國立臺灣大學生命科學院植物科學研究所 博士論文
Institute of Plant Biology College of Life Science
National Taiwan University Doctoral Dissertation
阿拉伯芥 A1 及 A2 型熱休克轉錄因子之功能研究 Functional Analysis of Class A1 and A2 Heat Shock
Transcriptional Factors in Arabidopsis 劉祥欽
Hsiang-chin Liu
指導教授:常怡雍 博士 Advisor: Yee-yung Charng, Ph.D.
共同指導教授:施明哲 博士 Co-advisor: Ming-Che Shih, Ph.D
中華民國一百零二年六月 June 2013
國立臺灣大學博士學位論文 口試委員會審定書
中文題目:阿拉伯芥 A1 及 A2 型熱休克轉錄因子之功能研究 英文題目:Functional Analysis of Class A1 and A2 Heat Shock
Transcriptional Factors in Arabidopsis
本論文係劉祥欽君(學號 D96B42010)在國立臺灣大學植物科 學研究所完成之博士學位論文,於民國 102 年 6 月 3 日承下列考試 委員審查通過及口試及格,特此證明
口試委員:中央研究院 農生中心
常怡雍 副研究員 (指導教授)
台灣大學 植物科學所 施明哲 教授 (共同指導教授) 台灣大學 植物科學所
靳宗洛 副教授 台灣大學 生化科技學系
王愛玉 教授 中央研究院 農生中心
邱子珍 研究員 中央研究院 農生中心
葉國楨 副研究員
致謝
謝謝常怡雍老師,您多年來對我孜孜不倦的教導以及親身示範治學該有
的嚴謹態度,讓我獲益匪淺。感謝老師建立起優良的實驗環境讓我的工作能
順利的完成,並對我的論文不厭其煩的詳加修改,使其更臻完善。
感謝中研院農生中心的施明哲老師、邱子珍老師及葉國楨老師、中研院
植微所的鄭萬興老師、台大植科所的靳宗洛老師以及台大生化科技系的王愛
玉老師細心審閱我的論文並惠予指導及建議。
感謝實驗室的成員廖秀婷、馮偉民、邱澗庚、賴韻如、王暐迪、林孟逸、
胡愷倫、阮郁婷、林修穎、林耀斌、吳亭穎、柴幗馨、葉碧珠及李宗遠在實
驗及生活上對我的多方協助。
最後我要感謝我的父母及妻子,你們給予我的愛與鼓勵,是我不斷向前
的寶貴能量。
謹以此論文獻給所有幫助過我的人,謝謝你們。
Table of Contents
Abstract in Chinese. ...X Abstract in English... XII Chapter 1 General introduction
1.1 Heat shock response and heat shock proteins. ...1
1.2 The identification of heat shock factor as the regulator of HSR. ...2
1.3 Functional domains and the post-translational modifications of heat shock factors....3
1.4 HSFs in yeast and animals. ...6
1.5 HSFs in plants. ...7
1.6 Progresses and goals. ... 11
Chapter 2 The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis 2.1 Abstract. ...12
2.2 Introduction. ...13
2.3 Materials and methods. ...17
2.3.1 Plant materials and growth conditions. ...17
2.3.2 Evaluation of growth rate and seed size...17
2.3.3 Thermotolerance assays. ...20
2.3.4 Oxidative, osmotic and salt stress treatments...20
2.3.5 RNA isolation and RT-PCR analysis. ...21
2.3.6 Microarray analysis...22
2.3.7 Immunoblotting ...23
2.3.8 Protein sequence alignment...23
...24
2.4.2 The QK mutant showed growth retardation and altered morphologies ...24
2.4.3 The QK mutant was very sensitive to HS treatments. ...27
2.4.4 Expression of a set of HSR genes was significantly decreased in the QK mutant at elevated temperature. ...32
2.4.5 HSFA1 mediated the induction of HSP genes under salt, osmotic, and oxidative stresses and conferred tolerance. ...38
2.5 Discussion...42
Chapter 3 Acquired thermotolerance independent of Heat Shock Factor A1 (HSFA1), the master regulator of the heat stress response 3.1 Abstract. ...51
3.2 Introduction. ...52
3.3 Materials and methods. ...54
3.4 Results and discussion. ...55
Chapter 4 Common and distinct functions of Arabidopsis class A1 and A2 Heat shock factors in diverse abiotic stress responses and development 4.1 Abstract. ...63
4.2 Introduction. ...64
4.3 Materials and methods. ...68
4.3.1 Plant materials and growth condition. ...68
4.3.2 Generating transgenic plants ...68
4.3.3 Evaluating seed size, seed abortion rate, and growth rate of young seedlings ...68
4.3.4 Thermotolerance assays. ...69
4.3.5 Oxidative, osmotic, and salt stress treatments...69
4.3.6 Immunoblotting and quantitative RT-PCR...70
4.3.7 Microarray analysis...70
4.4 Results. ...71
4.4.1 Ectopic expression of HSFA2 complements the defects of HSFA1 QK mutant in growth and development. ...71
4.4.2 Constitutive expression of HSFA2 induces expression of HSPs in the absence of HSFA1s, but is devoid of auto-regulation...75
4.4.3 Heat stress response genes are differentially regulated by the HSFA1s and HSFA2 ...79
4.4.4 Ectopic HSFA2 expression confers tolerance to diverse heat stress regimes and promoted heat-induced callus formation in the absence of HSFA1s...85
4.4.5 QK and hsf1a/b/d triple mutants are hypersensitive to prolonged exposure to temperatures as low as 27°C. ...89
4.4.6 The HSFA1s and HSFA2 have differential roles in tolerance to salt, osmotic, and oxidative stresses. ...94
4.5 Discussion...96
Future perspectives. ...104
References...106
List of Figures
Figure 1.1 Domain structure and activation of human heat shock factor 1 (HSF1) ...4 Figure 1.2 Phylogenetic relationship of plant HSFs ...8 Figure 2.1 Isolation of triple and quadruple knockout lines of Arabidopsis HSFA1s ...25 Figure 2.2 The QK mutant showed smaller seed size, growth retardation and altered
morphologies ...26 Figure 2.3 HSFA1s did not affect flowering time ...28 Figure 2.4 QK seedlings displayed diverse morphological phenotypes and a reduced HSP90
level...29 Figure 2.5 The QK mutant was very sensitive to various HS treatments...30 Figure 2.6 Thermotolerance phenotypes of wild-type and mutant seedlings...31 Figure 2.7 Adult plants of eTK and QK lines sown on soil were also sensitive to HS treatment
...33 Figure 2.8 Induction of HSR genes were affected in the QK and eTK mutants ...34 Figure 2.9 A major portion of genes up-regulated by HS are regulated by HSFA1s...37 Figure 2.10 HSFA1s mediated the induction of HSP genes under salt, osmotic and oxidative
stresses and conferred tolerance. ...40 Figure 2.11 Alignment of the N-terminal sequences of animal HSF1s and plant HSFA1s ...48 Figure 2.12 Working model of the HSFA1s-mediated regulatory cascade triggered by heat
stress in Arabidopsis ...49 Figure 3.1 Arabidopsis seedlings attained low levels of acquired thermotolerance in the
absence of the HSFA1s ...56 Figure 3.2 The expression levels of bZIP28, BIP1/BIP2, and UTR3 in the wild type and QK
mutant ...60 Figure 3.3 Simplified working model of the transcription network of the HSR in Arabidopsis.
...61 Figure 4.1 Transcript levels of the four HSFA1s and HSFA2 in transgenic plants overexpressing HSFA2... 72 Figure 4.2 Ectopic HSFA2 expression complements the defects of QK mutant in development
of seed and seedling ...73 Figure 4.3 Constitutive expression of HSFA2 induces expression of HSPs in the absence of
HSFA1s ...76 Figure 4.4 Effect of constitutive expression of HSFA2 on the transcript levels of HSP101,
HSA32, and endogenous HSFA2...77 Figure 4.5 Differential regulation of heat-shock induced genes by HSFA1s and HSFA2 ... 81 Figure 4.6 Confirmation of the target preference of HSFA1s and HSFA2 by qRT-PCR ...84 Figure 4.7 Constitutive expression of HSFA2 conferred high level of thermotolerance in the
absence of the HSFA1s under different heat stress regimes ...86 Figure 4.8 Constitutive expression of HSFA2 conferred a high level of thermotolerance in the
absence of HSFA1s ...87 Figure 4.9 Constitutive expression of HSFA2 promoted the formation of calluses after heat
treatment in the absence of the HSFA1s ...88 Figure 4.10 HSFA1a/b/d are required for tolerance to chronic heat stress at temperatures above 27°C in seedlings and flowering plants ... 90 Figure 4.11 Contributions of individual HSFA1s and constitutive expression of HSFA2 in
tolerance to salt, osmotic, and oxidative stresses ...92 Figure 4.12 HSFA2 is not essential for growth at 29°C, or tolerance to oxidative, salt, or
Figure 4.14 Constitutive expression of HSFA2 in wild-type background caused dwarfism with small rosette leaves ...99
List of Tables
Table 2.1 Primers used in this study. ...18 Table 2.2 Classification of HSFA1-dependent HSU genes...43 Table 4.1 Classifications of the heat-induced genes differentially regulated by HSFA1s and
HSFA2...83
摘要
在真核生物中,熱休克反應(heat shock response)主要是由 Heat shock factor(HSF)
所調控。但參與植物熱休克反應的主要調控者仍沒有廣泛地被鑑定出來。這個主要調控 者被認為可能是 HSFA1,因為共抑制(cosuppression)蕃茄中的 HSFA1a 會明顯的抑制 熱休克反應及降低熱耐受性。然而目前為止,仍沒有明確的證據顯示阿拉伯芥
(Arabidopsis)中的四個 HSFA1(HSFA1a/b/d/e)具此類似的功能。不論是單一或雙重 剔除的突變株,其熱休克反應及熱耐受性皆無受到明顯影響。這可能是因為多個類似功 能之基因所造成的互補效應。為了研究各別 HSFA1 的功能,我將 HSFA1a/b 及 HSFA1d/e 的雙重突變株互交,進而得到三重及四重突變株。實驗發現,HSFA1a/b/d 是熱休克反應 的主要調控者,而 HSFA1e 只有在種子耐熱能力的調控有部分功能。然而在植物的發育 上,這四個 HSFA1 皆具有重疊的重要功能。微陣列分析發現 HSFA1 調控了超過 65%的 熱誘導基因,但當植物喪失 HSFA1a/b/d 時,非致死高溫仍可以誘導出相對微弱但顯著的
後生耐熱能力(acquired thermotolerance),這個現象說明了可能有兩套以上的熱感應及熱
反應的機制存在於植物細胞中。
植物的熱誘導系統有一個不同於酵母菌及動物的特點就是他們擁有數個本身也會 受熱誘導的 HSF,而 HSFA2 是其中反應最強的一個。在蕃茄的系統中,HSFA2 跟 HSFA1a 會結合成異質聚合體來啟動熱休克蛋白(heat shock protein, HSP)基因的轉錄。但是到目 前為止,並沒有明確的證據指出 HSFA2 可以在沒有 HSFA1 的情況下獨立運作。因此,
我將 HSFA2 持續表現在 HSFA1a/b/d/e 同時被破壞的突變株中,藉此來闡明 HSFA2 的獨 立功能。我發現 HSFA2 與 HSFA1 個別偏好調控熱休克反應中的某些基因。另外,HSFA2 可以取代 HSFA1 在發育以及抵抗某些環境逆境的功能。有趣的是針對廣義的熱逆境(從 27°C 到 45°C)及氧化逆境,HSFA2 可以賦予植物良好的耐受性,但是對高鹽及高滲透
壓的逆境則較無作用。總結以上,我們的研究闡明阿拉伯芥中的 HSFA1 及 HSFA2 可以 獨立運行其功能,並指出它們功能上的共通性及特異性。
關鍵字 Key words 熱逆境 Heat stress
熱逆境轉錄因子 Heat shock transcriptional factor 非生物逆境 Non-biotic stress
植物耐熱能力 Plant thermotolerance 熱休克蛋白 Heat shock protein
Abstract
Heat shock factors (HSFs) are the well-known master regulators of heat shock response (HSR) in eukaryotes. In tomato, cosuppression of HSFA1a largely reduced HSR and
thermotolerance, which suggests the master role of HSFA1 in plants. However, the seeking of a master regulator of HSR in Arabidopsis is not yet accomplished that the single or double KO mutation of four HSFA1s (HSFA1a/b/d/e) in previous studies did not obviously abolish the HSR and thermotolerance. In this study, triple and quadruple KO mutants of HSFA1s clearly demonstrated their overlapping roles not only in HSR but also in development and the tolerance to other stresses. About 65% heat-upregulated genes were under the control of HSFA1s as revealed by microarray data, which indicated that a small portion of HSR was independent of HFSA1s. Indeed, a minimal but significant level of acquired thermotolerance was still induced without HSFA1a/b/d by a relatively milder heat stress regime. This
phenomenon suggests at least two heat-sensing and -responding mechanisms exist in plant cells.
Plants have several heat-induced HSFs, which is a unique feature only found in plants but not in yeast and animals. Among them, HSFA2 is the most induced one. In tomato, HSFA2 together with HSFA1a form the hetero-oligomeric complexes to activate the transcription of HSP genes. However, no direct evidence showed whether HSFA2 could function
independently without HSFA1s. To this end, HSFA2 was constitutively expressed in the HSFA1a/b/d/e quadruple knockout mutant for evaluation of its independent function. I found that HSFA1s and HSFA2 differentially regulated the HSR genes. Moreover, HSFA2 could replace the functions of HSFA1s in development and tolerance to a wide range of heat stress from 27°C to 45°C, and to H2O2 but not salt and osmotic stresses. Taken together, the findings reveal that HSFA1s and HSFA2 can work independently and have overlapping and distinct functions in Arabidopsis.
Chapter 1
General introduction
1.1 Heat shock response and heat shock proteins
Temperature is one of the limiting factors in determining the growth and survival of organisms. Optimal growth temperature in general determines the distribution of organisms on earth. Although the optimal growth temperature varies from species to species, triggering of heat stress response (HSR) inside cells is universal when ambient temperature exceeds the permissive temperature zone.
HSR is manifested by a rapid and transient induction of protective proteins such as the evolutionally conserved molecular chaperones called heat shock proteins (HSPs). HSPs, classified as HSP100s, HSP90s, HSP70s, HSP60s, HSP40s and small heat shock proteins (sHSPs) according to their molecular weight. In maintaining protein homeostasis, HSPs refold denatured proteins and prevent their aggregation at stress conditions, and assist the folding of de novo synthesized proteins at the optimal condition (Richter et al., 2010; Hartl et al., 2011;
Koga et al., 2011). HSP100s form a hexameric ring structure to disaggregate the denatured protein aggregates after stress with the assist of HSP70/HSP40. The aggregated polypeptides are pulled through the central pore of HSP100 complex into the unfolded state at the expense of ATP hydrolysis, gaining a chance to refold again. HSP90s, HSP70s, HSP40s/J-domain proteins (the cofactors of HSP70s) and HSP60s (chaperonins) are the main chaperones involving in the folding of newly synthesized proteins and refolding of denatured proteins.
They bind to the exposed hydrophobic regions of unfolded proteins to block their aggregation and promote their correct folding in the ATP-dependent manner. The family of sHSPs is
HSP90, HSP70/HSP40, and HSP60, the functions of sHSPs are not dependent on ATP. When the cellular environment is suitable for folding, denatured proteins are released from sHSP complex and refolded in the assistance of the ATP-dependent chaperones.
1.2 The identification of heat shock factor as the regulator of HSR
In eukaryotes, HSR is mainly controlled by the heat shock transcription factor (HSF) through recognizing the heat shock element upstream the TATA box of HSP genes (Wu, 1995).
The studies of HSR were initiated by the accidental discovery of Ferruccio Ritossa in 1962. He observed the heat-shock induced puffing patterns at specific sites of polytene chromosomes in the larvae salivary glands of Drosophila melanogaster (Ritossa, 1962). This phenomenon indicated enhanced transcription of distinct genes during heat stress. It provided the ideal system for studies of heat-shock induced genes and eventually led the isolation of the HSP genes (Schedl et al., 1978; Holmgren et al., 1979; Corces et al., 1980; Craig and McCarthy, 1980; Voellmy et al., 1981) and the cis-element at the promoter regions (Ingolia et al., 1980;
Török and Karch, 1980; Holmgren et al., 1981; Ingolia and Craig, 1981; Karch et al., 1981). A palindromic nGAAn sequence upstream the TATA box of Drosophila HSP70 was found to be necessary for the heat induction and named as heat shock element (Pelham, 1982). The heat shock element (HSE) was generally found in the promoter regions of eukaryotic HSP genes (Bienz and Pelham, 1987). Subsequent studies on the HSE-interacting proteins enabled the identification and purification of HSFs in Drosophila and yeast (Parker and Topol, 1984; Wu, 1984; Sorger and Pelham, 1987; Wiederrecht et al., 1987; Wu et al., 1987). The purified HSFs provided the information for cloning of the single copy HSF genes in yeast and Drosophila (Sorger and Pelham, 1988; Wiederrecht et al., 1988; Clos et al., 1990). The follow-up works in cloning the HSFs of tomato and human revealed the multiplicity of HSF genes in plants and vertebrates (Scharf et al., 1990; Rabindran et al., 1991; Schuetz et al., 1991).
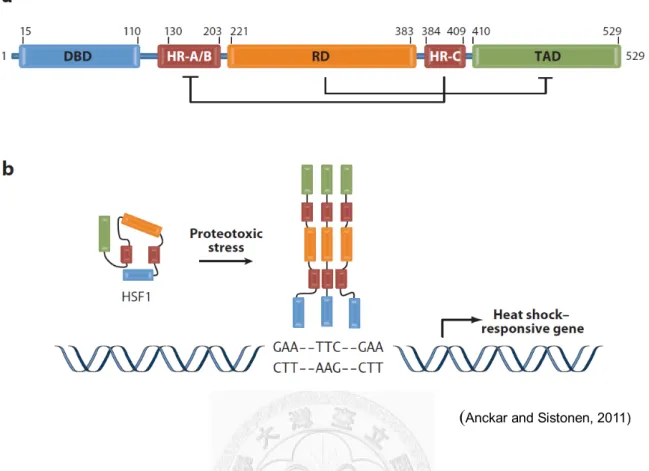
1.3 Functional domains and the post-translational modifications of heat shock factors The cloned HSF genes enabled researchers to analyze their homology and identify the evolutionarily conserved functional domains (Fig. 1.1). Transcription factors in eukaryotes can recognize the specific promoter sequences through the diverse types of DNA binding domain (DBD) and HSFs are no exception. Although the size and sequence of HSFs are variable among or within species, the DBDs near the N-terminus of HSFs are conserved with the globular structure of “winged” helix-turn-helix consisting of a core of three α-helices and a four-stranded β-sheet (Harrison et al., 1994; Vuister et al., 1994; Schultheiss et al., 1996;
Littlefield and Nelson, 1999; Cicero et al., 2001). The third α-helix directly binds to the guanine site of HSE through the formation of hydrogen bond (Littlefield and Nelson, 1999).
Consequently, the guanine and cytosine are the most conservative positions in the HSEs recognized by mammal HSF1 (Xiao and Lis, 1988; Trinklein et al., 2004).
The high-affinity binding of HSF to the HSE in higher eukaryotes upon heat-activation requires not only the recognition by DBD but also the assembling of trimer through the oligomerization domain (OD) (Xiao et al., 1991; Wu, 1995). The OD adjacent to the DBD is composed of two alpha-helical heptad repeats (HR-A and HR-B) (Peteranderl and Nelson, 1992; Peteranderl et al., 1999). Unlike the typical coiled-coil domains as the dimer type, the ODs of three HSF molecules form an unusual triple-stranded pattern. Another heptad repeat called HR-C near the C-terminus was suggested to suppress the trimerization of HSFs via interacting with the HR-A/B domain and maintains the inactive monomer (Rabindran et al., 1993; Wu, 1995). Accordingly, a single amino acid mutation disrupted the HR-C motif of human HSF1 and caused the constitutively trimeric form and DNA binding (Zuo et al., 1994).
The HSF of yeast and human HSF4 lack the HR-C domains and therefore are present as
Figure 1.1 Domain structure and activation of human heat shock factor 1 (HSF1).
(a) Human HSF1 is composed by the DNA-binding domain (DBD), HR-A/B/C regions, regulatory domain (RD) and trans-activation domain (TAD). (b) The activation of HSF1 by protein-damaging stresses. Upon activation, HSF1 trimerizes through the HR-A/B domains, and binds to HSEs comprised by the inverted repeats of the nGAAn pentamer in the promoters of heat shock-responsive genes.
(Anckar and Sistonen, 2011)
The C-terminal regions of HSFs are less conserved in sequence and size compared to the highly conserved DBD and OD across species (Åkerfelt et al., 2010; Fujimoto and Nakai, 2010;
Scharf et al., 2012). In general, these regions confer the regulation of transactivation and the protein localization. HSF1 of human, the best characterized member in HSFs as an example here, contains two separate transactivation domains (TAD1 and TAD2) at the extreme
C-terminal region enriched in hydrophobic and acidic residues. The activation domains interact with the basal transcription factor to activate the transcription of target genes (Green et al., 1995; Newton et al., 1996). Similar activation domains called AHA motifs (Aromatic, large Hydrophobic, and Acidic amino acid residues) are also found in A type HSFs of plants (Nover and Scharf, 1997). The activity of transactivation domains in HSF1 is restricted by the
regulation domain located between HR-A/B and HR-C of OD (Green et al., 1995; Shi et al., 1995; Zuo et al., 1995). This regulation domain contains several amino acid sites identified as the targets of posttranslational modifications (PTMs) (Åkerfelt et al., 2010; Anckar and Sistonen, 2011). Phosphorylation at different serine and threonine residues of the regulation domain has either positive or negative effects on the transactivation mediated by HSF1 (Chu et al., 1996; Knauf et al., 1996; Kline and Morimoto, 1997; Holmberg et al., 2001; Guettouche et al., 2005). In addition to phosphorylation, the sumoylation and acetylation at the regulation domain are involved in the repression of transactivation and DNA-binding, respectively (Hietakangas et al., 2006; Westerheide et al., 2009). PTMs were also identified in HSFs of plants. In Arabidopsis, phosphorylation was shown to regulate the HSE-binding activity of HSFA1 and the heat-induced nuclear localization of HSFA2 (Liu et al., 2008; Evrard et al., 2013). Sumoylation was identified in HSFA1d, HSFA2, and HSFB2b (Cohen-Peer et al., 2010) and negatively regulates the transactivation of HSFA2 (Miller et al., 2010). The negative
active trimer state of HSFs (Abravaya et al., 1992; Shi et al., 1998; Voellmy, 2004; Yamada et al., 2007; Hahn et al., 2011). In addition to the feedback regulation by HSPs, HSF Binding Protein (HSBP) was found to interact with trimeric HSFs to disassociate them into inactive monomeric form (Satyal et al., 1998; Fu et al., 2006; Hsu et al., 2010).
1.4 HSFs in yeast and animals
Generating the knock out (KO) mutant is one of the most direct methods to evaluate the functional significance of a gene in a particular species. The eukaryotic invertebrate organisms such as yeast, Drosophila, and Caenorhabditis elegans contain single copy of HSF in their genomes. Thus, disruption of their HSF gene enables the comprehensive estimation of their contribution. Yeast can not survive when the sole HSF is disrupted, which means HSF1 is required for the general cell growth and viability (Sorger and Pelham, 1988; Wiederrecht et al., 1988; Gallo et al., 1993). In Drosophila, the conditional KO allele showed that HSF is not required for cell growth or viability but for development at specific stages such as oogenesis and early larval development, and for the survival at severe heat stress (Jedlicka et al., 1997).
The chromatin immunoprecipitation and microarray (ChIP-chip) approach demonstrated that the HSF of Drosophila bound both the heat-shock responsive genes and the genes involved in development and reproduction (Gonsalves et al., 2011). In C. elegans, the mutants of HSF are also more sensitive to high-temperature stress due to impaired HSR and have decreased longevity (Hajdu-Cronin et al., 2004).
Mammals have three HSFs named HSF1, HSF2, and HSF4. Among them, HSF1 is dominant for HSR that the HSF1-KO mice could not induce HSPs at heat stress condition and the thermotolerance was abolished in HSF1-KO cells (McMillan et al., 1998; Xiao et al., 1999).
Moreover, the HSF1-KO mice showed multiple developmental defects such as female infertility, placental insufficiency, and growth retardation (Xiao et al., 1999). HSF2 is not the
major regulator of HSR but interplay with HSF1 to alter the expression patterns of major HSP genes (Östling et al., 2007). The HSF2-KO mice also showed abnormal development in brain and testis, and reduced quality of sperm and ovule (Kallio et al., 2002; Wang et al., 2003). The HSF4-KO mice has abnormal lens fiber cells, indicating the role of HSF4 in lens development (Fujimoto et al., 2004). Altogether HSF1, HSF2, and HSF4 contribute in the normal
development of mammal but only HSF1 plays the principal role in regulating HSR.
1.5 HSFs in plants
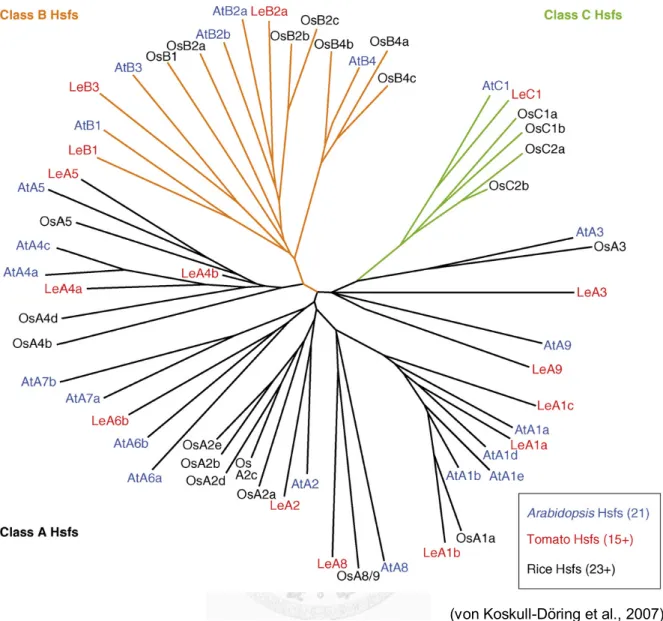
In yeast and animals, the KO strategies efficiently revealed the critical and non-redundant functions of HSFs in development and thermotolerance. However, the achievements of these strategies in plants were limited by the much higher multiplicity compared to yeast and animals that only have one to three HSFs (Fig. 1.2). In general, higher plants have more than twenty HSFs in their genomes (Scharf et al., 2012). For example,
Arabidopsis has 21 HSFs assigned as class A (15 members), B (5 members), and C (1 member) based on the homology of their HR-A/B regions (Nover et al., 2001). Among them, the class A HSFs were suggested as the primary activators of HSR but not class B and class C HSFs because only class A HSFs have the conserved AHA activation domain (Czarnecka-Verner et al., 2000; Kotak et al., 2004). Accordingly, only HSFA1a and HSFA2 but not HSFB1 of tomato could replace the functions of yeast HSF1 (Boscheinen et al., 1997).
Figure 1.2 Phylogenetic relationship of plant HSFs.
The phylogenetic relationship based on the comparison of the conserved DNA-binding domains and HR-A/B regions. For the detail, see the reference (von Koskull-Döring et al., 2007).
(von Koskull-Döring et al., 2007)
T-DNA insertion mutants of Arabidopsis are relatively convenient to obtain for the studies on the biological significance of genes (Alonso et al., 2003; Rosso et al., 2003).
However, the effect of single-gene disruption is often limited due to the existence of other homologous genes with overlapping or redundant functions. In fact, approximately 90% genes in Arabidopsis have at least one homolog (Cannon et al., 2004; Armisén et al., 2008). In the case of class A HSFs, the 15 members were further classified as nine groups (A1-A9) (Nover et al., 2001). The A1-type HSF subfamily has four members (A1a, A1b, A1d, and A1e), while A4, A6 and A7 groups have two members each (A4a/b, A6a/b and A7a/b, respectively). The other groups such as A2, A3, A5, A8, and A9 contain single member. The overlapping functions may be common among class A HSFs and therefore, until 2011, only a few studies focused on the single-HSF-KO mutants such as HSFA2 and HSFA3, showing the thermotolerance defects (Charng et al., 2007; Schramm et al., 2008). HSFA2 and HSFA3 both are heat-induced, which is a unique feature only found in plants HSFs but not in yeast and animals (Nover et al., 2001);
however, the response of HSFA2 to heat is much faster than HSFA3 (Nishizawa et al., 2006;
Schramm et al., 2006; Schramm et al., 2008; Yoshida et al., 2008). The strong heat-inducibility makes HSFA2 the dominant HSF after heat-shock treatment, and its importance was revealed by the specific thermotolerance assay with a extended recovery phase between the 37°C pretreatment and 44°C challenge (Charng et al., 2007). 37°C is a mild, non-lethal heat stress for Arabidopsis that can induce strong HSR including HSFA2. The highly expressed HSFA2 enables plants to sustain the expression level of HSPs for a longer duration after switching to non-stress condition. Hence, the heat-shock induced HSPs decline faster in the HSFA2 KO mutant and therefore the mutant has lower ability to tolerate the later challenge of lethal temperature at 44°C. The heat-shock induction of HSPs as well as the acquired
is its delayed heat-shock response depending on the function of DREB2A, an heat-induced AP2-domain transcription factors important for the drought and salt stress signalling (Sakuma et al., 2006; Schramm et al., 2008). The heat-inducibility of these important HSR regulators such as HSFA2 and the DREB2A-HSFA3 cascade indicates the existence of some upstream transcriptional factors. Four constitutively expressed HSFA1 members (HSFA1a/b/d/e) are the potential candidates of major regulators because co-suppression of HSFA1a in tomato strongly reduced the expression of HSPs and the level of thermotolerance (Mishra et al., 2002).
However, it is not clear whether the effect of co-suppression was limited in HSFA1a or expended to the other HSFA1 members such as HSFA1b and HSFA1c or further to other constitutively expressed HSFs. The double KO mutants of HSFA1a/b or HSFA1d/e in Arabidopsis showed only partial decrease in thermotolerance, which may be due to the functional redundancy of the remained HSFA1 members (Lohmann et al., 2004;
Nishizawa-Yokoi et al., 2011).
It is well-known that HSFs in yeasts and animals have very important roles in multiple developmental processes. In contrast, the function of HSFs in the development of plants is still largely unknown. HSFA9 of Arabidopsis and sunflower is specifically expressed during
embryogenesis and seed maturation (Almoguera et al., 2002; Díaz-Martín et al., 2005; Kotak et al., 2007). Ectopically expressing a dominant negative form of sunflower HSFA9 in tobacco caused significant reduction of seed longevity; however, only minor effects were found in seed maturation and germination (Tejedor-Cano et al., 2010). Notably, several reports showed that overexpressing HSFs retarded the growth of plants that may be due to the decrease of cell proliferation (Ogawa et al., 2007; Yokotani et al., 2008; Yoshida et al., 2008; Zhu et al., 2009).
These observations suggested a correlation between HSFs and plant developments; however, genetic evidence was lacking.
1.6 Progresses and goals
In this thesis, I have elucidated the individual roles of four HSFA1 member and HSFA2.
The multiplicity and redundancy of plant HSFs indeed complicate their functional genetic studies. The phenotypic effects of single KO mutation of HSFs are weak due to the functional redundancy. For the case of HSFA1s in Arabidopsis, the thermotolerance phenotypes were not obvious whether in single or double KO mutants. It may be due to the functional redundancy or weak contribution in thermotolerance of the HSFA1 genes. Therefore, generating the triple and even quadruple mutants of HSFA1s is required to address their overall and individual functions.
In Chapters 2 and 3, I describe the generation of the triple and quadruple mutants of HSFA1s, and partition of HSR as HSFA1s-dependent and HSFA1s-independent parts. We also show the novel function of HSFA1s in the development of seeds and seedlings.
For the individual roles of HSFA2, the study on single KO mutant already demonstrated its unique function in extending the acquired thermotolerance. However, it is not clearly addressed whether HSFA2 could trigger HSR without HSFA1s. In chapter 4, I describe the effects of constitutively expressed HSFA2 in the quadruple mutant of HSFA1s, and
demonstrate the novel role of HSFs in the chronic heat stress at temperature as low as 27°C.
Chapter 2
The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis.
The contents of this chapter are published in Plant, Cell & Environment (Liu et al., 2011).
2.1 Abstract
In Arabidopsis, there are four homologs of class A1 heat shock factor (HSFA1) genes, which likely encode the master regulators of heat shock response (HSR). However, previous studies with double knockout (KO) mutants were unable to confirm this point probably due to functional redundancy. Here, we generated a quadruple KO (QK) and four triple KO mutants to dissect their functions. Our data show that members of the HSFA1 group not only play a
pivotal role in HSR but also are involved in growth and development. Alterations in morphology and retardation in growth were observed in the quadruple but not in triple KO mutants. The basal and acquired thermotolerance capacity was dramatically decreased in the QK mutant but varied in triple KO mutants at different developmental stages. The
transcriptomics profiles suggested that more than 65% of the heat stress (HS)-up-regulated genes were HSFA1 dependent. HSFA1s were also involved in the expression of several HS genes induced by H2O2, salt and mannitol, which is consistent with the increased sensitive phenotype of the QK mutant to the stress factors. In conclusion, the Arabidopsis HSFA1s function as the master regulators of HSR and participate as important components in other abiotic stress responses as well.
2.2 Introduction
In eukaryotes, the conserved heat shock transcription factors (HSFs) are the major regulators of heat stress (HS) responsive genes encoding molecular chaperones and other stress proteins (Wu, 1995). Higher plant genomes contain an expanded HSF family as compared to other eukaryotes (Nover et al., 2001; Shiu et al., 2005). For example, Arabidopsis and rice have at least 21 and 25 HSF homologues, respectively (Nover et al., 2001; Wang et al., 2009).
However, to date, only one and four HSFs can be found in the genomes of invertebrates and vertebrates, respectively (Åkerfelt et al., 2010). The large HSF family in higher plants may be associated with fitness of the sessile organisms that have to constantly face recurring or multiple environmental challenges including extreme temperature, light intensity, flooding, drought, and soil salinity. Understanding the function and complexity of HSF network in higher plants is important in the contexts of the global climate change, which may significantly increase the frequency of duration of extreme heat in some regions (Hayhoe et al., 2004).
Based on structural characteristics and phylogenetic analysis, Arabidopsis HSFs are allocated into three major classes (class A, B, and C) and 14 groups (A1-9, B1-4, and C1;
(Nover et al., 2001). The rice HSF family has representatives in most of the groups (von Koskull-Döring et al., 2007; Wang et al., 2009). Moreover, the number of HSFs in each group varies in different species. For example, rice possesses only one member in the group A1, while Arabidopsis has four (A1a, b, d, e) and tomato has at least three members (A1a, b, c). By contrast, Arabidopsis has only one member in the group A2, whereas rice contains five
members (A2a-e) (von Koskull-Döring et al., 2007). In general, class A HSFs but not class B or class C HSFs contain a transactivator motif (Nover et al., 2001; Kotak et al., 2004).
Due to the availability of T-DNA knockout (KO) lines, considerable progress has been
knockdown of HSFA1a expression dramatically reduce thermotolerance in tomato (Mishra et al., 2002). In Arabidopsis, HSFA1a and HSFA1b were shown to work redundantly on a subset of HS response genes in the early phase of HS response (Lohmann et al., 2004; Busch et al., 2005). However, the HSFA1a/1b double KO mutant does not show substantial defects in thermotolerance (Lohmann et al., 2004). Thus, a unanimous agreement on whether HSFA1 act as the master regulator of HSR in plants has not yet been reached.
Among the HSF family, HSFA2 is the most heat-inducible one (Busch et al., 2005). Its expression is suppressed in the HSFA1a knockdown tomato plants (Mishra et al., 2002).
However, the expression of Arabidopsis HSFA2 is not affected in the HSFA1a/1b double KO line (Busch et al., 2005). Recent studies pointed out the pivotal role of HSFA2 in the late phase of HSR (Schramm et al., 2006; Charng et al., 2007; Wunderlich et al., 2007), suggesting that HSFA2 is a secondary regulator under the control of at least one master regulator to form a transcriptional cascade or network that covers the early and late expression of HS genes.
HSFA2 was also shown to be involved in other environmental stresses (Nishizawa et al., 2006;
Banti et al., 2010). Recently, components that modulate the activities of HSFA1 and A2 in Arabidopsis were reported. Calmodulin-binding protein kinase 3 (AtCBK3) was shown to phosphorylate HSFA1a in vitro and enhance the binding activity of HSF to the heat shock element (Liu et al., 2008). The HSF binding protein (HSBP) was shown to interact with HSFA1a, HSFA1b and HSFA2 and negatively regulate HSR (Hsu et al., 2010). ROF1 and ROF2, which are homologs of peptidyl prolyl cis/trans isomerase, were shown to play positive and negative roles, respectively, in regulating the activity of HSFA2 (Meiri and Breiman, 2009;
Meiri et al., 2010).
Arabidopsis HSFA3 is a HS-induced gene, which is under the control of DREB2A, a HS-inducible gene itself that functions in both HS and water-deficit stress response (Sakuma et al., 2006; Schramm et al., 2008). A moderate defect in thermotolerance was observed in the
DREB2A KO as well as the HSFA3 KO mutants (Sakuma et al., 2006; Schramm et al., 2008;
Yoshida et al., 2008). So far, the upstream regulator of DREB2A and HSFA3 under HS condition remains to be identified.
Biological functions of the members of the A4 group have been demonstrated in rice (Shim et al., 2009). The spl7 mutant of OsHSFA4d leads to spotted leaf phenotype under elevated temperature and solar radiation (Yamanouchi et al., 2002). Recent studies show that OsHSFA4a is involved in cadmium tolerance (Shim et al., 2009). In Arabidopsis,
overexpressing a dominant-negative HSFA4a suppresses the expression of the cytosolic H2O2-scavenging ASCORBATE PEROXIDASE 1 (APX1) that is induced under moderate light stress (Davletova et al., 2005). It was postulated that HSFA4a acts as an H2O2 sensor (Miller and Mittler, 2006).
Arabidopsis HSFA9 is regulated by the seed-specific transcription factor ABI3 and is expressed exclusively in the late stages of seed development (Kotak et al., 2007). It has been suggested that HSFA9 plays a specialized role in regulating HSP genes during seed maturation (Kotak et al., 2007). In agree with this idea, overexpression of sunflower HSFA9 fused a transcription repressor motif in transgenic tobacco caused a reduction in the levels of seed HSPs and seed basal thermotolerance (BT) (Tejedor-Cano et al., 2010).
Compared to the class A HSFs, the biological functions of class B and class C HSFs are less understood. Tomato HSFB1 was shown to be a general transcription co-activator that interacts with HAC1, a plant ortholog of CREB-binding protein (Bharti et al., 2004). However, there are evidences showing that members of the class B HSFs act as transcriptional repressors in Arabidopsis (Czarnecka-Verner et al., 2000; Ikeda and Ohme-Takagi, 2009; Kumar et al., 2009) and soybeans (Czarnecka-verner et al., 2004). Recently, HSFB1 and HSFB2b were
Identifying the master HSF(s) in Arabidopsis is essential in deciphering the complex crosstalks between different stress components known in this important model plant. In this study, we generated a quadruple KO (QK) and four triple KO lines, of which the latter contain only one intact HSFA1 gene. We found that HSFA1a, A1b and A1d work redundantly as the master regulators of HSR. A large portion of the HSR genes is directly or indirectly regulated by these HSFs. They were also required for the induction of several heat-induced transcription factors such as HSFA2 and DREB2A. Moreover, the induction of HSP genes by salt, osmotic, and oxidative stresses were dependent on the function of these HSFs. Interestingly, we also observed a role of these HSFs in early Arabidopsis development. To our knowledge, this is the first report on the role of HSFs on plant growth and development under well-controlled
environmental conditions.
2.3 Materials and methods
2.3.1 Plant materials and growth conditions
The HSFA1a/1b double KO and wild-type (wt) seeds in the ecotype Wassilewskija (Ws) background were kindly provided by Dr Fritz Schöffl (University of Tübingen, Germany) (Lohmann et al., 2004; Busch et al., 2005). The HSFA1d/1e double KO line was generated by crossing the T-DNA insertion line SALK_022404 (hsfA1d) and SALK_094943 (hsfA1e) obtained from the Arabidopsis Biological Resource Center (Ohio State University). The
Arabidopsis Genome Initiative (AGI) numbers of HSFA1a, A1b, A1d and A1e are AT4G17750, AT5G16820, AT1G32330 and AT3G02990, respectively. The QK and triple KO mutants were obtained by crossing the HSFA1a/1b and HSFA1d/1e double KO mutants. The T-DNA
insertions were confirmed by PCR amplification, and then homozygous lines of the mutant allele were isolated. SALK_066374 for HSP101 (AT1G74310) was obtained as previously described (Charng et al., 2006). The sequences of gene-specific primers used in this study are listed in Table 1. For propagating seeds, Arabidopsis plants in soil were grown in walk-in growth chambers at 22°C and 16 h of light (120 mol m-2 s-1). For seedlings grown in plates, Arabidopsis seeds were sterilized and sown on solid medium (25 mL of 0.5x Murashige and Skoog (MS) medium containing 0.8% agar and1%sucrose) in a Petri dish (90 x 15 mm),
incubated for 3 d at 4°C for synchronized germination, then grown to desired stage at 22°C and 16 h of light (120 mol m-2 s-1).
2.3.2 Evaluation of growth rate and seed size
The growth rate of the wt and mutants from the seed to the four rosette leaves stage was measured on agar plates containing 0.5x MS medium and 1% sucrose. Growth stages of
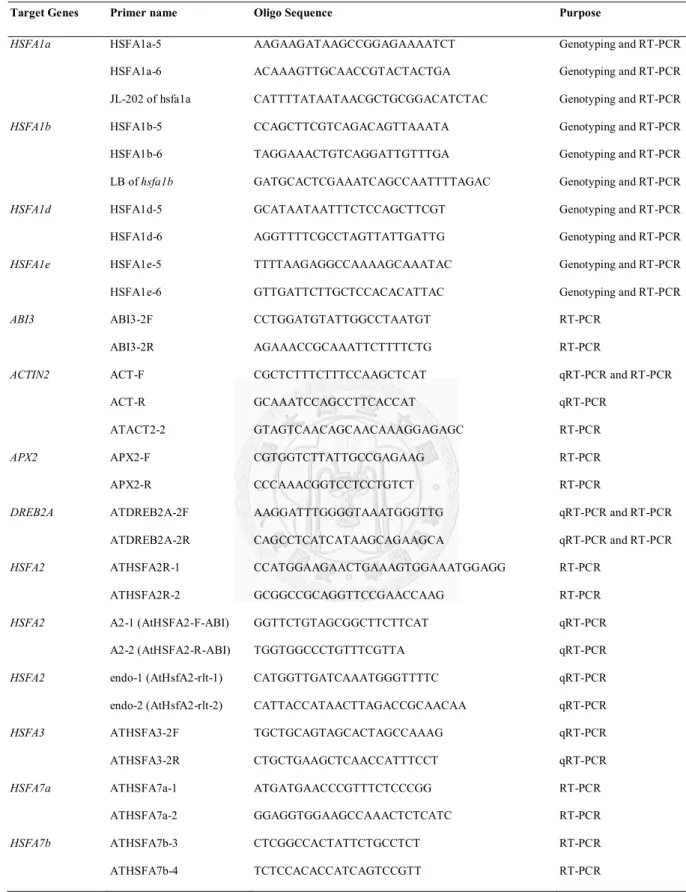
Table 2.1 Primers used in this study.
Target Genes Primer name Oligo Sequence Purpose
HSFA1a HSFA1a-5 AAGAAGATAAGCCGGAGAAAATCT Genotyping and RT-PCR
HSFA1a-6 ACAAAGTTGCAACCGTACTACTGA Genotyping and RT-PCR JL-202 of hsfa1a CATTTTATAATAACGCTGCGGACATCTAC Genotyping and RT-PCR
HSFA1b HSFA1b-5 CCAGCTTCGTCAGACAGTTAAATA Genotyping and RT-PCR
HSFA1b-6 TAGGAAACTGTCAGGATTGTTTGA Genotyping and RT-PCR LB of hsfa1b GATGCACTCGAAATCAGCCAATTTTAGAC Genotyping and RT-PCR
HSFA1d HSFA1d-5 GCATAATAATTTCTCCAGCTTCGT Genotyping and RT-PCR
HSFA1d-6 AGGTTTTCGCCTAGTTATTGATTG Genotyping and RT-PCR
HSFA1e HSFA1e-5 TTTTAAGAGGCCAAAAGCAAATAC Genotyping and RT-PCR
HSFA1e-6 GTTGATTCTTGCTCCACACATTAC Genotyping and RT-PCR
ABI3 ABI3-2F CCTGGATGTATTGGCCTAATGT RT-PCR
ABI3-2R AGAAACCGCAAATTCTTTTCTG RT-PCR
ACTIN2 ACT-F CGCTCTTTCTTTCCAAGCTCAT qRT-PCR and RT-PCR
ACT-R GCAAATCCAGCCTTCACCAT qRT-PCR
ATACT2-2 GTAGTCAACAGCAACAAAGGAGAGC RT-PCR
APX2 APX2-F CGTGGTCTTATTGCCGAGAAG RT-PCR
APX2-R CCCAAACGGTCCTCCTGTCT RT-PCR
DREB2A ATDREB2A-2F AAGGATTTGGGGTAAATGGGTTG qRT-PCR and RT-PCR
ATDREB2A-2R CAGCCTCATCATAAGCAGAAGCA qRT-PCR and RT-PCR
HSFA2 ATHSFA2R-1 CCATGGAAGAACTGAAAGTGGAAATGGAGG RT-PCR
ATHSFA2R-2 GCGGCCGCAGGTTCCGAACCAAG RT-PCR
HSFA2 A2-1 (AtHSFA2-F-ABI) GGTTCTGTAGCGGCTTCTTCAT qRT-PCR
A2-2 (AtHSFA2-R-ABI) TGGTGGCCCTGTTTCGTTA qRT-PCR
HSFA2 endo-1 (AtHsfA2-rlt-1) CATGGTTGATCAAATGGGTTTTC qRT-PCR
endo-2 (AtHsfA2-rlt-2) CATTACCATAACTTAGACCGCAACAA qRT-PCR
HSFA3 ATHSFA3-2F TGCTGCAGTAGCACTAGCCAAAG qRT-PCR
ATHSFA3-2R CTGCTGAAGCTCAACCATTTCCT qRT-PCR
HSFA7a ATHSFA7a-1 ATGATGAACCCGTTTCTCCCGG RT-PCR
ATHSFA7a-2 GGAGGTGGAAGCCAAACTCTCATC RT-PCR
HSFA7b ATHSFA7b-3 CTCGGCCACTATTCTGCCTCT RT-PCR
ATHSFA7b-4 TCTCCACACCATCAGTCCGTT RT-PCR
Target Genes Primer name Oligo Sequence Purpose
HSFA9
ATHSFA9-1F ATHSFA9-1R
TCGTCAGCTTAACTCTTACGGTTT TGTTTTTGCTTCTCCTCTTGATG
RT-PCR RT-PCR
HSFB1 ATHSFB1-1 ATGACGGCTGTGACGGCG RT-PCR
ATHSFB1-2 GCAGACTTTGCTGCTTTTCCA RT-PCR
HSFB2a ATHSFB2a-1 ATGAATTCGCCGCCGGTTGACG RT-PCR
ATHSFB2a-2 CAACCACGGCGTCTCCTCATCGG RT-PCR
HSFB2b ATHSFB2b-1 ATGCCGGGGGAACAAACCGG RT-PCR
ATHSFB2b-2 CCGAGTTCAAGCCACGACCC RT-PCR
HSP17.6-CI ATHSP17.6-1 ATGTCTCTAATTCCAAGCATCTTCG RT-PCR
ATHSP17.6-2 TTAACCAGAGATATCAATGGACTTAAC RT-PCR
HSP17.7-CII HSP17.7-CII-F GCAAGAAAGTTAACACAA RT-PCR
HSP17.7-CII-R CACGATCACAAACAAACTC RT-PCR
HSP25.3-p HSP21-F-EJ TGGACGTCTCTCCTTTCGGAT RT-PCR
HSP21-R-EJ TGATCGAGTCCTACTGAATCTGGA RT-PCR
mtHSC70-2 At mtHsc70-2-1F CTCTATCGCTCGGTATTGAAAC qRT-PCR
At mtHsc70-2-1R CTCTTCTTTGTGGGGATGGTTG qRT-PCR
HSP90.1 ATHSP90.1-1F AATACGCTGTTGGACAATTGAA RT-PCR
ATHSP90.1-1R GATTCTCGAAGGACTTCTTCTTC RT-PCR
HSP90 HSP90-1 TTTGGTGTTGGTTTCTACTCTGCTTA qRT-PCR and RT-PCR
HSP90-2 TCGTTCTGACCTTCCTTCATCCTTGT qRT-PCR and RT-PCR
HSA32 HSA32-F-ABI GGAAGAGTTTCGAGGAGAACGA qRT-PCR
HSA32-R-ABI GACCTCGCATCTCCGTAACAC qRT-PCR
HSA32 HSA32-F-EJ AAAGACTATGTGGAGGAGTG RT-PCR
HSA32-R-EJ CACATAGAGATTCACATTTG RT-PCR
HSP101 HSP101-F-ABI TGCATTTAGCTGGTGCTTTGAT qRT-PCR
HSP101-R-ABI CCACCGGCACTAGAGATTGC qRT-PCR
HSP101 Hsp101-F-EJ CACCAGGGTATGTTGGTCACG RT-PCR
Hsp101-R EJ GCACCATACACCGGGTCATAA RT-PCR
TIL1 TIL1-F (lipocalin-F) TTTATCGAAGGCAGCGCCTATA qRT-PCR
TIL1-R (lipocalin-R) GCTTGGCTTCGTCGCTTTTA qRT-PCR
MIPS2 AT2G22240-1F GTAGCTAGTAATGGCATCCTCTTTGA qRT-PCR
were equal to or greater than 2 mm long when the primary florescence reached 1 cm in length, as described previously (Wu et al., 2008). Six individual plants for each line grown in soil pots were measured. The seed size was measured by using 50 and 60 mesh sieves (Sigma-Aldrich, St Louis, MO, USA). The exclusion sizes of the 50 and 60 mesh sieves are 297 and 250 mm, respectively. Seed were divided into three sizes: >297 mm, 297~250 mm and <250 mm.
2.3.3 Thermotolerance assays
For thermotolerance assays, seeds were sown on 0.5x MS plates with 1% sucrose and imbibed at 4°C for 3 d in the dark. After imbibition, the plates were first incubated for 2 h at 22°C and then placed for 2 h in a water bath (44°C). The germination rate was measured after recovery under normal growth conditions for 2-14 d. For the BT assay, 7-day-old seedlings were treated for 26 min at 43°C. For the short-term acquired thermotolerance (SAT) assay, 7-day-old seedlings were acclimated for 1 h at 37°C, recovered for 2 h at 22°C and then treated for 140 min at 44°C. For the long-term acquired thermotolerance (LAT) assay, 5-day-old seedlings were acclimated for 1 h at 37°C, recovered for 2 d at 22°C and then treated for 50 min at 44°C. The above heat treatments were performed in the dark to avoid photo-oxidative stress, and the details were described in (Charng et al., 2006). For the tolerance against moderately high temperature (TMHT) assay, 7-day-old seedlings were treated for 7 d at 35°C under a light/dark cycle of 16 h/8 h (120 mol m-2 s-1). After applying the different HS regimes, plants were recovered for 7 d at 22°C. At the end of recovery, pictures were taken and the survival rates were measured. For thermotolerance assays of adult plants grown in potted soil, 15-day-old plants were placed in a growth chamber and subjected to TMHT conditions. After treatment for 5 d at 35°C, the plants were moved to room temperature and pictures were taken.
2.3.4 Oxidative, osmotic and salt stress treatments
For measurement of the transcript levels of HSR genes after oxidative, osmotic or salt stress treatments, 7-day-old seedlings sown on 0.5x MS plates with 1% sucrose under normal conditions were transferred into solutions containing 150 mM NaCl, 300 mM mannitol or 5 mM H2O2 for 2 h treatment. Subsequently, samples were frozen in liquid nitrogen for RNA extraction. For germination rate assays, seeds were sown in 0.5x MS plates containing 1%
sucrose with 300 mM mannitol or 150 mM NaCl, or 0.1% sucrose with 10 mM H2O2, imbibed at 4°C in the dark for 3 d, and then grown at 22°C for 2, 4 and 7 d. For survival rate assays, seeds were sown on 0.5x MS medium containing 1% sucrose with 300 mM mannitol, 100 mM NaCl or 0.1% sucrose with 5 mM H2O2, imbibed at 4°C for 3 d, and then grown at 22°C for 15 d.
2.3.5 RNA isolation and RT-PCR analysis
Total RNA of plant tissues was isolated as previously described (Charng et al., 2006).
The RNA quantity of samples was determined on a NanoDrop ND-1000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). Semi-quantitative RT-PCR analysis of tested genes was performed as previously described (Charng et al., 2007). PCR reactions were performed for 27 cycles in a volume of 10 μL using cDNA obtained from 12 ng total RNA. The
sequences of the primers for RT-PCR analysis for each gene are described in Supplemental Table 1. Quantitative RT-PCR (qRT-PCR) was performed in a 7300 Real-Time PCR System using the SYBR Green RT-PCR reagent kit following the manufacturer’s protocol (Applied Biosystems, Carlsbad, CA, USA). Each reaction was run in triplicates in a volume of 20 mL with an initial denaturation step at 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. Data were analyzed according to the manufacturer’s instructions using the 7300
ACTIN2 from the CT value of the GOI. The amplification efficiency (E), calculated based on a previous method (Peirson et al., 2003), was larger than 0.95 in each reaction.
2.3.6 Microarray analysis
Seven-day-old seedlings of Col-0, Ws and QK mutant grown at 22°C on 0.5x MS plates containing 1% sucrose were incubated for 1 h at 37°C. Subsequently, samples were collected for RNA extraction. The experiment was repeated and two biological replicates were processed for analysis. RNA extraction, quantification and microarray analysis using the ATH1
GeneChip arrays containing 22 810 probe sets (Affymetrix, Santa Clara, CA, USA) were performed as previously described (Chi et al., 2009). Assessment of experimental quality and statistical analyses were performed using remote analysis computation for gene expression (RACE) (Psarros et al., 2005), where individual probe results files (CEL files) were used as input and normalized using the robust multi-array average (RMA) algorithm (Irizarry et al., 2003). Four thousand two hundred eighty-one features with an absent detection call and 64 control features were removed for all chips analyzed. In total, data for 18,465 probe sets were used as input data for statistical analysis. Genes whose expression changed more than threefold between mutant and wt seedlings at a P value of <0.05 were selected. False discovery rate (FDR) at the stringent level of 0.05 was used to declare statistical significance and account for multiple tests. The relative change of these 18,465 features between wt and QK mutant with the same selection criteria was compared. Annotations and gene ontology classifications of
Arabidopsis genes were obtained from The Arabidopsis Information Resource database (TAIR, http://www.Arabidopsis.org). The microarray data reported here can be accessed in the Gene Expression Omnibus at the National Center for Biotechnology Information (NCBI; accession number GSE26266).
2.3.7 Immunoblotting
Protein extraction and immunoblotting were performed as previously described (Charng et al., 2006). Rabbit antiserum against AtHSFA2 was produced by employing a synthetic peptide with the sequence ‘HLLKNIKRRRNMGLQNVN’ as antigen. Immunization and serum collection were performed by commercial service (LTK Biotechnology, Taoyuan, Taiwan). The specificity of the AtHSFA2 antibody was confirmed by probing against protein samples extracted from heat-treated HSFA2 KO plants. The antibodies against HSP101, HSP90, HSA32, sHSP-CI and tubulin used herein were described previously (Charng et al., 2006; Chi et al., 2009).
2.3.8 Protein sequence alignment
Protein sequences of different species were obtained from NCBI
(http://www.ncbi.nlm.nih.gov). The alignment of the amino acid sequences of HSFs from different species was done by using the constraint-based multiple alignment tool (COBALT, http://www.ncbi.nlm.nih.gov/tools/cobalt) in NCBI.
2.4 Results
2.4.1 Isolation of triple and quadruple knockout lines of Arabidopsis HSFA1 genes To test the hypothesis that HSFA1 genes are functionally redundant in Arabidopsis, we produced a QK mutant of the Arabidopsis four HSFA1 genes by crossing the hsfA1a/1b (Ws ecotype background) and hsfA1d/1e (Col-0 ecotype background) mutants. In addition, to elucidate the function of each HSFA1 subtype, we isolated four triple KO mutants, named as aTK, bTK, dTK and eTK, where the prefixed letters represent the remaining functional HSFA1.
The genotypes and HSFA1 transcripts of these mutants were confirmed by PCR and RT-PCR, respectively (Fig. 2.1). Since the Col-0 and Ws genomic backgrounds were employed for generating the mutants, we included the wt of both ecotypes as controls.
2.4.2 The QK mutant showed growth retardation and altered morphologies
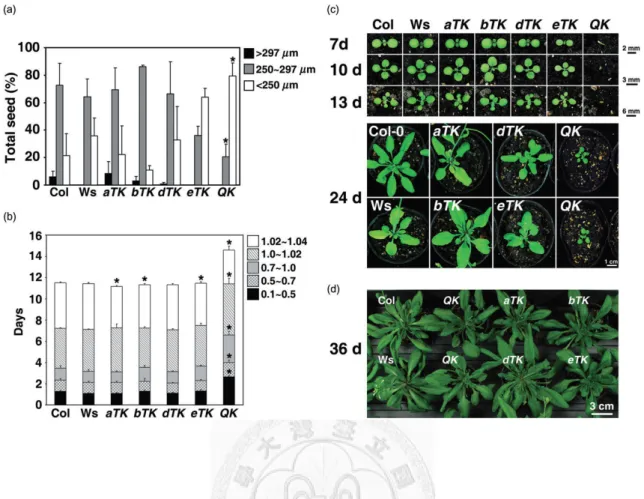
The Arabidopsis HSFA1 genes are not essential for viability and reproduction because the QK mutant could generate viable seeds at normal condition for several generations during our studies. However, in the absence of functional HSFA1 genes, plant growth and development were substantially affected. First, we noticed that the size of the mature seeds were
significantly decreased in the QK mutant but not in the triple KO mutants (Fig. 2.2a). We then compared the growth rates on solid medium plates based on the developmental stage defined by Boyes et al. (2001). The time needed for reaching stage 0.5 (emergence of radicle) and stage 1.0 (cotyledon fully opened) from stage 0.7 (hypocotyl and cotyledon emergence) was about 1.5 d longer in the QK mutant than in the other lines (Fig. 2.2b). After stage 1.0, the difference between the QK mutant and the wt plants became smaller. The size of the QK seedlings was smaller than that of the wt and triple KO plants at the same growth stage on solid medium plate (data not shown). This difference was even more obvious when the plants were grown in soil under long-day photoperiods (Fig. 2.2c). However, under short-day periods, the
Figure 2.1 Isolation of triple and quadruple knockout lines of Arabidopsis HSFA1s.
(a) Structures and the T-DNA insertion sites of the HSFA1 genes. Exons and introns are indicated as black and grey blocks, respectively. T-DNA insertion sites are marked by triangles with corresponding ecotypes. The arrows indicate the locations of primers used for PCR analysis of genotypes. (b) The genotypes of QK and triple KO mutant (aTK, bTK, dTK and eTK) lines were confirmed by multiplex PCR using the primers shown in (a). ACTIN2 was included as a positive control. (c) RT-PCR analysis of the transcript levels of HSFA1 genes using the primers shown in (a). The RT-PCR product of ACTIN2 is shown as a loading control.
Figure 2.2 The QK mutant showed smaller seed size, growth retardation and altered morphologies.
(a) Distribution of seed sizes of the wild types (Col-0 and Ws) and mutants. Results are presented as mean values of three replicates ± SD (n ≥ 150 each). (b) The growth rate of the wild type and mutants from imbibed seed to four rosette leaves stage grown on 0.5× MS medium plates containing 1% sucrose.
Growth stages are defined as follows: 0.1, seed imbibition; 0.5, radicle emergence; 0.7, hypocotyl and cotyledon emergence; 1.0, cotyledons fully opened; 1.02, two rosette leaves >1 mm; 1.04, four rosette leaves >1 mm. Results are presented as mean values of six replicates ± SD (n = 20 each). *P < 0.01 (versus wild-type plants, Student’s t-test). (c) 7- to 24-day-old plants grown in soil under long-day conditions (16 h light). (d) 36-day-old plants grown in soil under short-day conditions (12 h light).
rosette of QK mutant plants reached a size similar to that of the wt and triple KO plants (Fig.
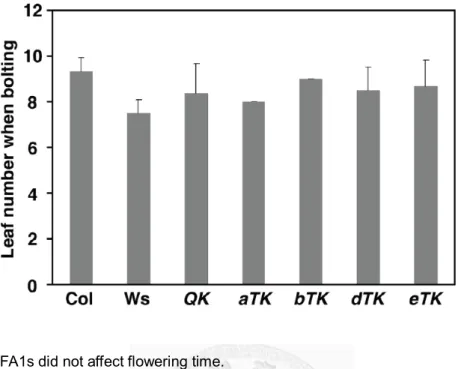
2.2d). There were no significant differences between the wt and the QK and triple KO mutants on flowering time (Fig. 2.3) and in the morphology of flowers and siliques (data not shown).
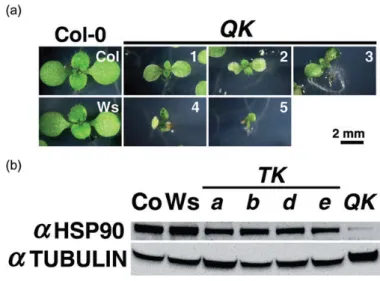
Another interesting finding is that the QK mutant showed diverse morphological phenotypes at the seedling stage. More than half of the QK seedlings showed pale greenish cotyledons with flimsy and wrinkled edges (Fig. 2.4a). The remaining mutant seedlings showed an array of different morphologies such as extruded root, meristemless shoot or concaved cotyledons, which were similar to that of the HSP90-RNAi lines (Sangster et al.
2007). Since these morphologies have been shown to associate with a lower HSP90 expression, we measured HSP90 levels by Western blotting. We found that the level of HSP90 was much lower in the QK mutant than in the wt and in the triple KO mutants (Fig. 2.4b).
2.4.3 The QK mutant was very sensitive to HS treatments
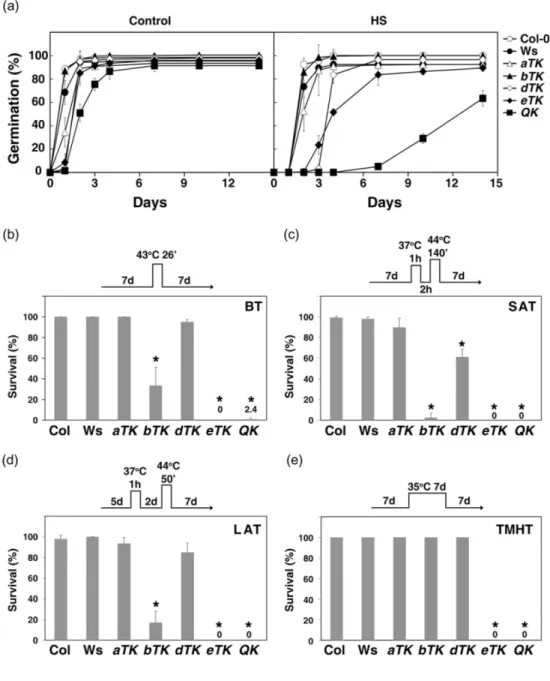
To investigate whether the HSFA1s constitute the master regulators of thermotolerance in Arabidopsis, we compared the tolerance levels of the QK and four triple KO mutants, as well as wt (Ws and Col-0) plants at different developmental stages under different HS regimes.
Firstly, we tested the thermotolerance of the seeds. Following heat treatment of imbibed seeds for 2 hr at 44°C, the germination rates of the QK mutant seeds were severely reduced, while rates of dTK and eTK seeds were moderately decreased (Fig. 2.5a). However, the
thermotolerance of aTK and bTK seeds was not significantly different from that of the wt.
Secondly, we tested the thermotolerance of the seedlings under four different HS regimes to reveal the BT, SAT, LAT and TMHT. The LAT condition has previously been used to reveal the phenotypes of several HS genes (Charng et al., 2006; 2007; Meiri and Breiman, 2009;
Figure 2.3 HSFA1s did not affect flowering time.
The flowering times of the wild-type (Col-0 and Ws) and mutant plants are shown as the number of rosette leaves at bolting. Results are presented as mean values ± SD (n = 4).
Figure 2.4 QK seedlings displayed diverse morphological phenotypes and a reduced HSP90 level.
(a) Five morphological types of the QK seedlings grown under normal conditions: (1) slightly pale greenish cotyledons (21.6%); (2) pale greenish cotyledons with flimsy and wrinkled edge (54.1%); (3) extruded root from medium (5.4%); (4) concaved cotyledons (3.6%); and (5) shoot meristemless;
delayed development (15.3%). Pictures were taken on day 9 after imbibition. The scale bar applies to all the pictures. (b) Western blot analysis of the cytosolic HSP90 level in 7-day-old seedlings grown under normal conditions. In each lane, approximately 100 mg of total protein was loaded. Tubulin serves as a loading control. Co, Col-0; TK, triple knockout; a, b, d, and e indicate aTK, bTK, dTK and eTK,
respectively.
Figure 2.5 The QK mutant was very sensitive to various HS treatments.
(a) The seed germination rate of the wild type (Col-0 and Ws) and mutants after HS treatment. The seeds were imbibed at 4°C for 3 d on 0.5× MS plates containing 1% sucrose followed by treatment at 44°C (HS) or 22°C (Control) for 2 h. The samples were returned to normal growth conditions and the germination rates were determined at indicated days after treatment. (b-e) The survival rates of seedlings after the treatments at different HS regimes schematically shown on top of each panel. (b) Basal thermotolerance (BT). (c) Short-term acquired thermotolerance (SAT). (d) Long-term acquired thermotolerance (LAT). (e) Thermotolerance to moderately high temperature (TMHT). For each treatment, three biological repeats were performed. Results are presented as mean values of three
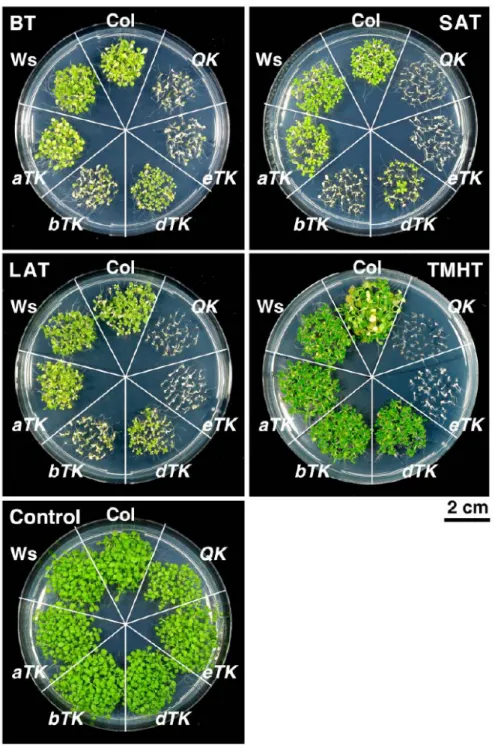
Figure 2.6 Thermotolerance phenotypes of wild-type and mutant seedlings.
BT, basal thermotolerance; SAT, short-term acquired thermotolerance; LAT, long-term acquired thermotolerance; and TMHT, tolerance against moderately high temperature. The different HS treatments are described in 2.3.3.
were generally non-viable (Fig. 2.5b-e and Fig. 2.6). The bTK mutant performed better than the eTK and QK but significantly worse than the aTK and dTK mutants, both of which exhibited no or small differences in comparison with the wt under the BT, LAT and TMHT regimes (Fig.
2.5b-e). Under SAT conditions, the aTK mutant performed slightly better than the dTK mutant (Fig. 2.5c). We also tested the thermotolerance of 20-day-old soil-grown plants; all QK and eTK plants were dead after exposure to 35°C for 5 d, whereas the other three triple KO mutants did not differ much from the wt (Fig. 2.7). The dramatic loss of thermotolerance of the QK mutant was unlikely due to its slow development. Because the development of the eTK mutant was similar to that of the wt plant, it was as heat-sensitive as the QK mutant. As a comparison, we noticed that the eTK and QK mutants had a much lower acquired thermotolerance level than that of HSP101 KO mutant (data not shown). These results suggest that HSFA1a, A1b and A1d play a major but partially redundant role in thermotolerance, whereas HSFA1e confers
thermotolerance to some extent at the seed stage but not at seedlings and adult plant stages.
2.4.4 Expression of a set of HSR genes was significantly decreased in the QK mutant at elevated temperature
To explain the dramatic loss of thermotolerance of the QK mutant, we first examined the expression of several HSR genes by RT-PCR in the wt and mutant lines. In imbibed seeds, the transcript levels of HSP101, HSA32 and HSP17.7-CII were significantly reduced as compared to aTK and bTK mutants or the wt (Fig. 2.8a). The transcript levels of these HSP genes in dTK and eTK were higher than in the QK mutant but lower than in the aTK and bTK mutants. On the other hand, expression of the seed-specific ABI3 and HSFA9 was not substantially affected in the QK mutant. These results suggest that HSFA1a and HSFA1b regulate the expression of the tested HSP genes in the seed independently of AIB3 and HSFA9.
Figure 2.7 Adult plants of eTK and QK lines sown on soil were also sensitive to HS treatment.
Fifteen-d-old plants of wt and mutants were treated in a growth chamber for 5 d at 35°C with a 16-h light/8-h dark cycle and a light intensity of 120 μmol m-2 s-1. After treatment, the photographs of plants were immediately taken.
Figure 2.8 Induction of HSR genes were affected in the QK and eTK mutants.
(a) The transcript levels of ABI3, HSFA9 and HSR genes in imbibed seeds (imbibed for 4 h). (b) The transcript levels of HSR genes in the 7-d-ayold seedlings under control and HS (37°C for 1 h) conditions.
The transcripts of each gene were detected by RT-PCR. The RT-PCR product of ACTIN2 is shown as a loading control. (c) qRT-PCR analysis of relative transcript levels of HSR genes after HS treatment.
Histograms show transcript levels relative to ACTIN2, normalized to the wild type with the highest expression level, and shown in percentage. Results are presented as mean values of three replicates ± SD. *P < 0.01 (versus wild-type plants, Student’s t-test). ND, not detected. (d)Western blot analysis of HSPs in the wild-type and QK mutant plants. Eighteen-day-old plants grown on medium plates were treated at 22°C (Control) or 37°C (HS) for 1 h and recovered at 22°C for 2 h before protein was extracted.
In each lane, approximately 100 mg of total protein was loaded. Tubulin serves as a loading control. Co, Col-0; a, b, d and e indicate aTK, bTK, dTK and eTK, respectively.