I
目錄
中文摘要………II 英文摘要………III
前言 ... 1
研究目的 ... 2
文獻探討 ... 3
研究方法 ... 5
結果與討論 ... 7
參考文獻 ... 9
附表 ... 11
附圖 ... 13
中文摘要
赤 桉(Eucalyptus camaldulensis)為 造 紙 重 要 的 來 源 樹 種 , 自 紙 漿 工 業 轉 移 至 熱 帶 之 後 , 利 用 桉 樹 所 生 產 的 紙 漿 需 求 大 增 。 本 計 劃 已 選 殖 出 與 纖 維 素 合 成 相 關 的 基 因 纖 維 素 降 解;KORRIGAN (EucKor) 基 因 之 cDNA。其 全 長 為 2379bp,含 ORF 為 1869bp,5’端 約 有 149bp UTR; 3’端 為 361 bp;約 為 623 個 胺 基 酸。與 基 因 對 應 全 長 之 gDNA 約 為 3391 bp, 含 六 個 exons 與 五 個 introns。 並 進 一 步 藉 定 量 RT-PCR, 發 現 主 要 表 達 區 域 為 莖 段 , 少 量 於 木 質 部 與 韌 皮 部 。 EucKor 基 因 已 經 由 農 桿 菌 成 功 地 轉 殖 到 菸 草 與 赤 桉 優 良 營 養 系 , 研 究 成 果 預 期 將 深 入 瞭 解 影 響 紙 漿 生 產 的 相 關 基 因 調 控 , 目 前 臺 灣 林 業 試 驗 所 已 在 赤 桉 中 分 離 出 數 個 影 響 紙 漿 生 產 的 木 質 素 生 合 成(lignin biosynthesis) 基 因 (如 4CL1, C4H) , 所 以 , 藉 由 農 桿 菌 轉 殖 也 可 再 轉 殖 到 具 有 木 質 素 基 因,或 纖 維 素 合 成 基 因(CesA)的 赤 桉 中 , 由 此 獲 得 兼 具 紙 漿 生 產 多 價 轉 基 因(gene stacking)林 木 , 這 些 基 因 轉 殖 苗 木 將 會 是 研 究 基 因 調 控 最 佳 的 材 料 , 也 會 在 全 球 能 源 短 缺 之 際 , 充 分 瞭 解 林 木 碳 源 配 置 , 提 供 林 木 生 質 能 源 最 有 效 益 的 使 用 方 式 。
關鍵字
Eucalyptus camaldulensis, KORRIGAN, gene-stacking trees
III
英文摘要
Eucalyptus is one of the most valuable plantation tree due to its extremely fast growth rate, superior fiber and pulping properties. We have clone a KORRIGAN gene which is 2379 bp containing a ORF 1869bp and 149bp for 5’UTR, 361 bp for 3’-UTR. There are five introns and six exons in its genomic structure. To characterize their expression pattern by RT-PCR, we found the expression is highest in stems (4~9 internodes) and lowest in xylem and phloem. in situ hybridization could not differentiate the expression pattern at cellular level. To deliver EucKOR genes in sense orientation into E. camaldulensis and tobacco via
Agrobacterium-mediated gene transformation, we had selected several putative E.
camaldulensis calli and tobacco transformants for further characterization. In
future, we plan to transfer EucKOR genes into wood-formation related transgenic E.camaldulensis to generate gene-stacking trees in the future. Those transgenic Eucalyptus trees would offer the potential for the development of sustainable bioenergy and may lead us to a new manufacturing system.
關鍵字
Eucalyptus camaldulensis, KORRIGAN, gene-stacking trees
前言
桉樹 (Eucalyptus) 即一般耳熟能詳的尤加利樹,其具有生長快速(巴西生長最快 約為每年每畝7.8 立方公尺,中國海南島生長最快約為每年每畝 4 立方公尺),木 材耐腐性強,輪伐期短 (約 5~7 年) 等特性,可作為傢俱、紙漿及纖維等工業用 途;而枝葉可提煉精油,作為醫療之用途,所以為熱帶最重要的造林樹種之一。
由桉樹生產的紙漿用量大增,如何提高桉樹紙漿產量,是近來林木生物科技的焦 點。由於臺灣林業試驗所已投入相當的資源,並協助產業提升紙漿之產量;但礙 於傳統的林業無法滿足需求量增高的紙漿工業。本計畫利用生物科技技術,希望 能加速林業於優良品系上的選拔,並進行基因轉殖苗木提供業界,除了產生高質 量的纖維素以增加紙漿廠收益外;另一方面,希望發展出林木纖維素以外之材料,
以做為生質能源的原料,而這些都是林木科學迫在眉睫的研究主題。本計畫希望 能快速地完成纖維素合成相關基因篩選,將其特性與功能以分子生物方式加以鑑 定並結合基因轉殖技術,使基因的特性能以活體的方式呈現,對此基因特性的分 析將更精確。在紙漿產量提升研究上,可由最重要的兩個因子下手:減低木質素 生產與提高纖維素含量;而針對生質能源的目標,則可由纖維素相關基因下手,
了解其調控機制,進而操縱其碳源的分佈,提高林木的利用價值。因此本計畫期 望藉由已建立的赤桉平台,在現有臺灣林業試驗所的研究之外,同時投入赤桉於 纖維素相關基因的研究團隊,經由林業試驗所育林組陳振榮與鍾振德博士等人協 助,利用已建立的赤桉 (Eucalyptus camaldulensis) 基因轉殖體系,未來將完成更 多與纖維素生產相關基因的選殖及轉殖研究。
2
研究目的
為提高木材在紙漿產業之利用價值,臺灣林業試驗所已由赤桉中分離出數個 影響木質素生合成途徑(lignin biosynthesis pathway)的相關基因 (例如 4CL1, C4H),並經由農桿菌將基因轉殖到不同的赤桉 (Eucalyptus camaldulensis) 品種,
也已達到隔離造林試驗階段;然而、影響紙漿產量可經由減低木質素生產或是提 高纖維素含量。所以、除了上述的基因之外,仍有許多涉及紙漿生產的因子,例 如纖維素、及半纖維素基因等相關生合成途徑之基因。 纖維素是細胞壁的主要 成分之一,由數千至上萬個D-葡萄糖分子,藉由 β-1,4 糖苷鍵連接的鏈狀聚合 物,也是世界上最豐富且擁有巨大商業價值的生物多聚體之一。近幾年來,隨著 基因組學的發展,關於高等植物纖維素生合成及相關基因表達調控的研究也有增 加的驅勢。雖然纖維素是由葡萄糖形成的多聚體,但是其合成機制卻相當複雜,
同時涉及到多種酶的調控。有關植物纖維素生合成機制,相關研究已有四十多年,
但進展十分緩慢;一直到近二十年,研究者開始由基因操控著手,1996 年在棉 花發現與纖維素相關的基因CesA,至今已有十餘年,期間水稻、阿拉伯芥、玉 米等植物中陸續選殖出CesA 的基因。而更多的證據相信植物除了纖維素 CesA 基因在纖維素合成中具有重要作用,其它酵素 (例如類纖維素合成酶,
KORRIGAN,蔗糖合成酶 SuSy,細胞的骨架蛋白,Rac13 蛋白等) 也同等重要,
需共同協調才能完成如此複雜之調控(Lerouxel et al, 2006)。所以此計畫的目標,
將選殖與纖維素生合成相關 KORRIGAN 基因(KOR)之 cDNA 、基因及其啟動子,
藉由定量RT-PCR,原位雜交等技術探討其基因表現,進而探討纖維素生合成調 控機制,同時將 KOR 轉殖進入赤桉中,期望研究成果在未來林木產業上可獲得 最佳的經濟效益。
文獻探討
纖維素 (cellulose) 是目前已知自然界中存量最多的一種生物性多醣類化合 物,廣泛分佈於植物界中,為植物體細胞壁 (cell wall)的主要組成成份之一,由 數千至上萬個D-葡萄糖分子,藉由β-1,4 糖苷鍵連接的鏈狀聚合物,也是世界 上最豐富且擁有巨大商業價值的生物多聚體之一。1838 年法國化學家佩恩 (Anselme Payen) 首次發現並證明纖維素是由D-葡萄糖 (glucose) 單體所組成,
經過β-1,4 糖苷鍵 (glycosidic bond) 鍵結而成,由500~1,400 個葡萄糖單體,進 而形成長鏈型的高分子聚合物。近幾年來,隨著基因組學的發展,關於高等植物 纖維素生合成及相關基因表達調控的研究也有增加的驅勢。雖然纖維素是由葡萄 糖形成的多聚體,但是其合成機制卻相當複雜,同時涉及到多種酶的調控。1996 年在棉花發現與纖維素相關的基因CesA,至今已有十餘年,期間水稻、阿拉伯 芥、玉米等植物中陸續選殖出CesA 的基因。而更多的證據相信植物除了纖維素 CesA 基因在纖維素合成中具有重要作用,其它酵素(例如纖維素合成酶,
KORRIGAN,蔗糖合成酶SuSy,細胞的骨架蛋白,Rac13 蛋白等)也同等重要,
需共同協調才能完成如此複雜之調控(Lerouxel et al, 2006)。而有關植物纖維素生 合成機制,相關研究已有三十多年,但進展十分緩慢;一直到近代,研究者才開 始由基因操控著手,並用了電子顯微鏡來對纖維素進行研究。在1983年發現水棉 (Spirogyra) 的微纖維中含有36條的葡聚醣鏈,因此,與研究人員假設的六到十 個次單元組合而成的蓮花狀球體,將六條寬2 nm的原纖維 (fibrils) 結合形成微纖 維 (microfibrils) 的想法一致,纖維素又開始成研究者著手探討的主題。
赤桉纖維素降解酶(KOR)為一種位於細胞膜上與纖維素合成有關的酵素,
其可能作用於細胞膜及細胞壁之交界,因此被認為與細胞壁的修 飾 有 關 。 基 因 序 列 比 對 的 結 果 發 現,K O R 為 endo-1,4–β-D glucanase巨大家族的成員,
也就是一種纖維素酶(cellulase),一般而言,被認為可水解具有1,4–β-D glucan骨 架的多醣體。而根據序列分析,KOR具有穿膜蛋白序列,具有LL和YXX中的 polarized targeting signals,顯示KOR可能位於細胞膜上,且在細胞分裂時位於細 胞板上,與細胞分裂有極高的相關性。1998年由Nicol等人分離出KOR基因,發 現序列約為2.8kb,可能為一個小的成員基因家族,到目前為止,KOR主要都是 以阿拉伯芥來進行較為深入的研究。目前,KOR基因被證實在阿拉伯芥中扮演 纖維素降解酶(cellulase)的角色,其主要作用於纖維素合成之前驅物;固醇糖苷 (steryl-glucoside,SG)之解離,能使纖維素鏈不斷的延長。而在不同的KOR基因 成員中,有發現到其組織的專一性,例如阿拉伯芥中的KOR2主要表現於根的分 生區域中;而KOR基因也可見於木質部、頂端分生組織與維管束等組織中。研 究也發現,當其基因表現量下降時,會導致植物在生長中造成中度至重度的缺陷,
如木質部的不規則表現和影響纖維素的超微結構。將KOR基因的啟動子區域插 入一段序列,發現基因表現量下降,而產生了不正常的擴大生長,此研究也暗示
4
KOR可能與細胞壁的生長有關,可是並無明顯的改變細胞壁和植物體本身的生 長,但確實影響了纖維素中超微結構的產生,導致樹木擁有較少的結晶纖維素的
含量(18,19)。除此之外,在楊樹的受壓木組織中也可以見到不同的KOR基因表達量
(Bhandari et al﹐2006),科學界對於纖維素與其調節機制具有非常高的興趣,但 是,目前還是以阿拉伯芥及棉花的研究最為深入。目前已有楊樹已進行KOR基 因轉殖,發現KOR與初生細胞壁及細胞伸長有關,當KOR基因突變時也會造成 細胞壁組成的改變。
再者,根據許多林木科學家的經驗,阿拉伯芥這類模式植物的研究結果,並 不能全然套用在林木系統上,主要是因為演化的過程中基因體大小、植物生活期、
開花授粉屬性等差異而造成。 赤桉是關係臺灣紙漿產業最直接的樹種之一,但 文獻資料中,至今並無赤桉相關的纖維素生合成基因研究在臺灣進行。本計劃為 了解赤桉纖維素合成機制,除選殖 KOR 基因之外,還會使用赤桉 KOR 先在菸 草中進行基因轉殖,主要是因為林木基因轉殖苗所需的時間較長,雖然模式植物 的成果不盡然能忠實地在赤桉重建,但在取得赤桉轉殖苗前將可獲得初步在植物 中in vivo 中所呈現之現象。所以本研究將以赤桉為材料,將 KOR 基因以正向送 入植物體內,將建立之轉殖株進一步分析,期望能更了解KOR 在纖維素合成的 角色,將來可應用於木材的產業利用。
研究方法
1. 選殖KOR cDNA 由於已知的KOR 可由木質部取得,本計畫也以赤桉嫰葉等 組織的RNA為材料,企圖選殖出不同KOR 成員。根據已發表的基因庫阿拉伯 芥,楊樹與松樹KOR 基因,設計位於不同物種間保留區域(conserved region) 的引子,以RT-PCR 的方式取得數個KOR 基因片段。經由RT- PCR取得的赤 桉KOR 基因片段(命名為EucKOR 等)將放入TA 載體進行定序與序列比對。
所獲得的EucKOR 基因片段定序後,以GenBank 的blastn 找出全長序列的候 選EucKOR 基因,同時可藉由其它軟體輔助取得初步比對序列資料。
2. 選殖KOR genome DNA 以葉片gemone DNA為材料,由獲得的KOR cDNA 序 列中設計出不同引子對,以期獲得基因全長,藉由PCR 的方式取得KOR 基 因片段。
3. 選殖全長KOR cDNA
i. EucKOR 3’端的互補DNA(cDNA) 將以3’RACE 技術完成。以種間保留區為正 股引子(sense primer)位置,另外反轉錄產物已帶有anchor 的poly-T 為反股引 子(antisense primer)。在RT- PCR 反應中即加入種間保留區為正股引子(sense primer)與adapter 專一性的反股引子。
ii. EucKOR 5’端的互補DNA 區將使用SMART RACE Kit (Clontech)完成。參照 套組的操作手冊,經由PCR 反應,經由數次反應與引子設計,以克隆出5’端 的cDNA。
iii. 為了確定這些EucKOR 基因為同一核酸序列,將從3’及5’端的未轉譯區域設 計正股引子及反股引子,則可得到全長的EucKOR 基因。預計全長約為2.4kb 左右,ORF約為1.8kb,而全長基因將由primer walking 的方式完成定序。
4. 定量RT-PCR 為取得EucKOR 在不同組織部位的基因表現,將萃取木質部、
嫩葉、靭皮部與根部組織的RNA並反轉錄為cDNA,以種間變異區為PCR 引 子,經由兩個步驟定量出的數據進行分析。
5. EucKOR 之雙元載體構築 (binary vector) 根據上述實驗取得的全長EucKOR cDNA,經評估後先將其置於持續作用的啟動子(CaMV 35S promoter)之後,以 正轉式方向插入以pBI121(Jefferson 1987)為架構的雙元載體。
6. 農桿菌為媒介的基因轉殖
EucKOR
基因進入菸草與赤桉雙元基因載體將用於基因轉殖
EucKOR
基因赤桉的生產。主要將以臺灣林業試 驗所轉殖系統為主,將培植體與農桿菌液共培養,以抗生素初步由培養基篩 選出可能帶有EucKOR
基因的組織。經由一系統的癒合組織培養、器官形成等6
將可獲得再生且帶有
EucKOR
基因的植株。7. 定期全程生長與形態觀察
由於基因轉殖KOR 基因,可能會改變碳水化合物生合成,而轉殖植物於癒合 組織形成到植株再生的過程都可觀察其形態或生長變化,所以將記錄芽的生 長抽長,芽數量等介質可提供KOR 轉殖苗在進入田間隔離試驗的參考。
8. 瓶苗進行PCR 基因轉殖分子鑑定
由分子鑑定技術,包含PCR 選出帶有合成纖維素相關基因(KOR)的瓶苗,進一 步微體繁殖產生瓶苗。
結果與討論
目前已成功選殖出KOR 之cDNA全長,其中所設計之引子位置與所獲得之 基因長度如表1與圖1所示。由於KOR基因為EGase巨大家族成員之一,在選殖過 程中需克服不斷選殖出類纖維素合成酶Csl之問題,經由數次引子設計與克隆,
最終選殖出一個片段cDNA,此片段基因命名為EucKOR7,其片段長度為950bp。
由於此區域含KOR專一性序列之基因變異區,經由此片段序列方可不斷往5’與3’
設計引子並克隆基因。在3’端的克隆部份,經由一次PCR即獲得包括UTR之序列;
然而在5’端之選殖又出現不同EGase成員,致使朝5端的克隆每次僅能獲得約 200bp之片段資訊。經由大量克隆篩選之後,最終獲得之全長資訊約為2,400bp。
為確保所獲得之EucKOR全長cDNA為完整存於赤桉之基因,且可後續成為 genomic DNA克隆之骨架,試驗中又設計位於UTR區域數個引子,企圖將完整 ORF選殖出(如圖1,表1所示)。
本計畫所獲得之全長EucKOR其長度為2,379bp,其中包含ORF為1,869bp,預 期由623個胺基酸組成,具有5’與3’ UTR分別為149bp與361bp (圖2與圖3)。為確 定已克隆之cDNA與預期序列相符,此序列已經過雙向定序,並確定序列無缺失,
且可經由軟體完整轉譯出胺基酸序列,以利後續基因轉殖工作。實驗之初,以木 質部與嫩葉為材料,嘗試選殖出不同EucKOR之成員,但試驗期間尚無法在嫩葉 中選殖出EucKOR基因片段,初步判斷此基因在嫩葉表達量極低,未來在研究工 作可負擔之狀況下,將再嘗試選殖不同成員。所選殖出之全長EucKOR與已克隆 出之3端片段藍桉之相似度為95%,目前所有林木基因庫中僅有楊樹已選殖出 KOR全長cDNA,故將顫楊、阿拉伯芥、稻米等涵蓋雙子葉與單子葉之ORFs比對,
結果顯示不同物種間之相似度由57%~78%不等(表2,3),所分析出之親緣關係如 圖4所示。 KOR之genomic DNA全長選殖工作已完成,此部份工作所費時間遠比 估計長,主要是因為Kor基因為 cellulase 隻巨大家族成員,而序列上又與 Csl 相 近,所以在基因的選殖上,需要嘗試不同的引子對經過許多次的PCR擴增,才能 選殖出對應Kor基因的序列。全長之Kor genomic DNA已完成雙向定序,其中 CSRII 區已成功篩選出並定序完成,其cDNA長度為945bp,而此區基因為1849bp,
具有三個exons 與四個introns。目前與阿拉伯芥之基因結構相同,全長約 3 kb,
具有六個exons與五個introns(圖5)。
為了解Kor基因在不同植物組織的表達量,我們設計不同的引子進行
real-time PCR定量與半定量PCR,結果如圖6所示。初步結果顯示,EucKOR基因 於根部表現量最高,其次為嫩葉,而在靭皮部與木質部的表達量最低;此試驗不 論是用半定量PCR或是real-time PCR,其結果一致,但根部樣品之誤差值較大,
且材料取得不易,未來將再以組織培養苗之根部RNA萃取,期能獲得較具意義 之根部基因表現。除此之外,使用不同區域之引子進行定量PCR,發現靠近5’端 的引子其專一性最高,可忠實呈現EucKOR基因在不同部位之表達模式。而研究
8
人員也利用相同引子進行原位雜交試驗,但因EucKOR表達量較低,無法經由目 前的原位雜交資料了解不同細胞間表達的差異。第三年的赤桉基因轉殖工作,所 遭遇的困難在於將基因轉入雙元載體 pBI121,我們利用PCR增加全長 EucKOR 的限制酶切位,以利雙元載體之構築,因pBI121載體之連接位點少,質體產量低,
且載體較大,所以第三年計畫雖如期進入基因轉殖,同時也增加菸草之基因轉殖 希望可以更早觀察與分析Kor基因於植物體中可能扮演的角色。將菸草及赤桉以 農桿菌介導轉殖法進行基因轉殖後,由於菸草生長非常快速,因此在培養兩至三 周就已經有芽體的產生,且在培養四周時,基因轉殖株的葉片已經成長至8到9 公分且具有3到4對葉片(圖7),已可用來抽取gDNA,因此將葉片進行gDNA的萃 取,並且利用PCR的方式,利用引子對(KOR_AS11和KOR_S11 & NOST2和35SP1) 嘗試將KOR片斷擴增出來,目前實驗進行至此尚無片段的產生,未來,將嘗試 以其他不同的PCR條件,包含改變引子對、Tm值、Taq的改變或是gDNA的重新 萃取,持續嘗試將KOR片斷擴增出。同時,這些轉殖菸草也帶有GUS基因,所 以將會持續以GUS染色再確定可能的轉殖株。赤桉也已進行六次,但皆在第二個 月出現嚴重褐化;經改善共培養時間,於近期開始有癒合組織出現 (圖8),此與 本實驗室其他纖維素相關基因轉殖結果相似,癒合組織之出現比對照組緩慢。本 計畫已經以農桿菌介導轉殖法,重複對菸草及赤桉進行多次的基因轉殖,希望可 以增加轉殖苗木,以利未來分析與實際應用。
迄今文獻尚無任何桉樹全長KOR cDNA序列發表,本計畫已於今年(2011)將 EucKor 基因之cDNA上傳至NCBI資料庫,一旦論文發表後將會開放其他研究學 者使用。而本計畫所選殖出KOR之ORF與其它物種相似度約為75%,而與AtKOR2 則僅有57%,是否意味在桉樹中仍有不同作用功能之其它成員存在,這將是值得 探討的現象。近期所刊出之文獻,已將KOR基因與CesA基因合併研究,而本實 驗室已於今年完成CesA相關研究資料,相信在論文發表上應可取得較好評價。
本計畫已訓練三位專題生,兩位畢業同學已進入其他國立大學研究所就讀,由於 此國科會計畫之支持,方能達成人才培育之目的。
參考文獻
1.
甘偉航、胡大維、楊政川。五種桉樹在東臺地區造林適應性初步試驗。林業 試驗所研究報告季刊。1990;5(2):99-109。2.
陳正豐、楊政川、張添榮、洪富文。二十種桉樹在台灣造林適應性試驗。林 業試驗所研究報告季刊。1995;10(3):283-292。3.
汪大維、王兆桓。本省桉樹人工林幼齡生長和收穫之研究。台灣林業科學。1996;11(2):123-136。
4.
陳正豐、洪富文、張添榮。三種桉樹在台灣東部地區生長之研究。台灣林業 科學。1996;11(3):233-238。5.
陳文茜。文茜世界周報。廣西造林運動創造綠色奇蹟。2011 Oct.網址http://www.youtube.com/watch?v=oj6onxDUgjc
6.
Bhandari S, Fujino T, Thammanagowda S, Zhang D, Xu F, Joshi CP (2006) Xylem-specific and tension stress-responsive coexpression of KORRIGAN endoglucanase and three secondary wall-associated cellulose synthase genes in aspen trees. Planta. 2006; 224(4):828-37.7.
Balatinecz JJ, Kretschmann DE, Leclerc A. Achievements in the utilizationof poplar wood—guideposts for the future. For Chron. 2001;77:265-269.8.
Brown Jr RM, Saxena IM, Kudlicka K. Cellulose iosynthesis in higher plants.Trends Plant Sci. 1996;1:149-156.
9.
Delmer DP. Cellulose biosynthesis: exciting times for a difficult field of study.Annu Rev Plant Physiol Plant Mol Biol. 1999;50:245-276.
10.
Festucci-Buselli1 RA, Otoni1 WC, Joshi CP. Structure, organization, and functions of cellulose synthase complexes in higher plants. Braz J Plant Physiol.2007;19:1-13.
11.
Jarvis MC. Interconversion of the Iα and Iβ crystalline forms of cellulose by bending. Carbohydr Res. 2000;325:150-154.12.
Jianru Zuo, Qi-Wen Niu, Naoko Nishizawa, Yan Wu, Benedikt Kost, and Nam-Hai Chua.KORRIGAN, an Arabidopsis endo-1,4-ß-glucanase, localizes to the cell plate by polarized targeting and is essential for cytokinesis. Plant Cell.2000; 12:1137-1152.
13.
Herth W. Arrays of plasma membrane “rosettes” in cellulose icrofibril formation of Spirogyra. Planta. 1983;159:347–356.14.
Lerouxel O, Cavalier D, Liepman A, Keegstra K. Biosynthesis of plant cell wall polysaccharides-a complex process. Current Opinion in Plant Biology.2006;9:621-630 3.
15.
Maloney VJ, Mansfield SD. Characterization and varied expression of a membrane-bound endo-beta-1,4-glucanase in hybrid poplar. Plant Biotechnol J.10
2010; 294-307
16.
Molhoj M, Pagant S, Hofte H. Towards understanding the role of membrane-bound endo-beta-1,4-glucanases in cellulose biosynthesis. Plant Cell Physiol. 2002; 43(12):1399-406.17.
Nicol F, His I, Jauneau A, Vernhettes S, Canut H, Hofte H. A plasma membrane-bound putative endo-1,4-beta-D-glucanase is required for normal wall assembly and cell elongation in Arabidopsis. EMBO J. 1998; 17(19):5563-76.18.
Rajai HA. The structures of cellulose. Washington D.C.: American Chemical Society; 1987.19.
Somerville C. Cellulose synthesis in higher plants. Annu Rev Cell Dev Biol.2006;22:53-78.20.
Wickholm K, Hult EL, Larsson PT, Iversen T, Lennholm H. Quantification of cellulose forms in complex cellulose materials: a chemometric model.Cellulose.2001;8:139-148.
附表
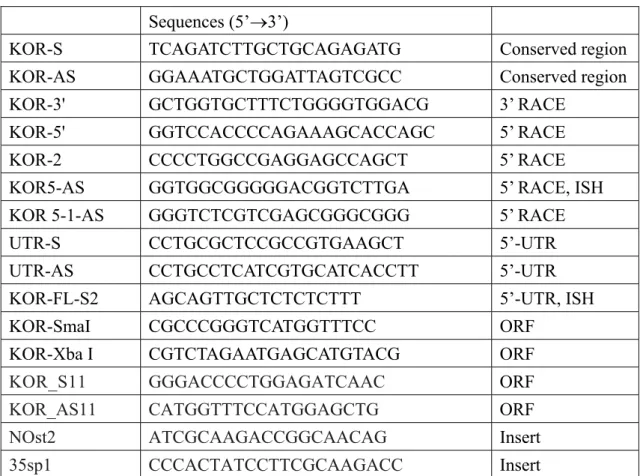
表 1. 用於選殖與分析 Kor 基因與基因轉植苗之引子與探針
Sequences (5’3’)
KOR-S TCAGATCTTGCTGCAGAGATG Conserved region KOR-AS GGAAATGCTGGATTAGTCGCC Conserved region KOR-3' GCTGGTGCTTTCTGGGGTGGACG 3’ RACE KOR-5' GGTCCACCCCAGAAAGCACCAGC 5’ RACE KOR-2 CCCCTGGCCGAGGAGCCAGCT 5’ RACE KOR5-AS GGTGGCGGGGGACGGTCTTGA 5’ RACE, ISH KOR 5-1-AS GGGTCTCGTCGAGCGGGCGGG 5’ RACE
UTR-S CCTGCGCTCCGCCGTGAAGCT 5’-UTR UTR-AS CCTGCCTCATCGTGCATCACCTT 5’-UTR KOR-FL-S2 AGCAGTTGCTCTCTCTTT 5’-UTR, ISH KOR-SmaI CGCCCGGGTCATGGTTTCC ORF
KOR-Xba I CGTCTAGAATGAGCATGTACG ORF
KOR_S11 GGGACCCCTGGAGATCAAC ORF KOR_AS11 CATGGTTTCCATGGAGCTG ORF NOst2 ATCGCAAGACCGGCAACAG Insert 35sp1 CCCACTATCCTTCGCAAGACC Insert
12
表 2. EucKOR cDNA 與其他物種之相似度比較表
Accession
number U37702 CAB51903 AAC49704 AL078637 AC001229 EucKOR U37702.1
(At Kor OR16)
72% 66.8% 65.4% 58.3% 73.7%
CAB51903
(Bn Kor) 72% 89.7% 87.5% 74.6% 73.3%
AAC49704
(Sl Kor) 66.8% 89.7% 84.4% 74.3% 74.3%
AL078637
(At T22A6) 65.4% 87.5% 84.4% 73.3% 69.8%
AC001229
(At F5114) 58.3% 74.6% 74.3% 73.3% 63.9%
EucKOR 73.7% 73.3% 74.3% 69.8% 63.9%
表 3:EucKOR amino acid 與其他物種之相似度比較表
Accession
number U37702 CAB51903 AAC49704 AL078637 AC001229 EucKOR U37702
(At) 98.2% 88.7% 85% 72.3% 89.4%
CAB51903
(Bn) 98.2% 89% 85.4% 72.4% 89.6%
AAC49704
(Sl) 88.7% 89% 82.1% 71.3% 91.1%
AL078637
(At) 85% 85.4% 82.1% 70.4% 81.9%
AC001229
(At) 72.3% 72.4% 71.3% 70.4% 74.4%
EucKOR 89.4% 89.6% 91.1% 81.9% 74.4%
*At: Arabidopsis; Bn: Brassica napus; Sl: Solanum lycopersicum
A T G C C G G C T G T C T C T G C A C G C G C A C A T A G C A T T T T C G A C C T G T C A G C G G G T A C A G G T T G C C T T G T T G A T C G A C C G G A G A G G C T C T C G T C C
G A G C A T G T A C G A G G A C G A G C T C G A C G A G A C C G A C C T G G G C T G C G G C C T C C A C G G T C C C C C C T C T G G G A A G A A G T C C G A T T T C A A G T T C A A C G C C A A G T A C T A C T T C C T C A C T G G G G A T A C C A T T G A T T A T G C T G C T T T A G A C G G T G C C A A G G A A G C T G C A T T G T A C T A T G A T G C T G G T G C T G C T C A G G T G C T C A G G A C A T T C A A T A G A A C T G T C A A T G C A T G G T A C T G C G A T T A C A T C C T G A A A C A T G T G G G A T G G A A A T G T C C C G A T A A C G C A G G G A A T G A C A A G A A C A C A T G G A A A C C
圖1 EucK
C G G A C G G G A C G C A G C C G C C C C A G C A G G C T G C A T C A T C T C G C C G G C C C G A C A A C T G C T C C C C A A T C G T C C T G G A C T T C C C G A C G A G G C C G C A A G A C C T T C C T T C T G G A G T A A A C G G C C G C T T C T G C A A A A C G C T G T A A C T T T C T A G C A A C C G G C C T T T C T G G G G C T C C T A A G T T T C A C A A T C C A A A G G T G A G C C T T C C T G G A C C C A C T T T G G G A A A A G C A T C A C A G T G G A G G G A C A G C A T G A C G T G C A G G A C T A C A A T A T T C C A T G A
圖
KOR 基因選
A C C C C T G G G C A A C C T C A A G G C T G G C T C T C G T C A G C C C C T C A T C A C T A C A C C C T C A G C A T A A C A G G T G C T G A A A A G T C T T T C C C G G G G A G C C A A T T C C T C G G A A G C A C T C T G T C T T T G A T C C A T T G T T T C C A T T T T A T A A C T C C A C A A T A A C T C G G T G G A C C A G C C G T C T G A T C A G G C C A G G G C T T G A T C T G G C C A C T G T T T C T A T T C A A T C C T C G G G G G G T G C C T C A C A T C G A A G G C T T C C A T T G G T T G C C G C T C T G C G G T
圖2. EucKO
附圖
選殖引子位置
G G G G A C C C C C G A C T A C G A C T C G G C C A G G C A A G A T C T C C T C A T C G T G C C C T C C A C A T G T G T C G T G G G T C T G G T G C C A T G A C C T C G C C C A C G T G C C G A C A C A C C C C A A A T C T T G T A G C A T T T C A A A G A T C G A G G C A A C T A G T T A T T T T A T T T G C A G A T T A T G G T G G C T G T T C C C A T A A T C A T C A G T T G A A C T A T T C A G T G T A C T G A T G T A A A A T G A G C C C A T C C C A A G C C A A A C C C G A C G T C C G T C A C T C G T G G T C C T C C C A T
OR cDNA O
置與RACE
C T G G A G A T C A C C G C G C C G G G G G G A G C A T T C G T C T G G T C A A G A C C G C A A G G C C C T T G G A G G G G C T G G G A G G G T C A T G C T C A G G T C A A G G A G C C A T C G A T A T G A T C A T T A A G T T G C T C A A C A A C A A G G A C A G C G A G G T G G G A T G A G A A C T T G C T A T G T A C T T A G C T C A G C C C T T G T G C T C G T C C A T G G A A G G A C T A C C T T T C T T G C G T G C T A C A T C G T A A G A A T A A G C G A A C A C G T T A C T A A C T A G C C T T A T C A T G T T T C C C A
RF 全長序列
E 相對長度
A A C G C C G C C G C C C T C T C C A A G A A G A A G A A G A C C G T C G G C G T C C C C C G C C C A T G T T C T T A A C T C C G G C A C T A C G A C G G C T G G A G C G T G A T C A T C A A G A A C A T C G C T G A C T G C T G G A T A G A T C T T G C T G C C T A C T C T C G G A G A T A C A G G T T T G T C T G G A C C A C T C C T G G C T G G G A C A A G G G T A T C C A A C C T G C C T G G A C C C C A G C C G A T G C T G C G G A T T T T G C C A C G G C T T T G G G G T C A A G T A C T G G T G G G A G A C A A C T A C A C A G G C G A T G G G A C T C C G C C G C
列
C G A C T C C G C A A C T C C C G C A G A A G A A G T C T C C A T C C T C A C C A C G G C T C A A C G C C C C C T T C A A G A G C C G G G G A C T C A T C G A G T G T G G G G C A C G C T C A G G T C T G C G T C C C G T G C T G A G A T C A G A A G C T A G T G C T G G T T G G G C G G G G C G G T A T C G C A A C A A G C T T G A T A C G A G G A G T C T T C A C A C C C T T C A A T G G A T A C T C C A A G A G G C A G G C A A T C A T T C A A C T G C A A G C A A T G G T T C A G A A C C C A G A G C G C T A A C C A C C C C C A
C C A C C C T A C T C G C A A C A G A C G C G C T A C C C G C G G G A A T G G A G T T C A C T T A A A G C T A A A A A G T A T C T G G A T T A C A A G T G C A C T A G A A G C
M S N S W T L H V L Y E G S A A R G Y L S R R T P G F G N T A L
圖4 拉伯
M Y G R D R P L D E V G S I L K A L M F K G L V G A A G E L G D T S G E M A A A R Y S A G Q L A T T L R L F L K G G L I W Y C G P N H Y P K L V G A M V A L S G
4 不同植物 伯芥、SL:蕃
P W G G P T Q Q G W V C G L L F N A Q R G Y Y D A A H V K E G S T T P L A S A S S A E A A P G I A K S P G Y P Q L N H G T F Y S T H V H H R V A G P D D G S A K
物間 KOR cD 蕃茄、PT:顫
L E I N A L L G Q G A G L I T S G K L P G D A I K I I K W G N D H Y C I V F K D T F Y N S H A G A F Y E E M L R P Q P L D V L R D G A S I P K H D G F I D K N T
圖3. EucK
DNA ORFs 顫楊、GH:棉
14 A D S A T E Q K K K L I V K T K H N N V F N F P K T D Y F L W M R P E N K A Y S T S Y W D W G G P D R T F H N Q Y V V N F A K R Q K N K V K H D V R T I F S A V
KOR amino a
之Phylogr 棉花。
E D E R S K K Y V D V P R H H S W R G N S F A M T K T F N S D I D Y K Q K L V H E F V W G Y G V L S Q A S I I A A F L A I D Y I L Y N C K G N Y N Y T P P M F P
acid sequence
ram。 Eu:
R N L N D L G C I I G K H G P S G L Q D M L S W S S A D T I R P V F A G A K T L G A W L Y W D N K L M C S Y L T V F S D G K N P R G W K W R E P T L A T P P P P
e
赤桉、ZM
Y D R A A V S R K I P D N Y T G K S D S V I E Y S D N I A A A C S S C S F H F S R Y A T G N A G A Q V P V F T S Y L D A A R K M S Y I R D T S K P A G N A G L P A P W K
M:玉米、AT
A L S F V L A S W A K A Q V D L R Q Q N S V L L F N A D T V G N P V A K P
T:阿
ATGAGCATGTACGGACGGGACCCCTGGGGGGGACCCCTGGAGATCAACGCCGCCGACTCCGCCACCGAGGACGAGC GCAGCCGCAACCTCAACGACTACGACCGCGCCGCCCTCTCCAACTCCCGCCCGCTCGACGAGACCCAGCAGGGCTG GCTCCTCGGCCAGGGGGAGCAGAAGAAGAAGAAGAAGTACGTCGACCTGGGCTGCATCATCGTCAGCCGCAAGATC TTCGTCTGGACCGTCGGCTCCATCCTCGTCTGCGGCCTCCTCGCCGGCCTCATCACCCTCATCGTCAAGACCGTCC CCCGCCACCACGGCAAGCACGGTCCCCCCGACAACTACACCCTCGCCCTCCACAAGGCCCTCATGTTCTTCAACGC CCAGCGCTGTAAGCCTCTTCCCACCCTCGCAGATCTGGCCATCCCACTTCAGGACTCGTACACTTCCCATCTGGGT TTTTCCTAATTTGTTGTGTTGTGGATTAAATGAGCGATCTTGGATAGTGTAGAGACTGTCTACTTGCTATTTTAAG CAAAAATTTCGGTTTTTTTTTAAACGTTTGCAGCTGGGAAGCTCCCCAAGCATAACAATGTGTCGTGGAGGGGCAA CTCCGGCCTTCAAGACGGCAAGTCCGATTCGTCCTGGGTGCTGAAGGATCTGGTGGGAGGGTACTACGACGCCGGG GACGCCATCAAGTTCAACTTCCCGAAGTCTTTCGCCATGACCATGCTCAGCTGGAGCGTCATCGAGTACAGCGCCA AGTACGAGGCCGCCGGGGAGCTCGCCCACGTCAAGGAGATCATCAAGTGGGGCACCGATTACTTCCTCAAGACCTT CAATTCCTCTGCCGACACCATCGATAACATCGCTGCTCAGGTACTGAAGTGGATAGAGTGATGTGTTATGACATCG CTAGCGGAGAATTTACCATTTCTGGGTCACTTCTGATTACCTAGCTAAAGGAAGAAAAATCTCGCCTCTAATTTGC TTATATATGCCACCTTGAGTGGAATTAAGTGTAAATGAACAAGATCCGGGCCAAACAATTGAATCTTAAATTGTTA AGTCCAACCTAAGTTTCACCCGATGGCCCGAAAAGCTGTTTGAAAAACATTGATTGTCAAGATCAAAGATTGCTTA GGATTGTGGCAGCTTTCAATTTAACTTTCTAGCCTGAGTGTTTGGGGAGAAGACTCGGTGGACTCCCCTGGGTGTT ACATTTCCAAGAAAATCCTGTTTGATTTCTTCAGATTGAAATGCTTCTCGTAGAGATCCCTATCGCATTTTCACCA AATTCTATTTTAATTGATTGCCCCATTTGATCGATTTGCACCTGTTTTCCCCTTTTTTCATCAGGTCGGTTCTGGG GATACTTCTGGAGGAAGCACTACCCCAAATGATCATTACTGCTGGATGCGTCCCGAAGACATTGATTATAAACGGC CTGTCTTTGCTTGTAGCAGTTGCTCAGATCTTGCTGCTGAGATGGCTGCTGCTTTAGCTTCTGCATCCATTGTTTT CAAAGACAACAAGGCCTACTCTCAGAAGCTAGTTCACGGTGCCAAAACGCTGTTCCATTTTTCGAGGCAACAGCGA GGGAGATACAGTGCTGGTTCAGCGGAAGCTGCAACTTTCTATAACTCCACTAGTTATTGGGATGAGTTTGTCTGGG GCGGGGCTTGGTTGTACTATGCAACCGGCAATAACTCTTATTTGCAACTTGCTACCACTCCTGGTATCGCAAAACA TGCTGGTGCTTTCTGGGGTGGACCAGATTATGGTGTACTTAGCTGGGACAACAAGCTTGCTGGTGCTCAGGTAAGT TTAGTCTCTCCTAGAGGTCTTTCCCCCCTTGTTATTGAGATATATGCCTAACATGCTACTTATCATTCAGTTGTCG ATATGTTAAAGGCTGACATACTATGTCCATGTTTGAAAATCCAAAAGGTGTATAATTTGCCACTCTTTCAGATCTA AAAGTATCTCTAGCAAGTTTGGTCACTGAGCCATTCAATTATCCTTAAATGCCAACTGTTCATGTTCTTGAGGAGT TAAATAAGGCAACTGCTCCATTACAATGCAAAGATCTTTGTATTTGAATTCATTTCATGTAGGAAGGAATGAGAAA AAAAACGAGAGCCCTCCATGCACTCAAAGTGAAAACTGTTGACCCATCTCTCTGTGAAACTCTTTTGAGGGGGGAC CTTGACTGGATGCTGTGGTAGCTTCAGAAACTGATTCATCTGTCTTCACTGTACCCAACCCTTAACTCTAATGCAT GAGATGTGACTGGATTTGTTTGGTCCATATGCATGCAGCAGCAGATGCTCTATTGATTTGTCTTTTGTTGAGAGAG ATGGGTTTAGGAGGTATTAAGTTCATCATTTTGAAGCATAAACTCTGAACAATGCAATGACCTTGATATACCTGTC GCAAACTTGTAGGTGCTCCTAAGCCGTCTGAGGCTGTTCCTCAGCCCTGGGTATCCATACGAGGAAATGCTCAGGA CATTTCACAATCAGGCCAGCATAATCATGTGCTCGTACCTGCCTGTCTTCACAAGCTTCAATAGAACCAAAGGTAA TGTCTCTTCAGACATGGCATAGGGTGGAAGGGGACGACAAATGCATGTAGTGATGAAACTAGTTGGCAGCTCAGTC TCATGTTCCGTTTTTGCCTTGGTTACAGGTGGCTTGATCCAGTTGAACCATGGAAGACCCCAGCCCCTTCAATATG TTGTCAATGCAGCCTTCCTGGCCACTGTATTCAGTGACTACCTTGATGCTGCGGATACTCCTGGATGGTACTGCGG ACCCACTTTCTATTCTACTGATGTCTTGCGTGATTTTGCCAAGAGGCAGGTATGTCATTTCACGCCTCTCAATTTT TCTAAAAACATTTTGGTATTGCCTCTTTTTTTTTTTTTTAGCTAAGATGCTGAAAAACAATGGATGATAACTCCAC CTGTGATTCTCTTATGGGATCTGTAGATCGATTACATCCTTGGGAAAAATCCTCGGAAAATGAGCTACATCGTTGG CTTTGGCAATCATTACCCGAAACATGTGCATCACAGGGGTGCCTCCATCCCAAAGAATAAGGTCAAGTACAACTGC AAAGGAGGATGGAAATGGAGGGACACATCGAAGCCAAACCCGAACACGTTGGTGGGAGCAATGGTTGCAGGTCCCG ATAAGCATGACGGCTTCCATGACGTCCGTACTAACTACAACTACACAGAACCCACTCTCGCAGGGAATGCAGGACT GGTTGCCGCGCTCGTGGCCTTATCAGGCGATGGGACCGCTAAGATCGACAAGAACACAATATTCTCTGCGGTTCCT CCCATGTTTCCCACTCCGCCGCCACCTCCAGCTCCATGGAAACCATG
圖5 EucKOR gDNA 序列與結構,其中包括六個 exons (彩色區域) 與五個 introns (無底 色區域)。
(A)
(B)
圖6 PCR
6. EuKOR R 之定量結
R mRNA 於 結果。
於不同組織中
16
中之表現量量。(A) 半定定量方式與與(B) Real-tim
me
圖 77.轉殖菸草 導出芽體
草在不同時期 體
期階段的記記錄,經過數
數次不定期的的繼代菸草草由癒傷組織織誘
圖 88.此為轉殖 有抗生素
殖赤桉在不同 素培養基中兩
同時期階段 兩個月。
18
段的記錄,經經過繼代培養養,赤桉之之癒合組織已已在