996. Effect of food availability on incubation period in the Pied Flycatcher (Ficedula hypoleuca). Auk 113: 249-253.
SAS Institute. 1989. SAS/STAT user’s guide, version 6, fourth ed., vol. 1. SAS Institute, Cary, NC.
Sedinger, J. S., and P. L. Flint. 1995. Environmental influence on life-history traits : growth survival, and fecundity in Black Brant (Branta bernical). Ecology 76:
2404-2414.
Severinghaus, S. R., and K. T. Blackshaw. 1976. A new guide to the birds of Taiwan.
Mei Ya Publication, Inc., Taipei. 222 pp.
Skutch, A. F. 1949. Do tropical birds rear as many young as they can nourish? Ibis 91:
430-458.
Smith, H. G., and M. Bruun. 2002. The effect of pasture on starling (Sturnus vulgaris) breeding success and population density in a heterogeneous agricultural landscape in southern Sweden. Agriculture, Ecosystems & Environment 92: 107-114.
Sockman, K. W. 1997. Variation in life-history traits and nest-site selection affects risk of nest predation in the California Gnatcatcher. Auk 114: 324-332.
Stearns, S. C. 1977. The evolution of life history traits : A critique of the theory and a review of the data. Annual Review of Ecology and Systematics 8: 145-171.
Steel, B. B. 1993. Selection of foraging and nesting sites by Black-throated Blue warblers: Their relative influence on habitat choice. Condor 95: 568-579.
Williams, A. J. 1980. Variation in weight of eggs and its effect on the breeding biology of the Great Skua. Emu 80: 198-202.
Williams, T. D. 2001. Experimental manipulation of female reproduction reveals an intraspecific egg size-clutch size trade-off. Proceedings: Biological Sciences 268:
423-428.
Wilson, R. R., and R. J. Coopers. 1998. Acadian Flycatcher nest placement: Does placement influence reproductive success? Condor 100: 673-679.
圖 1. 研究樣區地圖—台東市果園,有 T-1、T-2、T-3 及 T-4 四個樣區
T-1
T-2
T-4
T-3
圖 2. 研究樣區地圖—龍田果園,有 L-1、L-2 及 L-3 三個樣區
L-1 L-3
L-2
圖 3. 烏頭翁巢構造各部份示意圖。a:巢外口徑寬,b:巢內口徑寬,c:巢高,d:巢深。
圖 4. 巢位特徵之樹冠位置指標示意圖。e: 通過巢與樹心的軸為長軸,f: 通過巢與 長軸垂直的軸為短軸,樹冠位置指標 g=f/e。
b a
d
c
e
f
0 20 40 60 80 100 120 140
早期 中期 晚期
繁殖季時期
孵卵期巢數
2005 2006
圖 5. 2005-2006 年台東縣烏頭翁繁殖季不同時期之孵卵期巢數頻度分布
0 20 40 60 80 100 120 140
早期 中期 晚期
繁殖季時期
育雛期巢數
2005 2006
圖 6. 2005-2006 年台東縣烏頭翁繁殖季不同時期之育雛期巢數頻度分布
0 20 40 60 80 100 120 140
2005年 2006年
巢數
窩卵數2 窩卵數3 窩卵數4
圖 7. 2005-2006 年烏頭翁在不同年間之窩卵數巢數頻度分布
0 20 40 60 80 100 120 140
台東市果園 龍田果園
巢數
窩卵數2 窩卵數3 窩卵數4
圖 8. 2005-2006 年烏頭翁在不同果園間之窩卵數巢數頻度分布
0 20 40 60 80 100 120 140
繁殖季早期 繁殖季中期 繁殖季晚期
巢數
窩卵數2 窩卵數3 窩卵數4
圖 9. 2005-2006 年烏頭翁在繁殖季不同時期間之窩卵數巢數頻度分布
15.5 16.5 17.5 18.5 19.5 20.5 21.5
2005 2006
cm
2.6 2.7 2.8 2.9
g
蛋長蛋寬 蛋重
圖 10. 2005-2006 年烏頭翁在不同年間蛋重、蛋長及蛋寬比較
15.5 16.5 17.5 18.5 19.5 20.5 21.5
台東市果園 龍田果園
cm
2.6 2.7 2.8 2.9
g
蛋長 蛋寬 蛋重
圖 11. 2005-2006 年烏頭翁在不同果園間蛋重、蛋長及蛋寬比較