國立臺灣大學工學院應用力學研究所 碩士論文
Graduate Institute of Applied Mechanics College of Engineering
National Taiwan University Master Thesis
槲皮素對肝癌的影響
The effect of Quercetin in hepatocellular carcinoma
陳界甫 Jie-Fu Chen
指導教授:陳兆勛 博士 Advisor: Chao-Hsun Chen, Ph.D.
中華民國一零一年七月
July, 2012
i
致謝
本論文研究過程中,承蒙吳師 陳兆勛教授悉心指導,詳細的修改與校閱指正,
以及國立中山大學生物醫學所 戴明泓老師和其博士班學生 儲天輝學長在研究上 的建議,使得本文得以順利完成。另外還要感謝 戴明泓老師以及長庚大學醫檢系 賴信志教授撥冗參加本人的碩士學位考試,並給予肯定。在此獻上最衷心的敬意 與感謝。
此外,在台大研究所求學期間,有歡笑也有淚水,感謝同是奈米複材實驗室 的同學 徐東陞、陳文祥、陳忠澤,我們互相學習、互相勉勵及互相支持,一同在 求學過程中成長茁壯。另外也要感謝我大學時的摯友 簡齊毅、康肇元的一路支持 與陪伴,最後要感謝父母的栽培和不曾間斷的關愛。謹將本文獻給所有愛護我的 人。
ii
摘要
肝癌在全世界是一種常見的癌症之一,在台灣也是導致許多癌症病患的死亡 原因。為了提高肝癌病人的存活率,或降低手術過後的復發率,尋找有效的抗癌 藥物是重要並且需要的。而廣泛存在於植物界的槲皮素因為具有抗癌變及抗增生 的效果,且有強大的抗氧化能力以及在細胞訊號傳導、細胞週期、細胞凋亡的調 控上有一定的能力而受到注意。本研究目的是希望藉由相關實驗,來觀察槲皮素 是否能抑制人類肝癌細胞株Hep3B、Huh7,並以基因晶片做檢測分析,針對被調 控到且會影響細胞增殖生長、細胞週期、細胞移動及細胞凋亡的基因做進一步探
討,闡明槲皮素抑制人類肝癌細胞的藥理機制。此實驗以人類肝癌細胞株Hep3B
及Huh7 為材料進行體外培養,以各濃度槲皮素對細胞進行 Proliferation、Colony formation、Wound-healing 及 Flow cytometry 實驗,並以槲皮素影響效果較顯著的 Huh7 作進一步微陣列分析。實驗結果顯示,Hep3B 及 Huh7 在 48 小時槲皮素濃度 分別為500、50μM 時,開始有顯著抑制細胞增殖效果;在濃度 50、10μM 時,
顯著抑制細胞菌落發展;在同樣濃度100μM 時,經過 12 小時就開始有顯著抑制 細胞移動效果以及濃度分別在50、10μM 能分別誘導 Hep3B、Huh7 產生細胞凋 亡,且對細胞週期G1 phase 都有阻滯效果,以上實驗效果都與槲皮素濃度成正相 關;在來從進一步的Huh7 微陣列分析實驗結果發現,槲皮素能調控 Cell apoptosis、
Cell cycle、Cell migration 相關基因,而這些受到調控的基因可能參與了 Apoptosis pathway、JAK/STAT、PI3K/AKT、MAPK/ERK pathway。而這些被影響調控的基 因以及所參與的訊號傳導路徑就是槲皮素能夠抑制癌細胞生長增殖生長、細胞週 期、細胞移動以及誘導凋亡背後最強而有力的證據。未來槲皮素或許可用於臨床 上治療肝癌之輔助治療藥物,提高肝癌病人的存活率、降低化療藥物所產生的副 作用以及降低手術過後的復發率。
關鍵詞:肝癌、槲皮素、細胞凋亡、細胞週期、基因晶片
iii
Abstract
Hetapocellular carcinoma is one of the common cancers in the world, it is also the main reason that makes people die in the cancer. To increase the survival rate or reduce the recurrence rate, it is necessary and important to find those efficacious anti-cancer drugs. Quercetin is common existing in plantwith great effect of anti-carcinogenesis and anti- proliferation. It also has good capability on anti-oxidant. Besides, Quercetin has been noticed by the abilities on effecting cell signal transduction and regulating of cell cycle and inducing cell apoptosis. The purpose of this research is trying to observe whether Quercetin can inhibit HCC cell growth by several experiments. Moreover, to find that the mechanism of inhibiting the proliferation、growth、cell cycle、migration and inducing apoptosis on HCC cell by Quercetin, we used microarray so that we can analyze the suppression effect on HCC cell by gene expression. This experiment uses HCC cell line- Hep3 and Huh7 as the materials in vitro culture, and different concentrations of Quercetin are using on experiments of Proliferation, Colony formation, Wound-healing and Flow cytometry. Huh7 is used on microarray analysis due to the better influence on Quercetin as well. Experiment results show that Hep3B and Huh7 have significantly inhibiting proliferation when the concentration of Quercetin is 500, 50 μ M on 48 hours; it also shows inhibition on colony formation while the concentration is 50、10μM. And Quercetin has good inhibition effect on cell migration after 12 hours when the concentration is 100μM, and it can also induce Hep3B、Huh7 apoptosis amd arrest G1 phase in cell cycle when the concentration is 50、10μM. All effect by Quercetin during the experiments is depending on the concentrations of Quercetin. Moreover, the experiment of microarray analysis for Huh7 also shows that Quercetin can regulate the gene relative to Apoptosis、cell cycle and migration。And these gene regulated by Quercetin may be involve Apoptosis pathway and several signal
iv
transduction pathway:JAK/STAT、PI3K/AKT、MAPK/ERK pathway. All the effect on signal transduction pathway or gene regulated by Quercetin are the strong evidence that Quercetin can inhibit the growing of cancer cells、arrest cell cycle、suppress cell migration and induce the apoptosis of cancer cells. Perhaps people can use Quercetin on clinical medicine for curing liver cancer, increase the survival rate and also decrease the side effect and recurrence rate.
Keywords:Hetapocellular carcinoma, Quercetin, Apoptosis, Cell cycle, Microarray
v
目錄
第一章 前言 ... 1
第二章 文獻回顧 ... 3
一、 肝癌 ... 3
二、 槲皮素 ... 6
三、 細胞凋亡 ... 10
四、 細胞週期 ... 14
五、 基因晶片 ... 17
第三章 材料和方法 ... 20
一、 實驗材料 ... 20
1. 細胞培養 ... 20
2. 細胞計數 ... 22
3. 藥物處理 ... 23
4. Alamar Blue cell viability reagent ... 23
5. 流式細胞儀 ... 25
6. Affymetrix® Human Genome U219 Array ... 26
二、 實驗方法 ... 27
1. Cell proliferation assay ... 27
2. Colony formation assay ... 27
3. Wound-healing assay ... 28
4. Flow cytometry assay ... 28
5. Microarray analysis ... 29
vi
第四章 結果 ... 31
一、 Cell proliferation assay ... 31
二、 Colony formation assay ... 32
三、 Wound-healing assay ... 32
四、 Flow cytometry assay ... 33
五、 Microarray analysis ... 34
第五章 討論 ... 36
第六章 結論 ... 41
參考文獻 ... 62
vii
圖目錄
FIG.1Two major apoptotic pathways in mammalian cells. ... 12
FIG.2 人類肝癌細胞株 ... 20
FIG.3 血球計數器 ... 22
FIG.4 血球計數器上的方格 ... 22
FIG.5Resazurin 還原成 Resorufin ... 24
FIG.6Resorufin 的吸光度與放射螢光光譜 ... 24
FIG.7AlamarBlue protocol ... 24
FIG.8 細胞經雷射光激發產生的散射光及螢光 ... 25
Fig. 9 槲皮素對 Hep 3B 的 Proliferation assay 結果 ... 42
Fig. 10 槲皮素對 Huh 7 的 Proliferation assay 結果 ... 43
Fig. 11 槲皮素對 Hep 3B 的 Colony formation assay 結果 ... 44
Fig. 12 槲皮素對 Huh 7 的 Colony formation assay 結果 ... 45
Fig. 13 槲皮素對 Hep 3B 的 Wound-healing assay 結果 ... 46
Fig. 14 槲皮素對 Huh 7 的 Wound-healing assay 結果 ... 47
Fig. 15 槲皮素對 Hep 3B 的 Flow cytometry assay 結果 ... 48
Fig. 16 槲皮素對 Huh 7 的 Flow cytometry assay 結果 ... 49
Fig. 17 RNA Electropherogram ... 50
Fig. 18 RNA Purity test ... 51
Fig. 19 aRNA Quality Control ... 51
Fig. 20 Microarray Image of Huh7-0μM ... 52
Fig. 21 Microarray Image of Huh7-50μM ... 53
FIG.22 槲皮素對肝癌調控兩倍以上基因之影響 ... 61
viii
表目錄
Table. 1 槲皮素上調 2 倍以上 Apoptosis 相關基因 ... 54
Table. 2 槲皮素下調 2 倍以上 Apoptosis 相關基因 ... 56
Table. 3 槲皮素上調 2 倍以上 Cell cycle 相關基因 ... 57
Table. 4 槲皮素下調 2 倍以上 Cell cycle 相關基因 ... 58
Table. 5 槲皮素上調 2 倍以上 Migration 相關基因 ... 59
Table. 6 槲皮素下調 2 倍以上 Migration 相關基因 ... 60
1
第一章 前言
自民國七十一年起,癌症死亡率已連續三十年位居國人十大死因之首位 (行政 院衛生署,2012)。而肝細胞癌(hetapocellular carcinoma;HCC)更為我國男性與女 性癌症死亡原因的第一及第二位。肝癌一般分為原發性肝癌與轉移性肝癌兩種,
原發性肝癌是指由肝臟內的細胞經由癌化而形成惡性腫瘤,轉移性肝癌則是由其 他器官的癌細胞經由轉移到肝臟而形成。原發性肝癌通常與慢性肝炎所造成的肝 硬化有關,而有80%是因為B型肝炎病毒(HBV)、C型肝炎病毒(HCV)所導致,另外,
暴露於致癌因子,或者吸菸、喝酒也會引起肝癌的產生。目前治療肝癌最有效的 方法,是透過手術摘除腫瘤,但對於腫瘤較大或多發性的肝癌,卻有一定的手術 治療限制。在經過手術摘除腫瘤後,肝癌的復發率也是相當高。因此,為了提高 肝癌病人的存活率,或降低手術過後的復發率,尋找有效的抗癌藥物是重要並且 需要的。近年來研究發現,大部分的抗癌藥物均以造成癌細胞走向細胞凋亡路線 為其藥理作用,而開發可選擇性促進癌細胞凋亡的藥物是目前抗癌藥物研究的新 方向。可運用分子醫學如癌細胞與正常細胞在訊息傳遞上的差異來開發新的分子 藥理學作用機轉,透過改變癌細胞訊號傳導的狀態以調節基因表達,抑制細胞增 殖或誘導腫瘤細胞凋亡,標靶藥物對於腫瘤的治療,被認為是合理而且有希望的 策略。
槲皮素是一個典型的黃酮化合物(flavonoid),廣泛存在於植物界。因為具有抗 癌變及抗增生的效果,且有強大的抗氧化能力以及在細胞訊號傳導、細胞週期、
細胞凋亡的調控上有一定的能力,而受到注意。而根據諸多文獻顯示,槲皮素可 以透過不同的機制來達到抑制癌症的效果,包括抗氧化作用、抑制會活化致癌物 的酵素、 訊息傳遞途徑的修正、和受體及一些蛋白質的交互作用。近年來研究發 現,大部分的抗癌藥物均以造成癌細胞走向細胞凋亡路線為其藥理作用,而開發 可選擇性促進癌細胞凋亡的藥物是目前抗癌藥物研究的新方向。槲皮素可以透過
2
影響細胞凋亡路徑上的受體誘導細胞凋亡,例如誘導肺癌細胞的death receptor 5(DR5)表達,造成癌細胞毒殺,或是抑制表皮生長因子受體表達,抑制大腸癌細 胞生長及誘導凋亡;也可以透過調控細胞凋亡路徑上的基因,例如透過Survivin 和Bcl-2 蛋白下調和 p53 上調,激活 caspase-3 和 caspase-9,增加 Bax 易位到粒腺 體模,誘導凋亡;除了促進凋亡路徑上的表達,也可以抑制AKT、ERK 生存訊號 的表達,阻止細胞生長。
隨著人類基因體研究計畫展開後,傳統的核酸分子雜交技術所花費的時間與 效率早已不足以應付大量基因訊息的產生。隨著半導體技術的進步,人們也提出 了結合生物科技和半導體技術的構思。1990 年代,史丹佛大學結合了探針固相原 位合成技術(in situ synthesis)和照相平版印刷技術(Photolithography),首先製造出世 界上第一張基因晶片,可以一次同時分析大量的基因資訊。基因晶片擁有高統計 輸出量、操作容易、分析信賴度及精確性高、使用檢體樣品少、應用範圍廣可獲 得整體性的實驗數據等優越特性,解決了傳統操作技術繁雜、耗時耗工、低效率 之不足。
本研究目的是希望藉由細胞增殖、菌落發展、細胞移動及細胞凋亡實驗,來觀 察槲皮素是否能抑制人類肝癌細胞株Hep3B、Huh7,並以基因晶片做檢測分析,
選擇兩倍上調與兩倍下調的基因作Ontology 分析,以 Biological process 與 Molecular function 的分組,從中挑選與 Apoptosis、Cell cycle、Migration 有關的基因做初步 探討。瞭解基因可能參與的訊號傳導途徑及所造成的生物功能影響,闡明槲皮素 對人類肝癌細胞抑制增殖生長、細胞週期、細胞移動及促進凋亡背後的藥理機制。
3
第二章 文獻回顧
一、 肝癌
肝癌(Hepatocellular carcinoma,HCC)在全世界是一種常見的癌症之一,在台 灣也是導致許多癌症病患的死亡原因[1]。根據行政院衛生署之相關數據顯示,自 民國七十一年起至今惡性腫瘤一直居於十大國人死因之首位,其中肝癌又為國內 癌症病患死亡肇因之第一或第二名。根據流行病學研究調查統計,肝癌發生之相 關危險因子有性別差異、地理區佈、慢性肝炎感染與否及生活形態等。在性別方 面,男性較女性為罹患肝癌之高危險群;而以地理分佈觀之,東南亞、南非及一 些西非國家有較高之罹患率;另開發中的國家有較高於已開發國家二至三倍的肝 癌罹患率;在慢性肝炎方面如病毒性的B 型及 C 型肝炎感染;生活形態方面如酗 酒、抽煙或化學致癌物的攝入,如黃麴毒素[2][3],均為使肝癌罹患率提高之危險 因子。肝癌一般分為原發性肝癌與轉移性肝癌兩種,原發性肝癌是指由肝臟內的 細胞經由癌化而形成惡性腫瘤,轉移性肝癌則是由其他器官的癌細胞經由轉移到 肝臟而形成。原發性肝癌通常與慢性肝炎所造成的肝硬化有關,而有80%是因為 B 型肝炎病毒(HBV)、C 型肝炎病毒(HCV)所導致[4]。感染B 型肝炎或 C 型肝炎之 後,有一部份人變成慢性肝炎,慢性肝炎再演變為肝硬化,最後再形成肝癌[5][6]。
致癌因素
1. B 型、C 型肝炎病毒
肝炎的發生會長期不斷地破壞肝臟組織,使肝臟逐漸纖維化,並經由再生而 形成肝硬化,在此發炎、硬化的過程中,肝細胞的複製及再生發生了問題,因而 產生肝癌細胞。B 型肝炎的患者約有 15~40%會發展成肝硬化或肝癌[7],在台灣,
臨床上有八成以上的肝癌病人曾經被HBV 所感染[8]。HBV 為 DNA 病毒曾有研 究發現B 型肝炎病毒的 DNA 會嵌入宿主基因中,可能因此導致宿主 DNA 受損
4
或影響宿主的基因表現而引發肝癌[9]。HCV 為 RNA 病毒,一般認為 HCV 造成 肝細胞癌化的主因為HCV 的病毒蛋白。許多研究指出 HCV 的病毒蛋白能透過調 控細胞內的訊息傳導途徑來抑制細胞死亡與增加細胞生長[10][11]。
2. 黃麴毒素 (aflatoxins)
黃麴毒素主要是由真菌黃麴菌(Aspergillus flavus)及寄生麴菌(Aspergillus.
parasiticus)所產生的代謝產物[12]。台灣因氣候潮濕,儲藏不良的花生、玉米等是
黴菌的溫床。在許多的動物實驗中已證實黃麴毒素可以引發動物的腫瘤[13]。
3. 酗酒
過度飲用含酒精的飲料會引發酒精性肝炎與肝硬化。,而 B 型與 C 型肝炎病 毒帶原的病患飲用酒類則會增加2 到 4 倍罹患肝癌的機率[14]。
4. 其他因素
服用過量類固醇、動情素或荷爾蒙,還有抽煙及受到化學物質暴露等等,曾被 指出與肝癌有關[15]。
治療方法
其治療方法不外乎手術切除、肝臟移植、化學藥物治療、免疫治療、放射線 治療、冷凍治療、基因治療、肝動脈拴塞、酒精局部注射等[16][17]。目前治療肝 癌最有效的方法,是透過手術摘除腫瘤,但對於腫瘤較大或多發性的肝癌,卻有 一定的手術治療限制。在經過手術摘除腫瘤後,肝癌的復發率也是相當高[18]。肝 臟移植仍是目前治療肝癌的方法中,五年存活率最好的[19],但不是每個肝癌病患 都可以等到相符合的肝臟可以做移植。而化學治療是將一些抗癌藥物利用靜脈注 射的方式使其進入全身循環而殺死癌細胞。5-fluorouracil(5-FU)和 carboplatin 目前 為治療肝癌上廣泛使用的化療藥物。5-FU,類似尿嘧啶(uracil)的相似物,是一種 抗代謝物[20]。而carboplatin 能引起 DNA 的破壞造成毒殺細胞的效果[21]。但是 這些用於高劑量治療的抗癌藥物,會造成腫瘤細胞的抗藥性及嚴重的副作用。1995);
5
肝動脈拴塞與局部酒精注射則受限於肝腫瘤的大小與分布,另外病人的肝功能必 須正常才會有較好的療效[22][23]。
因此,為了提高肝癌病人的存活率,或降低手術過後的復發率,尋找有效的 抗癌藥物是重要並且需要的。可運用分子醫學如癌細胞與正常細胞在訊息傳遞上 的差異來開發新的分子藥理學作用機轉,透過改變癌細胞訊號傳導的狀態以調節 基因表達,抑制細胞增殖或誘導腫瘤細胞凋亡,標靶藥物對於腫瘤的治療,被認 為是合理而且有希望的策略。天然植物抗癌或防癌成分之研究也是未來努力的新 方向。本文目的在於探討存廣泛存在於植物界的槲皮素,在抑制人類肝癌細胞過 程中之藥理機轉,尋找在癌細胞在訊號傳導中被調控的基因,闡明槲皮素在抗癌 方面的作用機制。
6
二、 槲皮素
化學名
2-(3,4-Dihydroxyphenyl)-3,5,7-trihydroxy-4H-1-benzopyran-4-one 3,3',4',5,7-Pentahydroxyflavone
結構
分子式:C15H10O7
分子量:302.24 g/mol
簡介
槲皮素(Quercetin)是一個典型的黃酮化合物(flavonoid),廣泛存在於植物界。
黃酮類化合物為多酚類化合物,主要由diphenylpropane (C6–C3–C6)當作基本骨架。
槲皮素對預防癌症來說是一種具有吸引力的自然化合物,因為具有抗癌變及抗增 生的效果,且有強大的抗氧化能力以及在細胞訊號傳導、細胞週期、細胞凋亡的 調控上有一定的能力。槲皮素通常當作苷元(glycoside),有相當多的量可以被消化 道吸收並進行後續代謝轉換[24],在腸子吸收後會轉變為glucuronide/sulfate
7
conjugates,這些代謝產物可在血管循環系統中發現。而槲皮素的代謝產物 glucuronide 也被證實在人類肺癌細胞株 NCI-H209 中,能透過促凋亡蛋白的上調 以及抗凋亡蛋白的下調來促進細胞凋亡,抑制癌細胞的增殖[25]。槲皮素及其他相 關黃酮類化合物廣泛存在於水果及蔬菜,因具有淺力的抗癌特性而受到注意。他 們可以透過不同的機制來達到預防癌症的效果,包括抗氧化作用、抑制會活化致 癌物的酵素、 訊息傳遞途徑的修正、和受體及一些蛋白質的交互作用[26][27][28]。
抑癌機制
1. 與受體交互作用
槲皮素可與一些受體發生交互反應,特別是芳烴受體(aryl hydrocarbon receptor,
AhR),AhR 負責調控與可活化致癌前驅物的 CYP1 家族的基因表現[29],被認為 與肺癌的發展及大腸癌有關[30]。Death receptor 與細胞凋亡路徑有關,槲皮素可以 誘導肺癌細胞death receptor 5(DR5)表達,造成癌細胞的毒殺[31]。ErbB 蛋白家族,
又稱表皮生長因子受體(EGFR)家族,過度的 ErbB 訊號與各種腫瘤的形成有關,槲 皮素能降低ErbB2 和 ErbB3 的表達,抑制大腸癌細胞(HT-29)的生長和誘導凋亡
[32]。
2. 影響訊息傳導途徑
許多研究報告指出,槲皮素與相關類黃酮化合物在與癌變過程中的訊號傳導 途徑有很大的關係。包括細胞週期的調控、細胞凋亡、發炎和血管新生。例如,
槲皮素誘導肝癌細胞HepG2 凋亡是透過兩種途徑,粒腺體途徑及抑制細胞生存訊 號途徑Akt/ERK[33]。在人類白血病細胞(HL60)中能誘發 DNA 的裂解,Bax 的上 調以及Bcl-2 的下調[34]。Cyclooxygenase-2(COX-2)的異常表達和發炎及腫瘤形成 有重要的關係,過度的表達已被證明在一些癌症的侵襲有高度正相關[35] 。槲皮 素在乳腺癌細胞(MDA-MB-231)中透過抑制 p300 信號,抑制 COX-2 的表達和 PGE2
8
的產生以及COX-2 啟動子的活性,且在 primary human umbilical vein endothelial cells (HUVECs)可顯著抑制由血管內皮生長因子(VEGF 165)所誘導的血管新生[36]。
Phospholipase D (PLD)被認為與細胞增殖、腫瘤生成有關,而槲皮素能透過誘導 Phospholipase D1(PLD1)的下調及抑制 metalloproteinase-2 (MMP-2)活性,抑制人類 神經膠質瘤(U87 glioma cells )的細胞增殖及入侵[37]。槲皮素還能透過抑制Leptin、
Leptin receptor protein, Leptin mRNA and Leptin receptor mRNA 的表達,抑制人類胃 癌細胞(MGC-803)的細胞增殖。許多研究都可以證明槲皮素確實可以透過各種不同 的機制來達到預防癌症的效果。
槲皮素與肝癌
槲皮素加上TNF-related apoptosis-inducing ligand (TRAIL)能加速對
TRAIL-resistant 的肝癌細胞凋亡,透過顯著上調 DR5(TRAIL 的 death receptor)的 mRNA 和蛋白水平,及下調 cFLIP(caspase 8 抑制劑)的蛋白水平,提高了 TRAIL 在各種肝癌細胞中的敏感性,促進肝癌細胞的凋亡[38]。槲皮素也可以加上加上重 組的腺病毒載體(表達 p53, GM-CSF and B7-1 的基因),讓抑制肝癌細胞(BEL-7402, HuH-7 and HLE)增殖效果更佳[39]。除了透過細胞凋亡Death Receptor 途徑外,槲 皮素也可以透過促進細胞凋亡粒腺體途徑,透過caspase-3 和 caspase-9 的激活,增 加Bax 易位到粒腺體模,誘導細胞凋亡,以及抑制 Akt and extracellular regulated kinase (ERK)生存訊號,讓槲皮素能依劑量依賴性的誘導 HepG2 死亡[40]。還可以 透過Survivin 和 Bcl-2 蛋白下調和 p53 上調,促使細胞凋亡[41]。而在抑制細胞週 期方面,用流式細胞儀分析發現細胞週期G0/G1 phase 增加,S phase 下降,subG0/G1 上升,槲皮素對肝癌細胞(HA22T/VGH and HepG2)有劑量依賴性的抑制效果,且 annexin V 增加,加上化療試劑 paclitaxel 對肝癌細胞造成凋亡的效果更好[42]。在 動物實驗方面,利用NDEA(誘導大鼠肝癌)加上槲皮素,發現槲皮素是通過減少氧 化應激和減少抗氧化劑活性,發揮其在肝癌上的預防作用[43]。槲皮素目前也被拿
9
來搭配治療肝癌的化療藥物使用,例如化療藥物5-fluorouracil (5-FU) and
carboplatin 會使 Hsp27、Hsp40 上調,因此搭配槲皮素(熱休克蛋白 Hsp27、Hsp40 抑制劑),用 western blot 分析發現 pro-caspase 3 水平降低。兩者聯合能有效增加對 HepG2 與 Hep3B 的毒殺[44]。槲皮素還可以加上2-methoxyestradiol(hormonal agents),增加 annexin V 的 binding,造成 MnSOD RNA 的上升,對肝癌細胞 (HA22T/VGH and HepG2)有更佳的毒殺作用[45]。除了搭配化療藥物,也可以搭配 roscovitine(CDK 抑制劑),透過抑制 Akt pathway 下調 caspase3、caspase9,增加對 HepG2、Hep3B 的凋亡效果[46]。另一方面,Wnt pathway 的突變及失調可能導致 許多疾病,在癌變過程中扮演重要角色,槲皮素在Huh7 扮演 Wnt 抑制劑的角色,
針對Wnt 下游負責傳遞訊號的 βcatenin 作抑制[47]。
10
三、 細胞凋亡
在多細胞生物中,細胞的死亡有兩種不同的形式:第一類是Necrosis(細胞壞 死),第二類是 Apoptosis(細胞凋亡)。Necrosis 主要是因為外在因素如局部貧血、
高熱以及物理、化學損傷和生物的侵襲,造成細胞極速死亡而結束其生命。
Apoptosis 又稱為程序性的細胞死亡,它是用來描述伴隨細胞死亡的一系列形態學 上特定的變化[48]。在希臘文中apoptosis 為樹葉或花瓣凋落的意思[49]。它是指像 秋天樹葉凋謝一樣,細胞在一定的生理或病理條件下,遵循自身的程序,自己結 束生命的過程,最後細胞脫落離體或裂解為若干凋亡小體(apoptotic bodies)而被其 他細胞吞噬。
形態特徵
細胞凋亡的形態特徵與細胞壞死的形態特點不同。細胞壞死的形態特徵首先 是膜通透性增加,細胞外形發生不規則變化,內質網擴張,核染色質不規則的位 移,進而粒線體及核腫脹,溶酶體破壞,細胞膜破裂,胞質外溢。這種細胞死亡 過程常常引起炎症反應[50]。而細胞凋亡,是細胞內在的有規律的機制引起。主要 是受細胞本身之DNA 調控而作用,主要目的在消除老化、受傷及 DNA 受到破壞 的細胞。當細胞收當凋亡訊號時,細胞首先變圓,與周圍細胞脫離,胞質濃縮,
內質網擴張成泡狀病與細胞膜融合,粒線體無大變化,核染色質密度增高成半月 形,並凝聚在核膜周邊,核仁裂解,核固縮,進而細胞膜內陷將細胞自行分割為 多個外膜包裹,內含物不外洩的的細胞凋亡小體。由於這種過程不導致溶酶體及 細胞膜破裂,沒有細胞內含物外洩,固不引起炎症反應。生化特徵為細胞核內的 DNA 會被核酸內切酶分解 180-200bp(base pair),細胞膜上的 PS 磷脂(Phosphatidyl serine)外翻,這是讓巨噬細胞辨識細胞凋亡的訊息,當此訊息出現時,巨噬細胞會 進行吞噬作用吞噬apoptotic bodies[51]。與壞死(necrosis)程序最大的區別在於細胞
11
凋亡並不會引起周圍組織的發炎[52]。
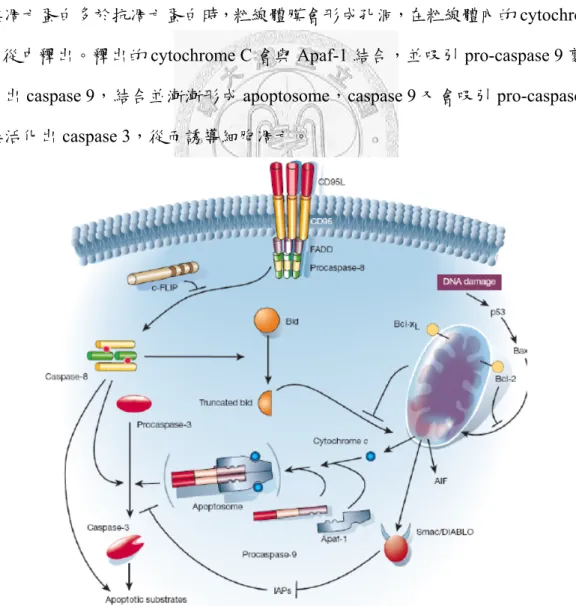
訊號傳導路徑
細胞凋亡傳導路徑主要分為兩條(如 Fig. 1)[53]:第一條是Death receptor pathway,第二條是 Mitochondrial Pathway。而細胞凋亡最後的訊息傳遞最後都是 會匯集到caspase 蛋白來執行細胞自我凋亡。Caspase 蛋白(Cysteine-Aspartate Specific ProteASES)屬於 cysteine protease 蛋白家族,在哺乳動物主要是以沒有活性 的pro-enzyme (zymogen)形式存在,當 caspase 受到 pro-apoptotic 訊息刺激時,其 結構會發生改變而變成有活性的caspase[54]。已活化的caspase 會切斷細胞內的結 構蛋白質,也會破壞某些參與代謝的酵素和訊息傳遞的蛋白質。Caspase 大致上可 分為兩類:一類為執行者(effector),如 caspase-3、6、7,它們可直接降解(degradation) 結構蛋白和功能蛋白,引起凋亡,但不能通過自我催化,或自我剪接的方式活化;
另一類為啟動者 (initiator),如 caspase-8、9,當細胞受到凋亡訊息後,能透過自 我剪接的方式活化,然後引起caspase 連鎖反應,如 caspase-8 可依次活化 caspase-3、
6、7[55]。
1. Death receptor pathway
Death receptor pathway 是由胞外腫瘤壞死因子(TNF)家族的配體如 TNF-α、
FasL/CD95L、TWEAK、TRAIL 與細胞膜上的受體結合(分別是 TNF-R、Fas/CD95、
DR3、DR4/5)。配體與受體結合後會活化細胞膜下的 death domain,開始吸引 FADD(Fas-Associated protein with Death Domain)或 TRADD(Tumor necrosis factor receptor type 1-associated DEATH domain protein),FADD 又會繼續吸引 pro-caspase 8 來結合形成複合物,稱為 death-inducing signaling complex,DISC。pro-caspase 8 在DISC 中局部濃度升高,會開始自我剪接並活化成 caspase 8,活化的 caspase 8 釋放到胞質中啟動caspase 的級聯反應,激活下游的 caspase 3,caspase 3 則會切割 ICAD/CAD complex,使兩者分開,讓 CAD 切割 DNA 形成降解導致細胞凋亡。而
12
活化的caspase 8 能使胞質中的 Bid 斷裂成 tBID,tBID 會轉移到粒腺體上,誘導粒 腺體釋放cytochrome C 進入胞質,開啟 Mitochondrial Pathway,有效增加了細胞凋 亡的訊號。
2. Mitochondrial Pathway
Mitochondrial Pathway 除了由 Death receptor pathway 活化的 Bid 傳遞凋亡訊息,
另一條受控於Bcl-2 家族蛋白 (B cell lymphoma-2)。Bcl-2 家族蛋白群依據 Bcl-2 homology domain (BH)的不同可大致區分為:(1)antiapoptotic proteins (包括: Bcl-2、
Bcl-xL、Bcl-w、mcl-1);(2)proapoptotic proteins (包括: Bax、Bcl-Xs、Bad、Bak、
Bid)[56]。當Bad 收到凋亡訊息,會被釋放出來與抗凋亡蛋白因子 Bcl-2 結合。若 促進凋亡蛋白多於抗凋亡蛋白時,粒線體膜會形成孔洞,在粒線體內的cytochrome C 會從中釋出。釋出的 cytochrome C 會與 Apaf-1 結合,並吸引 pro-caspase 9 剪接 活化出caspase 9,結合並漸漸形成 apoptosome,caspase 9 又會吸引 pro-caspase 3 剪接活化出caspase 3,從而誘導細胞凋亡。
Fig. 1 Two major apoptotic pathways in mammalian cells.
13
細胞凋亡除了受到Death receptor pathway 及 Mitochondrial Pathway 的基因調控 外,p53 也是一個很重要的調控因子。p53 是一種分子量 53kDa 的轉錄因子,位於 人體第17 對染色體上。在正常的情況下,p53 的表現量很低,但當 DNA 異常時,
p53 蛋白及轉錄活動明顯增加,並刺激 p21 蛋白的表現,使細胞停留在 G0/G1 期 而不能進入S 期,以進行 DNA 修補[57],直至 DNA 修復後才進入正常的週期。
若DNA 損傷嚴重無法修補時,則誘發細胞走向凋亡途徑,防止細胞癌化,因此 p53 有腫瘤抑制基因之稱。
大多數抗癌藥物,如拓撲異構酶抑制劑、抗代謝物和激素拮抗劑等,都可在 不同類型的腫瘤細胞中誘導細胞凋亡。抗癌藥物的療效不僅取決於這些藥物與各 自特意靶細胞的相互作用,也取決於這些藥物誘導細胞凋亡的能力。因此,腫瘤 細胞對細胞凋亡的敏感性是決定化療效果的關鍵因素,如何在治療中促進腫瘤細 胞凋亡已成為新的化療目標和篩選抗癌新藥的標準。
14
四、 細胞週期
細胞增殖是細胞生命的一個基本特徵,單細胞生物(細菌和酵母)以此來繁衍後 代,而多細胞生物(如高等真核生物)通過細胞增殖完成個體發育。細胞增殖與機體 的生長、在生、修復、細胞凋亡腫瘤發生都有密切關係,它也是細胞分化形成組 織、器官、系統的基礎。細胞生長到一定階段開始分裂,細胞分裂後產生的新細 胞生長增大,此後又平均的分裂成兩個遺傳性相同的子細胞。這種周而復始的生 長分裂週期稱為細胞週期(Cell Cycle)。換言之,細胞週期是指親代細胞分裂結束稻 子代細胞分裂結束所經歷的過程,而這個過程所需的時間稱為細胞週期時間。細 胞週期基本任務就是確保DNA 在合成期(S phase)時有確實複製一次,以及兩個相 同的染色體在有絲分裂期期(M phase)時有被平均的分配到子細胞中[58]。
細胞週期與檢查點
細胞週期可分為間期(Interphase)和有絲分裂期(Mitosis)。間期又可分為三個時 期:G1(Gap1 phase)、S(Synthesis phase)、G2(Gap2 phase)。當細胞進入 G1 期時,
細胞代謝活化,複製所需胞器以及一些細胞質的組成,以供下一階段複製染色體 使用。細胞在檢查過大小、生長因子和DNA 後,即通過 G1 checkpoint,進入 S 期 開始進行DNA 複製。複製完後進入 G2 期,此時細胞已具有兩倍的遺傳物質,並 為有絲分裂期作準備。當細胞確定DNA 完全複製後,即通過 G2 checkpoint,並進 入有絲分裂期(M phase)。細胞在有絲分裂期時,進行核裂(染色體分離)和質裂(細 胞質分裂)。細胞在 M 期時,也有一個 checkpoint,稱為 Spindle assembly checkpoint,
主要檢查染色體是否附著在紡錘體上。Checkpoint 對細胞週期調控相當重要,在 G1 和 G2 checkpoint 時發現 DNA 受損或複製不完全,細胞就會進入 G0 期,暫時
停止分裂,待DNA 修補完後在進入下一個時期。若細胞週期失去調控,造成染色
體變異,導致部分基因流失、重組,往往會演變成癌細胞,無限制的分裂形成腫
15
瘤。另外,細胞週期也受到生長環境(如細胞密度)的影響。當培養基長滿細胞後,
細胞即進入G0 期,不會進行分裂,若去除其中一些細胞,細胞又會開始進入細胞
週期直到長滿。
調控因子
在細胞週期中除了受到checkpoint 調控[59]之外,還受到兩個關鍵調控因素,
週期素(Cyclin)和週期素依賴激酶(cyclin dependent kinase,Cdk)[60]。這是由2001 年諾貝爾生理醫學獎得主利蘭.哈特威爾(Leland Hartwell)、保羅.納斯(Paul Nurse) 和蒂莫‧漢特(Timothy Hunt)的研究所發現。
1. 週期素 (Cyclin)
在 1983 年時,學者藉由蛙卵與海膽的研究,發現了一些會隨著細胞週期進 行而分解又出現的蛋白質,濃度在有絲分裂前增加,有絲分裂後消失,由於這種 蛋白質周期性的出現,因此將它命名為週期素cyclins (指週而復始出現的蛋白 質)[61],之後許多學者紛紛展開對週期蛋白的研究,在不同的生物體中分離出數 十種的週期蛋白。目前已發現的週期素有數十餘種,如酵母細胞中Cln1、Cln2、
Cln3 及哺乳動物細胞中的 Cyclin A、B、C、D、E、F 及 H 等。依據它們在細胞週 期中調控階段的不同分別歸為(1)G1 期週期素(G1 cyclins 或 START cyclins),如 Cyclin C、D、E,(2)M 期週期素(mitotic cyclins),如 Cyclin A、B。而 Cyclin H 則 會和 CDK 7 形成一具有酵素活性的複合物,進而活化 cdc 2 (CDK1) 及 CDK2。
在週期素分子中都含有一段高度保守的氨基酸序列稱為cyclin box,負責辨認不同 的cyclin-dependent kinases (CDKs)並與之結合,才能在調控細胞週期中發揮作用。
2. Cyclin-dependent kinases (CDKs)
Cyclin-dependent kinases (CDKs)是一群蛋白激酶家族,會和特殊的Cyclin 結合 而活化。目前已知共有10種CDKs 被發現,分別是cdc 2 (CDK1)、CDK2、CDK3、
16
CDK4、CDK5、CDK6、CDK7、CDK8、CDK8、CDK10。在真核細胞中,藉由連 續的訊息,經過活化,與隨後一系列特殊的Cyclin-CDK 複合物(Cyclin-CDK complex)來調控細胞週期進行,讓細胞增生。例如,Cyclin D 與CDK4 形成之複 合體與G1 期的進行有關,Cyclin D 的表現及其與CDK2 的結合則是與細胞進入S 期相關;Cyclin A與CDK2 主導了S 期的進行,而Cyclin B 與CDK1 則和細胞進 入M 期的調控有關。
細胞週期與癌細胞
由於細胞週期是一相當複雜的過程,因此小心且綿密的調控乃細胞分裂是否成 功的保證,如果此一調控機制在任一環節失去控制,細胞即可能發生不正常之生 長調節、未成熟分裂,甚且影響到後續的細胞生理及細胞代謝,也可能導致個體 特定細胞數不正常增生,或許因此導致腫瘤之發生[62]。對大部分癌細胞而言,它 們在檢查點的調控作用通常是有缺陷的[63]。還有p53 的突變在癌細胞中也是很常 見[64]。一旦這些調控的蛋白質表現異常,導致癌細胞不受控制,則會不斷地進行 細胞週期而增殖分裂。因此,在抑癌機轉方面,槲皮素如何影響癌細胞之細胞週 期,用於癌症之預防與治療之可能方向,調控細胞週期之機轉、細胞週期素(cyclin)
或細胞週期素依賴激酶(cyclin dependent kinases, Cdks)表現如何受到調控以及抑 制癌症活性物質在細胞週期中扮演之角色等之探討,都是此實驗的研究課題。
17
五、 基因晶片
生物體中基因的表現千變萬化,在瞭解生命體的生理發育,病理發展過程中,
扮演一個非常重要的角色。人體約有38,500 個基因,隨著人類基因體研究計畫展 開後,傳統的核酸分子雜交技術所花費的時間與效率早已不足以應付大量基因訊 息的產生。隨著半導體技術的進步,人們也提出了結合生物科技和半導體技術的 構思。在1980 年代,Bains 將短的 DNA 片對固定在支持物上,藉由雙股 DNA 分 子雜交(Hybridization)進行序列測定[65]。1990 年代,史丹佛大學結合了探針固相 原位合成技術(in situ synthesis)和照相平版印刷技術(Photolithography),首先製造出 世界上第一張基因晶片[66],可以一次同時分析大量的基因資訊,解決了傳統操作 技術繁雜、耗時耗工、低效率之不足。
基因晶片之分類
依據基因探針點印固定於晶片表面方法的不同,可分成兩種,一種是原位合 成(in situ)的基因晶片,又稱寡核苷酸晶片(oligonucleotide chip)[67],在晶片的特定 部位合成寡核苷酸而製成。第二種是先合成DNA,再由機器點印(printing)到晶片 上。
第一種原位合成技術又分為兩類。(一)光引導原位合成技術(light-directed in situ synthesis)[68]。由史丹佛大學發明,Affymetrix 公司採用。製作方式是利用光 罩蝕刻(Photolithography)的方式,進行核苷酸的合成,將長度約 25 個鹼基的核苷 酸合成在約一平方公分大小的晶片上。(二)壓電打印法(piezoelectric printing)。其方 法是用噴墨打印機將特定種類的四種鹼基合成試劑噴印到晶片的預定區域上,然 後再沖洗、去保護,進行寡核甘酸合成的下一個步驟[69]。第二種晶片外合成探針,
是先用分子生物學技術合成探針,在利用機械打點法點印到晶片上去[70]。
18
基因晶片之原理
基因晶片之原理,是將大量已之基因序列之基因探針,依照特定排列方式,
固定在載體,然後將特殊標記處理後之檢體RNA 與基因晶片上之探針進行雜交
(hybridization)。DNA 四種鹼基 A、T、C、G 會根據鹼基配對法則,AT 與 TA、CG
與GC 配對,雜交訊號強弱代表基因表現強度。最後由電腦及分析軟體,對數以萬
計的基因進行表現量分析。不論是細胞生命週期、生化調控路徑等基礎研究,還 是藥物研發中對於藥物作用位置的篩選,臨床的疾病診斷預測,癌症基因的檢測 等,都是基因晶片可以應用的範圍[71][72][73]。
Affymetrix 晶片介紹
Affymetrix 公司乃是寡核苷酸微陣列晶片的先驅,他們的晶片,有多項特有的設 計,較以往傳統的微陣列晶片,有更準確的實驗結果,將其特點分別敘述如下:
1. 光罩蝕刻法(Photolithography)
過去微陣列晶片需依賴自己實驗室持有的有限基因樣本作為基因探針,數量 及品質有時候不能完全符合實驗要求。光罩蝕刻法可自行設計探針,在一小晶片 中探究生命體所有基因變化。
2. 探針組(Probe Set)的採用
在生物演化上表現的基因常會出現類似轉錄樣本,以往在實驗室有限的基因 樣本中所選擇的探針,其專一性及代表性不夠,造成非專一性雜交,容易高估訊 號表現量,影響到分析結果的可信度。因此,採用多個探針來偵測基因的表現與 否變成為可以減少訊號錯誤的一個方式。
3. 完美配對及不完美配對探針設計(Design of perfect match/mismatch probe pairs) 設計一對獨特的25 mer 寡核苷酸探針,其中一個是完全與 Target sequence 互 補的完美配對,另一個則是除了第13 個核苷酸不互補配對外其餘序列與 Target
19
sequence 互補配對。此種設計可針對每一個點均有相對的陰性控制點,作背景值計 算,提高探針的靈敏性及特異性,尤其在檢測低表現量時更能增加訊號判斷的正 確性。
本實驗利用基因晶片高統計輸出量、操作容易、分析信賴度及精確性高、使 用檢體樣品少、應用範圍廣可獲得整體性的實驗數據等優越特性[74─79],探討人 類肝癌細胞在槲皮素作用下,細胞的基因表現影響及其作用機轉。
20
第三章 材料和方法
一、 實驗材料
1. 細胞培養
Cell line
人類肝癌細胞株Hep3B、Huh7,如Fig. 2左為Hep3B;右為 Huh7。
(ATCC,由國立中山大學生醫所戴明泓老師提供)
Fig. 2 人類肝癌細胞株
Medium
DMEM+10%CS+1%(penicillin +streptomycin)+1% L-glutamine
1. 將血清瓶經高壓滅菌釜滅菌及烘箱烘乾後放入 Laminar flow 開 UV 殺菌。
2. 使用量筒裝入 1 公升三次水,並加入 13.4g DMEM、3.7g NaHCO 、5.98g Hepes Acid Free,放在 Stirrer 上開啟自動攪拌。
3. 使用 PH meter 調溶液 PH 值為 7
4. 在血清瓶上裝 Filter 過濾剛泡好的 DMEM 溶液,分裝至血清瓶各 450ml 5. 加入 50ml(10%) CS、5ml(1%)(penicillin +streptomycin)、5ml(1%) L-glutamine,
放入4℃冰箱
21
解凍培養
1. 用 pipet 吸取 medium 10cc 至 dish
2. 將細胞從-80℃冰箱取出(含有 medium 及 DMSO),放入恆溫水槽 37℃下解凍 變液體
3. 加入 1cc 細胞(Hep3B p20,Huh7 p19)至 dish,稍微搖晃,用顯微鏡觀察細胞 呈現懸浮,放入incubator(37℃,95% air +5%CO ) 24hr
4. 從培養箱取出前天培養細胞的 dish
5. 用 suction 吸乾 medium,glass pipet 吸取 PBS 10cc,加入 dish(對 wall),稍微 搖晃(洗掉死細胞)
6. 用 suction 吸乾 PBS,glass pipet 吸取 medium 10cc 加入 dish(對 wall),放入 incubator
繼代培養
1. 用 suction 吸掉 medium
2. 先用 10ml PBS wash cell (洗掉含有血清的 medium,因為血清會降低 trypsin 作 用),用 suction 吸乾
3. 加入 trypsin 1ml 至 dish,shake 後放入 incubator 2-3 min,取出並拍打 4. 加入 5ml medium 至 dish,在用 pipet 吸起放入離心管(共有 6ml) 5. 將兩管離心管對秤放入離心機,1500rpm,5min
6. 用 5ml PBS wash dish ,suction 吸乾,加入 10ml medium 至新 dish 7. 離心管離心後,用 suction 吸乾上清液
8. 加入 500μl medium 至離心管,並拍散(subculture 1:5)
9. 從離心管吸取 100μl 加至新 dish,放入 incubator 繼續 maintain cell,剩餘細胞 實驗用
22
細胞冷凍
1. 用 trypsin 將細胞打下
2. 置入離心機離心 1500rpm,5min,吸乾上清液 3. 加入 4.5ml CS 及 0.5ml(10%) DMSO
4. 分裝各 1ml 至冷凍管,冰在−80℃冰箱
2. 細胞計數
1. 96 well 的其中 3 孔各加入 100μl trypan blue(稀釋用),稀釋時取 100μl 未知濃 度cell,在孔中吸放 10 次, 為一次稀釋,連續二個孔作吸放動作為第二次稀 釋,連續三個孔作吸放動作就是三次稀釋
2. 放入血球計數器(如 Fig. 3),在顯微鏡下計算在 4 個 4×4 方格(如 Fig. 4)內的總 細胞數
Fig. 3 血球計數器
Fig. 4 血球計數器上的方格
23
Cell counting foumular: 計數器上 數目× 2稀釋次數× 10 = cell 個數 cc
3. 藥物處理
槲皮素(Sigma, St Louis, USA) 1. 加入 0.3g 的槲皮素至離心管
2. 加入 DMSO 至 10cc,振動至溶解,濃度為 0.1M(stock solution) 3. 分裝各 1ml 至 ependorf(避光小 tube),共 10 瓶,冰在−20℃冰箱
4. Alamar Blue cell viability reagent
Introduction
細胞健康狀況有很多種方法可以監測。細胞膜的完整性,DNA 合成,DNA 含 量,酶的活性,ATP 的存在,和細胞減少的這些狀況,被稱為細胞活力和細胞死 亡的指標。作為一個細胞健康指示劑,alamarBlue 細胞活性試劑能測量各種人類、
動物細胞株、細菌、植物、真菌的增殖(proliferation)能力。Resazurin,為 alamarBlue®
試劑的活性成分,是一種無毒,細胞滲透性化合物,顏色是藍色的,幾乎無螢光。
一旦進入細胞,Resazurin 會被還原成 resorufin,顏色是紅色,高度螢光性的一種 化合物。活的細胞會不斷的將Resazurin 還原成 resorufin,增加螢光性以及顏色的 變化。
alamarBlue®細胞活性試劑用於評估細胞活力。只需添加 alamarBlue®試劑 10%
的sample 量(即,加入 10μl alamarBlue®試劑至 100μL 的 sample),送至37 度 C 培 養箱1-4 小時。較長的時間有較高的敏感性,而且並不會損害細胞健康。所產生的 螢光可用fluorescence spectrophotometer 讀取,或是用 spectrophotometer 讀取 alamarBlue®試劑的吸光度。最後,繪出螢光強度(或吸光值)與濃度的關係圖。
24
Fig. 5 Resazurin 還原成 Resorufin
AlamarBlue®作為一個細胞活性及增值的指示劑,透過 resazurin 變成 resorufin(如 Fig. 5);Resazurin,一個無螢光性的染料透過活細胞的還原反應變 成紅色且高度螢光性的resorufin(如 Fig. 6)。產生的螢光量與活細胞數量呈正 比。
Fig. 6 Resorufin 的吸光度與放射螢光光譜
Protocol
Fig. 7 AlamarBlue protocol
1. add reagent to cells
2. incubate 1-4hrs at 37C
3. read fluorescence(or absorbance)
4. process data
25
5. 流式細胞儀
流式細胞技術(flow cytometry)是指在流體狀態下觀測細胞的一種技術,而流式 細胞儀(flow cytometer)則是指細胞於流體狀態下移動時,能夠觀測及記錄細胞特質 的儀器。flow cytometry 技術主要是為了快速偵測一顆接著一顆流動於液體水柱中 的顆粒或細胞,當細胞被雷射光機發後,會產生0.5~5°之前散射光(forward scatter,
FSC)及15~150°之側散射光(side scatter 或 right-angle scatter 或 SSC)。由於入射光 的波長小於螢光的波長,因此由光的波長及強度變化,即可測出顆粒的大小(與 FSC 成正比),顆粒性(與 SSC 成正比)。當顆粒本身帶有螢光物質(fluorochrome),
或被帶有螢光物質的抗體或被其它螢光物質染上的話,則會產生波長不等的螢光 (如 Fig. 8)。Propidium iodide(PI)是核酸染劑,若單一細胞中核酸含量愈高,則與此 類螢光物質結合的量也愈高,因而受雷射激發後產生的螢光強度也就愈強。利用 此性質,可用來決定核酸量因凋零死亡時核酸量減少的變化或瞭解細胞分裂週期 (cell cycle)的變化。
Fig. 8 細胞經雷射光激發產生的散射光及螢光 SSC
入射光 細胞
FSC
螢光
26
6. Affymetrix® Human Genome U219 Array
Inroduction
Affymetrix® Human Genome U219 Array 擁有 49,411 個探針組,探針長度為 25 mer。是一個易於使用,低成本,且在人類全基因表達分析中最流行,最廣泛使用 的基因晶片。此晶片是專門為GeneAtlas® System 所設計,此系統結合了低成本、
簡單的工作流程、易用性、優異的性能,同時配合工業標準基因表達陣列設計。
這些人類全基因組表達陣列全面覆蓋在一個方便的格式,可同時處理四個樣本,
這種格式也降低了基因晶片處理的複雜性。
特點如下:
1. 最多引用─數以千計的研究使用此晶片的設計。
2. 方便的格式─在最少手動操作陣列的情況下,可同時處裡四種樣本。
3. 高轉錄覆蓋─9 至 10 個探針對一個轉錄因子,能得到高準確的表達。
4. 高重複性─訊號的 correlation 等於或大於 95%。
5. 高敏感性─以 1:100,000 檢測超過 70%的 RNA 轉錄因子。
27
二、 實驗方法
1. Cell proliferation assay
(1) 將人類肝癌細胞株 Hep3B、Huh7 分別種入 96 孔盤(Nunc, Denmark),每孔 50λ 的medium 包含5000個細胞,放入37℃ 、5%CO 的 incubator 至貼壁生長。
(2) 待細胞貼壁後加入50λ 不同濃度槲皮素的 medium(0、20、100、200、400、
1000μM),使之各孔藥物濃度為 0、10、50、100、200、500μM。
(3) 經過 24、48、72hr 後,用顯微鏡(Olympus)觀察並拍照。之後在各孔各加入 10λ 的Alamar Blue,放入incubator 4-6hr 反應至變色。
(4) 利用 Elisa Reader(Dynatech Laboratories, Chantilly, VA, USA)讀取吸光值,測 量波長570nm、參考波長 620nm。
(5) 將各濃度讀取的吸光值對控制組做百分比,並將 data 表示成 mean±SD,在 與控制組做t-test 求出 p value 並作圖,P<0.05 視為有顯著意義。(*表示 P<0.05;
**表示 P<0.001)。
2. Colony formation assay
(1) 將人類肝癌細胞株 Hep3B、Huh7 分別種入 6 孔盤(Nunc, Denmark),每孔3000 個細胞,放入37℃ 、5%CO 的 incubator 至貼壁生長。
(2) 待細胞貼壁後吸掉 medium,加入不同濃度槲皮素的medium(0、1、5、10、
50、100μM)。
(3) 經過10 天培養後,吸乾medium,各孔各加入 1cc Crystal violet(Sigma, USA),
固定細胞並染色,放置2-3hr 後用水沖洗並晾乾。
(4) 計算每個well 被染色的 colony 數目,將data 表示成 mean±SD,在與控制組 做t-test 求出 p value 並作圖,P<0.05 視為有顯著意義。(*表示 P<0.05;**表 示P<0.001)。
28
3. Wound-healing assay
(1) 將人類肝癌細胞株 Hep3B、Huh7 分別種入 6 孔盤(Nunc, Denmark),每孔 1 × 10 個細胞,放入 37℃ 、5%CO 的 incubator 至貼壁生長。
(2) 待細胞長至滿盤後,吸乾 medium,使用無菌的 pipet 10λ tip(gilaon,
Inc.:Middleton, WI)的尖端,在貼滿單層細胞的 well 底盤畫上十字,刮起在附 著在底盤的細胞,進行wound 的動作。加入 PBS 清洗被刮起的細胞及碎片,
並重複兩次清洗。
(3) 小心加入不同濃度槲皮素的 medium(0、10、50、100μM),以免細胞被沖起。
利用顯微鏡觀察細胞移動情形,並在0、12、24、36、48 hr 時拍照。
(4) 將照片中wound 的寬度進行量化,將 data 表示成 mean±SD,在與控制組做 t-test 求出 p value 並作圖,P<0.05 視為有顯著意義。(*表示 P<0.05;**表示 P<0.001)。
4. Flow cytometry assay
(1) 將人類肝癌細胞株 Hep3B、Huh7 分別種入 6 孔盤(Nunc, Denmark),每孔 2 × 10 個細胞,放入 37℃ 、5%CO 的 incubator 至貼壁生長。
(2) 加入不同濃度槲皮素的 medium(0、10、50、100μM)培養。48 小時候用 trypsin 收集細胞,以離心機1500rpm/5 分鐘去除上清液,加入 PBS 清洗細胞,以 4℃
離心機2000rpm/5 分鐘去除 PBS,加入冰冷的 70% ethanol 冰在−20℃冰箱。
(3) 24 小時候以 4℃ 離心機 2000rpm/5 分鐘去除 ethanol,加入 PBS 清洗細胞,
以4℃ 離心機 2000rpm/5 分鐘去除 PBS。
(4) 避光加入 staining solution 1ml,staining solution:1% Triton X-100(Sigma; St.
Louis, MO),0.1 mg/ml RNase A(Sigma; St. Louis, MO), 4g/ml propidium iodide (PI)(Sigma; St. Louis, MO)。
29
(5) 利用 Cell Lab Quanta SC Flow Cytometer (Beckman Coulter)分析不同 DNA 含量的細胞分佈情形。
(6) 將 data 表示成 mean±SD,在與控制組做 t-test 求出 p value 並作圖,P<0.05 視為有顯著意義。
5. Microarray analysis
因從前面四個實驗Proliferation Assay、Colony formation assay、Wound-healing assay 及 Flow cytometry assay 結果得知,同樣濃度的槲皮素對 Huh7 不管在抑制細 胞增殖、抑制細胞菌落發展、抑制細胞移動、抑制細胞週期以及促進細胞凋亡的 效果均比Hep3B 佳;而且槲皮素在濃度 50μM 時,對 Huh7 細胞增殖、菌落發展、
細胞移動、細胞週期和細胞凋亡均已經有顯著的效果,因此此實驗選擇槲皮素濃 度為50μM,細胞株為 Huh7 作微陣列分析(槲皮素濃度 0μM 為控制組,濃度 50 μM 為實驗組),並對分析完後有被調控的基因,選擇兩倍上調與兩倍下調的基因 作Ontology 分析,以 Biological process 與 Molecular function 的分組,從中挑選與 Apoptosis、Cell cycle、Migration 有關的基因做初步探討。
(1) 首先先將人類肝癌細胞株 Huh7 種入 10cm dish,約2 × 10 個細胞,放入 37℃ 、 5%CO 的 incubator 至貼壁生長。
(2) 加入不同濃度槲皮素的 medium(0、50μM)培養。48 小時候用 trypsin 收集細 胞,以離心機1500rpm/5 分鐘去除上清液,加入 PBS 清洗細胞,以 4℃ 離心 機2000rpm/5 分鐘去除 PBS,加入 1mL Trizol 放置 4℃冰箱 overnight。
(以下交給賽亞基因科技公司)
(3) 細胞萃取 RNA 後,以 Agilent 2100 Bioanalyzer electropherogram 測試 RNA 品質。
(4) 加入 poly-A RNA contols,在加入 T7 oligo(dT)引子,合成包含 T7 啟動子序列
30
的第一股cDNA。
(5) 把單股 cDNA 轉換成轉錄用雙股 DNA 模板,合成第二股 cDNA。
(6) 利用雙股 cDNA 模板,產生大量 biotin-modified aRNA 的複製,此為擴增步 驟。
(7) 擴增完後去除雜質,改善 biotin-modified aRNA 的穩定度,純化 aRNA。
(8) 將標記的 aRNA 進行切割,準備作為基因晶片雜交用的 target,進行雜交。
31
第四章 結果
一、 Cell proliferation assay
目前許多中草藥開發成為抗癌藥物之研究,其初步大量篩選皆是利用此方法。
以細胞體外培養的方式,可以簡便的篩選藥物對腫瘤細胞具毒性的物質。首先將
細胞種植在96 孔盤,不含槲皮素的培養液當作控制組,在含有各濃度的槲皮素的
培養液培養24、48、72 小時,讓細胞生長並用顯微鏡觀察。利用 AlamarBlue 試劑 在570nm 量測吸光值,以評估細胞的存活度。從結果可以發現槲皮素能有效抑制 細胞的增殖,並呈現劑量依賴性。我們將讀取的吸光值做統計分析,各個劑量的 的數據均表示成mean±SD,並與控制組作 t-test,P<0.05 視為有顯著意義(*代表 P<0.05;**代表 P<0.001)。
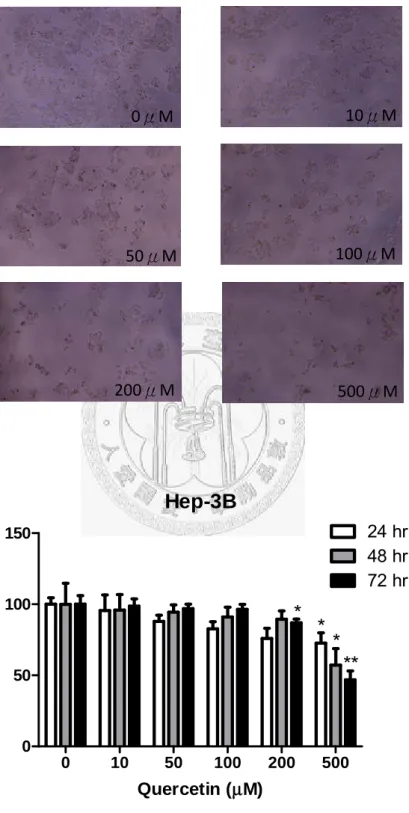
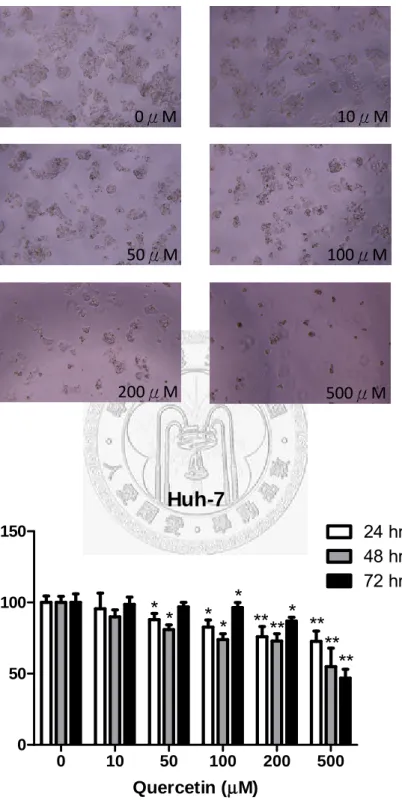
Fig. 9 (A)、Fig. 10 (A)分別代表 Hep3B、Huh7 在槲皮素各濃度(0、10、50、100、
200、500μM)培養 48 小時顯微鏡下(10X)的情形。從照片中可以很明顯看出槲皮濃 度越高,Hep3B 和 Huh7 的細胞增殖抑制效果越明顯。Fig. 9 (B)、Fig. 10 (B)分別 代表槲皮素濃度與Hep3B、Huh7 細胞存活度的關係圖。從圖中可以發現兩株細胞 的存活度都與槲皮素劑量呈現依賴性的關係,濃度越高,細胞存活度越低。Hep3B 在槲皮素濃度200μM、72hr 開始有顯著效果(存活度 86.9%),濃度 500μM 時,在 三個時間點24、48、72hr 皆有明顯效果(存活度 72.7、57.13、46.8%),尤其在 72hr 效果最為顯著。而Huh7 在槲皮素濃度 50μM、24 及 48hr 即開始有顯著效果(存活 度87.9、80.9%),濃度在 100、200、500μM 時,在三個時間點(24、48、72hr)皆有 明顯效果(100μM 三個時間點存活度分別為 82.6、73.9、96.4%;200μM 三個時間 點存活度分別為75.9、72.9、86.9%;500μM 三個時間點存活度分別為 72.6、54.9、
46.8%),尤其在 200μM,48、72hr 以及 500μM,24、48、72hr 效果最為顯著。相 較之下,槲皮素對Huh7 的細胞增殖整體抑制效果較 Hep3B 佳。
32
二、 Colony formation assay
在此方法中,目的是在研究槲皮素對人類肝癌細胞株colony 形成的能力。首 先將細胞種植在六孔盤,不含槲皮素的培養液當作控制組,在含有各槲皮素濃度 的培養液中生長十天,使細胞菌落發展。發現槲皮素能有效抑制細胞的菌落生長,
並呈現劑量依賴性。我們將Colony 的數目做量化統計分析,各個劑量的的數據均 表示成mean±SD,並與控制組作 t-test,P<0.05 視為有顯著意義(*代表 P<0.05;**
代表P<0.001)。
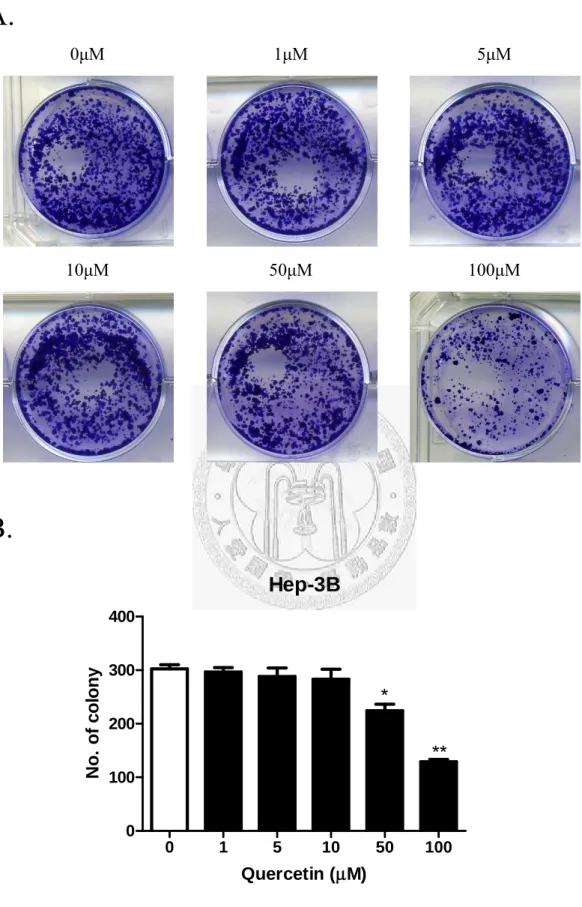
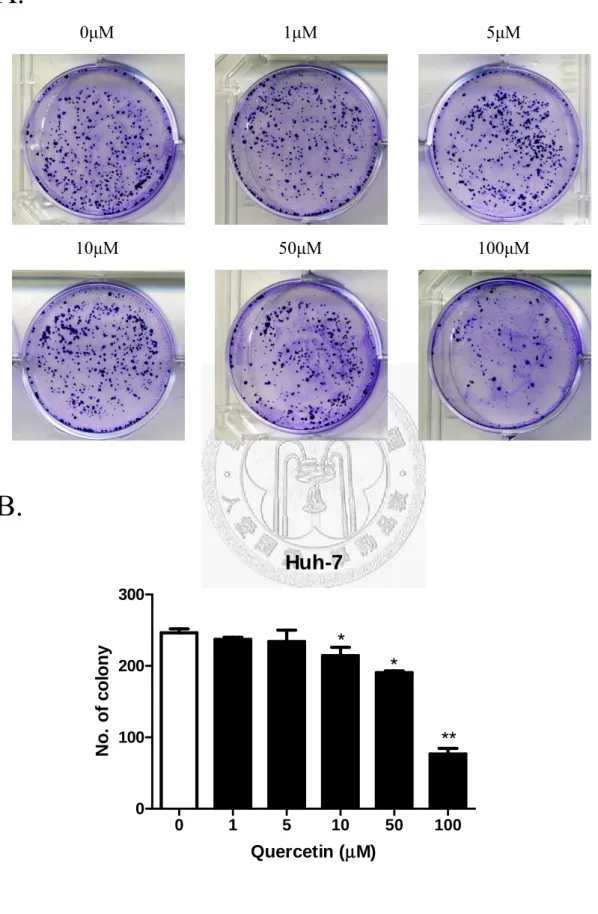
Fig. 11 (A)、Fig. 12 (A)分別代表 Hep3B、Huh7 在槲皮素各濃度(0、1、5、10、
50、100μM),被 Crystal violet 所染色的 Colny 影像圖。從圖中可以看出槲皮素濃 度越高,兩株細胞形成的colony 數目就明顯越少。而 Fig. 11 (B)、Fig. 12 (B)分別 代表Hep3B、Huh7 的 colony 數目與槲皮素濃度的關係圖。從圖中可以發現,藥物 濃度越高,Hep3B 及 Huh7 的 colony 的數目就越少,表示藥物槲皮素的濃度跟形 成colony 的數目呈現一定依賴性的關係。濃度在 0、1、5、10、50、100μM 時 colony 數分別為302、296、288、283、224、129 個。而 Hep3B 在藥物濃度 50μM 時有顯 著抑制細胞colony 的形成效果(占控制組 74.1%),藥物濃度為 100μM 時最為顯著(占 控制組42.7%),對控制組而言減少一半以上的 colony 形成。而 Huh7 效果較 Hep3B 好,濃度在0、1、5、10、50、100μM 時 colony 數分別為 246、237、234、214、
190、76 個。在 10、50μM 即有顯著效果(占控制組 86.9、77.2%),一樣在藥物濃度 為100μM 時最為顯著(占控制組 30.8%),跟控制組相比,只剩下三分之一不到的 colony 形成。
三、 Wound-healing assay
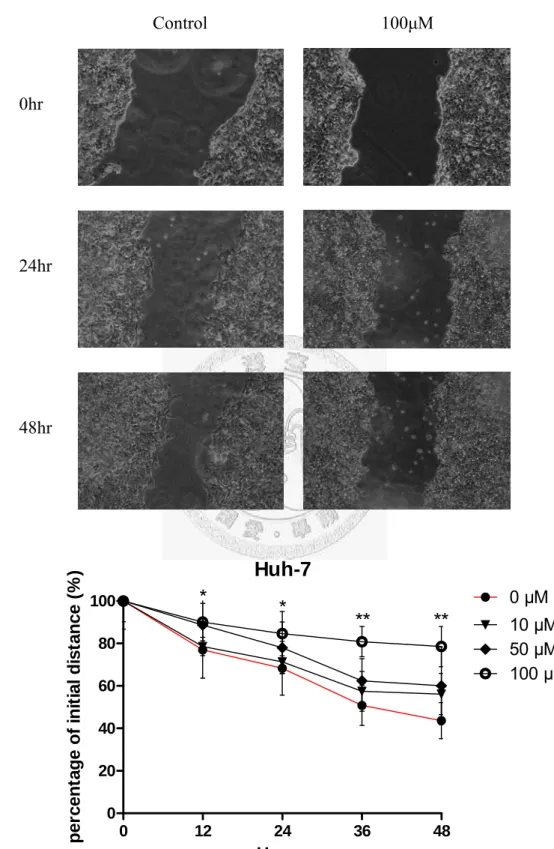
此方法簡單,便宜,是最直接研究體外細胞移動的方式。這種方法模擬體內 傷口癒合中的細胞移動情形。目的是在研究槲皮素對人類肝癌細胞株migration 的
33
能力。首先將細胞種植在六孔盤,等待其長滿後,用tip 的尖端在單層細胞上建立 一個傷口,加入各濃度的槲皮素,並在細胞移動過程中定期觀察照相,並比較圖 像,量化細胞移動速率做統計分析。數據均表示成mean±SD,實驗組(100μM)與 控制組作t-test,P<0.05 視為有顯著意義(*代表 P<0.05;**代表 P<0.001)。
Fig. 13 (A)、 Fig. 14 (A) 分別代表 Hep3B、Huh7 在槲皮素濃度 100μM 與控制 組傷口寬度與時間(0、24、48hr)的影像圖。從圖中可以發現,Hep3B 及 Huh7 在 48 小時候,控制組與實驗組在傷口的寬度上都有些微差距,但 Huh7 較為明顯,
表示槲皮素對兩株細胞均有抑制細胞移動之效果,而且對Huh7 抑制效果較好。Fig.
13 (B)、Fig. 14 (B) 分別代表 Hep3B、Huh7 在槲皮素各濃度(0、10、50、100μM) 與時間(0、12、24、36、48hr)與寬度的關係圖。從關係圖中可以發現,濃度 100μM,
Hep3B 在四個時間點與控制組的距離為 12、27.8、24.78、22.9%;Huh7 為 13.1、
16.3、30、34.9%。槲皮素對兩株細胞同樣在濃度 100μM、12 小時即有顯著抑制細 胞移動效果,36 及 48 小時候效果最為顯著。
四、 Flow cytometry assay
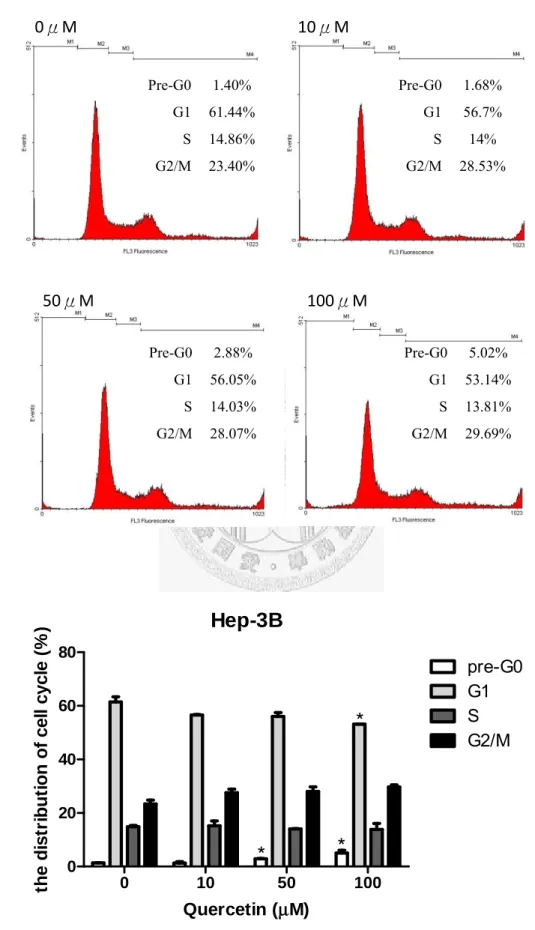
Propidium iodide(PI)是核酸染劑,若單一細胞中核酸含量愈高,則與此類螢光 物質結合的量也愈高,因而受雷射激發後產生的螢光強度也就愈強。此實驗目的 就是利用此性質,用螢光強度來決定核酸量因凋零死亡時核酸量減少的變化,來 判斷細胞是否正處於細胞凋亡狀態,以及了解槲皮素對整個細胞週期調控的影響。
首先將Huh7 種植在六孔盤,加入各濃度槲皮素(0、10、50、100μM),培養 48 小 時候將細胞打下,用Cold Ethanol fix overnight,在加入 PI 染劑,用流式細胞儀分 析。數據均表示成mean±SD,並將實驗組與控制組作 t-test,P<0.05 視為有顯著意 義(*代表 P<0.05;**代表 P<0.001)。
Fig. 15 (A)、Fig. 16 (A)分別為槲皮素對 Hep3B、Huh7 經流氏細胞儀分析後各 phase 的分佈圖。其中 Pre-G0 phase 代表細胞正處於細胞凋亡狀態。槲皮素濃度 0、
34
10、50、100μM 時,Hep3B 在 Pre-G0 phase 分別為 1.4%、1.68%、2.88%、5.02%,
Huh7 為 9.37%、28.06%、60.93%、67.69%。在 G1 phase 方面,槲皮素濃度 0、10、
50、100μM 時,Hep3B 分別為 61.44%、56.7%、56.05%、53.14%;Huh7 為 44.13%、
30.06%、11.77%、11.22%。在 S phase 方面,槲皮素濃度 0、10、50、100μM 時,
Hep3B 分別為 14.86%、14%、14.03%、13.81%;Huh7 為 16.87%、10.87%、6.15%、
5.63%。在 G2/M phase 方面,槲皮素濃度 0、10、50、100μM 時,Hep3B 分別為 23.4%、28.53%、28.07%、29.69%;Huh7 為 29.63%、31.39%、21.63%、15.99%。
整體來看,槲皮素的濃度提高會造成Pre-G0 phase 的上升,G1、S phase 的減少,
表示槲皮素對兩株細胞均有濃度與凋亡率、G1 phase、S phase 的依賴關係。由 Fig.
15(B)、Fig. 16(B) 可得知槲皮素對 Hep3B 在濃度 50μM 開始有促進細胞凋亡顯著 效果,在100μM 有抑制 G1 phase 效果。而 Huh7 在濃度 10μM 時就有細胞凋亡顯 著效果,50、100μM 的效果更好,以及在濃度 10μM 以上可顯著抑制 G1 phase,
在濃度50μM 以上可顯著抑制 S phase。
五、 Microarray analysis
基因晶片,是近年來生命科學研究的一大利器。它可經由一次實驗,得到生物 體所有基因表現量的數據。本實驗目的在於,由基因表現的觀點來探討槲皮素對 人體肝癌細胞的基因變化,瞭解槲皮素對人類肝癌細胞能有效抑制背後的藥理機 轉。首先先將人類肝癌細胞株Huh7 種植在 10cm dish,加入濃度 50μM 的槲皮素 培養48 小時(濃度 0μM 當作控制組),在將細胞打下加入 Trizol 後送至賽亞基因科 技公司作RNA 萃取及微陣列分析。
Fig. 17 為控制組與實驗組萃取出來的 RNA 電泳圖。圖中 18S 與 28S RNA 的 band 清晰明顯。而且在 Fig. 18 圖中控制組與實驗組的 RIN(RNA Integrity Number ) 值高達9.8 及 9.5(10 為 RNA 完整性最好、.0 為最差),表示萃取出來的 RNA 的品
35
質很高,完整性佳。Fig. 19 為Labeled aRNA 的 Quality 檢測,控制組於實驗組 OD 260/280 值分別為 1.95、1.97(需介於 1.8─2.0),濃度分別為 1.5463μg/μl、1.3198μg/μl。
Fig. 20 及 Fig. 21 分別為控制組(Huh7-0)與實驗組(Huh7-50)雜交完後微陣列的影 像。
基因晶片的數據十分龐大,為減低分析的複雜度,兼顧通盤了解槲皮素對於人 類肝癌細胞的機轉,我們選擇兩倍上調與兩倍下調的基因作Ontology 分析,以 Biological process 與 Molecular function 的分組,從中挑選與 Apoptosis、Cell cycle、
Migration 有關的基因做初步探討。Table. 1 為槲皮素上調 2 倍以上 Apoptosis 相關 基因,被調控的基因分別為IGFBP3、CAV1、FOSL1、LGALS1、PMAIP1、ETS1、
C8orf4、SOX4、TNFRSF12A、EGLN3、SOS1、SERPINE1、TNFSF9、TNFRSF21、
SOCS2、PHLDA2、BCL2L11、ANGPTL4、GCLC、BEX2、IER3、SH3RF1、BIRC3、
GCLM、ADM、DUSP6、BIK、MCL1、DUSP6、BMP2、KRT20、DUSP6、MAX、
SQSTM1、ABR、AMIGO2、DDIT3、SOD2、PHLDA1、TSPO、EPHA2、FOXC1、
NR4A1、FGF2、TNFRSF10A、DFFA、F3、TGFA。Table. 2 為槲皮素上下調 2 倍 以上Apoptosis 相關基因,被調控的基因分別為 IGF2、B4GALT1、MAGED1、PRLR、
PCSK9、PLG、TNFSF10、F7、KNG1、LCN2、COL2A1、GSN、FAM3B。Table.
3 為槲皮素上調 2 倍以上 Cell cycle 相關基因,被調控的基因分別為 CYP1A1、
CDKN2B、C13orf15、FOSL1、ETS1、DST、RGS2、NEDD9、CDKN1C、BEX2、
PLK2、BMP2、DDIT3、TRNP1、FOXC1、FGF2、NOLC1。Table. 4 為槲皮素下 調2 倍以上 Cell cycle 相關基因,被調控的基因分別為 CASC5、MCM6、G0S2。
Table. 5 為槲皮素上調 2 倍以上 Cell migration 相關基因,被調控的基因分別為 IGFBP3、TNFRSF12A、SERPINE1、SERPINE2、TUBB2B、JAG1、HBEGF、CTGF、
ITGA2、VCAN、EPHA2、CEACAM1、FOXC1、FGF2、TRIB1、F3。Table. 6 為 槲皮素下調2 倍以上 Cell migration 相關基因,GAB1、AMOT、IGF2、B4GALT1、
SUN2、APOA1、F7、APOA1、VTN。
36
第五章 討論
隨著基因體研究的盛行,以及愈來愈繁雜的基因體資料,生物學家們於是需 要一個工具,可以系統性的整合基因的資料,基因的產物,甚至是基因所具備的 功能。在這種需求下,Gene Ontology[80][81]就誕生了。為了統整各個生物基因體 資料,方便互相溝通比對,瞭解生物體共通的運作法則,它提供了一個統一的字 彙結構來描述基因的三個層面:Molecular Function、Biological Process 和 Cellular Component。Molecular Function 是以分子的角度,描述基因產物的活動,例如催化
(catalytic activity)或轉錄調控(transcription regulator activity)等;Biological process 則是描述在細胞內發生的,可以定義開始和結束的事件或行動;Cellular Component 表示細胞的某個部分或位置,該分支裡面的字彙用來描述基因在細胞的哪個位置 發揮它的功能。應用此資料庫,我們可快速的歸納基因晶片的資料,做初步機轉 探討。對於槲皮素作用於Huh7 的微陣列分析結果,我們首先採取 2 倍上調與 2 倍 下調以上的基因作ontology 分析,以 Biological process 與 Molecular function 的分 組,從中挑選與Apoptosis、Cell cycle、Migration 有關的基因做初步討論。
有關癌症與細胞凋亡的相關研究,顯示癌症的發生是由於癌細胞無法正常的 進行細胞凋亡(apoptosis),造成漫無止境的大量增生,而妨礙到正常的細胞生理機 能,因此細胞凋亡、不受控制的增生與癌症關係成為相當熱門的研究課題。在槲 皮素影響Apoptosis 的兩倍上調基因裡,最突出的基因為 IGFBP3,上調了 8.53 倍,
其基因產物為類胰島素生長因子結合蛋白3(insulin-like growth factor binding protein 3),Lee KW 等人發現,IGFBP3 藉由與 retinoid X receptor-alpha (RXRalpha) 結合後,RXRalpha 的結合伙伴核受體 Nur77 會迅速從細胞核易位到粒腺體啟動凋 亡級聯反應,造成caspase 的激活[82],誘導細胞產生凋亡。另外上調7.52 倍的 CAV1 及2.23 倍的 SOD2,Trimmer C 等人發現這兩個基因在基質微環境(stormal
microenviroment)中有腫瘤抑制功能。缺乏 CAV1 的成纖維細胞會促進乳癌細胞 (MDA-MB-231)的腫瘤生長,而 SOD2 的過度表達可以抑制缺少 CAV1 的成纖維細
37
胞的促進腫瘤影響[83]。Han F 等人發現,CAV1 的過度表現會抑制 EGFR-MAPK pathway,造成抑制細胞的侵襲和促進細胞凋亡[84]。另外槲皮素上調2.05 倍的 BMP2 在骨骼及軟骨發育中扮演一個重要的角色,有研究顯示此基因可抑制胃癌細 胞的增殖以及造成CDK4 的下調,讓癌細胞在細胞週期 G1 phase 受到阻滯[85]。
此外在Apoptosis 兩倍上調基因裡,發現槲皮素也參與了 Apoptosis signal
transduction pathway 的調控,在 Death Receptor Pathway 中上調腫瘤壞死因子配體 TNFSF9,腫瘤壞死因子受體 TNFRSF12A、 TNFRSF21(Death receptor 6)、
TNFRSF10A(Death receptor 4);在 Mitochondrial Pathway 中上調促凋亡基因 PMAIP1(NOXA)、BCL2L11(BIM)、BIK,最終都會導致 caspase 的激活,產生及 聯反應,造成細胞凋亡。而由我們實驗四Flow cytometry assay 結果 Fig. 16 可以證 明槲皮素的確能促進細胞凋亡,Fig. 16 (A)表示槲皮素在濃度 0、10、50、100μM 時,細胞凋亡率分別為9.37%、28.06%、60.93%、67.69%,從 Fig. 16 (B)可知槲皮 素在10μM 即有顯著促進細胞凋亡效果,50、100μM 效果更顯著。
在槲皮素影響Apoptosis 兩倍下調中最突出的基因為 PRLR,下調了 4.63 倍。
PRLR 其基因產物為催乳素受體(Prolactin receptor),屬於 Cytokine receptor 家族成 員,會與催乳素(Prolactin)作用當作跨膜受體。當催乳素與受體結合後,會誘導激 活以下三個途徑:JAK / STAT 途徑、MAPK /ERK 途徑和 PI3K/Akt 途徑[86]。
JAK/STAT 及 MAPK /ERK 途徑訊號途徑是透過細胞膜傳送細胞外的化學訊號到細 胞核內DNA 上的基因啟動子,啟動 DNA 的轉錄,影響細胞功能如生長、分化[87]。
PI3K/AKT 途徑中,透過激活 AKT,通過對其下游的靶蛋白進行磷酸化而發揮其 抗凋亡的作用,如AKT 能通過磷酸化 Bcl-2 家族成員 BAD 阻止凋亡,還有可以磷 酸化mTOR,讓細胞生長、增殖,此外還能透過對 N F-κB 和 P53 的間接作用影響 細胞存活[88]。Granado-Serrano 等人發現,槲皮素能透過調控 Akt、ERK 生存訊號 途徑,讓槲皮素能依劑量性的抑制細胞增殖[40]。因此我們推測槲皮素藉由下調 PRLR,減少 JAK / STAT 途徑、MAPK /ERK 途徑和 PI3K/Akt 途徑的激活,抑制