國立臺灣大學醫學院藥學研究所 碩士論文
Graduate Institute of Pharmaceutical Sciences College of Medicine
National Taiwan University Master Thesis
Doxorubicin 誘導 MCF-7/WT 轉變為抗藥性細胞株的過 程中基因表現變化之探討
The gene expression profile in the induction of doxorubicin-resistant MCF-7 cells
蕭惠婷 Hui-Ting Hsiao
指導教授:陳燕惠 博士 Advisor: Yen-Hui Chen, Ph.D.
中華民國 100 年 9 月
September, 2011
誌謝
記得才剛踏進藥學系的那一刻,結果轉眼間,兩年又兩個月的時間已經過去,
來到了尾聲。在這些日子裡,實驗室裡的共同生活點滴、學術上的討論、言不及 義的閒扯,現在都成為我的回憶與經驗的一部分。雖然論文不盡完美,但至少是 一個階段的註記,能順利的完成都要感謝眾多學長姊、同學學弟妹的共同砥礪,
以及老師們的教導。
首先誠摯的感謝指導教授陳燕惠老師,在我實驗上有疑問時不厭其煩地 與我討論,以及在論文撰寫過程中的重要建議,讓我能順利把論文完成。此外,
也感謝口試委員許麗卿老師及楊家榮老師,對我的論文提供了許多寶貴的意見,
使論文得以更加完善而嚴謹;感謝顧記華老師和孔繁璐老師大方的提供我實驗上 需用到的抗體,讓我能順利的完成實驗。
另外要感謝軒宇,很幸運能進入同一間實驗室,在實驗上、生活上等等 的大小事情都幫我非常多,也是我這兩年來一起努力的好夥伴!感謝尚勳學長提 供我細胞株,讓我可以順利完成實驗。感謝瑞苓學姊及佳純學姊,在我實驗卡關 時,給我許多實驗上的指導,讓我能順利解決問題。感謝文彥和可名還有思妤,
和我分享我不會的實驗步驟。感謝韋婷也常協助我解決一些實驗上的需求,以及 在口試的時候幫我記錄。感謝與我同屆的碩二同學們,大家一起努力與互相鼓勵,
讓我更有前進的動力;最後也感謝家人。
即將離開這熟悉的環境,心中充滿了不捨,期望在往後的日子也能夠認 真努力地朝自己下一步的目標邁進!
中文摘要 中文摘要 中文摘要 中文摘要
癌細胞對多種化療藥物產生抵抗力,此種現象稱為多重抗藥性,常為造成化 療失敗的原因。細胞產生抗藥性的原因一直被熱烈的探討,且也可以使用細胞毒 性藥物來篩選並建立抗藥性細胞株。藉由減少、喪失或變更藥物標靶、使細胞凋 亡的訊息傳遞受阻、增進細胞DNA受藥物損害後的修復能力、活化更多與細胞內 解毒及藥物代謝相關的酵素、增強排除藥物至細胞外的功能、減少藥物進入細胞 的量,癌細胞可對單一種藥物或者一系列藥物產生抗藥性。
MCF-7/ADR 為 對 doxorubicin 具 抗 藥 性 的 人 類 乳 癌 細 胞 株 , 相 較 於 MCF-7/WT,其過量表現P-glycoprotein減少了doxorubicin在細胞內的量並展現多重 抗藥性的樣貌。本實驗室將MCF-7/WT培養在以1 nM doxorubicin為起始濃度的培 養基中,待細胞生長穩定後,增加為兩倍藥物量,篩選對doxorubicin有抗藥性的細 胞 稱 MCF-7/ADR-n, n 為 1 nM 的 倍 數 。 目 前 分 別 建 立 了 MCF-7/ADR-1、 MCF-7/ADR-2、MCF-7/ADR-4、MCF-7/ADR-8、MCF-7/ADR-16、MCF-7/ADR-32、 MCF-7/ADR-64、MCF-7/ADR-128、MCF-7/ADR-256、MCF-7/ADR-512、MCF-7/ADR-1024
共11種對不同濃度的doxorubicin具有抗藥性的細胞株,細胞培養的終點止於 P-glycoprotein的出現為MCF-7/ADR-1024,其與MCF-7/ADR具有相同程度的抗藥性 (對doxorubicin的IC50分別為10.3 µM及12.9 µM)。
本研究利用一系列不同抗藥性程度的細胞,來探討doxorubicin對MCF-7產生抗 藥性的過程中,在ABC轉運蛋白、細胞凋亡、抗氧化及藥物代謝、DNA修復、細 胞增生等方面之相關基因表現有無變化。在ABC轉運蛋白方面,從MCF-7/ADR-32
細胞開始MRP1運輸蛋白的mRNA表現量開始漸升,在MCF-7/ADR-256 中MRP1的 mRNA 表 現 量 為 最 高 點 , 然 而 在 MCF-7/ADR-512、 MCF-7/ADR-1024中 下 降 至 與 MCF-7/WT相同程度,而P-gp的mRNA卻僅在MCF-7/ADR-1024才大量表現。 MRP1 出現及消失與P-gp的出現兩者之間是否有相關,目前尚無法證明,而MRP1運輸蛋 白mRNA的大量表現可能為細胞對doxorubicin具有較低程度抗藥性的原因。在細胞
凋亡方面,Bcl-2的mRNA在MCF-7/ADR-1024時表現量減少,GCS的mRNA表現量則 是增加;在DNA修補方面,BRCA1/2的mRNA表現量隨著對doxorubicin抗藥性程度 增加而減少,野生型p53在MCF-7/ADR-1024及MCF-7/ADR中大量減少,突變型p53 基因表現量在MCF-7/ADR-1024及MCF-7/ADR中大量增加,而總p53的基因及蛋白表 現量均呈現上升的趨勢,顯示突變可能會增加p53的半衰期,而使總p53蛋白量增 加。在抗氧化還原方面,GCL及轉錄因子Nrf2在基因表現量上並沒有明顯變化,
GSTπ 只 在 MCF-7/ADR-1024及 MCF-7/ADR 中 大 量 表 現 。 在 EMT 相 關 基 因 中 , E-cadherin 的 mRNA 表 現 量 在 MCF-7/ADR-1024及 MCF-7/ADR 中 大 量 減 少 , N-cadherin、Vimentin及可抑制E-cadherin表現的轉錄因子ZEB1、ZEB2、Twist1、
Slug 表 現 量 在 MCF-7/ADR-1024 及 MCF-7/ADR 中 增 加 , 細 胞 的 型 態 也 在 MCF-7/ADR-1024時與MCF-7/WT(類似上皮細胞)有所不同,較類似間質細胞。另外,
PKCα的基因表現量隨著對doxorubicin抗藥性程度增加而增加,ER-α的mRNA在 MCF-7/ADR-1024及MCF-7/ADR中幾乎沒有表現。
總而言之,以建立的11種不同程度的doxorubicin抗藥性細胞,分析比較細胞間 的基因表現差異,可使我們對MCF-7細胞對doxorubicin產生抗藥性的原因有更多了 解,也提供往後其他與抗藥性相關研究的基礎。
關鍵字:Doxorubicin、MCF-7、MCF-7/ADR、MDR1、MRP1、多重抗藥性
Abstract
The ability of cancer cells to become simultaneously resistant to different drugs — a trait known as multidrug resistance — remains a significant impediment to successful chemotherapy. Cellular mechanisms of drug resistance have been intensively studied, as experimental models can be easily generated by in vitro selection with cytotoxic agents.
Cancer cells in culture can become resistant to a single drug, or a class of drugs with a similar mechanism of action, by altering the drug’s cellular target or by increasing repair of drug-induced damage, frequently to DNA.
MCF-7/ADR is a doxorubicin-resistant cell line. In comparison with MCF-7/WT, MCF-7/ADR overexpresses P-gp that lowers the intracellular concentration of doxorubicin and exhibits the multidrug resistnace. Resistant cells were selected by doxorubicin at the starting concentration 1 nM. After the cells were tolerable, doubling the drug concentration was applied till the cells acquired tolerance. Followed by repeated treatments, 11 cell lines were established with incremental resistance to doxorubicin. The series of cells are so-called MCF-7/ADR-n and named as MCF-7/ADR-1、MCF-7/ADR-2、MCF-7/ADR-4、MCF-7/ADR-8、MCF-7/ADR-16、 MCF-7/ADR-32、MCF-7/ADR-64、MCF-7/ADR-128、MCF-7/ADR-256、MCF-7/ADR-512、 MCF-7/ADR-1024, respectively.
To investigate the gene expression profiles during resistance induction, the expression levels were measured for genes involving in ABC transporter protein, apoptosis, antioxidant and drug metabolism, DNA repair and cell proliferation in the series of MCF-7/ADR-n. Regarding ABC transporters, MRP1 expression increased slightly in MCF-7/ADR-32 and a peak was observed in MCF-7/ADR-256, then declined afterwards. P-gp mRNA bursted only in MCF-7/ADR-1024. The association of MRP1 with P-gp is still unclear. Regarding apoptosis genes, Bcl-2 expression decreased with
the elevated resistance to doxorubicin, but GCS mRNA expression was in the opposite way. Regarding DNA repair system, BRCA1/2 expression levels decreased with the elevation of doxorubicin resistance. Wild-type p53 mRNA importantly dropped in MCF-7/ADR-1024, while mutant p53 mRNA markedly appeared in MCF-7/ADR-1024. Levels of total p53 mRNA and protein were elevated with the increased doxorubicin resistance. Longer half life of mutated p53 may contribute to the elevation. Regarding antioxidant and drug metabolism, GCL and transcription factor Nrf2 showed slightly changes in mRNA expression, and GSTπ was overexpressed only in MCF-7/ADR-1024. Regarding EMT related genes, E-cadherin mRNA almost vanished in MCF-7/ADR-1024, but N-cadherin、Vimentin and E-cadherin transcription suppressors (i.e. ZEB1 、 ZEB2 、 Twist1 and Slug) were overexpressed in MCF-7/ADR-1024. MCF-7/ADR-1024 is mesenchymal-like and is different from MCF-7/WT. PKCα mRNA rised with the elevation of doxorubicin resistance, while ER-α mRNA dropped in MCF-7/ADR-1024.
In conclusion, a series of MCF-7 cells with incremental resistance to doxorubicin were established. Eleven cell lines were employed for measurement of gene expression to better understand gene profiles in the induction of doxorubicin resistance in MCF-7.
Key word:Doxorubicin、MCF-7、MCF-7/ADR、MDR1、MRP1、Multidrug resistance
目錄 目錄 目錄 目錄
誌謝 ... iii
中文摘要 ...i
英文摘要 ... iii
目錄 ... v
圖目錄 ... vii
表目錄 ...ix
縮寫對照表 ... x
第一章、緒論 ... 1
1.1 多重抗藥性 ... 1
1.2 ABC 轉運蛋白家族在多重抗藥性中扮演角色及功能 ... 2
1.3 其他可能使癌細胞表現抗藥性或侵入組織性之相關分子 ... 4
1.4 Doxorubicin 之作用機制與影響 ... 7
1.5 MCF-7/ADR 細胞及 MCF-7 Wild Type 細胞之差異 ... 8
1.6 利用 doxorubicin 篩選並建立具抗藥性之 MCF-7 細胞 ... 9
1.7 研究目的 ... 9
第二章、材料與研究方法 ... 11
2.1 實驗儀器 ... 11
2.2 實驗材料 ... 12
2.3 培養液、培養基與緩衝液之配製 ... 13
2.4 實驗方法 ... 15
一、 培養基之製備 ... 15
二、 細胞株培養及抗藥性細胞株之建立 ... 16
三、 逆轉錄-聚合酶連鎖反應法 (Reverse-Transcription PCR) ... 16
四、 即時定量-聚合酶連鎖反應法 (Real-Time PCR) ... 18
五、 西方點墨法 (Western blot) ... 19
六、四甲基偶氮唑鹽比色法 (MTT assay) ... 21
七、Rhodamine 123/calcein-AM 螢光染劑法 (MDR1/MRP1 functional assay)21 2.5 統計檢定 ... 22
第三章、實驗結果 ... 23
第四章、討論 ... 29
第五章、結論 ... 35
第六章、圖表 ... 36
第七章、參考文獻 ... 60
圖目錄
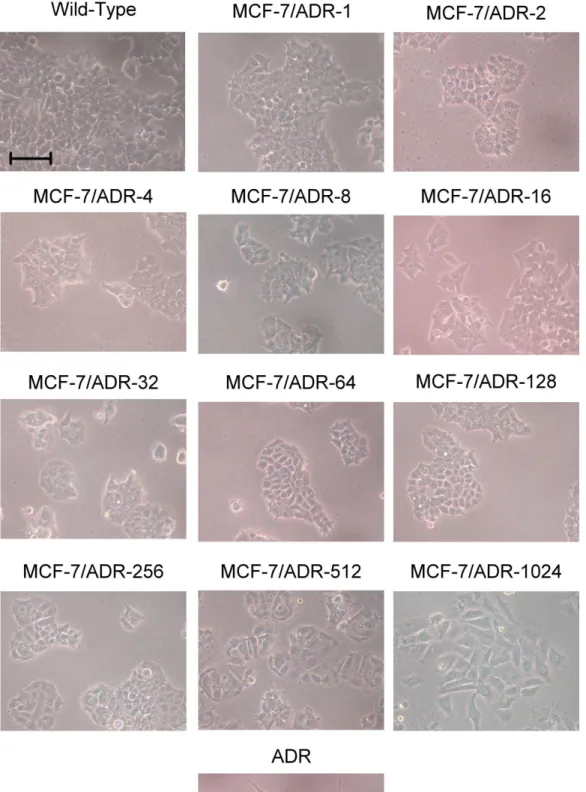
圖 1.各種 MCF-7 細胞株型態照 ... 37
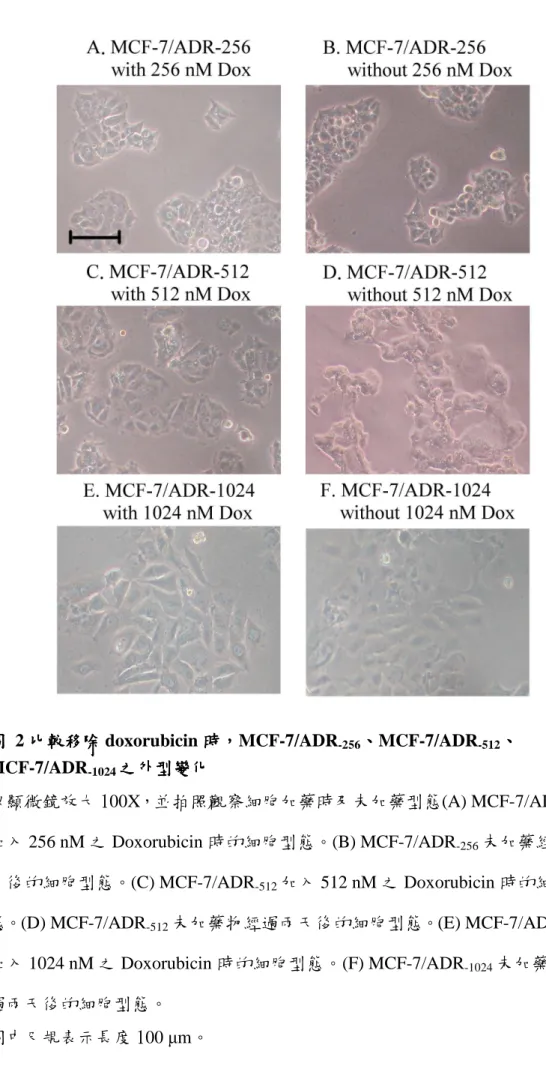
圖 2 比較移除 doxorubicin 時,MCF-7/ADR-256、MCF-7/ADR-512、MCF-7/ADR-1024 之外型變化 ... 38
圖 3 以 RT-PCR 及西方點墨法分析 MDR1 基因及蛋白表現 ... 39
圖 4 MRP1 基因表現定量及蛋白表現 (Real-time PCR) ... 40
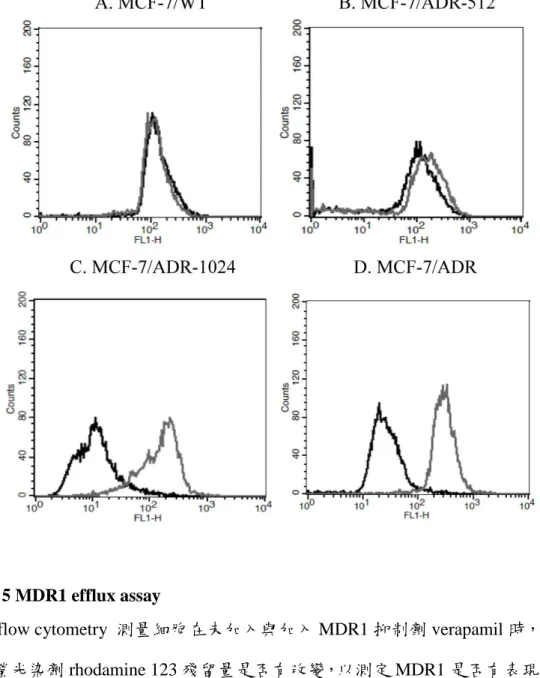
圖 5 MDR1 efflux assay ... 41
圖 6 MRP1 efflux assay ... 42
圖 7 以 RT-PCR 分析抗細胞凋亡相關基因 Bcl-2、c-FLIP、GCS 之 mRNA 在不同 抗藥性程度的 MCF-7 中之表現量 ... 43
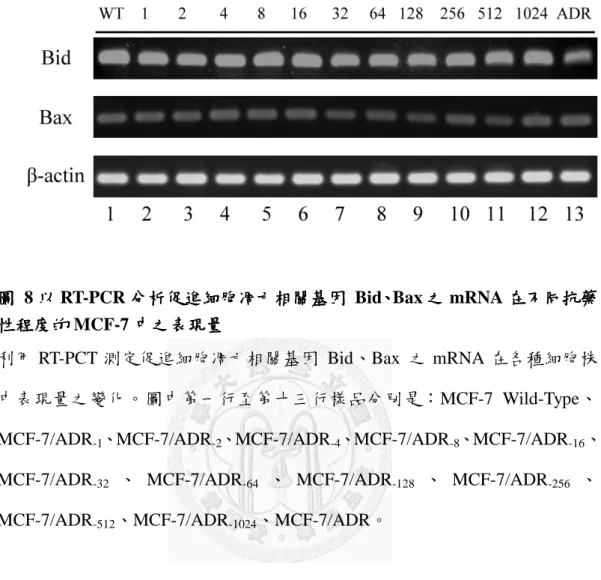
圖 8 以 RT-PCR 分析促進細胞凋亡相關基因 Bid、Bax 之 mRNA 在不同抗藥性程 度的 MCF-7 中之表現量 ... 44
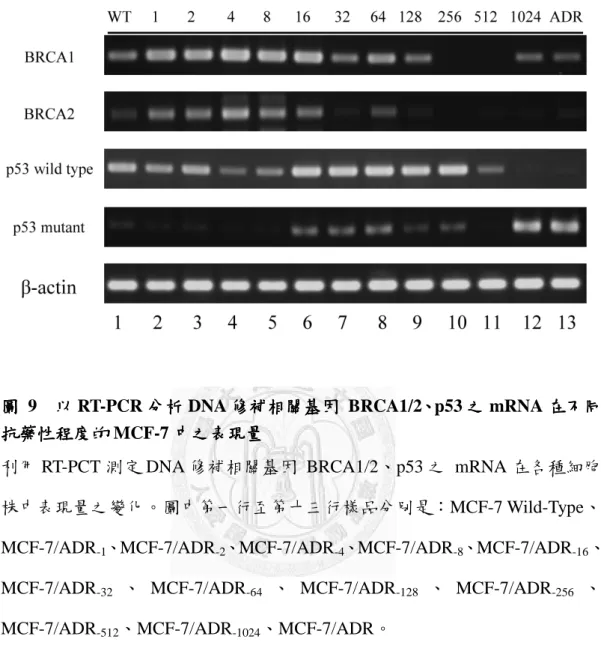
圖 9 以 RT-PCR 分析 DNA 修補相關基因 BRCA1/2、p53 之 mRNA 在不同抗藥性 程度的 MCF-7 中之表現量 ... 45
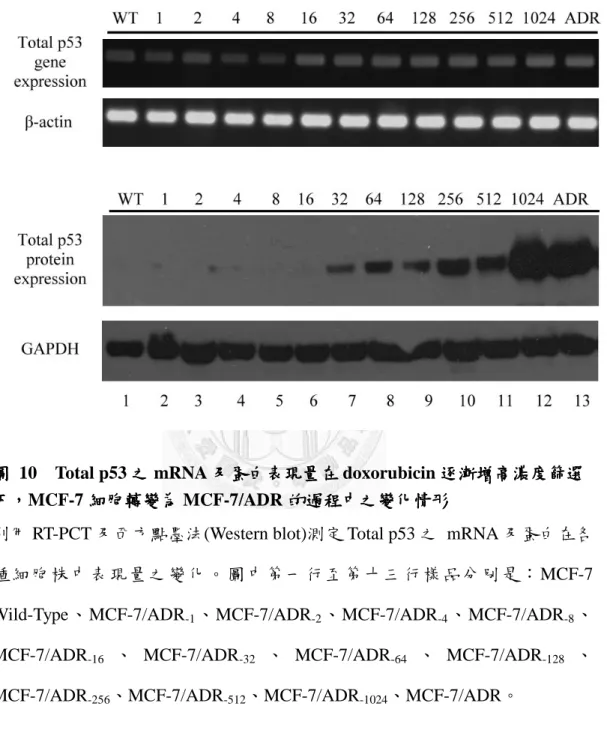
圖 10 Total p53 之 mRNA 及蛋白表現量在 doxorubicin 逐漸增高濃度篩選下,MCF-7 細胞轉變為 MCF-7/ADR 的過程中之變化情形 ... 46
圖 11 以 RT-PCR 分析 Nrf2 轉錄因子及藥物代謝、解毒、抗氧化相關基因 GST-π、 GCL 之 mRNA 在不同抗藥性程度的 MCF-7 中之表現量 ... 47
圖 12 以 RT-PCR 分析 EMT 相關基因 E-cadherin 及 ZEB1、ZEB2、Snail 等轉錄因 子之 mRNA 在不同抗藥性程度的 MCF-7 中之表現量 ... 48
圖 13 以 RT-PCR 分析 EMT 相關基因 N-cadherin、Vimentin 及 Twist1、Slug 等轉 錄因子之 mRNA 在不同抗藥性程度的 MCF-7 中之表現量 ... 49
圖 14 以 RT-PCR 分析 ABC 轉運蛋白 BCRP 之 mRNA 在不同抗藥性程度的 MCF-7 中之表現量 ... 50 圖 15 以 RT-PCR 分析促進細胞增生轉錄因子 ER-α 及蛋白激酶 PKCα 之 mRNA 在
不同抗藥性程度的 MCF-7 中之表現量 ... 51
附圖 1 癌細胞產生抗藥性之機轉圖 ... 52
附圖 2 Sphingolipids (Ceramide/GCS)引起細胞產生抗藥性的途徑 ... 53
附圖 3 p53 蛋白參與的調控 ... 54
附圖 4 上皮細胞與間質細胞藉由 EMT 機制轉換途徑圖 ... 55
附圖 5 Nrf2 藉由與 ARE 的結合活化 Phase II 解毒酵素及抗氧化酵素而造成抗藥性 之機轉圖 ... 56
附圖 6 MCF-7/ADR 中之突變 p53 在其基因 exon 5 處的 126-133 codon 有 21 bp 的 缺失 ... 57
表目錄
表 1 Doxorubicin-resistant 細胞株的 IC50值 ... 36 表 2 PCR 和 Real-time PCR 所使用之核酸引子 ... 58
縮寫對照表 縮寫對照表 縮寫對照表 縮寫對照表
英文 縮寫
Antioxidant-response element ARE
ATP-binding cassette transporter ABC transporter
B-cell lymphoma 2 Bcl-2
Breast cancer resistance protein BCRP
Cellular FLICE-inhibitory protein c-FLIP
E-cadherin CDH1
Epithelial–mesenchymal transition EMT
Estrogen receptor ER
Excision repair cross-complementing protein ERCC1
Glucosylceramide synthase GCS
Glutamate cysteine ligase GCL
Glutathione GSH
Glutathione peroxidase GPx
Glutathione S-transferases GSTs
GSH disulfide GSSG
Heme oxygenase-1 HO-1
Leukotriene C4 LTC4
MCF-7 adriamycin resistance MCF-7/ADR
MCF-7 wild type MCF-7/WT
Michigan Cancer Foundation – 7 MCF-7
Multidrug resistance MDR
Multidrug resistance-associated protein 1 MRP1
NAD(P)H quinone oxidoreductase-1 NQO1
N-cadherin CDH2
Neuroblastoma NB
NF-E2-related factor 2 Nrf-2
Peroxiredoxin Prx
P-glycoprotein P-gp
Protein kinase C α PKCα
Steroid and xenobiotic receptor SXR
Thioredoxin Trx
Thioredoxin reductase TrxR
UDP-glucuronosyltransferase UGT
X-ray cross complementing 1 XRCC1
Y-box binding protein 1 YB-1
第一章 第一章 第一章
第一章、 、 、 、緒論 緒論 緒論 緒論
1.1 多重抗藥性多重抗藥性多重抗藥性多重抗藥性
化學治療為有效治療癌細胞的方式之一,然而當癌細胞對藥物產生抗藥性時,
使化療的效果明顯受阻。當細胞對一種藥物產生抗藥性之後,可能也會對同系列 類似結構的藥物,甚至完全不相關的藥物有抗藥性,這種現象稱為多重抗藥性。
細胞產生抗藥性的原因一直被熱烈的探討之外,有些研究甚至直接以細胞毒性藥 物來篩選並建立抗藥性細胞株。培養中的癌細胞經由下列幾種可能的機制,產生 對單一種藥物或者一系列藥物的抗藥性:減少、喪失或變更藥物標靶、使細胞凋 亡的訊息傳遞受阻、增進細胞 DNA 受藥物損害後的修復能力、活化更多與細胞內 解毒及藥物代謝相關的酵素、增強排除藥物至細胞外的功能、減少藥物進入細胞 的量[1, 2](附圖 1)。
當藥物引起死亡及細胞週期的訊息受阻或產生改變時,可使細胞生長情況受影 響或者免於走向細胞凋亡路徑。例如,有些癌細胞在長期接觸化療藥物時,與細 胞週期及 DNA 損傷修復相關的 p53 蛋白會突變或失去功能,導致細胞週期的改 變;另外也有報導指出,ceramide 為一種可引起程序性細胞死亡的物質,當細胞接 觸藥物後,大量表現酵素 glucosylceramide synthase 將 ceramide 醣基化之後,可使 細胞不走向死亡的途徑,此醣基化的動作也讓細胞具有抗藥性[3]。
Cytochrome P450 3A4 是一種與藥物代謝相關的重要酵素,其基因的表現可被 steroid and xenobiotic receptor (SXR) 調節[4]。當 cytochrome P450 3A4酵素的表現 量增加,可增進藥物分子的分解,使藥物失去活性、造成抗藥性的發生。在DNA 損傷的修復方面,有研究指出在對烷化劑 (akylating agents) 具抗藥性的癌細胞 中 , 與 DNA 斷 裂 修 復 相 關 蛋 白 excision repair cross-complementing protein (ERCC1)[5] 或 X-ray cross complementing 1 (XRCC1)[6]大量表現。
而ABC轉運蛋白的表現,通常為造成藥物在細胞內殘留量減少,對藥物表現抗 藥性的主因之一。
1.2 ABC 轉運蛋白轉運蛋白轉運蛋白轉運蛋白家族在多重抗藥性中扮演角色及功能家族在多重抗藥性中扮演角色及功能家族在多重抗藥性中扮演角色及功能家族在多重抗藥性中扮演角色及功能
多重抗藥性轉運蛋白最廣為人所知的功能,就是可將抗癌藥物癌移出癌細胞 外,造成化療的失敗。在的多重抗藥性癌細胞中,通常伴隨著 ABC 轉運蛋白大量 表現[7]。
ATP-binding cassete (ABC) transporter 為一龐大穿膜蛋白家族,根據其序列及結 構分為七類,由 A 至 G 命名。ABC 轉運蛋白藉由消耗 ATP 運送多種外源物質 (xenobiotics) 包括藥物、脂質、細胞代謝物;而 ABCB1、ABCC1、ABCG2 最常 與抗藥性的引起相關[8]。
P-glycoprotein ( P-gp;MDR1;ABCB1)為 MDR1 基因轉譯而來,為一磷酸化、
醣基化的膜蛋白;分子量 170-kDa,共 1280 個胺基酸組成。P-gp 在人體許多正常 組織中都有表現,例如血腦屏障 (blood brain barrier, BBB)、小腸等處,功能為運 輸一些磷脂質及膽鹽,因此 P-gp 在正常組織中可以避免其過度吸收外源物質,而 最近也有研究指出 P-gp 也可扮演運送膽固醇的角色[9]。
P-gp 除了位於各種不同具有多重抗藥性細胞株的細胞膜上之外,也存在於高基 氏 體 膜 上 , 這 些 抗 藥 性 細 胞 株 包 括 乳 癌 細 胞 (MCF-7) 、 人 類 淋 巴 血 癌 細 胞 (CEM)、口腔上皮癌細胞 (KBC4)、骨肉瘤細胞 (U2OS)等細胞。
P-gp的受質有許多種,包含 colchicine, anthracyclines, epipodophyllotoxins, vinca alkaloids, taxanes, camptothecins, bisantrene, imatinib, mitoxantrone, saquinivir, methotrexate and actinomycin D。由於P-gp可與多種脂溶性藥物結合,因此多種P-gp 的抑制劑隨之發展。Verapamil與cyclosporin A為目前實驗室中最常用到的兩種P-gp 抑制劑;verapamil在臨床上用為鈣離子通道抑制劑 (calcium channel blocker),而 cyclosporin A為免疫抑制劑 (immunosuppressant)。然而這兩種藥物需在高濃度下才 具有逆轉MDR現象的作用 (EC50,2-10 mM),在臨床上此濃度下藥物的毒性會限 制它們的作用,因此更有效的P-gp抑制劑尚待發展[10]。
在許多研究中指出,許多種複雜的機制參與調節MDR1基因在人類癌細胞中的
表現。人類的MDR1基因啟動子 (promoter)上包含許多轉錄因子SP1、NF-Y、YB-1 的結合位,而這些轉錄因子曾經被報導過可促進MDR1基因啟動子的活性。而有研 究指出人類癌細胞在壓力誘導下,MDR1基因上的 inverted CCAAT box (Y-box) 可 被YB-1轉錄因子直接接上,促進其表現上升。另外有研究指出,NF-κB/p65及c-Fos 蛋白複合物藉由與MDR1啟動子CAAT位置處結合,對MDR1基因的啟動子活性有 負向調節作用,但此現象只在MCF-7/WT見到,在MCF-7/ADR中則無。有研究也 指出, p53蛋白的突變而失去活性,也可能誘導P-gp表現,引起多重抗藥性的產生 [11, 12]。
MRP1 (multi-drug resistance-associated protein,ABCC1)為與P-gp 相似的ABC 運輸蛋白,MRP1一開始是在由doxorubicin篩選的human small cell lung carcinoma cell line中發現,因此與此細胞多重抗藥性的造成有關。
可被MRP1運送的藥物受質大部分與P-gp重疊,包含anthracyclines, mitoxantrone, vinca alkaloids, imatinib, epipodophyllotoxins, camptothecins, colchicine, saquinivir and methotrexate[13]。但除此之外,MRP1在正常組織中也有表現,正常生理作用 下負責運輸穀胱甘肽glutathione (GSH)、穀胱甘肽衍伸(glucuronide-conjugates)重金 屬、GSH disulfide (GSSG)、白三烯素C4 (leukotriene C4,LTC4),大多為藥物代謝 過程中 phase II酵素的產物,在藥物代謝及解毒的過程扮演重要的角色,另外也可 以避免細胞在受到藥物刺激後,被誘導而產生的氧化壓力傷害[13]。
有研究指出,doxorubicin可經由c-jun N-terminal的途徑誘導人類非小細胞肺癌 中MRP1的表現[14];正常p53為腫瘤抑制基因,但當其突變時,也可能導致MRP1 上升[15];在神經母細胞瘤中(neuroblastoma, NB),MYCN oncogene可藉由接到 MRP1 啟 動 子 上 的 E-box 而 使 MRP1 表 現 上 升 [16] 。 而 近 來 轉 錄 因 子 Nrf-2 (NF-E2-related factor 2也被報導具有調節MRP1的功能,Nrf-2蛋白與抗氧化酵素的 調 節 與 表 現 息 息 相 關 [17] , 它 可 藉 由 接 至 MRP1 啟 動 子 的 ARE-element (antioxidant-response element)上而促進MRP1的表現[18]。
BCRP (breast cancer resistance protein,BCRP,ABCG2,MXR1),為近來在乳 腺癌細胞株MCF-7/AdrVp 中檢測到一種跨膜轉運蛋白,但在其他種類的癌細胞中 也有BCRP的表現,例如骨隨瘤細胞(8226 myeloma cell)、大腸癌細胞(HT29RNOV colon cancer cell)。BCRP只有655個胺基酸,大小為P-gp及MRP1的一半,因此又稱
為Half transporter。除了癌細胞外,BCRP在胎盤、血腦屏障、腸胃道等正常組織中 也有分布[19]。
BCRP的受質也包含anthracycline類抗癌藥物,以及topotecan、 mitoxantrone
或 doxorubicin,但通常表現BCRP的細胞株大多經由mitoxantrone篩選而得。BCRP 在癌細胞中過度表現也會引起多重抗藥性的現象,但有關此新的運輸蛋白質和臨 床藥物性之間的關聯仍需進一步的研究來確認。
1.3 其他可能使癌細胞表現抗藥性或侵入組織性之相關其他可能使癌細胞表現抗藥性或侵入組織性之相關其他可能使癌細胞表現抗藥性或侵入組織性之相關其他可能使癌細胞表現抗藥性或侵入組織性之相關分子分子分子 分子
PKCα (protein kinase C α)的表現和乳癌細胞的生長速率、抗藥性的產生、雌激 素受體功能的調控、細胞轉移能力和細胞凋亡都有極大相關性[20]。當 PKCα 受到 刺激表現上升時,會促進細胞生長、增加細胞的侵入力、促使 MDR 相關基因的表 現並且使雌激素受體表現下降[21]。先前有研究指出,MCF-7/ADR 在細胞核中的 PKCα 表現量與活性比 MCF-7/WT 細胞核中的高,推論 PKCα 可能與 MCF-7/ADR 表現的抗藥性特徵有相關[22]。
在細胞凋亡(apoptosis)方面,Bcl-2 (B-cell lymphoma 2)是Bcl-2 family的一員常 見的抑制 apoptosis 的分子(apoptosis suppressor),在肺癌、攝護腺癌、乳癌細胞等 都有表現。Bcl-2 family 中除了 Bcl-2 之外,還有其他抗細胞凋亡(anti-apoptotic)及 促進細胞凋亡(pro-apoptotic)的蛋白[23]。促進細胞凋亡的蛋白包含 Bax, Bak 及 Bid 等,抗細胞凋亡的蛋白有 Bcl-2, Bcl-xL 及 Bcl-W。促細胞凋亡的蛋白 Bid 可活化 Bax、Bak,使其接上粒線體外膜並改變粒線體膜的通透性,造成 cytochrome c 釋
放及 caspase 梯瀑反應(caspase cascade),引起細胞的死亡;而 Bcl-2 可與促細胞凋
亡蛋白 Bax、Bak、Bid 形成異二聚體(heterodimer),抑制細胞凋亡的發生。Bcl-2 在很多種癌細胞中都過度表現,與腫瘤的形成、抗藥性產生都有相關。雖然 Bcl-2 過度表現在某種程度上表示細胞較易存活,但在某些研究上卻指出有過度表現 Bcl-2 的腫瘤細胞有較好的預後[24]。例如乳癌細胞中,有 40%-70%的乳癌都有過
度表現 Bcl-2,有過度表現 Bcl-2 的乳癌細胞對荷爾蒙藥物治療的反應較佳[24]。
有趣的是,在一篇過去的報導發現,在 MCF-7/WT 細胞中的 Bcl-2 有過度表現,
但在 MCF-7/ADR 中 Bcl-2 的量卻少很多;而 Bcl-xL 及 Bax 的量在此兩種細胞中 的表現並沒有差異[25]。
c-FLIP (cellular FLICE-inhibitory protein)、GCS (glucosylceramide synthase)也在 細胞凋亡中扮演重要的角色。在許多腫瘤細胞中發現有大量的 c-FLIP 表現,可能 是藉由抑制 caspase-8 的活化而使這些腫瘤細胞可抗拒死亡受器(death receptor)所 引起的細胞凋亡[26]。Ceramide 為細胞內的一種脂質傳訊者(lipid messenger),可促 進 doxorubicin 引起的細胞凋亡。GCS 可對 ceramide 進行醣基化,使其變成 glucosylceramide,減少 ceramide 的量就可以減少細胞凋亡的發生[27]。GCS 在許 多具有抗藥性癌細胞(特別是對 doxorubicin 有抗藥性的癌細胞)都有過量表現,且 在乳癌、卵巢癌、大腸癌中都有其表現。也有研究指出,以 GCS 抑制劑來抑制 GCS 的表現或是功能,則可逆轉抗藥性[28],且可能也跟 P-gp 的表現有相關性 [29](附圖 2)。
在 DNA 修補方面,BRCA1 與 BRCA2 在 DNA 扮演重要的角色。BRCA1/2 在 乳癌細胞和其他組織中都有表現,與其他的腫瘤抑制基因(tumor suppressors)合稱 為 DNA 損傷之感測器(DNA damage sensors)。BRCA1/2 可進入細胞核中修補 DNA 雙股斷裂,也參與基因的轉錄[30]。當 BRCA1/2 有突變或表現量下降時,轉變為 癌細胞的風險提高許多,有篇研究指出,doxorubicin 濃度提高時,MCF-7 細胞中 的 BRCA1 之 mRNA 表現量會下降,顯示 doxorubicin 的濃度與 BRCA1 的表現可 能有相關性[31]。腫瘤抑制基因 p53 蛋白在 DNA 受損時,也可活化 DNA 修復蛋
白,另外可抑制細胞生長週期停留於 G1/S 的節律點上,以達成 DNA 損壞的辨識,
並讓 DNA 修復蛋白有更充裕時間修補 DNA 損壞部位,繼續細胞正常生長週期。
若細胞的 DNA 受損已不能修復,p53 蛋白能起始細胞凋亡程序,避免擁有不正常 遺傳資訊的細胞繼續分裂生長[32](附圖 3)。在受到壓力的細胞中,p53 蛋白的半衰 期 (half-life) 會突然的增加,造成 p53 蛋白在細胞中的累積。再來則是構型變化 (conformational change) 使得 p53 蛋白被激發成為轉錄調節因子 (transcription regulator),在前文中也提過突變的 p53 可能誘導 ABC 轉運蛋白 MDR1 的表現[12]。
有研究指出,經過化療藥物篩選過而產生抗藥性之細胞,比其親代不具抗藥性 之癌細胞更具轉移性 (metastatic)及侵入性(invasive)[33]。上皮-間葉細胞轉換現象 (epithelial–mesenchymal transition,EMT)為細胞改變其入侵、移動能力及細胞與細
胞間相互作用的重要過程。一般情況下,E-cadherin (CDH1)為不具轉移能力的上 皮細胞(epithelial cell)間黏著接連(adherens junctions)的主要成分,維持細胞間能緊 密連結成層狀,是抑制細胞移行(migration)、也是上皮細胞標記分子之一,因此 E-cadherin 的調節成為細胞是否具有侵入能力的關鍵[34]。附圖 4 中,當 EMT 發 生時,一些轉錄因子如 Snail、Slug、ZEB1/2、Twist1 會抑制 E-cadherin 的表現 (Snail、Slug、ZEB1/2、Twist 又可稱為 CDH1 的 transcription repressor),使上皮細 胞失去 E-cadherin,細胞間連結變疏鬆、無法成層、型態改變,漸成為間質細胞 (mesenchymal cell),誘導 N-cadherin (CDH2)及 vimentin 產生,此兩者為間質細胞 的標記[35]。近來有研究提到,EMT 相關基因在癌細胞中扮演的角色比想像中更 廣,例如:Slug 的表現可能與癌細胞抗藥性的產生有關聯、在乳癌細胞中 Slug 表 現會抑制 tumor suppressor BCRA2 的表現[36]、也會保護癌細胞避免走向細胞凋亡 [37]。
GSTs (glutathione S-transferases)主要在細胞中負責保護細胞免於受到毒物、外
在壓力侵害,為藥物代謝過程中參與phase II的酵素[38]。GSTs能將藥物或有機分 子接上glutathione,使其成為極性分子,再被MRP1運輸至細胞外。在抗藥性癌細
胞中,GSTs常常過度表現[12],例如MCF-7/ADR細胞中的GSTπ在mRNA、蛋白層 次 的 表 現 上 都 遠 高 於 MCF-7/WT , 儘 管 此 兩 種 細 胞 中 負 責 運 輸 GSTπ 作 用 後 glutathione鍵結產物的MRP1轉運蛋白的表現量都相當低[39]。這些藥物代謝解毒、
及抗氧化相關酵素的調控,近來被發現被Nrf2 (nuclear factor erythroid-2-related factor-2)有密切的關係[40]。Nrf-2為一種轉錄因子,平常與Keap1結合並受到Keap1 的抑制;當其受到外在氧化壓力刺激時,被誘導而脫離Keap1、進入細胞核中,與 轉錄因子Maf結合一起接到目標基因啟動子的ARE區域(antioxidant responsive element)上,進而促進目標基因的表現增加[41](附圖5)。Nrf2負責調控的基因可大 約 分 為 三 類 : (1) 細 胞 內 氧 化 還 原 平 衡 相 關 蛋 白 (intracellular redox-balancing proteins):glutamate cysteine ligase (GCL)、glutathione peroxidase (GPx)、 thioredoxin (Trx)、 thioredoxin reductase (TrxR)、 peroxiredoxin (Prx)、及 heme oxygenase-1 (HO-1),其中GCL為抗氧化物榖胱甘肽(glutathione, GSH) 的生成酵素。(2)phase II 解毒相關酵素(phase II detoxifying enzymes):glutathione S-transferase (GST),
NAD(P)H quinone oxidoreductase-1(NQO1),及UDP-glucuronosyltransferase (UGT)。
(3)運輸蛋白( transporters):multidrug resistance-associated protein (MRP)[42]。Nrf2 的作用可以使細胞增強抵抗外在壓力的能力,因此近來其在癌細胞中的影響也被 熱烈的探討。
1.4 Doxorubicin 之作用之作用之作用之作用機制機制機制機制與影響與影響與影響與影響
Doxorubicin (Adriamycin®)是一種廣泛使用及對多種癌症均有治療效果的抗癌 舊藥,屬於 anthracycline 類藥物。其主要作用機制是經由螯合 DNA、抑制 topoisomerase II 干擾 DNA 雙股的分離和螺旋的活性、以及產生氫氧自由基等作用 來毒殺細胞[7]。因此除了一般常見的化療副作用:如嘔吐、噁心、黏膜組織發炎 和脫髮外,由於心肌對氫氧自由基的傷害特別敏感,使 doxorubicin 容易對心臟產 生累積性的傷害而致命,因而限制了此藥物的多次使用。
癌 細 胞 對 doxorubicin 產 生 抗 藥 性 主 要 的 原 因 是 因 為 產 生 P-glycoprotein (MDR1),而增加藥物的排出量,另外也可能因為許多與 P-gp 無關之抗藥性機轉產 生抗藥性[43],例如活化細胞中的解毒代謝系統,大量表現 GSTπ[44]、GCL 以及 活化轉錄因子 Nrf2 的系統將藥物代謝而減少對細胞的毒性[45, 46]。p53 的突變也 在 doxorubicin 的抗藥性細胞中被發現[12],另外,GCS 的過量表現也被提過是造 成 doxorubicin 有抗藥性的原因之一[27, 28]。
1.5 MCF-7/ADR 細胞及細胞及細胞及 MCF-7 Wild Type 細胞之差異細胞及 細胞之差異細胞之差異 細胞之差異
MCF-7 全名為 Michigan Cancer Foundation – 7,1970 年從一年齡 69 歲的高加 索女性身上所分離的乳癌細胞株。MCF-7/ADR 為對 doxorubicin 類藥物具有抗藥 性的乳癌細胞株,其產生抗藥性的主要原因是因為表現 P-glycoprotein,可將癌細 胞內要物排出細胞外,因而對藥物產生抗藥性[47]。而前文在介紹可能與產生抗藥 性相關之分子的部分,也提到不少在 MCF-7 及 MCF-7/ADR 中表現量有差異的分 子,因此在此就不贅述。除了上述提過的分子之外,MCF-7 具有雌激素受體(estrogen receptor,ER-positive)[48],雌激素受體屬於核受體(nuclear receptor)超家族成員,
是 重 要 的 轉 錄 因子 ,可 被 雌 激 素 作 用形 成雙 元 體 鍵 結 至 下游 基因 啟 動 子 的 ERE(estrogen response element)上,促使細胞增生、避免走向細胞凋亡途徑,是治 療乳癌藥物的作用標靶之一[49]。例如常見的治療乳癌藥物 tamoxifen,即為一種 雌激素競爭型抑制劑(SERM),具有和雌激素競爭鍵結到 ER-α 蛋白的能力,而抑 制細胞增生[50]。另外,MCF-7 細胞也具有 E-cadherin、cytokeratin,與上皮細胞 較相似;沒有表現 P-glycoprotien,對 doxorubicin 的作用較敏感。MCF-7/ADR 表 現 Vimentin、N-cadherin,型態較類似間葉細胞;除了有表現大量的 P-gp 之外,
其不具有 Estrogen receptor (ER-negative),總 p53 蛋白的量比 MCF-7/WT 高,且在 p53 的基因第 126-133 codon exon5 的片段有 21 bp 的缺失(deletion),是突變的 p53[12](附圖 6)。Doxorubicin 引起 MCF-7/ADR 產生抗藥性的原因可能相當多,除
了 MCF-7/ADR 細胞中較多的 P-gp、GSTπ 表現極具有突變 p53 之外,還有許多可 能的機制值得繼續探討。
1.6 利用利用利用利用 doxorubicin 篩選篩選篩選並建立篩選並建立並建立具抗藥性之並建立具抗藥性之具抗藥性之 MCF-7 細胞具抗藥性之 細胞細胞細胞
在較早時期的研究中,已有人使用 doxorubicin 來篩選 MCF-7 細胞,使其對 doxorubin 產生抗藥性,成為 MCF-7/ADR 細胞,但其作法大多是直接以高濃度的 doxorubicin (1 µM)與細胞一同培養。此篩選方法在藥物剛加入的一週後,未具抗 藥性的 MCF-7 細胞有 99%會死亡,只有約少數 1 至 2 個 colony 留下;再以同樣的 高濃度 doxorubicin (1 µM)繼續對僅存的少數 MCF-7 細胞作用約三個月,並擴大繁 殖僅存的 MCF-7 細胞。作者在少數的 MCF-7 細胞數量增加後,觀察細胞型態可 發現其外型與 MCF-7/ADR 相近,且與 MCF-7/ADR 一樣有大量的 P-gp 表現[51]。
另外也有許多研究以相同方法得到抗藥性細胞[52],或者有表現 ABC 轉運蛋白的 細胞,但這些文獻只指出藥物持續刺激和篩選可得到最終的抗藥性細胞,與先前 已建立的抗藥性細胞非常相似,卻無法知道當癌細胞從無抗藥性慢慢轉變成有抗 藥性的中間的每一段不同時期的過程,有哪些基因的表現或蛋白的活性在中間發 生了變化。
1.7 研究目的研究目的研究目的研究目的
在本篇論文中,利用 doxorubicin 來篩選 MCF-7 細胞並建立對 doxorubicin 有抗 藥性的 MCF-7 細胞株。不同的是,藥物的起始濃度為 1 nM,當在此濃度下的細胞 生長穩定後,稱這株新細胞為 MCF-7/ADR sub-clone 1 (MCF-7/ADR-1),之後逐漸 以 2 之倍數增加藥物濃度 (2、4、8、16、32、64、128、256、512、1024 nM),命 名 以 此 類 推 (MCF-7/ADR-2、 MCF-7/ADR-4、 MCF-7/ADR-8、 MCF-7/ADR-16、 MCF-7/ADR-32、MCF-7/ADR-64、MCF-7/ADR-128、MCF-7/ADR-256、MCF-7/ADR-512、
MCF-7/ADR-1024),培養並保留不同程度抗藥性的細胞,最後得到共 11 種不同抗藥
性 程 度 的 MCF-7/ADR 細 胞 株 。 將 此 一 系 列 的 細 胞 以 聚 合 酶 連 鎖 反 應 法 (polymerase chain reaction,PCR)分析表現有差異之基因、以及可能參與多重抗藥 性產生的基因。另一方面,比較自己培養出來的抗藥性細胞與傳統的 MCF-7/ADR 細胞之異同。另外,從幾種可能造成多重抗藥性途徑相關的基因,去探討這些基 因由 MCF-7/WT 演變至 MCF-7/ADR 的過程中,其表現的情形,並推測除了 ABC 轉運蛋白之外,其他參與使 MCF-7/ADR 產生抗藥性的眾多可能原因。
第二章 第二章 第二章
第二章、 、 、 、 材料與研究方法 材料與研究方法 材料與研究方法 材料與研究方法
2.1 實驗儀器實驗儀器實驗儀器實驗儀器
1. 12 孔乾浴器 (Pantech Firefox Dry Bath 6100) 2. 3D 旋轉振盪器 (Sun Chion SH306)
3. 核酸定量 PCR 偵測系統 (Applied Biosystems Gene Amp) 4. 影像分析照相系統 (Pharmacia Biotech)
5. 超微量分光光度計(Thermo Scientific ND-1000) 6. pH 值測定儀 (Radiometer Copenhagen PHM-92) 7. 二氧化碳培養箱 (Sanyo MCO-17 AI)
8. 水浴鍋 (Yihder BH-230)
9. 加熱電磁攪拌器 (Corning PC-420) 10. 冷凝器 (Savant RT100)
11. 倒置顯微鏡 (Nikon Diaphot 300) 12. 桌上型離心機 (Eppendorf 5415 D)
13. 真空乾燥離心機 (Savant Speed Vac SC 110)
14. 蛋白質電泳裝置 (Bio-Rad Mini-PROTEAN Tetra Cell) 15. 蛋白質轉漬裝置 (Bio-Rad Mini Trans-Blot)
16. 落地式冷凍離心機 (Hitachi CF 150) 17. 電源供應器 (Bio-Rad PAC 3000)
18. 漩渦震盪混合器 (Scientific industries Vortex-2 Genie) 19. 懸吊式桌上型離心機 (Kubota 2010)
20. 核酸即時定量 PCR 偵測系統 (Roche Light Cycler 2.0)
21. 核酸即時定量 PCR 專用離心機 (Roche LC Carousel Centrifuge 2.0)
2.2 實驗材料實驗材料實驗材料實驗材料
1. 下列產品自美國 Sigma 公司購得
Ethium bromide (EtBr),Sodium dodecyl sulfate (SDS),Doxorubicin
, Bovine serum albumin (BSA) , Etoposide , DNase I , Tris-hydroxymethylaminomethane (Tris)
2. 下列產品自美國 Promega 公司購得
MgCl2,Go Taq DNA polymerase,MMLV(-) reverse transcriptase,RNasin
3. 下列產品自美國 Invitrogen 公司購得
Dulbecco’s Modified Eagle Medium (DMEM),Trizol,Agarose
4. 下列產品自美國 Merck 公司購得
40% Polyacrylamide gel,Sodium hydroxide,Glycerol
5. 下列產品自美國 Bio-Rad 公司購得
Ammonium persulfate (APS) , N,N,N’,N’-Tetramethylethylenediamine , Acrylamide/bisacrylamide
6. 下列產品自美國 GeneTeks 公司購得
dNTP,Oligo dT,100 bp DNA marker,Protein marker
7. 下列產品自 Millipore 公司購得 ECL
8. 下列產品自美國 Roche 公司購得 DNA Master SYBER Green I
9. PCR 和 Real-time PCR 所使用之核酸引子合成及定序委託明欣公司 (表 1)
10. 下列產品自 Santa cruz Biotechnology 購得
Mouse anti-β-actin monoclonal antibody (β-actin:sc-47778) MRP1 (C-20) : sc-7773
MDR1 (D-11):sc-55510
Donkey anti-goat IgG-HRP : sc-2020 Goat anti-mouse IgG-HRP : sc-2005 Goat anti-rabbit IgG-HRP : sc-2004
2.3 培培培培養液養液養液養液、、、、培養培養培養培養基基基基與緩衝液之配製與緩衝液之配製與緩衝液之配製與緩衝液之配製 1. 1X DMEM (1 L) ,pH=7.2
一包 Dulbecco’s Modified Eagle Medium (DMEM) 粉末 NaHCO3 3.7 g
將 pH 值調至 7.2,加入去離子水至 1L 0.22 µm 過濾膜過濾
2. 1X PBS (1L),pH=7.4 8 g NaCl
0.2 g KCl 1.44 g Na2HPO4
0.24 g KH2PO4
將 pH 值調至 7.2,加入去離子水至 1 L
3. 50 X TAE electrophoresis buffer (1L) 242 g Tris-base
57.1 mL glacial acetic acid 100 mL 0.5 M EDTA ( pH 8.0)
4. 1X Cell lysis buffer 0.4 g Tris (Base) 0.88 g NaCl
補 dH2O 至 80 ml,pH 調至 7.5 0.186 g EDTA
1 ml Triton X-100 補 dH2O 至 100 ml
5. Tris-glycine electrophoresis buffer (SDS unning buffer),pH=8.3 12.1 g Tris (base)
75 g Glycine 4 g SDS
dH2O 補至 4 L
6. Transfer buffer (1L) ,pH=8,3 3 g Tris (base)
8.8 g Glycine
0.2 g MeOH dH2O 補至 500 µL 7. Blocking buffer
5% 脫脂奶粉 (溶於 TBST buffer) 8. TBS-T wash buffer,pH=7.5 3 g Tris (base)
8.8 g NaCl 0.2 g KCl 1 ml Tween-20
先加入 800 ml dH2O,用 HCl 調 pH 至 7.4,最後再補至 1 L。
9. 4X Sample buffer
25 ml Tris˙HCl buffer (pH 6.8) 20 ml 2-Mercaptoethanol
40 ml Glycerol 8 g SDS
0.04 g Bromophenol Blue dH2O 補至 100 ml
2.4 實驗方法實驗方法實驗方法實驗方法 一
一 一
一、、、、 培養基之製備培養基之製備培養基之製備培養基之製備
本配置法採用 BRL 公司之方法:DMEM 粉末倒入 1 L 燒杯並加入滅菌之去離 子水 900 ml、及 3.7 克碳酸氫鈉,攪拌至完全溶解,調整 pH 值至 7.2,將水補至 1 L。使用 0.22 µm 過濾膜 (Millipore)過濾;分裝至 500 ml 已滅菌血清瓶中,保存於 2-8℃之冰箱中。
二 二 二
二、、、、 細胞株培養及抗藥性細胞株之建立細胞株培養及抗藥性細胞株之建立細胞株培養及抗藥性細胞株之建立細胞株培養及抗藥性細胞株之建立
實驗所用之細胞株為:MCF-7/WT、MCF-7/ADR、MCF-7/VP 及本實驗室所建 立之對 doxorubicin 有不同程度抗藥性之 MCF-7 抗藥性細胞株。MCF-7 細胞株為 人類乳癌細胞,其中 MCF-7/ADR 對 doxorubicin 具抗藥性,而 MCF-7/VP 則是對 VP-16 (etoposide)具抗藥性。本實驗室之抗藥性細胞株建立方法為:以 doxorubicin 1 nM 為起始濃度加入 MCF-7/WT 細胞株培養液中,至細胞株生長狀況對藥物穩定 之後,每次增加一倍藥物濃度,最中濃度至 1024 nM doxorubicin。細胞均以 DMEM 培 養 , DMEM 添 加 10 % 胎 牛 血 清 (fetal calf serum) 及 抗 生 素 (pn-strep-amphosolution)於 37℃、5% CO2之定溫培養箱內培養。
三 三 三
三、、、、 逆轉錄逆轉錄逆轉錄逆轉錄-聚合酶連鎖反應法聚合酶連鎖反應法聚合酶連鎖反應法聚合酶連鎖反應法 (Reverse-Transcription PCR) 1. 全全全全 RNA 之抽取之抽取之抽取之抽取
(1) HOMOGENIZATION
吸掉 medium,加入冰的 PBS 清洗,吸去 PBS
在 chemical fume hood 加入 1 ml TRIZOL reagent (1 ml TRIZOL/10 cm2,約 1 ml TRIZOL 加在 diameter 3.5cm dish 中)
(2) PHASE SEPARATION
在 15-30℃下靜置 sample 5 分鐘,將 lysate 裝入 Epp. tube (1.5 mL)中
加入 200 µl chloroform/ 1 ml TRIZOL reagent (20% chloroform)盡量用手搖至溶 液呈乳白色 (15 sec),在 15-30℃ (可置冰上) 下靜置 2-3 分鐘(溶液分成二層)
離心 12000 g,15 分鐘,2-8℃。將上層吸至新的 Epp tube 中 (RNA 在上層,
上層 60%體積為 TRIZOL,protein、chloroform 在下層) (3) RNA PRECIPITATION
加入 500 µl isopropyl alcohol
在 15-30℃下靜置 10 分鐘。離心 12000 g,10 分鐘,2-8℃,將 RNA 沉澱
(4) RNA WASH
倒掉上清液。加入 1 ml 75% ethanol 清洗 RNA pellet (vortex 混合,離心 7500 g,5 分鐘,2-8℃)
(5) REDISSOLVING THE RNA
吸去 ethanol 儘量使 RNA pellet 乾燥,加入 20 µl Rnase-free water 溶解 RNA 水浴 55-60℃ 10 分鐘,加入 DNase I 0.3 µl 混合
水浴 37℃ 15 分鐘。之後再度水浴 65℃ 8 分鐘 (inactivate DNase) 2. RNA 定量
測 OD,230:260:280 is 1:2:1 for RNA (polysaccharides: nuclear acid: protein).
3. 逆轉錄法
Reagent Volume Note
RNA 5 µL 1 µL 的 enzyme 可以轉出 5µg cDNA。
H2O (nuclease free) 30 µL RNA+H2O=35 µL
Oligo dT 1 µL 固定的體積
Total 36µL
70 °C 水浴 5 分鐘,置冰上 5 分鐘
加 10 µL RT buffer,2 µL dNTP、0.5-1 µL RNasin,最後才加 1µL RT enzyme (總 體積達 50 µL)。
在 40 °C 水浴槽中反應 1 小時,再移到 95 °C 水浴 5 分鐘,使 RT enzyme 失 活。
測 cDNA 濃度。
4. 聚合酶連鎖反應法
Reagent Volume Final conc.
Primer FR 各 1 µL (0.4µM) 0.25-2.5 µM H2O (nuclease free) 10.5 µL
cDNA 1 µL <250 ng
GoTaq®Green Master Mix, 2X
12.5 µL (2X) 1X
Total 25µL
5. 2% agarose gel
取 agarose 2 g 加 TAE buffer 100 ml 於微波爐煮 2.5 min 後倒入膠盤待冷卻凝 固,取 5 µL PCR 產物加 0.5 µL DNA loading dye 混合後,以 1X TAE buffer 進行 110 V、30 min 的電泳分析,接著將膠取出後以 Ethium bromide 染色、退染後照相。
四 四 四
四、、、、 即時定量即時定量即時定量即時定量-聚合酶連鎖反應法聚合酶連鎖反應法聚合酶連鎖反應法聚合酶連鎖反應法 (Real-Time PCR) 將下述樣品混勻後,裝入毛細管並離心,全程保持低溫。
Reagent Volume Final conc.
Primer FR 各 1 µL
H2O (nuclease free) 12 µL
cDNA 1 µL <250 ng
Master Mix, 5×
concentration
4 µL 1X
Total 19 µL
離心完後,將樣品放入機器,並設定下列條件進行反應。
Cycles ℃℃ ℃℃ Hold
Ramp rate (℃℃℃/s) ℃
Acquisition Mode
Analysis Mode Preincubation 1 95 10 min 20 None None
Amplification 45
95 10 sec 20 None
Quantification 58 10 sec 20 None
72 12 sec 20 Single
Melting Curves 1
95 0 sec 20 None
Melting curves 65 15 sec 20 None
95 0 sec 0.1 Continuous
Cooling 1 40 30 sec 20 None None
所得資料經由 LightCycler software 4.05 分析。
五 五 五
五、、、、 西方點墨法西方點墨法西方點墨法西方點墨法 (Western blot) 1. 細胞全蛋白之抽取細胞全蛋白之抽取細胞全蛋白之抽取細胞全蛋白之抽取
吸掉 medium,用 PBS 沖洗,吸掉 PBS。
以刮板將細胞刮下,裝入 eppendorf tube,離心 2000 rpm、3 分鐘、2-8℃。
去除上清液,加入 2-3 倍體積之 Cell lysis buffer,懸浮均勻。
(42 µL lysis buffer,7 µL protease inhibitor) 置冰上 20 分鐘,vortex 至少四次。
離心 12000 g、15 分鐘、2-8℃,將上清液移至新的 Epp tube 中,置於-80℃保 存。
2. 蛋白質濃度之測定蛋白質濃度之測定蛋白質濃度之測定蛋白質濃度之測定
dH2O 加入 Bio-Rad Dye Reagent (4:1)稀釋,混合均勻,稀釋後可在室溫下使用
兩星期。
以下均加入 200 µL 稀釋的 Bio-Rad Dye Reagent,反應五分鐘。
反應 5 分鐘,測 595 nm 吸光值,置於-80 ℃保存。
3. SDS-PAGE 電泳法電泳法電泳法電泳法
將 protein 與 sample buffer (1:3)放入微量離心管混勻,於沸水中煮 5 分鐘,置 冰上 5-10 分鐘。
將 15-30 µg 的蛋白樣品注入膠片空格中,先以 40 V 電壓使樣品跑過 stacking gel,之後改為 80 V,使樣品跑至 dye 離底部約 1 cm 為止,約 3-4 小時。
4. 蛋白質轉漬蛋白質轉漬蛋白質轉漬蛋白質轉漬
跑完電泳後,將膠裁成適當大小,並依照大小剪裁 NC membrane 及 filter paper。使用電壓為 400 mA,1.5-2 小時。
5. 抗原抗體交互作用分析抗原抗體交互作用分析抗原抗體交互作用分析抗原抗體交互作用分析
Blocking:5 %脫脂奶粉,溶於 TBST buffer,反應 1 小時。
Wash:3X TBST buffer,洗 10 分鐘。
加入 1 st 抗體(以 TBST 稀釋),反應 1 小時。
Wash:3X TBST buffer,洗 10 分鐘。
加入 2nd 抗體(以 TBST 稀釋),反應 1 小時。
Wash:3X TBST buffer,洗 10 分鐘。
NO. 1 2 3 4 5 6 7 8 blank
BSA 1 µg/µL (µL)
0 1 2 3 4 5 Sample
1 µL
Lysis buffer 1µL
dH2O (µL) 10 9 8 7 6 5 9 9
Final conc.
(mg/mL)
0 0.1 0.2 0.3 0.4 0.5
六 六 六
六、、、、u 四甲基偶氮唑鹽比色法四甲基偶氮唑鹽比色法四甲基偶氮唑鹽比色法 (MTT assay) 四甲基偶氮唑鹽比色法
將細胞依所需數目種入 24 孔盤中,並在隔日換培養基及加入藥物。
待藥物作用的天數完成後,將 5 mg/ml 的 MTT 50 µL 加入細胞中,作用兩個 半小時。
時間到後,將上清液抽掉,加入 DMSO 400 µL,作用 20 min 待紫色結晶溶解 後,測波長 570 nm 吸光值,及 690 nm 吸光值作為校正。
七 七 七
七、、、、Rhodamine 123/calcein-AM 螢光染劑法螢光染劑法螢光染劑法螢光染劑法 (MDR1/MRP1 functional assay) MDR1
將細胞分成兩組,一組先加入 24 小時的 MDR1 抑制劑 verapamil 20 µL,另 一組不加。
Verapamil 作用 24 小時後,兩組皆加入 rhodamine 123(MDR1 受質) 5 µM 一小 時。
以冰冷的 PBS 沖洗細胞,並用 trypsin 將細胞收集下來、離心、打散,最後裝 入 falcon 上機專用的試管中。
以 FL1 的 channel 接受 rhodamine 所發出的波長的螢光,並記錄其螢光強度,
比較有無加入 verapamil 的組別螢光強度的變化。
MRP1
將細胞分成兩組,一組先加入 MRP1 抑制劑 MK571 50 uL 作用 24 小時,另 一組不加。
MK571 作用 24 小時後,兩組皆加入 calcein-AM 0.3 µM 一小時,calcein-AM 本身並不會發出螢光,但其進入細胞後被細胞內 esterase 作用變成 calcein 才方為 MRP1 的受質,且可發出螢光被偵測。
以冰冷的 PBS 沖洗細胞,並用 trypsin 將細胞收集下來、離心、打散,最後裝
入 falcon 上機專用的試管中。
以 FL1 的 channel 接受 calcein 所發出的波長的螢光,並記錄其螢光強度,比 較有無加入 MK571 的組別螢光強度的變化[53, 54]。
2.5 統計檢定統計檢定統計檢定統計檢定
本研究的數據接假設為遵守常態分布,以 Office 2010 軟體 Excel 做變異數分 析,來檢定各組數據的差異性,並用學生氏 t 檢定(Student’s t-test)進行全組數據的 比較及兩兩組別的差異分析。以*表示 p value <0.05,認定數據間有顯著差異。圖 表上數值由平均數(mean) ±標準差(standard deviation)表示。
第三章 第三章 第三章
第三章、 、 、 、實驗結果 實驗結果 實驗結果 實驗結果
3.1 利用利用利用利用 MTT 法測定法測定法測定 doxorubicin 篩選之不同程度抗藥性細胞之法測定 篩選之不同程度抗藥性細胞之篩選之不同程度抗藥性細胞之 IC篩選之不同程度抗藥性細胞之 50值值值值
參照本實驗室之前發表的文獻,以及本篇研究測定的 MCF-7 之 IC50約為 0.1 µM,MCF-7/ADR 之 IC50約為 12.9 µM。其餘篩選出來之抗藥性細胞株,待每株細 胞皆在各自加的 doxorubicin 濃度下可穩定生長時,才以 MTT 法測定 IC50。測出來 之 IC50值分別為:MCF-7/ADR-1 (0.117µM)、MCF-7/ADR-2 (0.140 µM)、MCF-7/ADR-4
(0.145 µM)、MCF-7/ADR-8 (0.170 µM)、MCF-7/ADR-16 (0.215 µM)、MCF-7/ADR-32
(0.275 µM)、MCF-7/ADR-64 (0.366 µM)、MCF-7/ADR-128 (0.388 µM)、MCF-7/ADR-256
(0.575 µM)、MCF-7/ADR-512 (1.72 µM)、MCF-7/ADR-1024 (10.3 µM)(表 1)。
3.2 各種抗藥性細胞之型態變化各種抗藥性細胞之型態變化各種抗藥性細胞之型態變化各種抗藥性細胞之型態變化
將 各 種 不 同 MCF-7 細 胞 株 型 態 以 影 像 記 錄 下 來 , 可 發 現 幾 個 特 點 : MCF-7/Wild-Type 細胞間連結較緊密,形狀較寬短,當細胞數目眾多而擁擠時,細 胞與細胞間會產生堆疊現象,而 MCF-7/ADR-1、MCF-7/ADR-2、MCF-7/ADR-4、 MCF-7/ADR-8、MCF-7/ADR-16、MCF-7/ADR-32、MCF-7/ADR-64、MCF-7/ADR-128、 MCF-7/ADR-256 這 幾 株 細 胞 在 外 型 上 並 沒 有 與 MCF-7/WT 有 太 大 差 異 。 從
MCF-7/ADR-512 開始細胞外型開始有些微變化,細胞形狀拉長,且細胞間間隔較疏
鬆,堆疊的現象也減少。到了 MCF-7/ADR-1024細胞時,其外型與 MCF-7/ADR 相 似,形狀較長分支較多,細胞間不堆疊且在顯微鏡下觀看可發現其較透光,與培 養皿底部貼附較緊。由形狀來判斷,MCF-7/ADR-1024幾乎可說與 MCF-7/ADR 相 同,但仍需更進一步在基因表現型態的分析與比較才能確認此兩種細胞的異同(圖 1)。
3.3 移除移除移除移除 Doxorubicin 512 nM 時時時,時,,MCF-7/ADR, -512之外型之變化之外型之變化 之外型之變化之外型之變化
在培養細胞之過程中,發現只有 MCF-7/ADR-512 在加藥與未加藥的情況下會
有不同的細胞外觀。在加藥時,MCF-7/ADR-512細胞邊界較清晰,外型較健康、完 整;將藥物移除後,細胞邊界消失成雲朵狀,無法判斷外型,且此時細胞開始有 些許的死亡,生長情況變差,而其他株 MCF-7 抗藥性細胞並沒有這種情形。以 MCF-7/ADR-512前後兩株細胞 MCF-7/ADR-256及 MCF-7/ADR-1024做對照,這兩株 細胞在加藥時或不加藥的狀態下,細胞外觀與生長情況並不會有所改變(圖 2)。
3.4 以以以以 RT-PCR 與西方點墨法與西方點墨法與西方點墨法與西方點墨法分析分析分析分析 ABC transporter MDR1 mRNA 及蛋白及蛋白及蛋白之表現及蛋白之表現之表現之表現 情況
情況 情況
情況,,,,作為鑑定篩選出的抗藥性細胞株已轉變為作為鑑定篩選出的抗藥性細胞株已轉變為作為鑑定篩選出的抗藥性細胞株已轉變為作為鑑定篩選出的抗藥性細胞株已轉變為 MCF-7/ADR 之根據之根據之根據之根據
MCF-7/ADR 之主要特徵為其有表現 ABC 轉運蛋白,可將藥物排除,因而對 doxorubicin 有抗藥性。以 RT-PCR 及西方點墨法得到的結果發現,MDR1 mRNA 及 其 蛋 白 一 直 到 MCF-7/ADR-1024 細 胞 時 才 有 大 量 表 現 , 在 此 之 前 甚 至 連 MCF-7/ADR-512都沒偵測到 MDR1 mRNA 之訊號。此結果顯示 MCF-7/ADR-1024才 算是真正的 MCF-7/ADR 細胞,為 doxorubicin 篩選 MCF-7/WT 細胞之終點(圖 3)。
3.5 以以以以 Real-time PCR 及西方點墨法及西方點墨法及西方點墨法及西方點墨法分析分析分析 MDR1 之外的分析 之外的之外的之外的 ABC transporter MRP1 mRNA 及蛋白及蛋白及蛋白及蛋白之表現變化情況之表現變化情況之表現變化情況之表現變化情況
根據文獻,雖然在 MCF-7/WT 及 MCF-7/ADR 中的 MRP1 mRNA 及蛋白量都 相當少,但以 Real-time PCR 精確分析後發現在 MCF-7/ADR-32、MCF-7/ADR-64、 MCF-7/ADR-128、MCF-7/ADR-256、MCF-7/ADR-512中 MRP1 的 mRNA 表現量都有 明顯的增加,其增加的倍數(與 MCF-7/WT 相比)分別是:MCF-7/ADR-32 為 95 倍、
MCF-7/ADR-64 為 234 倍、MCF-7/ADR-128 為 777 倍、MCF-7/ADR-256 為 2132 倍、
MCF-7/ADR-512 為 140 倍。MRP1 mRNA 表現量在 MCF-7/ADR-1024細胞中又降至 僅有 MCF-7/WT 的 2 倍,而 MCF-7/ADR 也只有 MCF-7/WT 的 12 倍。特別值得注 意的是,作為正向控制組的 MCF-7/VP 細胞,在過去的文獻中指出其為對 etoposide (VP-16)具抗藥性的 MCF-7 細胞,表現大量的 MRP1 運輸蛋白,在此圖中其 MRP1
mRNA 表現量為 MCF-7/WT 的 64 倍,遠低於 MCF-7/ADR-256的 2132 倍(圖 4)。
另外,在西方點墨法的結果中可見 MCF-7/ADR-256細胞中的 MRP1 蛋白表現量最 多。
3.6 以以以以 Flow Cytometry 分析細胞株表現分析細胞株表現分析細胞株表現 MDR1 與分析細胞株表現 與與與 MRP1 兩種兩種兩種兩種 ABC 轉運蛋白是否轉運蛋白是否轉運蛋白是否轉運蛋白是否 具
具 具
具運輸功能運輸功能運輸功能運輸功能
在測定 MDR1 運輸蛋白的功能方面,使用 rhodamine123 作為螢光染劑,其為 MDR1 的受質,可經由細胞膜擴散進入細胞內,再被 MDR1 運輸出細胞外。加入 20 µL 的 MDR1 抑制劑 verapamil 可使 rhodamine 留在細胞中。因此若是有表現 MDR1 的細胞, 以流 式細胞儀 可偵測到 有 加入 verapamil 的組 別,其細 胞中 rhodamine 訊號會比其未加 verapamil 時強。結果發現在 MCF-7/WT、MCF-7/ADR-512
兩株細胞中,有無加入 verapamil 似乎都不影響 rhodamine 在細胞中的殘留量(圖 5A 及 5B);而在 MCF-7/ADR-1024及 MCF-7/ADR 中,有加 verapamil 的組別其細胞中 rhodamine 的量均比未加 verapamil 高(圖 5C 及 5D),顯示這兩株細胞有表現 MDR1,且其表現的 MDR1 有運輸功能。在測定 MRP1 運輸蛋白的功能方面,使 用 calcein-AM 作為螢光染劑,calcein-AM 進入細胞內後,被細胞中的 esterase 作 用完轉變為可發螢光之 calcein,且 calcein 為 MRP1 的受質,可被 MRP1 運輸至細 胞外,而使細胞內偵測到之螢光量降低;若加入 MRP1 的抑制劑 MK571 50 µL,
可使 calcein 留在細胞內而得到較高的螢光值。以流式細胞儀分析後,發現有無加 入 MK571 並不影響 calcein 在 MCF-7/WT 細胞中的量(圖 6A),而 MRP1 mRNA 表 現量最多及次多的 MCF-7/ADR-256 、MCF-7/ADR-128 中可見到在未加入 MK571 前,細胞內螢光值較低,加入 MK571 後,細胞內螢光值比各自未加 MK571 時變 高 了 約 3 倍 (MCF-7/ADR-128 ) 及 3.5 倍 (MCF-7/ADR-256 )( 圖 6B 及 6C) 。 MCF-7/ADR-512、MCF-7/ADR-1024 細胞中的 MRP1 表現量相當少,因此在加入 MK571 後,細胞內螢光值比各自未加 MK571 時變高了約 1.6 倍(MCF-7/ADR-512 )
及 1.7 倍(MCF-7/ADR-1024 )(圖 6D 及 6E)。作為正向控制組的 MCF-7/VP 細胞也與 MCF-7/ADR-256有相似的結果,比其自己在未加 MK571 時增高了約 5.8 倍(圖 6F)。
3.7 以以以以 RT-PCR 分析分析分析分析抗細胞凋亡相關基因抗細胞凋亡相關基因抗細胞凋亡相關基因抗細胞凋亡相關基因 Bcl-2、、、c-FLIP、、 、、、GCS 及促進及促進及促進細胞凋亡及促進細胞凋亡細胞凋亡細胞凋亡 相關基因
相關基因
相關基因相關基因 Bid、、、、Bax 之之之 mRNA 在不同抗藥性程度的之 在不同抗藥性程度的在不同抗藥性程度的 MCF-7 中之表現量在不同抗藥性程度的 中之表現量中之表現量 中之表現量
先前有研究指出在 MCF-7/ADR 中的 Bcl-2 表現量遠低於 MCF-7/WT,在本篇 結果中看到隨著對 doxorubicin 的抗藥性程度越高,Bcl-2 的 mRNA 表現量也逐漸 降低,至 MCF-7/ADR-1024細胞時表現量只有 MCF-7/WT 的 0.1 倍。c-FLIP mRNA 在不同細胞株中的表現量則是沒有變化;GCS 經常被指出在抗藥性細胞中有過量 表現,可避免細胞凋亡的產生,在圖 7 中可看到在 MCF-7/ADR-16之後的細胞 GCS mRNA 表現量有增高的現象,且 MCF-7/ADR 中的 GCS mRNA 表現量為 MCF-7/W T的約 1.4 倍。
而在促細胞凋亡相關基因 Bid 及 Bax mRNA 的表現量,在細胞對 doxorubicin 產生抗藥性越來越高的過程中,與 MCF-7/WT 相比並沒有太大的改變(圖 8)。
3.8 以以以以 RT-PCR 分析分析分析分析 DNA 修補修補修補修補相關基因相關基因相關基因 BRCA1/2、相關基因 、、p53 之、 之之 mRNA,之 ,,及以西方點,及以西方點及以西方點及以西方點 墨法分析
墨法分析
墨法分析墨法分析 Total p53 蛋白在不同抗藥性程度的蛋白在不同抗藥性程度的蛋白在不同抗藥性程度的蛋白在不同抗藥性程度的 MCF-7 中之表現量中之表現量中之表現量中之表現量
BRCA1 及 BRCA2 最早在乳腺細胞發現,負責 DNA 雙股斷裂的修補,為腫瘤 抑制基因,其突變或是表現量降低極有可能導致乳癌的發生。也有研究提出當癌 細胞接受化療藥物後,會減少 BRCA1/2 的 mRNA 和蛋白的表現量。由圖 9 可發現 隨著 MCF-7 細胞對 doxorubicin 的抗藥性程度越高,BRCA1/2 的表現有降低的趨 勢,在 MCF-7/ADR-1024及 MCF-7/ADR 中的 BRCA1/2 幾乎沒有 mRNA 的表現。
另外,同樣為腫瘤抑制基因的 p53 蛋白擁有多種功能,其中一項也是可調控 DNA 損傷的修補,但有研究指出在 MCF-7/ADR 中的總 p53 表現量比 MCF-7/WT 高,且在 ADR 細胞中的 p53 有 21 缺失的突變(codon 126-133 at exon 5)。在圖 9 可
看到在隨著 MCF-7 細胞對 doxorubicin 的抗藥性程度變高的過程中,野生型 p53 的 mRNA 表現量在 MCF-7/ADR-1024及 MCF-7/ADR 中大量減少,且具 21 bp deletion 的突變型 p53 在 MCF-7/WT 中原本相當少量,在 MCF-7/ADR-16至 MCF-7/ADR-256
的時期表現有些許上升,至 MCF-7/ADR-1024及 MCF-7/ADR 的表現則是大幅上升。
另外則分析細胞中總 p53 的 mRNA 及蛋白表現量變化,在基因表現量方面,
總 p53 的量隨著 MCF-7 細胞對 doxorubicin 的抗藥性程度變強而增高,而蛋白質的 表現亦是如此,且表現量增多的情況比基因的變化更加明顯(圖 10)。
3.9 以以以以 RT-PCR 分析分析分析分析 Nrf2 轉錄因子及藥物代謝轉錄因子及藥物代謝轉錄因子及藥物代謝轉錄因子及藥物代謝、、、解毒、解毒解毒解毒、、、、抗氧化抗氧化抗氧化抗氧化相關基因相關基因相關基因相關基因 GST-π、、、、 GCL 之之之之 mRNA 在不同抗藥性程度的在不同抗藥性程度的在不同抗藥性程度的在不同抗藥性程度的 MCF-7 中之表現量中之表現量中之表現量 中之表現量
轉錄因子 Nrf2 可參與許多與抗氧化、解毒代謝相關酵素的表現之調控,調控 方式是藉由進入細胞核內與被調控酵素基因啟動子上的 ARE 結合,誘導基因的轉 錄使其表現量上升。GST-π、GCL 及 MRP1 都被報導過具有 ARE 片段,可被 Nrf2 調控。但在圖 11 中可看到,隨著 MCF-7 細胞對 doxorubicin 的抗藥性程度越高,
Nrf2 基因表現量並沒有增加的趨勢,GCL 的 mRNA 表現量也沒有太大改變。GST-π 則是在 MCF-7/ADR-1024始出現大量的 mRNA 表現,為 MCF-7/WT 的約 101 倍,
MCF-7/ADR 的 GST-π mRNA 表現量為 MCF-7/WT 的 111 倍。
3.10 以以以以 RT-PCR 分析分析分析分析 EMT 相關基因相關基因相關基因相關基因 E-cadherin、、、、N-cadherin、、、、Vimentin 及及及及 ZEB1、、、、ZEB2、、、、Snail、、、、Twist1、、、、Slug 等轉錄因子之等轉錄因子之等轉錄因子之等轉錄因子之 mRNA 在不同抗藥性程度的在不同抗藥性程度的在不同抗藥性程度的在不同抗藥性程度的 MCF-7 中之表現量中之表現量中之表現量中之表現量
從圖 12 可見上皮細胞標記 E-cadherin 在 MCF-7/ADR-1024細胞中才大量減少至 MCF/WT 的 0.1 倍,在 MCF-7/ADR-1024之前的細胞都還有表現與 MCF-7/WT 約相 等的 E-cadhrin RNA。N-cadherin、vimentin 也是在 MCF-7/ADR-1024細胞中才大量 表現,MCF-7/ADR-1024之前的細胞雖然有些有 N-cadherin、Vimentin 的基因表現但
量並不多。轉錄因子 Snail 的 mRNA 表現量在隨著 MCF-7 細胞對 doxorubicin 的抗 藥性程度越高的過程中並沒有太大的改變,但轉錄因子 ZEB1、ZEB2、Twist1、Slug 等 轉 錄 因 子 的 mRNA 在 MCF-7/ADR-1024 之 前 表 現 量 都 相 當 少 , 直 到 MCF-7/ADR-1024才有上升的趨勢(圖 13)。
3.11 以以以以 RT-PCR 分析分析分析分析 ABC 轉運蛋白轉運蛋白轉運蛋白轉運蛋白 BCRP 之之之之 mRNA 在不同抗藥性程度的在不同抗藥性程度的在不同抗藥性程度的在不同抗藥性程度的 MCF-7 中之表現量中之表現量中之表現量中之表現量
雖然 BCRP 也常為造成癌細胞具有抗藥性的原因之一,但在本篇實驗中發現 BRCP 的 mRNA 表現量在隨著 MCF-7 細胞對 doxorubicin 的抗藥性程度越高的過 程中並沒有太大的改變(圖 14)。
3.12 以以以以 RT-PCR 分析促進細胞增生轉錄因子分析促進細胞增生轉錄因子分析促進細胞增生轉錄因子分析促進細胞增生轉錄因子 ER-α 及蛋白激酶及蛋白激酶及蛋白激酶 PKCα 之及蛋白激酶 之之之 mRNA 在在在在 不同抗藥性程度的
不同抗藥性程度的 不同抗藥性程度的
不同抗藥性程度的 MCF-7 中之表現量中之表現量中之表現量 中之表現量
先前文獻指出 MCF-7/WT 為 ER-α positive 細胞,而 MCF-7/ADR 為 ER-α negative 細胞,但 ER-α 的 mRNA 表現量在隨著 MCF-7 細胞對 doxorubicin 的抗藥 性程度越高的過程中直到 MCF-7/ADR-1024才大量減少至幾乎不表現,與 ADR 細胞 一樣變成 ER-α negative (圖 15)。PKCα 之 mRNA 及蛋白被報導過在有抗藥性的細 胞中會有較高的表現,實驗中,PKCα mRNA 表現量從 MCF-7/ADR-16開始有逐漸 上升的趨勢,到了 MCF-7/ADR-1024及 MCF-7/ADR 時分別為 MCF-7/WT 的 1.4 及 1.5 倍左右。
第四章 第四章 第四章
第四章、 、 、 、討論 討論 討論 討論
4.1 以緩慢漸進以緩慢漸進以緩慢漸進增高以緩慢漸進增高增高 doxorubicin 濃度的方法篩選增高 濃度的方法篩選濃度的方法篩選濃度的方法篩選 MCF-7/WT 而得到而得到而得到 MCF-7/ADR而得到 的過程
的過程 的過程
的過程,,,,細胞型態會產生變化細胞型態會產生變化細胞型態會產生變化細胞型態會產生變化,,,,可能與可能與可能與可能與 EMT 相關基因的相關基因的相關基因的相關基因的表現有關表現有關表現有關 表現有關
在前文提過,較早的研究直接以高濃度(1 µM) doxorubicin 持續刺激 MCF-7 細 胞,在強大的藥物壓力下篩選出表現 MDR1 且對 doxorubicin 有抗藥性的細胞[51],
較近的研究則有人以較低的 doxorubicin 濃度(低於 MCF-7/WT 的 IC50)持續刺激 MCF-7 細胞,得到較低程度抗藥性且沒有表現 MDR1 的細胞[55]。也有研究先以 低濃度( 10 nM) doxorubicin 作為刺激 MCF-7/WT 的起始濃度,待其生長穩定後再 增加藥物濃度,耗時約 31 個月,直到其篩選的細胞也表現 MDR1 為止[52]。但這 幾篇研究通常只分析最終表現 MDR1 的抗藥性細胞,或者有些只分析了在未表現 MDR1 前的較低程度抗 doxorubicin 細胞。因此本實驗室以 doxorubicin 篩選了一系 列 MCF-7 抗藥性細胞,將不同程度抗藥性由低至高的 MCF-7/ADR 細胞分別列出 來探討,分析當 MCF-7/WT 被 doxrubicin 刺激轉變為 MCF-7/ADR 的過程中,有 哪些基因的表現量產生變化,並推論除了 MDR1 的表現之外,尚有哪些基因可能 也參與多重抗藥性的發展。
一開始以 1 nM 之 doxorubicin 為起始濃度篩選 MCF-7/WT 細胞,每當細胞生 長穩定即增加兩倍濃度,得到了 MCF-7/ADR-1、MCF-7/ADR-2、MCF-7/ADR-4、 MCF-7/ADR-8、MCF-7/ADR-16、MCF-7/ADR-32、MCF-7/ADR-64、MCF-7/ADR-128、 MCF-7/ADR-256、MCF-7/ADR-512、MCF-7/ADR-1024共 11 種不同程度之 doxorubicin 抗藥性細胞。首先測定每種細胞之 IC50 值,以作為日後判別每一株細胞的依據。
在前面幾株低程度抗藥性細胞 MCF-7/ADR-1、MCF-7/ADR-2、MCF-7/ADR-4、 MCF-7/ADR-8 中之 IC50 並沒有與 MCF-7/WT 有太大變化,顯示這幾個濃度的 doxorubicin 對 MCF-7 細 胞 的 抗 藥 性 程 度 並 沒 有 太 大 的 影 響 及 篩 選 作 用 , MCF-7/ADR-32開始至 MCF-7/ADR-256細胞的 IC50比 MCF-7/WT 上升了約 3-6 倍,
這個時期的細胞在加藥情況下達到穩定生長的所需時間變長,大約需要兩個月才