國立台灣大學生命科學院生命科學系 碩士論文
Department of Life Science College of Life Science National Taiwan University
Master Thesis
Transcriptional Factors Responsive to CYCLOIDEA in zygomorphic flower of Sinningia speciosa
大岩桐兩側對稱花中受 CYCLOIDEA 調控之轉錄因子
王佩琦
Jocelin Muliawan
指導教授:王俊能 博士 Advisor:Chun-Neng Wang, Ph.D
中華民國 108 年 06 月
June 2019
口試委員會審定書
中文摘要
兩側對稱性花被認定是被子植物演化的主要趨勢,其花從正面可畫出單一個 對稱軸,將花分成兩個鏡像半部,背側,兩側和腹側花瓣沿著此對稱軸排列。兩側 對稱花使傳粉者從固定的角度進入花中,以促進精確的花粉傳播和柱頭接收,從而 大大提高繁殖成功率。在金魚草中,TCP 轉錄因子 CYCLOIDEA(CYC)在侷限在
背部花瓣上表現,CYC 透過調節細胞增殖和細胞延長的作用,促使背側花瓣發育,
使其在外型上與兩側及腹側花瓣相異。然而,CYC 啟動了那些下游基因,以及它 們如何合作以產生背部辨識的花瓣形狀和大小是未知的。野生型大岩桐(Sinningia speciosa)為兩側對稱花朵,然而在人為栽培的大岩桐中,兩側對稱卻可輕易地轉換 成輻射對稱,這說明了花對稱的發育模組可能是很容易改變。
為了找出CYC 可能的下游基因,我們從大岩桐 'Espirito Santo'(SsES)的轉錄
組(RNA-seq)中篩選出背腹側瓣之間的差異性表達的轉錄因子(DE-TFs)。其中,
篩出9 個背側高表達的轉錄因子(包括 SsCYC),其 5 端調節區(regulatory region)都
有鑑定出TCP 結合位點,同時也透過 qRT-PCR 再次驗證這 9 個轉錄因子確實侷限
在背側花瓣表現,因此,這9 個轉錄因子很有可能就是 SsCYC 的下游基因。為了
證明SsCYC 對這九個轉錄因子的調節能力,在煙草(Nicotiana benthamiana)原生質
體的暫時性表達系統中,以雙熒光素酶測定檢測 SsCYC 和報告子(候選 TF 的 5
端調節區)之間的相互作用。結果發現,SsCYC 能夠自我調節,並且活化 RADIALIS-
like(SsRL2)基因,該基因是金魚草中 RADIALIS 的直系同源基因,但其功能尚不
清楚。有趣的是,SsCYC 還活化乙烯反應轉錄激活因子 SsERF1 並抑制乙烯反應轉
錄抑制因子SsERF3 和 ovate 家族轉錄抑制因子 SsOFP6,其功能目前也尚未知。
SsERF1 和 SsERF3 的可以調控乙烯信號傳導途徑的下游基因。它們可能透過
調 控 EXPANXIN(EXPA)基因、木葡聚醣內轉葡糖基酶 /水解酶(xyloglucan
endotransglucosylase/hydrolase)基因和內切-1,4-β-D-葡聚醣酶(EGase)基因來使細
胞壁變的鬆散,進而改變背側花瓣細胞的延長。同時,這三個基因也在大岩桐轉錄 組中被鑑定為背側表達基因,這也符合我們在大岩桐中觀察到背側花瓣的細胞有 較大的細胞面積,因此背側花瓣相較於腹側花瓣長度較長,這也被認為是大岩桐花 發育成兩側對稱的原因之一。
關鍵詞:大岩桐; 兩側對稱性; SsCYC; 5 端調節區; TCP 結合位點;下游轉錄因子;
細胞延長
Abstract
Floral zygomorphy (bilateral symmetry), in which the dorsal, lateral and ventral petals are arranged along a single plane, dividing flower into two mirror-image halves, has been selected as the major trend in angiosperm evolution. Zygomorphic flowers allow the pollinators to enter the flower in fixed angle to facilitate exact pollen deposition and stigma reception, thus greatly enhance reproductive success. In Antirrhinum, TCP transcription factor, CYCLOIDEA (CYC) is strictly expressed at the dorsal petals and it can function to regulate cell proliferation and expansion for generating dorsal identity.
However, what the downstream of CYC are and how they cooperate to generate the petal shape and size for the dorsal identity are largely unknown. The wild type Sinningia speciosa exhibits zygomorphic symmetry, yet reversal to actinomorphic (radial symmetry)
is common, indicating that the developmental module for floral zygomorphy might be easily altered.
In order to discover CYC downstream, differentially expressed transcription factors (DE-TFs) between dorsi-ventral petals were screened from the RNA-seq data of S.
speciosa ‘Espirito Santo’ (SsES). Among them, nine TFs, including SsCYC itself, have
their 5’ regulatory regions been identified with TCP binding sites and their dorsal
restricted expression was confirmed by qRT-PCR. To demonstrate the possible regulation
of SsCYC on these TFs, dual-luciferase assay transiently expressed in protoplasts of Nicotiana benthamiana leaves was used to examine the interaction between the effector
(SsCYC) and the reporter (5’ regulatory region of the candidate TFs). It was found that
SsCYC was able to auto-regulate itself and also upregulate a RADIALIS-like (SsRL2) gene which is the orthologue of RADIALIS in Antirrhinum, but its function is unknown.
Interestingly, SsCYC also up-regulated the ethylene response transcriptional activator, SsERF1 and down-regulated the ethylene response transcriptional repressor, SsERF3 and
an ovate family transcriptional repressor, SsOFP6 whose function is unknown.
The finding of SsERF1 and SsERF3 as SsCYC responsive TFs could be linked to their function as downstream regulators of ethylene signaling pathway. They might alter dorsal cell expansion via regulation of EXPANXIN (EXPA) genes, xyloglucan endotransglucosylase/hydrolase (XTH) encoding gene and endo-1,4-β-D-glucanase (EGase) encoding gene to loosen the cell wall, since these three genes were identified as the dorsal expressed genes in the RNA-seq data of SsES. This suggestion is also reflected by the observation that the dorsal petals of SsES have larger cell area, thus are longer in length compared to the ventral petals, which is considered as one of the factors that generates floral zygomophy in this flower.
KEYWORDS: Sinningia speciosa; floral zygomorphy; SsCYC; 5’ regulatory region;
Table of Contents
口試委員會審定書 ... I 中文摘要 ... II Abstract ... IV Table of Contents ... VI List of Tables ... VII List of Figures ... VIII List of Supplementary Data ... IX Abbreviation ... XI
Introduction ... 1
Materials and Methods ... 9
Results ... 30
Discussion ... 42
Conclusion ... 59
References ... 61
Supplementary Data ... 67
List of Tables
Table 1 The list of 34 TFs identified from 630 DEGs and their BLASTX annotations ... 31 Table 2 The list of 19 TFs predicted to contain TCP binding sites at their 5’ regulatory
region ... 34 Table 3 Percentage of identity between the 5’ regulatory region of S. speciosa
‘Espirito Santo’ (SsES) and S. speciosa ‘Avenida Niemeyer’ (SsAN) ... 38 Table 4 Summary of TCP binding sites found at the 5' regulatory regions isolated from
S. speciosa ‘Espirito Santo’ (SsES) ... 38
Table 5 Summary of TCP binding sites predicted from the 5' regulatory region of S.
speciosa 'Avenida Niemeyer' (SsAN) ... 39
List of Figures
Figure 1 qRT-PCR confirmation of dorsal-expressed TFs that have been predicted to have TCP binding sites at their 5’ regulatory region... 35 Figure 2 Dual-luciferase assay result... 41 Figure 3 SsCYC and SsRAD-like genes expression profile in S. speciosa ‘Espirito
santo’ and S. speciosa ‘Avanti’ ... 46 Figure 4 Possible bias that might occur in the dual-luciferase assay ... 58 Figure 5 Hypothesis of S. speciosa floral zygomorphy regulation by SsCYC... 60
List of Supplementary Data
Supplementary Table S1 Summarized TCP binding consensus ... 67 Supplementary Table S2 Primer list for the isolation of S. speciosa ‘Espirito Santo’
transcription factor coding sequence ... 68 Supplementary Table S3 Primer list for qRT-PCR confirmation of dorsal-expressed
transcription factor of S. speciosa ‘Espirito Santo’ ... 69 Supplementary Table S4 Primer list for the isolation of S. speciosa ‘Espirito Santo’5’
regulatory region ... 70 Supplementary Table S5 Thermal Asymmetric Interlaced PCR (TAIL-PCR) ... 71 Supplementary Table S6 Self-Formed Adaptor PCR (SEFA-PCR) primer list for the
isolation of S. speciosa ‘Espirito Santo’ 5' regulatory region ... 72 Supplementary Table S7 Primer list for the construction of dual-luciferase vector .73 Supplementary Table S8 RNA-seq and BLASTx annotation result of dorsal expressed
genes encoding cell wall loosening agent ... 74 Supplementary Figure S1 The role of CYC in floral zygomorphy ... 75 Supplementary Figure S2 Phylogeny of TCP proteins from Sinningia speciosa,
Solanum lycopersicum and Arabidopsis thaliana ... 76
Supplementary Figure S3 Thermal Asymmetric Interlaced PCR (TAIL-PCR) ... 77 Supplementary Figure S4 Self-Formed Adaptor PCR (SEFA-PCR) ... 78 Supplementary Figure S5 Reporter and internal control construct for dual-luciferase
assay ... 79
Supplementary Figure S6 Construct of SsCYC tag GFP (by Yu-An, Shi) ... 80 Supplementary Figure S7 Construct of 35s-GFP (by Yu-An, Shi) ... 81 Supplementary Figure S8 Sequence alignment for the 5’regulatory region of S.
speciosa ‘Espirito Santo’ and “Avenida Niemeyer’ ... 82
Supplementary Figure S9 Expression profile of SsDIV in S. speciosa 'Espirito Santo' (SsES) ... 102 Supplementary Figure S10 Dorsal and ventral petals observation of S. speciosa
'Espirito Santo' (SsES) ... 103 Supplementary Table S9 Full coding sequences of TFs containing TCP binding sites at
their 5’ regulatory regions ... 104 Supplementary Table S10 5’ Regulatory region sequence of dorsal-expressed TFs of S.
speciosa ‘Espirito Santo’ ... 111
Abbreviation
AD primer Arbitrary degenerate primer bHLH basic helix-loop-helix
C:I Choloroform : Isoamyl alcohol CIB Cryptochrome 2-interacting bHLH CTAB Hexadecyl trimethyl-ammonium bromide
CYC CYCLOIDEA
DEGs Differentially expressed genes DE-TF Differentially expressed TF
DICH DICHOTOMA
DIV DIVARICATA
DRIFs DIV-and-RAD-interacting-factors EGase Endo-1,4-β-D-glucanase
ERF Ethylene response factor
EXPA EXPANSIN
GFP Green Fluorescence Protein GS Primer Gene specific primer
IPTG Isopropyl β-D-1-thiogalactopyranoside LB Luria Bertani
NaOAc Acetic acid sodium salt
NGAL NGATHA-Like
OFP OVATE FAMILY PROTEINS-like P:C:I Phenol : Choloroform : Isoamyl alcohol PCR Polymerase chain reaction
PEG Polyethylene glycol PVPP Polyvinylpolypyrrolidone
RAD RADIALIS
RL RADIALIS-like 2
SEFA-PCR Self-Formed Adaptor PCR SP primer Specific primer
SsA S. speciosa ‘Avanti’
SsAN S. speciosa ‘Avenida Niemeyer’
SsES S. speciosa ‘Espirito Santo’
SsPF S. speciosa ‘Pink Flower’
TAIL-PCR Thermal Asymmetric Interlaced PCR
TCP TEOSINTE BRANCHED1, CYCLOIDEA and PROLIFERATING CELL FACTORS
TF Transcriptional Factor TR Transcriptional regulator
X-gal 5-bromo-4-chloro-3-indolyl-beta-D-galacto-pyranoside XTH Xyloglucan endotransglucosylase/hydrolase
Introduction
Floral symmetry has been considered as the important feature that influences the interaction between plant and pollinator. Generally, there are two main types of floral symmetry, which are zygomorphic (bilateral/mono-symmetry) and actinomorphic (radial/poly-symmetry) that usually could be determined by face-on view of flower perianth. Zygomorphic flowers are characterized by having the dorsal, lateral and ventral petals arranged along a single plane, dividing flower into two mirror-image halves (one dividing plane) whereas the actinomorphic flowers have their perianth arranged into more than one dividing planes. The emergence of zygomorphic symmetry from its actinomorphic ancestral has been correlated with plant-pollinator specific interaction (Spencer and Kim, 2018; Hileman 2014).
The complexity of the floral image in zygomorphic flowers improves the pollinators recognition and discrimination, by limiting the pollination to particular species, preventing the inefficient pollinating species. This restriction then results in reproductive barriers that lead to speciation in both plants and pollinators, often suggested as plant- pollinator co-evolution. While the pollinators of actinomorphic flowers may approach the flowers from any direction, zygomorphic flowers provide these visitors additional
stigma contact with, the pollinator’s body. This precision thus results in a higher proportion of pollen reaching the stigma. Therefore, floral zygomorphy provides more efficient pollination, which is then suggested as a reproductive advantage during the angiosperm evolution. Although the zygomorphic flowers have evolved many times from the actinomorphic ancestors, the reversals to actinomorphic have also been observed. This suggests that the developmental module for floral zygomorphy might be easily altered (Neal et al., 1998; Spencer and Kim, 2018).
The molecular mechanism underlying the floral zygomorphy is centered on CYCLOIDEA (CYC) dorsi-ventral asymmetric expression. CYC is belong to TCP
transcription factor family, in ECE-CYC2 clade. This TF family is characterized by the amino acid basic helix–loop–helix (bHLH) motif in its encoded proteins. The TCP is named after TEOSINTE BRANCHED1 (TB1) from maize (Zea mays), CYCLOIDEA (CYC) from snapdragon (A. majus), and PROLIFERATING CELL
FACTORS 1 and 2 (PCF1 and PCF2) from rice (Oryza sativa). Based on the differences
within the TCP domain, TCP transcription factors are classified into TCP class I that consists of rice PCF proteins and TCP class II that consists of TB1 and CYC proteins.
Outside of the TCP domain, there is 18–20 residue arginine-rich motif (the R domain) which is found in some of class II TCPs, but absent in almost all of the class I TCPs. The TCP class II is then further divided into CYC/TB1 (ECE) and CIN clades. The ECE clade
is characterized with glutamic acid-cysteine-glutamic acid motif found between TCP and R domains. Upon duplication, this clade is divided into CYC1, CYC2 and CYC3. The CYC2 gene group is considered as the major regulator of floral symmetry (Martín-Trillo
and Cubas, 2009).
In Antirrhinum majus, the dorsal specific expression of AmCYC and its close related protein DICHOTOMA (AmDICH) generates specific dorsal shape and size by regulating the cell proliferation and expansion, and additionally inhibiting the stamens growth. Both of the genes inhibit the expression of ventral determinant gene, DIVARIVATA (AmDIV).
This inhibition is mediated through RADIALIS (AmRAD), as the AmRAD protein competes with AmDIV for the interaction with DIV-and-RAD-interacting-factors (DRIFs). Interaction of AmDIV with DRIF is important for the activation of genes that are important for ventral identity. In dorsal petal, AmDIV interaction with DRIF is distracted by the presence of AmRAD, thus making the AmDIV become restricted to be only in ventral petals (Supplementary Fig. S1A; Spencer and Kim, 2018). The absence of AmCYC and AmDICH in A. majus cyc;dich double mutant causes no restriction of AmDIV to the dorsal area, thus the mutant flowers become ventralized and have actinomorphic appearance. Moreover, cyc mutant alone produces semipeloric flowers and the dich single mutant only alters dorsal petal shape. As the mutation analysis shows that
the key regulator of floral zygomorphy (Corley et al. 2005; Luo et al. 1999). Besides in A. majus, other CYC2-like genes in the core eudicots also play the major function in
controlling floral zygomorphy due to the strong dorsoventrally asymmetric expression.
Species showing dorsal or along with lateral expression of CYC generally have zygomorphic flowers, whereas the absence of CYC or ubiquitous CYC expression in all petals results in actinomorphic flowers in some species. Therefore, the progression of CYC expression (absent- ubiquitous − dorsal/lateral − dorsal) plays an important role during the transition of actinomorphic to zygomorphic, and also its reversal (Supplementary Fig. S1B-E; Spencer and Kim, 2018).
The wild type of Sinningia speciosa flower exhibits zygomorphic symmetry, while the commercial type has actinomorphic symmetry. The floral zygomorphy in the wild type is regulated by the dorsal specific expression of a single copy of CYC2-like gene (Supplementary Fig. S2; Ye, 2018, unpublished work), SsCYC. The actinomorphic mutant of this flower is caused by 10 bases deletion in this gene, causing it to be inactive (Dong et al., 2018). Since CYC acts as the key regulator of floral zygomorphy in S.
speciosa as if in A. majus, then S. speciosa could be a comparable model to study the
floral zygomorphy regulation. However, the floral zygomorphy regulation of CYC through RAD, as described in A. majus is not conserved in all zygomorphic lineages (Baxter et al., 2007; Costa et al., 2005, Hsu et al., 2018) which means that CYC might
regulate other genes for the generation of zygomorphic symmetry. Yet, what these target genes of CYC are and how they cooperate to generate petal shape and size for floral zygomorphy are still largely unknown.
The bHLH domain of TCP TF has the capability for binding to GC rich DNA sequences and also for protein-protein interaction. The basic region of this domain mediates the interaction between the protein and targeted DNA sequences, whereas the HLH region provides protein-protein interaction by forming homo- or hetero-dimer (Atchley and Fitch 1997). Both two classes of TCP TF have distinct but overlapping consensus of DNA binding sequences (TCP binding sites); GGNCCCAC for class I and GTGGNCCC for class II, with GGNCCC serves as the core sequence (Koshugi and Ohashi, 2002). TCP binding sites have been reported to role as the cis-elements that mediate TCP TFs regulation of their targets, such as AmRAD (Costa et al., 2005), CYCLIN (Li et al. 2005), PCNA (Kosugi and Ohashi 1997), LIPOXIGENASE2 (Schommer et al.
2008), CIRCADIAN ASSOCIATED1 (Pruneda-Paz et al. 2009), etc. Therefore, the presence of TCP binding sites could be the indicator for determining TCP TFs’ targets, including CYC’s targets (Koshugi and Ohashi 2002).
The recent study in S. speciosa ‘Espirito Santo’ (SsES) has revealed that there were 630 dorsi-ventral differentially expressed genes (DEGs) (Pan, Z.J., unpublished data).
the floral zygomorphy of S. speciosa, including SsCYC downstream. In order to minimize the scope for the screening of SsCYC downstream, this study focused mainly on the TFs activated by SsCYC. TF is known for its effect on a single developmental module which influences only the morphology of a single organ. As the consequence, TF is naturally selected as the source of phenotypic variation. Therefore, mapping SsCYC target TFs will provide a better rationale of how the floral zygomorphy in S. speciosa is established. As TCP binding sites serve as the important elements that might mediate CYC regulation, the identification of SsCYC targets from the DE-TFs of SsES relied on the presence of the TCP binding sites at their 5’regulatory regions. In order to narrow down to SsCYC activation target TFs, the identification was focused on the TFs that had similar expression pattern with SsCYC, which were the dorsal-expressed TFs. The regulation of these TFs by SsCYC was then demonstrated by dual-luciferase assay, transiently expressed in the protoplasts of Nicotiana benthamiana leaves, with SsCYC as the effector and the 5’
regulatory region fused with firefly luciferase as the reporter.
Flowers with zygomorphic symmetry often have their petals could be distinguished into dorsal, lateral and ventral parts due to the different shape and size within these regions, which leads to the hypothesis that the petal identity of each region should have some effects to the establishment of floral zygomorphy. Petal identity itself is determined by two factors, which are cell elongation as well as rate and direction of cell division. The
cell elongation in the basal part is important for determining the final size and shape of the petal, while the rate and direction of cell division determine the shape and size of the distal region. This mechanism requires a quite complex hormonal regulation (Irish, 2008;
van Es, 2018). Jasmonic acid influences the petal size of Arabidopsis through post- transcriptional regulation of BIGPETAL (BPE), TF that regulates cell expansion (Brioudes et al., 2009). Auxin, ethylene and gibberellin also affect cell proliferation and elongation during petal development by integrating in certain TF regulations (Chandler 2011).
In this study, several TFs were found to be responsive to SsCYC. Instead of SsRAD, orthologue of Anthirrhinum RAD; another RAD-like gene (SsRL2) whose function was unknown, was identified as SsCYC downstream. Interestingly, two ethylene responsive factors (SsERFs) were also found to be regulated by SsCYC. ERFs are known as the
integral components of signaling cascade that regulate different kinds of downstream genes of various developmental and stress responsive pathways. As the downstream component of ethylene signaling pathway, ERFs also interact with other hormone pathways, such as jasmonic acid, ABA, auxin, salicylic acid, gibberellins, and brassinosteroids (Müller and Munné-Bosch, 2015). Moreover, an ovate family protein (SsOFP6) whose overexpression in Arabidopsis results in flat, thick and cyan leaves
SsCYC. Taken together, these results led to the suggestion that SsCYC might work through these TFs to affect the dorsal petals cell growth of SsES, developing the floral zygomorphy of this flower.
Materials and Methods
Plant material and growth condition
Sinningia speciosa ‘Espirito Santo’ was obtained from Dr. Cecilia Koo Botanic
Conservation Center, Pingtung, Taiwan. The seeds were cultivated under 16/8 hours (day/night) cycle at 24℃ with 70% relative humidity. Floral bud developmental stage was determined based on dorsal corolla tube length. Floral bud stage 5 which has 8-10 mm length of dorsal tube was used for transcription factor (TF) isolation. Dissected dorsal and ventral petals from floral bud stage 5 were used for expression pattern validation of the dorsal expressed TFs. Finally, the leaves were used for 5’ regulatory region isolation.
All samples were frozen in liquid nitrogen and stored at -80℃.
Prediction of transcription factor
RNA-seq data of S. speciosa ‘Espirito Santo’ floral bud stage 5 was provided and analyzed by Dr. Zhao-Jun, Pan. Based on RNA-seq analysis, 630 genes were found to have dorsi-ventral differential expression (DEGs) (p-value<0.05; log2FC ≥1). In order to find the TFs among these DEGs, TF prediction was performed using iTAK online (v1.6) (http://itak.feilab.net/cgi-bin/itak/online_itak.cgi) for nucleotide sequences. The
(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastx&PAGE_TYPE=BlastSearc h&LINK_LOC=blasthome) for annotation.
Prediction of TCP binding site
Since SsCYC downstream regulation might be facilitated by the presence of TCP binding sites at the 5’regulatory region of its target genes, screening for the binding sites was done for each of the predicted TF. The 5’ regulatory region of each TF was retrieved from S. speciosa ‘Avenida Niemeyer’ draft genome using RStudio software (Version 1.1.463; RStudio Inc., 2009) and Linux Interface (done by Ya-Chi, Nien). TCP binding consensus was summarized from the paper ‘TCP Transcription Factors: Evolution, Structure, and Biochemical Function’ (González-Grandío and Cubas, 2016) that has compiled most of TCP binding sites found in the in vitro and in vivo experiments in numerous studies. Screening for the presence of each of the summarized TCP binding consensus (Supplementary Table S1) was done for all the predicted regulatory regions using fuzznuc (http://emboss.bioinformatics.nl/cgi-bin/emboss/fuzznuc) for both strands of complementary sequence.
Total RNA extraction and reverse transcription
The total RNA from whole floral bud stage 5 and dissected dorsal and ventral petals of floral bud stage 5 were extracted using Trizol Reagent (Invitrogen, Waltham, MA, USA) according to manufacturer’s protocol. The RNA quality was measure using NanoDrop Spectrophotometer. Synthesis of complementary DNA (cDNA) was done using Superscript IV (Invitrogen, Waltham, MA, USA) according to manufacturer’s protocol.
Isolation of the transcription factor of S. speciosa ‘Espirito Santo’
TFs that have been predicted to contain TCP binding sites at their 5’ regulatory regions were isolated in order to get their full length coding sequences. The sequence of each TF was amplified with PCR using Phusion® High-Fidelity DNA Polymerase (New England Biolabs, Ipswich, MA, USA) (Supplementary Table S2) and the products were purified by gel extraction (Viogene, GP1002), following the manufacturer’s protocol. The purified products were proceed to A-tailing in order to increase ligation efficiency. A- tailing was done by adding 0.3 μL of TaKaRa Ex Taq DNA Polymerase (Takara Bio, USA), 3 μL of Ex Tag buffer and 0.6 μL of 2mM dATP into 10 μL of purified product and the mixture was then incubated at 72℃ for 1 hour. The A-tailed products were purified by PCR Clean Up system (Viogene, GP1002) and were ligated to T&A™ cloning
Ligation mixture component vector: insert molar ratio 1:3 Vector fragments end conc. 3-30 fmol Insert fragments end conc. 9-90 fmol 10x Ligation Buffer A 2.0 μL 10x Ligation Buffer B 2.0 μL
yT4 DNA ligase 1.0 μL
ddH2O to final volume of 20 μL
The ligation mixture was incubated overnight. The next day, transformation was done using the heat shock method. About 2 μL of vector containing DNA of interest was mixed with 20 μL of competent cell, Escherichia coli HIT-DH5α (Real Biotech Corporation,
Taipei, Taiwan) and was chilled on ice for 20 minutes. Then, the mixture was thawed at 42℃ for 1 minute for heat shock and quickly chilled on ice. After heat shock procedure, 50 μL of LB broth was added to the mixture, followed by incubation at 37℃ for 1 hour.
The bacterial solution was added with 100 μL of 0.1 M IPTG and 20 μL of 80 mg/mL X-
Gal, and spread on LB agar plate contained Ampicillin (100 μg/mL). The plate was incubated for 16-18 hours. Colonies containing the insert were selected by using colony PCR. After confirmation, the colonies containing the correct insertion size of DNA was cultured in 3 mL of LB broth contained Ampicillin (100 μg/mL) by shaking at 37℃ for 16-18 hours. The plasmids were extracted using Mini Plus Plasmid DNA Extraction System (Viogene, GF2002) according to manufacturer protocol and sent to sequencing (Genomics, New Taipei City, Taiwan).
Validation for the expression pattern of the dorsal-expressed TFs of S. speciosa
‘Espirito Santo’
Quantitative real time PCR (qRT-PCR) analysis was done to validate the RNA-seq
data of the dorsal-expressed TFs that have been predicted to contain TCP binding sites at their 5’ regulatory regions. qRT-PCR analysis was performed in Bio-Rad PCR machine (CFX-384) using KAPA SYBR® FAST qPCR Master Mix (2X) Kit (KAPA Biosystem, KR0389) (Supplementary Table S3). The recipe and program were listed below:
qRT-PCR mixture
Reagent Volume
ddH2O 1.0 μL
2x Master Mix 5.0 μL Forward Primer (1μM) 1.0 μL Reverse Primer (1μM) 1.0 μL cDNA (5ng/μL) 2.0 μL Total Volume 10 μL Thermal cycle program:
Step Temperature Time
1 95 ℃ 3 min
2 95 ℃ 10 s
3 55-57 ℃ 30 s
Plate read
4 Go to step 2, 39 cycles
5 95 ℃ 10 s
6 Melt curve 65 to 95℃, increment 0.5 5 s Plate read
After the running of PCR, the obtained data was analyzed using CFX Maestro™ Software for CFX Real-Time PCR Instruments (Version 1.1; Bio-Rad Laboratories Inc, 2017). The expression level of each TF was quantified as relative fold gene expression level (2-ΔΔCT), using 18s as reference gene and ventral petals as the control. The ∆Ct was calculated as Ct (dorsal/ventral) – Ct (reference gene) and the ∆∆Ct was calculated ∆Ct (dorsal petals) – ∆Ct (ventral petals).
Genomic DNA Extraction
Genomic DNA (gDNA) extraction was performed with Hexadecyl trimethyl- ammonium bromide (CTAB) method (Doyle, 1990). The collected leaves were homogenized in liquid nitrogen using mortar and pestle. The homogenized tissue was added with 1 mL of CTAB, 20 mg of PVPP and 5 μL of β-mercaptoethanol, proceed by
incubation at 65℃ for 30 minutes. Next, the mixture was added with 500 μL of PCI (phenol : choloroform : isoamyl alcohol, 25:24:1, pH = 8.0) and inverted for 15 minutes, followed by centrifugation at 13.000 rpm for 10 minutes. The upper layer of the solution was transferred to the new tube, added with 1 μL RNase A and incubated at 37℃ for 20- 30 minutes. The solution was added with 500 μL of C:I (choloform: isoamyl alcohol, 24:1)
and inverted for 15 minutes, followed by centrifugation at 13.000 rpm for 10 minutes.
The upper layer was transferred to the new tube and added with one to tenth volume of 3
M NaOAc (pH = 5.5), then precipitated with 0.7 volume of isopropanol. The mixture was
incubated at -20 ℃ for 1 hour, proceed by centrifugation at 13.000 rpm for 10 minutes.
The supernatant was discarded and the pellet was washed by the addition of 1 mL of 70%
ethanol and centrifugation at 13.000 for 5 minutes. The supernatant was discarded and the pellet was air dried. Finally, 30-50 μL of ddH2O was added to dissolve the pellet. The
quantity and quality of extracted gDNA was measured with Nanodrop Spectrophotometer.
The gDNA was stored at -20 ℃.
CTAB buffer (100 mL)
Reagent per reaction
Hexadecyl trimethyl-ammonium bromide (CTAB) 2.0 g
1M Tris (pH = 8.0) 10.0 mL
0.5 Ethylenediaminetetraacetic acid (EDTA, pH = 8.0) 4.0 mL
5 M NaCl 28.0 mL
ddH2O 56.0 mL
The pH was adjusted to 8.0 using NaOH and stored at room temperature
Isolation of the 5’ regulatory region of Sinningia speciosa ‘Espirito Santo’
There were several PCR based approaches used for isolating the 5’ regulatory region of each dorsal-high expressed TF. The regulatory region of Sispe038Scf1202g12026 (SsOFP6) was isolated using pair of primers designed directly
from the predicted regulatory region of S. speciosa ‘Avenida Nieyemer’ (SsAN). The regulatory region of Sispe038Scf1400g01001 (SsCYC) was isolated with forward primer
designed at the known coding sequence (CDS) of S. speciosa ‘Espirito Santo’ (SsES).
Another two regulatory regions, Sispe038Scf1061g02075 (SsERF3) and Sispe038Scf2159g01072 (SsCIB2), were isolated by nested PCR using two sets of primers.
The first set of primers contained the forward primer designed directly from the predicted regulatory region of SsAN and reverse primer designed on the known CDS of SsES. The second set of primers was design to amplify a secondary target within the first run product, thus reducing the non-specific binding in products. For the second round of the nested PCR, the product of the first PCR was diluted to 100 times. All of these three approaches were done with Phusion® High-Fidelity DNA Polymerase (New England Biolabs, Ipswich, MA, USA) according to manufacturer protocol (Supplementary Table S4). The amplified products were continued to cloning, using the same procedure described for transcription factor isolation and then sent to sequencing (Genomics, New Taipei City,
Taiwan). Last, the regulatory region of Sispe038Scf0228g08027 (SsERF17) was isolated with Thermal Asymmetric Interlaced PCR (TAIL-PCR), whereas for Sispe038Scf0170g01016 (SsRL2), Sispe038Scf2996g00029 (SsNGAL1) and Sispe038Scf5680g00016 (SsERF1), the regulatory regions were isolated with Self-
Formed Adaptor PCR (SEFA-PCR). All the isolated regulatory regions were screened for the presence of TCP binding sites.
Thermal Asymmetric Interlaced PCR (TAIL-PCR)
Thermal Asymmetric Interlaced PCR (TAIL-PCR) is used to amplify the unknown sequence, in this case the regulatory region that is adjacent to the known CDS. It uses two sets of primers which are the gene-specific primers (GS primers) that usually have high melting temperatures and arbitrary degenerate primers (AD primers) (Supplementary Table S5) that usually have low melting temperatures. By using the combination of these
primers, amplification of the expected sequence could be done from the known end and the unknown end, respectively. Specificity is obtained through subsequent rounds of TAIL-PCR, using nested gene-specific primers and alternate of high and low annealing temperatures cycles (Supplementary Fig. S3a). The TAIL-PCR used in this study was referred from Liu et al. (1995) and Liu and Whittier (1995) with modifications. The AD primers were adopted from Singer and Burke (2003). The recipe and program of TAIL- PCR were listed below:
Single reaction for primary TAIL-PCR
Reagent Volume
Phusion DNA polymerase (0.02 units/μL) 0.1 μL 5X Phusion HF or GC Buffer 2.0 μL
10 mM dNTPs 0.2 μL
6 x AD primer 2.0 μL
10 µM GS1 primer 0.5 μL
gDNA (20 ng/μL) 0.5 μL
ddH2O add to 10 μL
Thermal cycle for primary TAIL-PCR
Step Temperature Time
1 94 ℃ 2 min
2 94 ℃ 30 s
3 62 ℃ 1 min
4 72 ℃ 2.5 min
5 Go to step 2 for 4 cycles
6 94 ℃ 30 s
7 25 ℃ 3 min
8 Ramping from 25 to 72 ℃ (rate = 0.3℃/sec)
9 72 ℃ 2.5 min
10 94 ℃ 10 s
11 68 ℃ 1 min
12 72 ℃ 2.5 min
13 94 ℃ 10 s
14 68 ℃ 1 min
15 72 ℃ 2.5 min
16 94 ℃ 10 s
17 44 ℃ 1 min
18 72 ℃ 2.5 min
19 Go to step 10 for 14 cycles
20 72 ℃ 2.5 min
Single reaction for secondary TAIL-PCR
Reagent Volume
Phusion DNA polymerase (0.02 units/μL) 0.1 μL 5X Phusion HF or GC Buffer 2.0 μL
10 mM dNTPs 0.2 μL
6 x AD primer 2.0 μL
10 µM GS1 primer 0.5 μL
1:1000 diluted 1st reaction 0.5 μL
ddH2O add to 10 μL
Thermal cycle for secondary TAIL-PCR Step Temperature Time
1 94 ℃ 10 s
2 68 ℃ 1 min
3 72 ℃ 2.5 min
4 94 ℃ 10 s
5 68 ℃ 1 min
6 72 ℃ 2.5 min
7 94 ℃ 10 s
8 44 ℃ 1 min
9 72 ℃ 2.5 min
10 Go to step 1 for 11 cycles
11 72 ℃ 5 min
Single reaction for tertiary TAIL-PCR
Reagent Volume
Phusion DNA polymerase (0.02 units/μL) 0.1 μL 5X Phusion HF or GC Buffer 2.0 μL
10 mM dNTPs 0.2 μL
6 x AD primer 2.0 μL
10 µM GS1 primer 0.5 μL
1:1000 diluted 3rd reaction 0.5 μL
ddH2O add to 10 μL
Thermal cycle for tertiary TAIL-PCR Step Temperature Time
1 94 ℃ 15 s
2 44 ℃ 1 min
3 72 ℃ 2.5 min
4 Go to step 1 for 19 cycles
5 72 ℃ 5 min
After the 3rd round of PCR, the products from 1st, 2nd and 3rd PCR were run together in gel electrophoresis. The product from the 1st round might contain the non-specific
products which could be seen by the smear appearance on the gel. The expected specific products could usually be observed from the product of 2nd and 3rd round, with the 3rd round product having slight decreased in size. The largest band from the 3rd round product was isolated and continued to cloning, using the same procedure described for transcription factor isolation, then sent to sequencing (Genomics, New Taipei City,
Taiwan) (Supplementary Fig. S3b).
Self-Formed Adaptor PCR (SEFA-PCR)
Self-Formed Adaptor PCR (SEFA-PCR) is developed to overcome the drawbacks of TAIL-PCR, which is the product is usually less than 1.0 kb. It combines the advantages of ligation-mediated PCR in its specificity and of TAIL PCR in its simplicity. It uses four primers that are located sequentially on the known DNA sequences. SP1, SP2, and SP4 are the specific primers designed from the known region and have relatively high annealing temperatures (e.g., 70°C), whereas SP3 (e.g., 5′- TACCCAAAGAAGCAGGAANNNNNNNNGTGAAA-3′) is a partially degenerate primer which plays the key role in the process. First, a single cycle of PCR was carried out at a low annealing temperature (e.g., 35°C) with only primer SP3. At this low annealing temperature, SP3 can prime and elongate at many positions on the DNA template. A position probably exists somewhere downstream of the known DNA sequence
where SP3 primes and extends, thus creating a nascent single strand which has a binding site for SP1. After a single cycle of PCR, the annealing temperature is increased to the point (e.g., 70°C) corresponding to the annealing temperature of SP1. Then, SP1 is added to the reaction mixture. At this high annealing temperature, only SP1 can prime the target site efficiently, thus creating a pool of single-stranded DNA with the SP1 sequence at the 3’ end and the SP3 complementary sequence at the 5’ end. Finally, several cycles of a low annealing temperature (e.g., 55°C) are performed to facilitate the loop-back extension, thus creating an adaptor which contains binding sites for SP1 and SP2. Once the adaptor has been created, the target sequences can be amplified efficiently by SP1. After SEFA PCR, a second round of nested PCR was run with the single primer SP2. A third round of thermally asymmetric PCR was run to improve the specificity with primer SP4 (e.g., annealing at 70°C) and the other short primer, SP5 (e.g., annealing at 60°C), positioned between SP2 and SP3 (Supplementary Table S6; Supplementary Fig. S4a). The SEFA- PCR used in this study was adopted from Wang et al. (2007) with modifications. The recipe and program were listed below:
Single reaction for primary SEFA-PCR
Reagent Volume
Phusion DNA polymerase (0.02 units/μL) 0.2 μL 5X Phusion HF or GC Buffer 4.0 μL
10 mM dNTPs 0.4 μL
5 μM SP3 1.0 μL
gDNA (1000 ng/μL) 1.0 μL
ddH2O add to 20 μL
Thermal cycle for primary SEFA-PCR
Step Temperature Time
1 98 ℃ 30 s
2 35 ℃ 3 min
3 Ramping from 35 to 70 ℃(rate = 0.2℃/sec) 4 Add 3 μl of 5 μM SP1
5 98 ℃ 10 s
6 70 ℃ 3 min
7 Go to step 5 for 24 cycles
8 98 ℃ 10 s
9 70 ℃ 3 min
10 98 ℃ 10 s
11 70 ℃ 3 min
12 98 ℃ 10 s
13 65 ℃ 30 s
14 70 ℃ 3 min
15 Go to step 8 for 10 cycles
16 25 ℃ 10 s
Single reaction for secondary SEFA-PCR
Reagent Volume
Phusion DNA polymerase (0.02 units/μL) 0.1 μL 5X Phusion HF or GC Buffer 2.0 μL
10 mM dNTPs 0.2 μL
5 uM SP2 3.0 μL
1:10 diluted 1st reaction 0.5 μL
ddH2O add to 10 μL
Thermal cycle for secondary SEFA-PCR
Step Temperature Time
1 98 ℃ 30 s
2 98 ℃ 10 s
3 70 ℃ 3 min
4 Go to step 2 for 29 cycles
5 25 ℃ 10 s
Single reaction for tertiary SEFA-PCR
Reagent Volume
Phusion DNA polymerase (0.02 units/μL) 0.1 μL 5X Phusion HF or GC Buffer 2.0 μL
10 mM dNTPs 0.2 μL
5 uM SP4 3.0 μL
5 uM SP5 0.3 μL
1:10 diluted 1st reaction 0.5 μL
ddH2O add to 20 μL
Thermal cycle for tertiary SEFA-PCR
Step Temperature Time
1 98 ℃ 10 s
2 70 ℃ 3 min
3 98 ℃ 10 s
4 70 ℃ 3 min
5 98 ℃ 10 s
6 65 ℃ 30 s
7 70 ℃ 3 min
8 Go to step 1 for 9 cycles
9 25 ℃ 10 s
After the 3rd round of PCR the products from 1st, 2nd and 3rd PCR were run together in gel electrophoresis. The 1st and 2nd might contain some non-specific products with low molecular weight, and the desired product is usually expected to be seen in the 3rd product.
Therefore, the largest band from 3rd product was isolated and sent to sequencing
(Genomics, New Taipei City, Taiwan) (Supplementary Fig. S4b). New forward primers were design to amplify the desired regulatory region paired with SP4 primers, using the
same procedure as described in the transcription factor isolation (Supplementary Table S4).
Vector construction for dual-luciferase assay
The PJD301-firefly driven by the 5’ regulatory region of interest was used as the reporter (Supplementary Fig. S5a), whereas PJD301-renilla driven by 35s promoter was used as the internal control to normalized the transfection variability (Supplementary Fig. S5b) (Luehresen et al., 1995). The vector expressing SsCYC tagged with GFP was
served as the effector for the tested group (Supplementary Fig. S6), whereas vector expressing only GFP without SsCYC was used as effector for the control group (Supplementary Fig. S7).The isolated regulatory region sequence of SsRL1, SsERF17, SsOFP6, SsCYC, SsCIB2, and SsNGAL1 were amplified using PCR and cloned into the
BamHI and SalI restriction sites of the PJD301-firefly, whereas SsERF3 and SsERF1 were amplified by PCR to add HincII and NCO1 restriction site and cloned into the AfeI and NcoI restriction site of the vector (Supplementary Table S7). The general recipe for enzyme digestion was described as below:
Recipe for BamHI and Sal1 digestion
Reagent Volume
BamHI buffer 10.0 μL
BamHI (10 U/µ L) 2.5 μL SalI (10 U/µL) 5.0 μL
DNA 400-500 ng
ddH2O add to 100 μL
Recipe for HincII and NcoI digestion
Reagent Volume
1X Tango Buffer 10.0 μL HincII (10 U/µL) 5.0 μL NcoI (10 U/µ L) 5.0 μL
DNA 400-500 ng
ddH2O add to 100 μL
Recipe for Afe1 and NcoI digestion
Reagent Volume
2X Tango Buffer 20.0 μL HincII (10 U/µL) 2.5 μL NcoI (10 U/µ L) 2.5 μL
DNA 400-500 ng
ddH2O add to 100 μL
The reaction mixtures were incubated overnight and the desired digestion products were purified by gel purification (Viogene, GP1002), following the manufacturer’s protocol.
The purified products were ligated to the PJD301-firefly vector following the recipe described below:
Ligation reaction of PJD-firefly with the desired digestion product PJD301: insert molar ratio 1:3
PJD301 fragments end conc. 3-30 fmol Insert fragments end conc. 9-90 fmol 10x Ligation Buffer A 2.0 μL 10x Ligation Buffer B 2.0 μL
yT4 DNA ligase 1.0 μL
ddH2O to final volume of 20 μL
The ligation mixture was incubated overnight. The next day, transformation was done using the heat shock method into the Escherichia coli HIT-DH5α (Real Biotech
Corporation, Taipei, Taiwan). Amipicillin (100 μg/mL) plate was used as the selection medium. After 16-18 hours of incubation colony PCR was done to select the colony carrying the vector of interest. The colony that has been confirmed to carry the desired vector was cultured into LB contained Ampicillin (100 μg/mL) for maxi plasmid extraction (Viogene, GMV2002).
Protoplast isolation
Protoplast isolation was done according to ‘Arabidopsis mesophyll protoplasts protocol’ (Yoo et al., 2007) with modifications. Nicotiana benthamiana leaves were used
as the source of protoplasts instead of Arabidospsis. The plants were grown under 16/8 hours (day/night) cycle at 27℃ with 70% relative humidity. The leaves from 4-5 weeks- old-plant were chosen and cut into 0.5–1-mm strips from the middle part of a leaf using
a fresh sharp razor blade. The cut leaves were transferred into the prepared enzyme solution and digested in the dark for 3 hours at room temperature. After digestion, the solution was diluted with an equal volume of W5 solution. The enzyme solution containing the protoplasts was filtered through 75-μm nylon mesh into round-bottom tube.
The filtered solution was then centrifuged at 100 g for 2 minutes. The supernatant was removed and the protoplasts were re-suspended with W5 solution at 2 × 105 ml−1 after counting cells under the microscope (× 100) using a hemacytometer. The protoplasts were rested on ice for 30 minutes. After 30 minutes, the W5 solution was removed and the protoplasts were re-suspended in MMG solution 2 × 105 ml−1.
Protoplast DNA-PEG–calcium transfection
The protoplast transfection of vector mixture containing effector, reporter and
internal control was also performed following the method described in ‘Arabidopsis
mesophyll protoplasts protocol’ (Yoo et al., 2007). About 10 μL of vector mixture (the amount of each vector was 10 μg in 10 μL) was added into a 2-ml microfuge tube, followed by 100 μl protoplasts (2 × 104 protoplasts), then the mixture was mixed gently.
About 110 μl of PEG solution was added and the mixture was mixed gently by tapping the tube. The transfection mixture was incubated for 15 minutes at room temperature.
by inverting. The mixture was centrifuge at 100 g for 2 minutes and the supernatant was removed. The protoplasts were re-suspended in 0.5 mL of WI solution in each well of a 12-well tissue culture plate. Incubation was done for 16 hours.
Experimental design for SsCYC and dorsal-expressed TFs interaction analysis
Test Control
Vector Mixture
SsCYC-GFP effector GFP effector
5'regulatory region-PJD301 Firefly 5'regulatory region-PJD301 Firefly
PJD301 Renilla PJD301 Renilla
Dual-luciferase assay.
After 16 hours of incubation, the transfected protoplasts were collected by moving them to 2 mL microfuge tube, followed by centrifugation at 100 g for 2 minutes and the supernatant was removed. The dual-luciferase assay was done in 96-well white flat bottom plate according to the instruction of Dual-Luciferase® Reporter Assay System for product E1960 (Promega Corporation, USA). About 20 μl passive lysis buffer was added into the protoplasts and the mixture was transferred into the well of the plate. After 5 minutes, 100 μl of LAR II reagent was added into the mixture and the firefly luciferase activity was measured by luminometer by 10s measurement using i-control™ Microplate Reader Software (Version 1.8; Tecan, 2011). Then, 100 μL of Stop & Glo® was added and the renilla luciferase activity was measured by 10s
measurement. All reactions were run triplicate. The interaction of SsCYC and its downstream target was determined as normalized fold change (Δfold activity) by calculating the firefly to renilla activity ratio of the tested group divided to the control group. One-Way Analysis of Variance was used to assess the up/down-regulation significance level, using One-Way Analysis of Variance Calculator (https://goodcalculators.com/one-way-anova-calculator/).
Results
34 Transcription factors were predicted among 630 dorsi-ventral DEGs
The RNA-seq data has shown that there were 630 dorsi-ventral DEGs of S.
speciosa ‘Espirito Santo’. In order to screen for the TFs among these DEGs, iTAK was used as the identification and classification tool. Around 34 TFs were identified; 17 of them were the dorsal-expressed TFs and the others 17 were the ventral expressed TFs.
Based on NCBI BLASTX analysis, CYC (SsCYC) which was previously known as the major regulator of floral zygomorphy of A. majus was identified in the dorsal-expressed TF group (Table 1).
Table 1 The list of 34 TFs identified from 630 DEGs and their BLASTX annotations
19 out of 34 TFs were enriched with TCP binding sites at their predicted 5’
regulatory regions
TCP binding site has been known as the important element that mediates gene regulation of TCP TF family. Basically, TCP binding site is classified into two classes
with the consensus of GGNCCCAC for class I and GTGGNCCC for class II. Most of genes that are regulated by TCP-TFs are usually enriched with these binding sites.
Moreover, regulation of these genes through the binding of TCP TFs at these binding sites has also been confirmed either in the in vitro or in vivo analysis. It is also important to be noted that each TCP TF might have different preference of TCP binding sites. There are also some evidences that the recognized binding site motifs of TCP TF might not always follow the exact common consensus of GGNCCCAC or GTGGNCCC. For instance, some TCP-TFs have been found to bind to the motif GAGGGACCCT, TTGGGACCTC, GTGGGAACCA (classified as class I), tGGKMCCa, GGACCA, tGGGtCCAC, and TGGKGCC (classified as class II) which actually do not resemble class I or class II consensus. Another case is that some TCP TFs have also been reported to recognize the binding motif TGGGC(C/T) or GGNCCCNC which is the combination of both class I and class II consensus, thus classified as class I&II (González-Grandío & Cubas, 2016).
Since SsCYC belongs to TCP TF family, SsCYC downstream regulation might also be facilitated by the presence of TCP binding sites at the regulatory region of its downstream, suggesting that the presence of TCP binding sites at the 5’regulatory region is the important indicator to determine SsCYC downstream among the dorsi-ventral DE- TFs. Therefore, the 2 kb sequences of 5' regulatory region of each TF were retrieved from the draft genome of S. speciosa ‘Avenida Niemeyer’ as it was the only available genome
data. The retrieved sequences were then screened for the presence of TCP binding consensus, summarized from the paper ‘TCP Transcription Factors: Evolution, Structure, and Biochemical Function’ (González-Grandío & Cubas, 2016) which included TCP class I, TCP class II, combination of both class I and class II, as well as the unique sequences (the ones that not resemble both classes) that have been proved to be bound by TFs of TCP family.
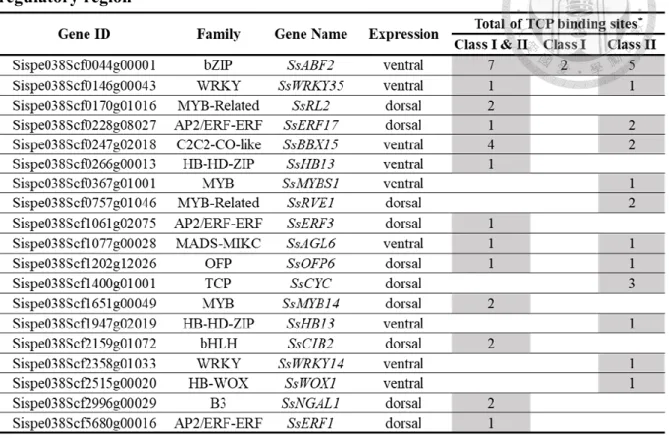
Among 34 DE-TFs, there were 19 TFs that were predicted to contain TCP binding sites at their 5’ regulatory regions; 9 of them, including SsCYC were the dorsal-expressed TFs and 10 of them were the ventral expressed TFs. Most of these TFs were enriched with TCP class I&II and class II binding sites (Table 2). This result suggested that these TFs might have the possibility as SsCYC downstream target. However, it is also possible that they might be regulated by other TCP TFs.
Table 2 The list of 19 TFs predicted to contain TCP binding sites at their 5’
regulatory region
*TCP binding consensus found:
Class I&II: GGNCCCNC and TGGGC(C/T) Class I: GTGGGNCC
Class II: tGGKMCCa, GGACCA, and TGGKGCC
The expression pattern of dorsal-expressed TFs was consistent with the RNA-seq
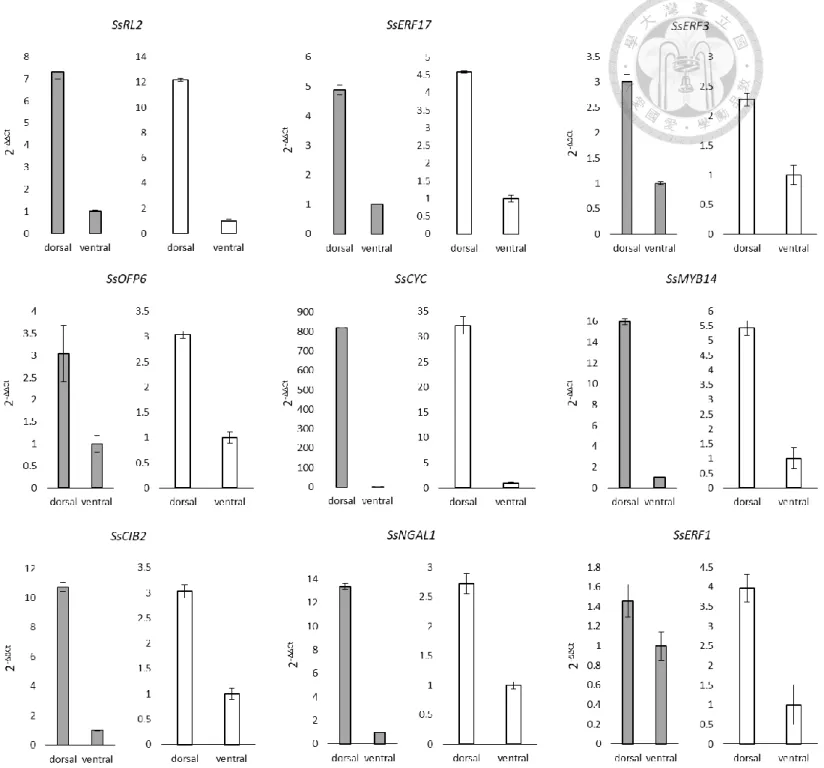
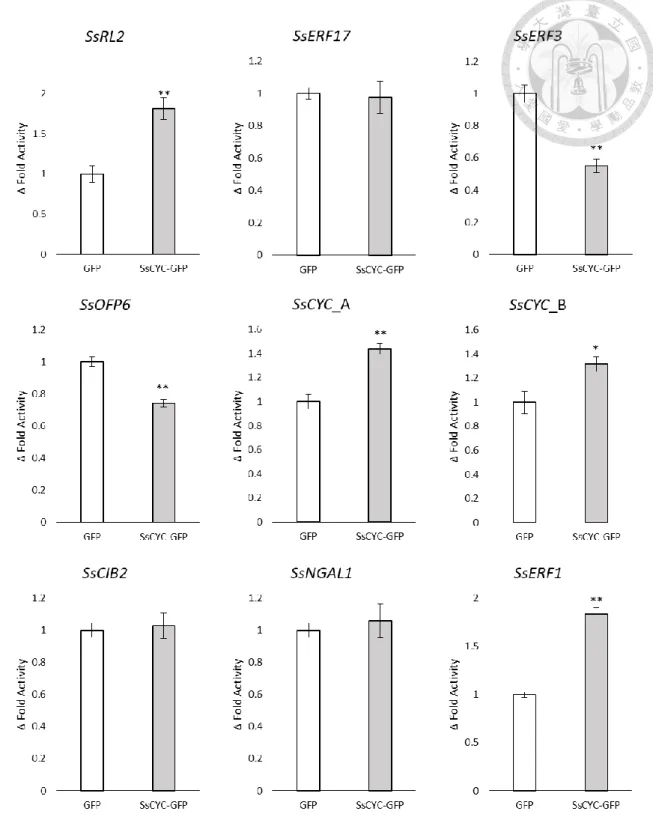
In order to narrow down the possible SsCYC downstream TFs, this study focused on those TFs that might be the activation targets of SsCYC. These TFs should be those that have the similar expression pattern with SsCYC, which then should be the dorsal- expressed TFs. The qRT-PCR result showed that the 9 dorsal-expressed TFs expression pattern was consistent with the RNA-seq data, confirming their possibility as SsCYC activation targets (Fig. 1).
Figure 1 qRT-PCR confirmation of dorsal-expressed TFs that have been predicted to have TCP binding sites at their 5’ regulatory region
The grey bars represent the qRT-PCR results and the white bars represent the RNA-seq results, expressed as the mean of relative fold gene expression level (2-ΔΔCt)± standard error of mean. 18s was used as reference gene and ventral expression level was used as control.
All the isolated 5’ regulatory regions of S. speciosa ‘Espirito Santo’ dorsal-expressed
TFs contained TCP binding sites
Since the 9 dorsal-expressed TFs have the consistent expression pattern with the RNA-seq data, then isolation of the 5' regulatory region of these TFs from S. speciosa
‘Espirito Santo’ (SsES) was conducted by PCR based methods. The reverse primer of each regulatory region was designed to facilitate overlap at the 3' with the beginning of the coding sequence (CDS) of the corresponding TF, except for SsOFP6.
The 5' regulatory regions that were successfully isolated were those belong to SsCYC, SsRL2, SsERF17, SsERF3, SsOFP6, SsCIB2, SsNGAL1 and SsERF1. Their
lengths were varied between almost 1 to 2 kb (Table 4). Each isolated regulatory region of SsES showed similarity ranging from 75% (SsERF1) to ~98% (SsCYC) when aligned with the predicted sequence of S. speciosa ‘Avenida Niemeyer’ (SsAN) (Table 3), indicating that regulatory sequence variations might appear within cultivars. All the isolated regulatory regions, with the exception of SsOFP6 also have their 3' sequences overlap with the beginning of the CDS of their respective TFs. These results confirmed that all the obtained regulatory sequences were belong to their respective TFs. The differences between SsAN and SsES regulatory regions were due to several point mutations and indels. Comparing to the other regulatory regions, SsERF1 showed significant differences between SsES and SsAN, which was characterized with frequent
large gaps, caused due to large insertions or deletions and point mutations. Unfortunately, the regulatory region of SsMYB14 was failed to be isolated using all the approaches, thus it did not continue to the remaining analysis. Moreover, the regulatory region of SsCYC in SsES showed two different alleles, also due to indels. The length of these two alleles only differed in 2 bp.
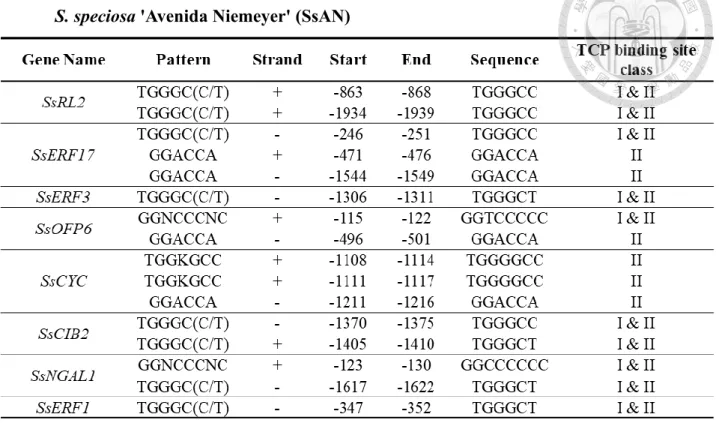
The isolated 5' regulatory regions were also screened for TCP binding sites using the same method described previously. All the isolated 5' regulatory regions contain TCP binding sites. Most of them were enriched with either TCP class I&II or/and class II.
Since the regulatory regions of most TFs were quite similar to the predicted ones, they also shared similar binding consensus at almost similar position, except for SsERF1.
However, the regulatory region of SsRL2 in SsES was lack of 1 binding site that caused due to the shorter length comparing to SsRL2 in SsAN, so that it could not cover the binding site found at the position between (-1934) and (-1939) of SsAN. Similar to SsRL2, SsERF17 in SsES was also lack of 1 binding site which was caused by long deletion so
that it missed the binding site found at the region between (-1544) and (-1549) of SsAN.
In the case of SsOFP6 of SsES, the lacking of 1 binding site was caused due to the change of one base from T to C at position -462, which eliminated this binding site in SsES. As the regulatory sequence of SsERF1 of SsES had pretty low percentage of similarity to
38
sequence and position (Table 3, 4 & 5; Supplementary Fig. S8). Yet, these results still suggested that these TFs might be the target of SsCYC or other TCP TFs.
Table 3 Percentage of identity between the 5’ regulatory region of S. speciosa
‘Espirito Santo’ (SsES) and S. speciosa ‘Avenida Niemeyer’ (SsAN) Gene Name Identity (%)
SsRL2 97.86
SsERF17 95.56
SsERF3 94.75
SsOFP6 97.02
SsCYC_A 98.28 SsCYC_B 98.23
SsCIB2 97.72
SsNGAL1 96.56
SsERF1 75.48
Analysis was done by Clustal MUSCLE tool (http://www.ebi.ac.uk/Tools/msa/muscle/)
Table 4 Summary of TCP binding sites found at the 5' regulatory regions isolated from S. speciosa ‘Espirito Santo’ (SsES)
*SsCYC_A and SsCYC_B refer to SsCYC 5’ regulatory region
Table 5 Summary of TCP binding sites predicted from the 5' regulatory region of S. speciosa 'Avenida Niemeyer' (SsAN)
SsCYC might have the ability to autoregulate itself and regulate other TFs
The interaction of SsCYC with its possible downstream targets was checked by dual-luciferase assay by co-transfecting effector, reporter and internal control into the same protoplasts of Nicotiana benthamiana leaves. The SsCYC-GFP was used as the effector to regulate the firefly luciferase activity driven by the 5’ regulatory region of interest. The detected firefly luciferase signal of each tested regulatory region was normalized by renilla luciferase signal to encounter the transfection variability. The firefly/renila luciferase signal ratio obtained using SsCYC-GFP effector was compared to GFP effector (control) to specify the interaction of SsCYC with the corresponding TFs,
expressed as normalized fold change (Δfold activity).
Some of the dorsal expressed TFs showed response to SsCYC effector. Significant up-regulation by SsCYC was observed in the SsCYC, SsRL2 and SsERF1 regulatory region construct, indicating that they might be the activation target of SsCYC. The ability of SsCYC to activate itself might be considered as a positive autoregulation. In contrast, significant down-regulation by SsCYC was also observed in the SsOFP6 and SsERF3 regulatory region construct, indicating SsCYC might repress these TFs expression. The remaining construct did not show neither activation nor repression by SsCYC (Fig. 2).
Figure 2 Dual-luciferase assay result
SsCYC regulation of the target TFs was expressed as mean of normalized fold change (Δfold activity) ± standard error of mean, determined by calculating the firefly to renilla luciferase activity ratio of the tested group (SsCYC-GFP effector) divided to the control group (GFP effector). The results were analyzed using One-Way Analysis of Variance; *P-value<0.05, **P-value<0.01.
Discussion
Floral zygomorphy study has mainly focused on the dorsi-ventral asymmetric expression of transcription factor (TF) CYCLOIDEA (CYC) which belongs to TCP TF family class II. The role of CYC in floral zygomorphy is early discovered in A. majus, where CYC is expressed in the dorsal petal of the flower, patterning the dorsal petal identity by affecting its size and shape so that it could be distinguished from the ventral petal (Costa et al., 2005; Hileman, 2014; Spencer and Kim, 2018). The phenomenon of CYC regulation of floral zygomorphy has also been observed in S. speciosa (Dong et al.,
2018). This study showed the discovery of TFs that were responsive to CYC in S. speciosa.
SsCYC was able to regulate certain dorsal-expressed TFs whose 5’ regulatory regions were enriched with TCP binding sites, elements that have been known to mediate TCP TF family gene regulation. SsCYC regulation of these TFs could be linked to their function as SsCYC downstream in patterning the dorsal identity of S. speciosa.
The floral zygomorphy establishment in S. speciosa ‘Espirito Santo’ might involve
another RAD-like gene
It has been well-known that the floral zygomorphy regulation in A. majus relies on AmCYC and AmDICH regulation of AmRAD in dorsal petal. The fact that AmCYC is
able to bind to the TCP binding sites found at the promoter and intron of AmRAD suggests that TCP binding sites are also the important elements that provide AmCYC regulation of AmRAD (Costa et al., 2005). The classic pattern of CYC-RAD-DIV regulation is generally
thought to be conserved in Lamiales, and even has been reported outside Lamiales which is in Dispacales (Pretson and Hileman, 2009; Boyden et al., 2013). For instance, the CYC- RAD regulation is found in Bournea leiophylla (Gesneriaceae) (Zhou et al., 2008),
Veronica montana and Gratiola officinalis (Antirrhinum close relatives) (Preston et al.,
2009), since their RAD genes are expressed in the similar manner with their CYC gene counterparts. Furthermore, study in Chirita heterotricha (Gesneriaceae) signifies CYC binding site enrichment at RAD promoter outside the Antirrhinum (Yang et al., 2010), which supports the hypothesis of CYC-RAD model conservation in establishing floral zygomorphy.
However, there are also evidences that the CYC-RAD model is actually not conserved. In both Antirrhinum and Arabidopsis, it has been found that there are some RAD-like genes. Observation of the 5 RAD-like genes in Antirrhinum shows that none of
them are expressed like AmRAD in dorsal regions of the flower. The same phenomenon is also occurred in the 6 RAD-like genes of Arabidopsis, which they are not expressed at the same region with TCP1, the Arabidopsis AmCYC orthologue. Moreover, when
endogenous RAD-like genes of Arabidopsis. Together, these studies suggest that there might be changes have occurred in the cis-regulatory elements of these RAD-like genes during the duplication which raise the possibility that the control of floral zyomorphy in other species does not always follow the CYC-RAD-DIV model of Antirrhinum (Baxter et al., 2007; Costa et al., 2005). In addition, it has also been reported that in Saintpaulia ionantha (Gesneraiceae), the RAD expression does not correlate with CYC (Hsu et al.,
2018). This evidence supports the suggestion that even in Gesneriaceae, CYC might co- opt other pathways in regulating floral zygomorphy.
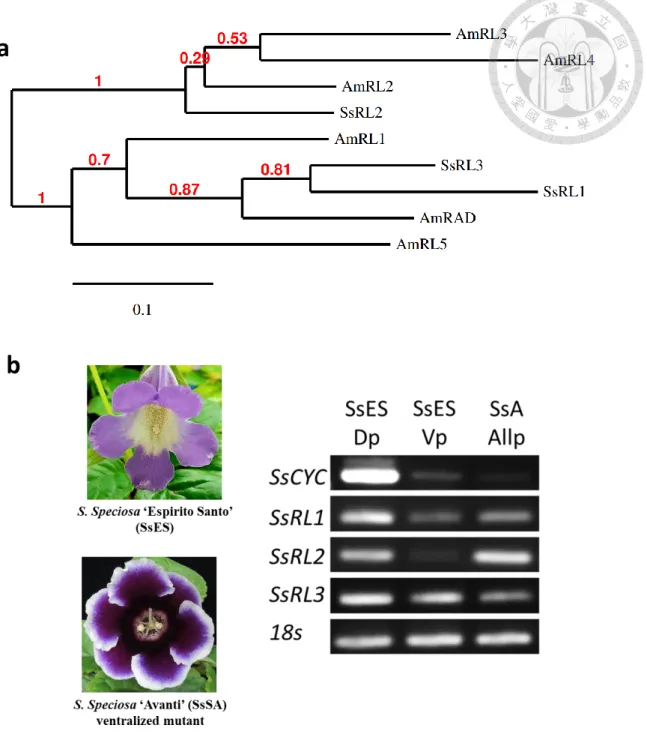
Although S. speciosa is belong to Lamiales, the RNA-seq data of S. speciosa
‘Espirito Santo’ (SsES) indeed only showed that SsCYC was differentially expressed while no SsRAD and SsDIV (Table 1), homologous of AmRAD and AmDIV were found to be dorsi-ventral differentially-expressed. Instead of RAD, other RAD-like gene (SsRL2) which has more similarity to the Antirrhinum RAD-like 2 (AmRL2) was found to be dorsi- ventral differentially expressed. In the case of SsES, the SsRL2 seemed to be activated by SsCYC since its 5’ regulatory region was enriched with the TCP binding site and showed up-regulation by SsCYC in the dual-luciferase assay (Table 4; Fig. 2). Although the function of this TF is still unknown, but it might have some influences in the floral zygomorphy regulation of SsES.
Based on this data, it was originally thought that the control of floral zygomorphy in S. speciosa might not mimic the model of A. majus, but later it was found that there were two other RAD-like genes (SsRL1 and SsRL3) in SsES that surprisingly did not pass the dorsi-ventral differential expression filter in the RNA-seq. In order to confirm the existence of CYC-RAD-DIV model in S. speciosa, the expression of these RAD-like genes and SsCYC was compared within the dorsal and ventral petals of SsES and also with the whole petals of S. speciosa ‘Avanti’ (SsA), the other cultivar that has ventralized actinomorphic symmetry caused due to 10 bp deletion of SsCYC. As expected, SsCYC exhibited high expression pattern at the dorsal petals of SsES but was expressed in low level at the ventral petals of SsES and the whole petals of SsA, supporting the previous suggestion that SsCYC is the major role of floral zygomorphy in S. speciosa. Interestingly, the SsRL1 was the only RAD-like gene that expressed almost in the similar manner with SsCYC, confirming that the establishment of floral zygomorhy in S. speciosa might
follows the model of Antirrhinum. The high expression pattern of SsRL2 in SsA indicated that there might be other regulation of this TF besides by SsCYC (Fig. 3).
Figure 3 SsCYC and SsRAD-like genes expression profile in S. speciosa ‘Espirito santo’ and S. speciosa ‘Avanti’
(a.) Phylogenetic tree of S. speciosa and Antirrhinum RAD and RAD-like genes. The tree was produced using Phylogeny.fr (http://www.phylogeny.fr/alacarte.cgi). Bootstrap values (red number) are based on 100 replicates. GenBank references: AmRL1, AJ791699;
AmRL2, DQ375230; AmRL3, DQ375227; AmRL4, DQ375228; AmRL5, AJ793240. (b.) Real-time PCR analysis to compare the expression pattern of SsCYC and SsRAD-like genes within the dorsal (Dp) and ventral (Vp) petals of S. speciosa ‘Espirito santo’ (SsES), and the whole petals (AllP) of S. speciosa ‘Avanti’ (SsA). SsA is the ventralized actinomorphic mutant of S. speciosa caused due to 10 bp deletion of SsCYC.