國立宜蘭大學園藝學系 碩士論文
Department of Horticulture National Ilan University
Master Thesis
孤挺花種球之刻溝雙鱗片切割繁殖法 對子球數量及生長速度之影響
Effect of notching twin-scaling propagation method of Hippeastrum hybridum on numbers and growing speed of
bulblets.
指導教授:朱 玉 博士 Yu Chu Ph.D
研究生:黃 延 萍 Yen-Ping Huang
中華民國九十八年七月
July,2009
誌謝
本論文得以順利完成,首先要感謝恩師 朱玉教授,感謝老師這三年來 對我的悉心指導與鼓勵,及劉孔生老師撥冗惠予指導,在實驗上提供許多 寶貴的意見和協助,以及在論文指導上悉心斧正,特誌卷首謹誌最深之謝 忱!。
進入園藝研究所這個以前從未探索的領域,想當初只是單純喜歡園藝 就踏入這個領域,而在這快一千多個日子裡,也經歷了開心、難過、掙扎 及決擇,在研究生活的適應上及做實驗過程中遇到瓶頸時,老師總是不吝 給予指導與信心。第二年休學時,在工作及生活上經歷最低潮難過時,曾 經一度想放棄了,然而老師一再的鼓勵及對我的寬容和關懷,使我沒有放 棄而能繼續完成,實讓我銘感於心,朱玉老師再次的感謝您!。而這一路走 來,覺得自己很幸運,承蒙師長、研究室學長信彥、學弟妹家榮、慧芳、
貿鴻、海天、冠宇及世俊在實驗過程中的大力幫忙,我真心的感謝你們在 這一路上的協助幫忙,沒有你們,我是絕對無法完成我的論文實驗。
在進修期間感謝光復國小同事間的幫忙與協助,讓我可以無後顧之憂 的前往進修,每當在學校工作和學業之間忙得不可開交時,你們總是給我 信心和鼓勵,感謝佳珮、宜臻、秋燕、美雲及惠珍姐妹常為我加油打氣。
最後,感謝我的家人媽媽、姐姐、弟弟及天上的父親,在我求學成長 的歷程中,一直以來都很尊重我的想法和選擇,讓我自由地去做我自己想 做的事。研究所是人生另一個里程碑,感謝這三年帶給我的寶貴經驗與收 穫,感謝出現在我生命中的每一個人!謝謝你們!
目 錄(contents)
頁次中文摘要(Summary) ---1
英文摘要(Summary) ---2
壹、前言(Introduction) ---4
貳、前人研究(Literature review ) ---6
一、 植物學分類及性狀 ---6
二、 育種歷史及目標 ---7
三、 鱗莖之形態 ---8
四、生長發育和開花---10
五、種球繁殖之方法---11
(一)、種子實生法 ---11
(二)、自然分球法 ---12
(三)、鱗片繁殖法 ---13
1.基底刻溝法 ---13
2.瓣狀鱗片法---14
3.雙鱗片繁殖法 ---14
(四)、微體繁殖法---15
六、影響子球數量及生長速度之因子---15
(一)、生理因子 ---15
1.品種特性 ---15
2.繁殖體之大小 ---16
3.鱗片位置及其成熟度 ---17
(二)、環境因子 ---18
II
1.繁殖時期 ---18
2.溫度 ---19
3.介質濕度 ---19
4.光線 ---20
5.消毒處理 ---20
七、 孤挺花育種及產業發展概況 ---21
(一)、全球 ---21
(二)、台灣 ---22
参、材料與方法(Materials and methods) ---23
一、 植物材料 ---23
二、 其它材料---24
三、 試驗環境---24
四、 試驗設計與統計分析---24
五、 試驗調查項目---25
六、 試驗方法 ---25
(一)、不同切割繁殖法之比較 ---25
(二)、刻溝雙鱗片繁殖法之試驗 ---26
1.品種間之比較 ---26
2.種球大小之比較 ---27
3.不同切割時期之比較---27
4.靜置不同天數之比較 ---27
5.鱗片切割不同份數之比較 ---28
(三)、刻溝雙鱗片產業大量生產之應用 ---29
1.種球不同大小之比較 ---29
2.刻溝、瓣狀及刻溝雙鱗片不同繁殖法之比較---29
III
肆、結果(Results) --- ---30
一、不同切割繁殖法之比較 ---30
二、刻溝雙鱗片繁殖法之比較 ---31
(一) 種球不同大小之比較 ---31
(二) 不同切割時期之比較---32
(三) 靜置不同天數之比較---33
(四) 鱗片切割不同份數之比較 ---33
三、刻溝雙鱗片產業應用之結果 ---34
伍、討論(Discussion) ---36
一、刻溝雙鱗片繁殖法之探討---36
二、種球大小、品種差異對子球數量及生長速度之影響---37
三、切割時期、栽培環境對子球數量及生長速度之影響---39
陸、結論(Conclusion)------41
柒、參考文獻(Reference) ------67
捌、附錄(Appendix) ------72
圖目次
圖1.孤挺花種子---42
圖2.子球自然分球之位置---42
圖3.孤挺花種球8等份刻溝法 ---43
圖4.孤挺花種球8等份瓣狀法---43
圖5.孤挺花種球雙鱗片繁殖法 ---44
圖 6 . 本 試 驗 所 使 用 之 孤 挺 花 品 種 ( A ) 紅 獅 子 ( B ) 蘋 果 花 ( C ) 珍 女 士 (D)雙記錄(E)花孔雀 ---45
圖 7.鱗片不同切割繁殖方式 (A)頂切 (B)刻溝 (C)刻溝穿透(D)刻溝雙鱗片 ---46
圖 8.刻溝雙鱗片繁殖法生長之情形(A)種球切割後靜置二天(B)蛭石介質培育 (C)四個月子球生長情形(D)小植株---47
圖 9.刻溝雙鱗片法切割成不同份數(A)16 等份(B)8 等份(C)8+1/2 等份---48
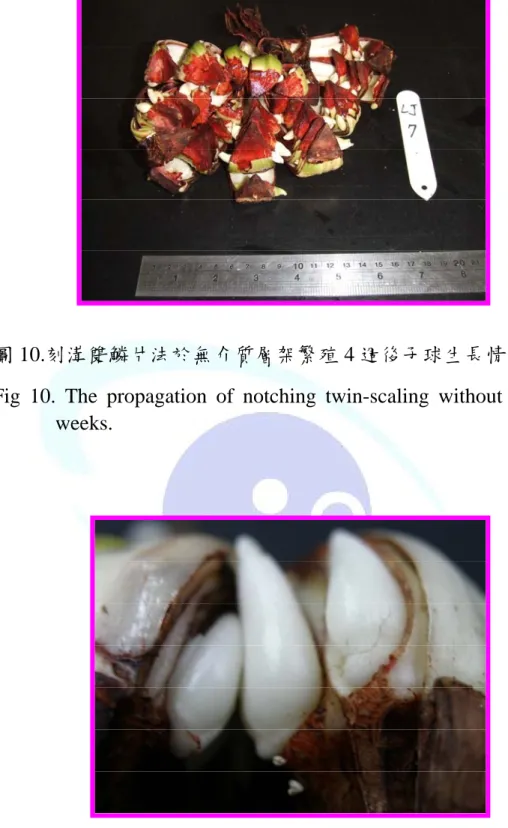
圖 10.刻溝雙鱗片法於無介質層架繁殖 4 週後子球生長情形 ---49
圖 11.刻溝雙鱗片繁殖法小鱗莖生長位置 ---49
圖 12.刻溝雙鱗片無介質放置於層架上繁殖之情形 ---50
圖 13. ‘Lady Jane’刻溝雙鱗片法繁殖四個月後子球生長情形---51
圖 14. ‘Apple Blossom’ 刻溝雙鱗片切割不同份數三個月後子球生長情形 (A)8 等份 (B) 8+1/2 等份 (C) 16 等份 ---52
圖 15. ‘Blossom Pecacok’ 刻溝雙鱗片切割不同份數三個月後子球生長情形 (A)8 等份 (B) 8+1/2 等份 (C) 16 等份 ---53
V
圖 16. ‘A210’鱗片繁殖四個月之情形 (A)瓣狀 (B)刻溝 (C)刻溝雙鱗片 ---54 圖 1 7 . ‘ A 2 1 0 ’ 種 球 利 用 刻 溝 雙 鱗 片 繁 殖 四 個 月 後 子 球 生 長 情 形 (A)大球(B)中球(C)小球---55 圖 18.不同品種利用不同切割繁殖法對子球數之影響---56 圖 19.‘Apple Blossom’種球不同大小利用刻溝雙鱗片對子球數之影響 ---57 圖 20.‘Lady Jane’利用刻溝雙鱗片繁殖於不同月份對子球數量之影響 ---58
表目次
表1. 不同切割繁殖法對子球數量之影響 ---59
表2.不同切割繁殖法對子球生長之影響---60
表3 'Apple Blossom'種球不同大小利用刻溝雙鱗片繁殖對子球數 之影響---61
表4. 'Apple Blossom'種球不同大小利用刻溝雙鱗片繁殖對子球生長 之影響---61
表 5. Lady Jane’利用刻溝雙鱗片繁殖於不同月份對子球生長之影響---62
表6. ‘Apple Blossom’ 和‘Blossom Pecock’利用刻溝雙鱗片切割不同份數 對子球數量及生長之影響---63
表 7. ‘Lady Jane’刻溝雙鱗片切割後放置不同天數對子球生長之影響---64
表 8. ‘A210’種球不同大小利用刻溝雙鱗片繁殖對子球數之影響---65
表 9.‘A210’不同種球大小利用刻溝雙鱗片繁殖對子球生長之影響---65
表 10. ‘A210’利用不同切割繁殖法對子球數之影響---66
表11. ‘A210’利用不同切割繁殖法對子球生長之影響---66
摘要
本研究藉探討孤挺花(Hippeastrum hybridum Hort.)之種球不同切割 繁殖法之改進,期增進子球數量及生長速度並尋求簡便操作之技術,以 達成種球生產最佳之經濟利益。切割方法分別為刻溝、刻溝雙鱗片、刻 溝穿透法及頂切四種方法。試驗結果顯示以刻溝雙鱗片繁殖法形成子球 數最多,刻溝和刻溝穿透法次之且無顯著差異,頂切法形成數最少。品 種間之比較,以‘Lady Jane’和‘Apple Blossom’形成率最高,‘Red Lion’次 之,`Double Record’最低。
本研究開發之刻溝雙鱗片繁殖法,為刻溝繁殖法之改良方式,先以 米字形將種球從底部切割成八等份,切割深度約為種球三分之二,再將 每等份鱗片以雙鱗片方式切割但與種球不分開,靜置二天後放置於保濕 的蛭石中培育四個月。不同季節之子球繁殖率比較,以一月及七月子球 繁殖率最高且無顯著差異,十月次之,四月最低。種球周徑愈大,繁殖 倍率愈高,周徑 20 公分之種球,即可達生產子球之標準。種球最適宜切 割份數為 8 等份。與雙麟片繁殖法相比較下,刻溝雙鱗片繁殖法其小球 初期生長較快速,所繁殖之子球較大,將可縮短子球生長至開花球的期 間。另外改良鱗片培育方式,培育初期採用無介質於層架上繁殖,待一 個月長出子球後再移至蛭石介質中栽培,此法可避免鱗片腐爛因而提高 子球形成率並可加速子球之生長速度,在操作管理上更為節省空間。
刻溝雙鱗片繁殖法具有節省空間、繁殖子球數量多及子球初期生長 快速之優點,可應用於簡易、大量及快速的孤挺花種球繁殖上,實為孤 挺花短期快速增殖之有效方法,希望能將此技術大量應用於種球生產,
以供產業界推廣之參考。
關鍵字:孤挺花、刻溝、刻溝穿透、刻溝雙鱗片、頂切、子球數量、子球生長
Summary
This study was for the purpose of investigating various cutting methods and their improvements in Hippeastrum, and thereby to increase the propagation rate and growth speed of the bulblets, thereafter by means of developing a simple manipulation technology to achieve the best economic benefit in bulblet production. The cutting methods underwent investigation including notching, notching twin-scaling, notching coring and top notching.
The results showed that notching twin-scaling was the highest in number of bulblet formation, then notching and notching coring came next and both had no significant difference, and top-notching was the lowest. In comparison among cultivars ,`Lady Jane'and`Apple Blossom'was the highest in rate of bulblet formation, then`Red Lion'came next, and`Double Record'was the lowest.
The notching twin-scaling developed in this study is the improved method of notching technique. The bulbs were first cut into eight parts from the base evenly. The cutting depth was approximately 2/3 of the bulbs length but each equal part was not separated from the mother bulb. After two days curing, bulbs were placed into the moisturized vermiculite for 4 month incubation. In comparisons for the bulblet reproduction percentage in different seasons showed January and July were the highest and had no significant difference,
October came next, and April was the lowest. The bigger the bulb circumference size was the higher the bulblet reproduction percentage obtained.
The bulb with a circumference size above 20 centimeters can be the standard size for reproduction. The most suitable bulb cutting number is 8 equal parts.
Compare with the twin-scaling, the notching twin-scaling technique had faster initial growth of bulblets and the bulblets were larger. Moreover it could shorten the growth in period from bulblet to flowering bulb. The result from the improvement on incubation method showed that initial period bulbs could put on non-medium incubation shelf, then place into moisturized vermiculite after one month later when bulblets reproduced. This method can avoid the scales rottenness , thus raising the bulblet formation rate and growth speed.
Furthermore can save the manipulation management space.
In conclusion, the notching twin-scaling propagation technique effectively save nursery space, obtain higher bulblet formation percentage and shorten the initial growth period of the bulblets, therefore it can be applied to a simple massive and fast bulblets production program, and can also performed as a reference for commercial propagation of Hippeastrum bulbs.
Key words:Hippeastrum ,notching , notching coring ,notching twin-scaling , top notching ,bulblet numbers ,bulblet growth
壹、前言(Indroduction)
孤挺花(Hippeastrum hybridum Hort.)為原產於熱帶中南美州之球根 花卉。園藝栽培品種花色與花型豔麗多采多姿,在歐、美、日為甚受歡 迎的球根花卉,用於盆花、切花及花壇植物,在台灣則多栽植於庭園或 盆花觀賞。台灣孤挺花栽培始於 1911 年,由日本人鈴木三郎氏自新台坡 引入,為花朵較小、花色少的劍瓣種,因生長容易、自然分球多而遍佈 全省,但並沒有形成商業性的消費(劉,1996)。近年來隨著新品種陸續由 國外培育並引進栽培,由於花色、花型變化豐富,加上孤挺花對環境的 適應力強,在臺灣已受各界愛花人士的注意,消費量有逐年增加的趨勢,
為具高度發展潛力的熱帶球根花卉(張,1998;林,2000)。
國內目前商業生產用孤挺花種球大多由國外進口,主要是以盆花生 產為主,而孤挺花在國內的種球生產仍處於起步階段,除了少數業者零 星栽培外,尚有各農業學校、種苗場及其他農業研究單位的培育、研究 及推廣。孤挺花主要繁殖方式有種子實生法、自然分球法、瓣狀鱗片繁 殖法及雙鱗片繁殖法。商業生產的種球繁殖主要是以雙鱗片繁殖法為主 (王,1992;Okubo,1993),若以雙鱗片繁殖商業生產的種球,子球增殖倍 率高,單位種球可產生較多子球數,但子球生長速度較慢,切割四個月 後子球周徑為 2~3cm(張,王,1998),生長至開花球需要三年期間;若將 種球平均切成 8-12 塊的瓣狀鱗片繁殖,與雙鱗片相較下,子球增殖倍率 較低,但可得較大周徑小球,子球生長一年半可得開花球。但雙鱗片繁
殖法在台灣粗放管理條件下,雙鱗片極易腐爛,增殖倍率大受影響,故 目前台灣孤挺花的繁殖主要是以瓣狀鱗片法為主,此方法與雙鱗片相比 較下其管理較容易、操作也較簡便。
球根花卉鱗片繁殖法之子球形成率及品質,常受植物種類、切割方 式及環境因子的影響,如何能在求得子球繁殖率、品質一致性及生長速 度上取得平衡,是生產者最大的挑戰。台灣地處亞熱帶氣候,對培育鱗 莖有良好的環境條件,適合孤挺花周年生長,若能自行開發培育新品種,
並整合種球繁殖、栽培、採後處理與花期調控等技術,而不必全仰賴進 口開花球,無論在生產盆花、切花或作庭園花壇觀賞,皆具有產業發展 潛力。
貳、前人研究(Literature review )
一、 植物學分類及性狀
孤挺花(Hippeastrum hybridum Hort.)為熱帶性球根植物,原產於中南 美洲,主要分佈於巴西、祕魯、墨西哥、智利、阿根廷(Rees,1985)。在植 物 分 類 上 屬 於 單 子 葉 亞 綱 (subclass Monocotyledonae) 石 蒜 科 (Amaryllidaceae) 孤 挺 花 屬 (Hippeastrum) 多 年 生 草 本 鱗 皮 鱗 莖 (tunicated bulbs)之球根花卉(劉等,2004)。中文別名為孤挺花、百枝蓮、石蒜花、喇
叭花、華冑蘭及鼓吹花 (王,1995;劉,2005)。
Hippeastrum 是由希臘語騎士(hippeus)和星星(astron)兩個字所組合而 成(Huely et al.,1992)。一般常用英文名稱為 Amaryllis,因原生種的不同,
又 有 Barbados Lily (H.equestre;H. puniceum) , Butterfly Amaryllis (H.papilio),Dutch Amaryllis,Green Amaryllis (H.calyptratum),Equestrian Star-Flower (H.Equestre;H.puniceum),South African Amaryllis (Okubo,1993) 等多種名稱。
孤挺花之球根屬於有皮鱗莖(tunicated bulb),為葉片基部肥大之鱗片 (leaf scales)所組成,最外層包覆之咖啡色皮膜是由養份耗盡之鱗片所組 成,具有保護及防止水份散失之功能(楊,2007),種球耐乾燥力強,成熟 的鱗莖周徑 20cm 以上即達開花球。依品種的不同,一般植株高度約 25~60cm(小型、中型花),大型花植株高度可達 90cm,葉為帶形或線形,
葉寬為 2.5~6cm(Okubo,1993)。花莖中空,小花由一對大苞葉(Spathe)包被 著,每一各別的花朵著生於更小絲帶狀苞葉的葉腋(axial)上,花序為繖形 花序(umbel),由 2~6 朵花所形成,每一朵花是由六枚花被(perianth)所組 成,花萼與花瓣不易區分(劉等,2004),花朵大而華麗但較欠缺香氣,花 色主要為紅色、粉紅色、白色、黃綠色或雙色;花柱較雄蕊為長,柱頭三 裂,心皮三個互相癒合而成,子房下位三室,果實為蒴果(董,1985;
Rees,1985),種子黑色扁平質薄,每個子房約 20~30 個種子,每個蒴果大 約 80 幾個種子。果實裂開後應即採收並馬上播種,種子過份乾燥易失去 萌芽力,種子壽命短(黃,1980)。
二、 育種歷史及目標
孤挺花的原生種約有 70 多種,現今之園藝栽培種乃經由原生種選種 後再經多年雜交育種而來。孤挺花之育種歷史已超過 200 年,培育出 300 多個栽培品種。而最早育出的雜交種為 Johnsonii 氏在 1977 年由 H.vittatum 和 H.reginae 於英國育成。孤挺花育種最主要的兩群為 Reginae 和 Leopoldii 之雜交群,Reginaef 群是由荷蘭 Jan de Graff 父子利用 H.vittatum、 H.striatum和H.striatum 和H.psittacinum 雜交選育出適合於歐洲之優良品 種。Leopoldii 則是由 Richard Pearce 發現的 H.leopoldii 和H.pardinum,從 南美洲安地斯山脈引進英國,具有花大、開展及對稱之生育特性。而目前 普 遍 栽 培 的 品 種 大 都 由 H.aulicum , H.reginae , H.reticulum , H.solandriflorum,及 H.vittatum 等原生種經長期雜交育種而來,所以用 hybridum 的種名(黃,1980;Okubo,1993)。
孤挺花基本的染色體數 N=11(Darlington and Wylie,1955),目前雜交栽 培種已從二倍體發展到三倍體和四倍體,四倍體品種如‘Apple Blossom’、
‘Lucky Strike’及‘Red Strike’ (Khalee and Siemsen,1989),因此花朵特性也多 傾向於大花、花型圓、花瓣寬大等,花型種類多計有喇叭型、圓形花、石 蒜花型、蘭花型、袖珍型、半重瓣及重瓣花型。花色從深紅、紅色、粉紅、
鮭魚色、橙色、黃綠色及白色等,或具有深淺雙色調之花色,花色亦有暈 色、斑點、條紋及鑲邊等變化(Rees,1985)。現今全球種球生產的主要產地 為荷蘭、以色列、南非、史瓦濟蘭(Okubo et al,1993)。孤挺花的育種及生 產目前以荷蘭的 Ludwing Warmenhove 公司及南非的 HADECO 公司為主 要的新品種培育機構。未來孤挺花主要育種目標方向為(1)耐寒品種(2)
具香味品種(3)新花色品種,尤其是金黃色、藍色到紫黑色系列(4)抗 病品種(劉等,2004)。
三、 鱗莖之形態
球根花卉種類很多,依其養份蓄積部位之不同,可分為鱗莖(bulb)、
球莖(corm)、塊莖(tuber)、塊根(tuberous root)、根莖(Rhizome)等。不同種 類球根花卉之間其繁殖法也不相同。孤挺花為石蒜屬之鱗莖花卉,故以探 討鱗莖花卉之形態為對象。
鱗莖形成是由厚肉質之鱗片包裹於一個特化、短縮莖上所形成之鱗 莖,具有貯藏養份與生殖生長(reproduction)等功能,依 Hartmann(1990)將 鱗莖花卉依鱗片組成形態及最外鱗片膜之有無可分為:
1.無皮鱗莖(non-tunicate):
鱗片分離的著生於基盤上,鱗片之最外層無乾鱗片膜,如百合。
2.有皮鱗莖(tunicate):
鱗片連續以頂芽為中心,作同心圓方式包裹為鱗莖,鱗莖之最外層有 乾鱗片膜包裹,可防止鱗莖乾枯及外力傷害,如孤挺花、風信子、西洋水 仙、金花石蒜等。有皮鱗莖依鱗片組成來源不同又可分為三類:
(1)鱗莖全由葉基部肥大之葉鱗片所組成,葉基部肥厚主因葉片養份蓄積的 結果,每一片葉片形成一鱗片稱葉鱗片(leaf scales),如孤挺花、金花石蒜 等。
(2)球鱗片之組成一部份為葉鱗片,另一部份為真鱗片(true scale)如西洋水 仙、風信子等。
(3) 鱗片全部由真鱗片所組成,如鬱金香。
孤挺花成熟的鱗莖周徑約為 20~30 公分,由葉基部肥大之葉鱗片所組 成,大約含有 6 個枝葉組(shoot units),每個枝葉組一般含有 4 片葉片(3 片全周葉及 1 片半周葉)和一個花序(花序的基部不會形成鱗片),最外部的 2 個枝葉組沒有地上部葉片,中間兩組有萌發的葉片,最內部的 2 組葉片 也不會萌發。鱗片在短縮的莖上包裹中心著生於全周,其短縮莖呈假軸分 枝系統(pseudo sympodial branching system)。新的小鱗莖(bulblet)會產生於 鱗莖最外部鱗片的葉腋處,子球會在第九片葉和第一個花序產生時萌發,
第一個花序通常會夭折。一般是由第九片葉(後 4 個枝葉組)開始萌發開 花,花序的位置在葉片萌發時側面相對位置,並相對形成一軸線,在此稱 為開花軸。其花芽分化在一年之中會發生數次並與營養生長同時進行,但
是有些花芽並不會達到開花階段,而在鱗莖內枯死(Rees,1985)。
四、生長發育和開花
鱗莖初期是營養性的生長,要到周徑大約 20 公分以上,才從幼年期 轉為成熟期,達到可開花的鱗莖。孤挺花在台灣可周年生長露地栽培,
但若夏季陽光太強,宜適度遮蔭,若在溫室的環境中,給予不斷的供水,
植株則呈常綠無休眠狀態,在一年當中可由莖頂生長點,有規律地生長 12 葉片(Rees,1985)。鱗莖生長最低溫限制為 8℃,最適合生長溫度為日 溫 23℃,夜溫 18℃(Hanks,1985)。鱗莖形成與肥大多為環境不適宜時於 休眠前期進行物質貯藏,為後期休眠作其準備,栽培田間的孤挺花所表 現出之地上部生育狀況及鱗莖肥大程度的差異會隨著春、秋季節溫度升 降而產生(林,1970)。冷涼的溫度會增加鱗莖內碳水化合物和醣類的累 積,使鱗莖的膨漲增大和抑制地上部的生長,溫度降低時同時也會提高 鱗莖內氮、鈣、磷、鎂和鉀營養的吸收。
孤挺花需種植於排水良好且富含有機質的砂質土中,PH值在6-7之 間。子球生長初期不需營養施肥,鱗莖愈大時增加營養需求及提高CO2
濃度能增進鱗莖的大小(Silberbush,2003)。在適當的環境條件下,影響 鱗莖發育的主要因素是光合作用,支持最佳光合成能力的生長環境能提 高子球生長速率。在肥培管理方面,由於孤挺花耐肥性頗佳,生長期間 每個月施加液態追肥,肥料比例以氮-磷-鉀=20-20-20之複合肥料為佳,
可促進種球肥大。水份管理則以土壤維持濕潤,不可積水及土壤過分乾
燥,約一年至一年半即可培養出周徑20公分以上之開花球。
一般孤挺花之開花期大都集中於3-5月,即台灣清明節前後。盆花和 切花生產以選擇周徑20公分以上之開花球,品種選擇宜考慮可抽出多花 莖之品種為主,以增加切花適量。切花採收時機為蕾期採收,當花苞著 色時即可進行切花採收,為避免花莖基部裂開外捲,可先以0.125M的蔗 糖溶液浸1天即可改善。促成栽培時,開花球先置於23~25℃通風環境下2 星期,後貯藏於13℃相對溼度80%環境下8~10星期,若貯藏於5~9℃環境 下,可貯藏較久之時間。在種球處理與貯藏期間,應保持根系活力,避 免種球凍傷及過於失水狀態。
五、 種球繁殖之方法
孤挺花其種球繁殖方法與其它石蒜科植物相似,有性繁殖為種子實 生法,而無性繁殖則有自然分球法、鱗片繁殖法及微體繁殖,可依植物 種類特性及利用目地不同採用不同的繁殖方式,以下分別簡述之:
(一)、種子實生法
孤挺花實生苗從種子播種至開花需3年以上才可達商業開花球標準 (黃,1980)。球根花卉由種子繁殖需要較長之幼年期,故至開花所需時 間較長,如金花石蒜(Lycoris)由播種至開花需4-5年(呂,1995),鬱金香 需要6年(Rees,1993),風信子需要4-6年(De Hertogh and Le Nard,1993)。孤 挺花以種子繁殖時需以新鮮種子播種(圖1)才能達最高之發芽率,貯藏愈
久發芽率愈低(黃,1980)。種子一般於晚春或早夏採收,採收後馬上播種 發芽率最高,且必須於5星期內播種完(Rogers,1976),種子於溫度25℃有 最高之發芽率,溫度超過25℃或低於20℃時發芽率會遞減,種子貯藏於 相對濕度11%(5℃)或52%(15℃)下,可維持種子活性長達1年(Carpenter and Ostmark,1988a);種子於低溫5℃貯藏若超過3個月以上,發芽率會快 速降低(呂,1995)。經由種子繁殖之後代會產生遺傳變異,植物花色及 株形等性狀不易控制,且生長所需時間較長,故通常只用於培育新品種 的育種時採用,少用於商業性種球生產。
(二)、自然分球法
一般鱗莖生長至某一階段會形成小鱗莖或分球(offsets),植物種類不 同分球數亦不相同,如鬱金香、鳶尾開花後母球會形成5個子球,少數大 的子球於第二年可開花;西洋水仙自然分球率為1.6倍(Hanks,1985);納 麗石蒜平均分球數為2個,溫度較低時分球數較高(賴,1995)。金花石蒜 自然分球率為1.8倍(呂,1995),金花石蒜在開花後之種球才會有子球產 生。納麗石蒜在未進行切割、生長點未受傷之情形下,不易自然分球 (Vishnevetsky et al.,2003)。孤挺花之自然分球數依品種而有所不同,De Hertogh 和Le Nard(1993)根據215個品種調查分球數介於0.1~17.3之間變 化,求得平均分球數為2.7個。其中只有8個品種(‘Brillant Red’、‘Carrara’、
‘Faust’...et.al)分球數超過10個。根據子球大小和生長環境的不同,子球 長至開花球需要1~2年。子球產生於鱗莖最外部鱗片的葉腋處,新的子球 會在第九片葉產生時萌發(Okubo,1993),子球生長於葉片萌發時之兩側
(圖2),並相對成一個軸線,與開花軸位置相同,未開花球或子球則少有 分球情形。此種繁殖方式所需的設備、技術性及勞力成本較低,可應用 於鬱金香、鳶尾等分球數較多,子球至開花期較短之作物,但對孤挺花、
西洋水仙、金花石蒜及納麗石蒜等自然分球率低的植物,無法應用於商 業性生產。
(三)、鱗片繁殖法
鱗片繁殖主要是將植株鱗片以切割的方式分為幾部份,以促進不定芽 及小植株的再生(Hertogh and Nard,1993)。由於石蒜屬鱗莖再生部位為背 軸面靠近基部的位置,小鱗莖是由鱗片背軸面接近基盤之表皮細胞及次 表皮細胞分裂所形成,小鱗莖之維管束走向與外鱗片維管束相接,在鱗 片基部接近兩邊切口處著生,但偶而也有小鱗莖著生於雙鱗片內側或外 側之鱗片基盤處。依種球切割方式主要可分為下列三種:
1.基底刻溝法(notching)
依種球大小從鱗莖基盤底部以放射狀切割8~16等份(圖3),不切開 球體,切割深度約為鱗莖之2/3,破壞頂芽優勢,促使不定芽的形成,
在鱗片基盤兩側可產生子球。風信子(Hyacinths)基底刻溝後,一個種球 可得12~24個子球,子球2~3年可達開花球;金花石蒜一個種球可形成 8~20個子球(呂,1995)。孤挺花種球利用刻溝法可產生8~17個子球 (Ephrath,2001),刻溝法及瓣狀鱗片不同切割方式對小球形成數之影響 差異不大,子球形成數約為12~25個(朱,2003)。
2.瓣狀鱗片法(chipping)
於鱗莖基部以放射狀切開球體成為瓣狀,依種球大小作8~16等份 切割,以此瓣狀鱗片為繁殖體。西洋水仙一個種球可得12個小鱗莖,
第二年約有14~71%鱗莖可開花(Hanks,1991);金花石蒜以8等份分割 後,一個種球可得13個小鱗莖(呂,1995);孤挺花以瓣狀繁殖可形成 12~25個小鱗莖,刻溝處理所形成之小球因生長空間受到擠壓,形狀較 不規則,瓣狀鱗片因生長空間較大,所形成小球較圓。刻溝及瓣狀鱗 片所形成小球生長期間較短,在17個月栽培可達30cm開花球(朱,2003 )。
3.雙鱗片繁殖法(twin-scaling)
雙鱗片繁殖法是由Luyten(1926)所開發之孤挺花人工繁殖技術,而後 經由Traub(1933)、Heaton(1934)和Vijverberg(1981)等人陸續改良建立標準 之雙鱗片繁殖技術(twin-scaling),並應用於孤挺花商業種球生產上。此 法為將種球以放射狀切割8等份後,再將瓣狀鱗片以兩個鱗片及其相連部 份基盤切割為雙鱗片(twin-scale)繁殖體。孤挺花一個種球約可切成40組 雙鱗片,一組雙鱗片平均得1.5個小鱗莖(Huang et al.,1990a),小鱗莖培養 3年可達開花球。西洋水仙一個種球平均可得48~90個小鱗莖,培養3-4 年可達開花球(Hanks,1997);金花石蒜一個種球平均可得50-75個小鱗 莖,經培養2-3年可達開花球(呂,1995)。納麗石蒜平均一個雙鱗片可得1.27 個小鱗莖(賴,1995)。目前除了孤挺花、西洋水仙外,金花石蒜、納麗石 蒜等石蒜科植物多以此法應用於商業上種球之繁殖。
(四)、微體繁殖法(micropropagation)
即利用組織培養技術繁殖種球,以鱗片或莖頂部份為培殖體進行無 菌培養,大量誘導不定芽分化。組織培養適用於大量標準化繁殖。水仙 以雙鱗片為培殖體,經組織培養六個月後,每個種球可得200-300小鱗莖 (Hussey,1982)。金花石蒜利用組織培養八個月後,得50~75株小鱗莖 (林,1987),瓶苗出瓶後溫室栽培4~5年可達開花球。微體繁殖法因生產成 本高、生長期間長及瓶苗出瓶後存活率等問題,只用於新品種之大量繁 殖或是莖頂培養無病毒種球,直接應用於商業性種球仍少見。
六、 影響子球數量及生長速度之因子
種球利用鱗片繁殖法時常受到內在生理及外在環境因子的影響,而 使繁殖倍率和生長發育速度有所不同,因此必須瞭解各因子條件對繁殖 體之影響,才能於種球繁殖時達最佳繁殖率及種球品質一致性,各項影 響因子說明如下:
(一)、生理因子
1.品種特性
對不同種類植物,雙鱗片繁殖法小鱗莖形成率會有差異,其中以孤 挺花較容易繁殖,納麗石蒜則不易成功(Rees,1992)。Pierik和Ippel(1977) 在納麗石蒜試驗中,比較五個不同營養系和三個雜交系之雙鱗片繁殖 法,小鱗莖形成率結果有顯著差異,小鱗莖形成率最高者可達100%,最
差者僅有45%。孤挺花不同品種間雙鱗片繁殖法比較,其中以‘Apple Blossom’的94.4%小鱗莖形成率最高,每組雙鱗片平均形成個數為1.28 個,‘Best Seller’次之為86.1%,而‘Red Lion’形成率最低只有76.4%,此 結果與田間栽培自然分球率的表現相同(張,1998)。
2.繁殖體之大小
種球的品質由鱗莖的大小所決定,種球的大小代表開花能力及其商 業價值,種球周徑達20公分以上具有開花之能力,孤挺花商業上的種球 依周徑大小來分級,可分為20-23、24-27、28-31及32cm以上。帶有鱗片 基部相連接之基盤部份稱為繁殖體(propagule),種球愈大,鱗片繁殖體 所提供之養份愈多,且繁殖體所帶鱗片數對於繁殖倍率有很大的影響,
金 花 石 蒜 鱗 片 繁 殖 , 鱗 片 數 愈 高 所 誘 導 之 子 球 數 及 發 根 率 愈 高 (呂,1995)。繁殖體大小可依鱗片寬度、鱗片長度、二方面探討如下:
(1)鱗片寬度
母球切割不同份數,會影響繁殖體鱗片基盤之寬度。當鱗片繁殖體 愈大,所得之子球愈大、子球數也愈多(Squires,1990)。西洋水仙母球以 4(8twin-scales)、8(16)、16(32)及24(48)等份切割,子球形成數以16、24 分割最多,8分割最少,但小鱗莖以8分割者最重,24分割最輕(Hanks and Rees,1978)。金花石蒜鱗莖以4(8twin-scales)、8(16)、16(32)及24(48)分割 時,不同寬度下小鱗莖形成率以4、8分割較高,16和24分割之形成率低,
推測原因為16和24分割之繁殖體較細小,易發生腐爛現象。納麗石蒜鱗
片寬度愈寬,可形成小鱗莖之基盤也較寬,可產生小鱗莖之空間較多,
小鱗莖形成數也較多(Pierik and Ippel.1977)。孤挺花鱗片基盤寬度與小鱗 莖形成數呈顯著正相關,隨著基盤寬度增加,小鱗莖形成數也增加,基 盤最寬之6.0~6.5mm平均可達3.92個,基盤最小之3.5~4.0mm平均有1.39 個;培育初期6~8週時各處理間小鱗莖形成數沒有差異,但第16週調查 時,基盤寬度小者,因相對生長空間及養分有限,有許多小鱗莖無法正 常生長發育,而使可用之小鱗莖數目減少(張,1998)。
(2)鱗片長度
納麗石蒜鱗片長度愈長,小鱗莖形成愈早且生長愈快,小鱗莖也愈 重(Pierik and Ippel,1977)。金花石蒜取2、4及6公分長度之雙鱗片繁殖體,
小鱗莖形成率無顯著差異,平均每個繁殖體之小鱗莖數為1.3 個,鱗片 長度對小鱗莖大小有顯著影響,隨著鱗片長增加而小鱗莖愈大,發根率 也愈高(呂,1995)。孤挺花雙鱗片之內、外鱗片長度均需大於5cm,才可 形成小鱗莖,且鱗片愈長,小鱗莖生長愈快;其中又以外鱗片長度對小 鱗莖之影響較大,外鱗片愈長小鱗莖葉片數也愈多(Okubo et al.,1990)。
3.鱗片位置及其成熟度
鱗莖之鱗片位置愈靠外側其鱗片愈早熟。西洋水仙將鱗片由外至內 分為1~ 5層,小鱗莖形成率以第2層最好,小鱗莖重以第3層為佳,第5層 之小鱗莖形成率及小鱗莖重均最差(Hanks,1980)。雙鱗片以外層鱗片(第 1~3層)小鱗莖形成較早,小鱗莖約12~16週可達體積高峰,愈內側之鱗片
(第4、5層)需24週後小鱗莖方達最大體積;其中以3~4層小鱗莖形成數最 多,重量也最重,第1層則表現最差(Hanks,1985)。金花石蒜小鱗莖形成 率第1~5層無差異,第6層較差;小鱗莖大小則以第1~2層最大,第3~5層 次之,第6層最小(呂,1995)。孤挺花鱗片依位置由外到內分為三層,以 最外層小鱗莖形成數較多,第二層小鱗莖形成數次之,第三層小鱗莖數 最少(Okubo et al.,1990)。將孤挺花鱗片由外而內分成6組(第PI~PVI)不同 成熟度之雙鱗片,小鱗莖形成率以第PII~PIV最好,可達94.4%以上,PI、
PV及PVI較差,在77.8%以下,小鱗莖形成數、重量及大小均以中層的 PII~PIV較佳,而最內層最差(張,1998)。
(二)、環境因子
球根花卉鱗片繁殖法常因培育環境而影響鱗片的成活率(或腐爛 率)、子球形成率、生長速度及大小等,其環境因子分別如下描述:
1.繁殖時期
西洋水仙雙鱗片繁殖於每月處理均可形成子球,以6月及9月之子球 形成數最多(Hanks,1977);金花石蒜於不同月份取雙鱗片做組織培養,結 果以5-7 月處理之出芽率高、腐爛率低,處理時期最好(林和馬,1987);
呂(1995)以金花石蒜雙鱗片繁殖,小鱗莖形成率以4-6月處理最佳,7月以 後處理略低,至11月處理時最低。孤挺花之鱗片組織培養處理時期於 1-3 月 進 行 為 佳 (Huang,1990) , 孤 挺 花 之 鱗 片 繁 殖 則 以 11-1 月 為 佳 (Rees,1992)。
2.溫度
Hanks和Rees(1978)以西洋水仙雙鱗片繁殖,栽培溫度以15-20℃小 鱗莖形數較多、重量較重,9℃溫度處理最差;雙鱗片栽培於13-20℃中,
溫度愈高小鱗莖數愈多。納麗石蒜雙鱗片繁殖之溫度以20-24℃之子球 數、子球重最佳(Hanks,1986)。金花石蒜雙鱗片繁殖以25℃處理較佳(呂,
1995)。小球生長環境中的溫度條件會影響小球的形成數及大小,孤挺花 在20/10℃及25/15℃的瓣狀鱗片處理中,鱗片成活率100%、小球形成數、
小球周徑及鮮重為各處理中最佳,在30/20℃下,成活率最低僅83.3%,
小球形成數為各處理中最小值(朱,2003)。28/23℃的高溫會造成孤挺花實 生 苗 球 根 肥 大 不 良 , 於 23/18℃ 、 18/13℃ 下 球 根 重 的 增 加 速 度 最 快 (Hayashi,1970);不同溫度下對鱗莖肥大之影響,鱗莖肥大以20℃最佳,
其次為15℃、25℃,地上葉之生育以25℃最旺盛,其次依序為30℃、20℃、
15℃(張,2004);鱗莖生長最低溫限制為8℃,而最佳生長溫度為日夜溫 23/18℃(Hayashi,1970)因此,可得知10-25℃的涼溫適合孤挺花小球生長 發育,而30℃前後的高溫易造成鱗片的腐爛及小球生長速度的減緩,故 秋冬至初春冷涼氣候較適合小球繁殖,若在夏季高溫期間繁殖時必須留 意溫度的控管(朱,2003)。
3.介質濕度
在鱗片繁殖上常將繁殖體直接扦插於栽培蛭石介質中或是直接放置 於保濕之塑膠袋中,栽培介質之相對濕度會影響鱗片之腐爛率及小鱗莖
形成率及重量大小,介質溼度太高其鱗片容易腐爛而影響小球形成率。
金花石蒜於不同溼度介質進行雙鱗片繁殖,結果顯示介質水分含量對小 鱗莖形成率無顯著影響,但對小鱗莖大小有影響,介質水分含量增加,
小鱗莖生長速度也較快(呂,1995)。孤挺花不同溼度介質處理中,其結果 亦同(張,1998)。
4.光線
金花石蒜雙鱗片繁殖中,光或暗處理對小球形成率並不顯著,但於 暗處理會形成較多的根,且小鱗莖也較大。雙鱗片繁殖期照光,會使鱗 片 及 所 形 成 的 小 鱗 莖 呈 現 綠 色 現 象 , 而 暗 處 理 的 小 鱗 莖 均 呈 白 色 (呂,1995)
5.消毒處理
鱗片繁殖時需栽培於高濕的環境下,以避免小鱗莖尚未長出而鱗片 已枯萎的情形,但於高濕環境下繁殖體易發生病原菌污染。一般鱗片腐 爛原因可分為三類(1)生理性-無病蟲害感染,可能為鱗片養分不足,而 不能形成子球。(2)真菌感染-以青黴污染較多,或母體本身之病源。(3) 蟲害薊馬危害-可能由母球或栽培環境傳染。西洋水仙鱗片繁殖時,種球 會先以54℃熱水處理1小時,既可有效防止污染,又不影響小鱗莖形成
(Linde,1992);納麗石蒜雙鱗片繁殖時,會先將培植體以45℃溫水處理 30分,再以70%酒精浸泡1分鐘表面消毒效果最好(賴,1995)。
七、孤挺花育種及產業發展概況
(一)、全球
現今全球種球之生產量約為 3000 萬球,主要供應國為荷蘭,約有 1500 萬球(Henk,2005),荷蘭種球生產量占全世界之一半。荷蘭的孤挺花 生產皆於溫室中,採用雙鱗片標準化生產商業種球,溫室夏季會以膠布覆 蓋或遮陰系統以降低溫度,冬季時則在底部或地上使用加熱或冷卻導管。
在南非及史瓦濟蘭,孤挺花生產均於戶外田間。在日本,鱗莖通常是在 3 月到 4 月中開始植栽於戶外,10-11 月再將鱗莖拔起,若在較溫暖潮濕的 氣候區,冬天仍然留在戶外。
荷蘭孤挺花種球生產公司為Amary Uiskwekerij,此家企業是家族企 業起家,目前已是第三代,共有三座種球培育場,總生產面積為93,000平 方公尺,以生產種球為主,此家並與另一家公司Nieuwkerk合作,Nieuwkerk 公司主要進行育種及盆花與切花生產,這二家公司聯合以分工方式生產孤 挺花行銷全世界。
(二)、台灣
行政院農委會台中種苗改良場自民國90年開始進行孤挺花品種收集 及育種工作,目前共已收集130個商業品種。孤挺花之雜交育種工作每年 皆會進行,初步篩選共選出具有潛力之孤挺花新品系100個,目前選育出
之「種苗1號-粉珍珠」、「種苗2號-紅豔」重瓣品種,已獲得品種權(劉,
2007)。民國96年農委會舉辦孤挺花新品系100單株觀摩會及民眾票選活 動,根據票選統計前幾名結果,可看出台灣南部民眾對於孤挺花顏色仍 較偏好花型大且紅色系之品種,但若顏色特殊少見,也很受民眾青睞,
此結果也可提供育種者日後選種相當重要之參考依據。從雜交後裔中,
有些單株於播種後培育一年半即可開花,具有早花性,有些親本所雜交 之後裔表現相當一致,可作為進一步研究,發展出F1種子販售。目前以 選拔出表現不錯之單株,配合溫室之栽培管理及花期調控,正進一步進 行量化之工作,期望創造出適宜台灣氣候發展及受一般大眾喜好之孤挺 花自有品種,以作為台灣進軍國際之商業品種。
在民間產業方面,久雅植科股份有限公司於2006年成立,專營孤挺 花的育種、種球大量生產、切花及花期調控等研究。經營者為楊德麟先 生,被喻為孤挺花達人,其育種、生產的苗圃場區設於宜蘭縣礁溪近郊,
面積約4200坪,設有溫室、網室、地面苗床等,在一般民間孤挺花培育 者中少有。其所擁有之種球來源搜集遍佈台灣各地,並結合了十七世紀 引入之早期原種與二十世紀的加入品系,其中也經過自由授粉、種子苗 產生大量新性狀,培育至今。久雅植科育種的優勢是不必仰賴國外進口 生產種球,因種球本已長久適應台灣氣候、土宜,故希望能創造台灣孤 挺花的培育風氣,並打入國際市場,為台灣花卉產業盡一份心力。
参、材料與方法 (Materials and methods)
一、 植物材料
本試驗所採用之材料,均取自國立宜蘭大學園藝學系培養 2-3 年之種 球,其鱗莖周徑為 20~32 公分。鱗莖由土中挖取後去除葉片及根系,鱗莖 留頸 2-3cm,洗淨表面且剝除最外層之乾枯鱗皮,置於室內環境靜置 1 天後 進行試驗處理。本研究所使用品種之特性如下:
1.‘Red Lion’紅獅子
屬於大花單瓣品種,花朵為單一鮮艷大紅色,花莖長度約可達 60cm,每 枝花莖可開 4-6 朵花,適合做為盆花栽培(圖 6A)。
2.‘Apple Blossom’蘋果花
屬於大花單瓣品種,花朵顏色淡粉有如蘋果而得名,花莖長度約可達 60cm,每枝花莖可開 4-6 朵花。此品種容易自然分球,可作為盆花及切花
(圖 6B)。
3.‘Lady Jane’珍女士
屬於大花重瓣品種,花朵深橘色帶潛白色線條,每一花莖約可開 4~6 朵花,適合做為盆花及切花栽培(圖 6C)。
4.‘Double Record’雙紀錄
屬於中型花重瓣品種,花朵外緣為紅白雙色相間,每一花莖約可開 4~6
朵花,適合做為盆花及切花栽培(圖 6D)。
5.‘Blossom Peacock’花孔雀
由日本選育而得,花為重瓣,花瓣外緣為紅色,中間為白色,每一花莖 可開 4~6 朵花,花莖長度約為 50~65cm(圖 6E)。
二、 其他材料
試驗所使用之栽培介質為二號蛭石(南海蛭石工業股份有限公司),栽培 容器為自華興園藝資材行所購得長方形植槽(長 68×寬 43×高 22cm)及四層置 物架(長 52×寬 33×高 108cm)。
三、 試驗環境
本試驗種球切割繁殖技術於本系館實驗室進行,之後栽培環境為本系簡 易溫室及宜蘭龍潭苗圃場區溫室。試驗期間控制澆水,維持介質之溼度,以 利植株之生長發育。
四、 試驗設計與統計分析
所有試驗皆採完成逢機設計(Complete Randomized Desigh,CRD),而本 研究所調查的各項資料,採用 Excel 和 SAS 統計分析軟體,以鄧肯氏多變 域分析(Duncan’s Multiple-Range Test),檢驗各處理間 5%的差異顯著性。
五、 試驗調查項目
1. 種球周徑:以皮尺測量鱗莖周長,以公分為單位,記錄小數第一位。
2. 種球鮮重:種球去除根葉後,鱗莖頸部留 2-3 公分,以電子天平測量種 球重量,記錄至小數第一位。
3. 子球直徑:以電子游標尺量取子球直徑,以公厘為單位,記錄至小數第 一位。
4. 子球數:測量自鱗片基盤長出之子球數目。
5. 小植株重:以電子天平測量小鱗莖全株重,包含根重、葉片重及子球重,
以公克為單位,記錄小數至第一位。
6. 最大葉長:以皮尺量取最長葉片之長度,由葉基部量至頂端,以公分為 單位,記錄至小數第一位。
7. 葉片數:計數每個子球所長出之總葉片數。
六、 試驗方法
(一)、不同切割繁殖法之比較
本試驗目的為利用不同切割繁殖方式來改進孤挺花之子球繁殖率及其 生長速度,以開發簡易操作、大量繁殖之孤挺花人工切割繁殖法。於 96 年 10 月 22 日取周徑為 26~29cm 之種球,品種為 Lady Jane、Double record 及 Red Lion 三個品種,分別進行刻溝、頂切、刻溝穿透、刻溝雙鱗片四種不同 種球切刻方式,每處理 3 重複,每重複 1 球,共 12 球。切割方式說明如下:
1.頂切(top-notching):將種球自鱗莖頸部往下切割成 8 等份,切割深度不超 過 2/3,每等份鱗片與種球不分開(圖 7A)。
2.刻溝(notching):將種球自基盤底部往上切割 8 等份,切割深度不超過 2/3,
每等份鱗片與種球不分開(圖 7B)。
3.刻溝穿透(notching coring):將種球作刻溝 8 等份切割後,再將種球中心穿 透挖除(圖 7C)。
4.刻溝雙鱗片(notching twin-scaling):將種球作刻溝 8 等份切割後,再將每 等份鱗片從基盤處由外往內切成雙鱗片,約可切成 5 層雙鱗片,鱗片與種 球不分開(圖 7D)。
種球切割後於室溫下放置二天後,再將種球種植於植槽中,每植槽種 6~8 球,介質使用保濕蛭石,栽培於本系簡易溫室。四個月後再將子球取出,
對各處理之子球形成數、子球直徑、小植株重、葉片數及最大葉長等項目進 行調查(圖 8)。
(二)、刻溝雙鱗片繁殖法之試驗
因於實驗(一)四種不同切割繁殖法中,刻溝雙鱗片繁殖法所形成子球數 量最多,其繁殖倍率於四種切割法中為最高,故此改良鱗片切割方式確實能 增進子球形成數量。本試驗目的即利用此法來探討刻溝雙鱗片繁殖法對子球 數量及生長速度之影響,本試驗分為下列五個方面來探討:
1.品種間之比較
試驗目的為比較品種間的差異在刻溝雙鱗片繁殖法上對子球數量之影 響,在各試驗中比較的品種有‘Lady Jane’、‘Double Record’、‘Red Lion’、
‘Apple Blossom’及‘Blossom Peacock’五個品種。
2.種球大小之比較
本試驗目的為比較種球不同大小利用刻溝雙鱗片繁殖法對子球數量及 生長之影響,在種球大小與繁殖倍率之間找出最適經濟利益。於 97 年 7 月 4 日取‘Apple Blossom’周徑分為四組 30cm、26cm、22cm、15cm 不同大小之 種球,進行刻溝雙鱗片繁殖法,每組處理為三重複,每重複 2 球,四組處理 共 24 球。種球先刻溝 8 等份後靜置一天,種球略為向外張開後,再進行刻 溝雙鱗片處理,於室溫下靜置二天,再將種球種植於植槽中,每植槽種 6~8 球,介質使用保濕蛭石,培育於本系簡易溫室。繁殖四個月後再將子球取出,
調查子球形成數、子球直徑、小植株重、葉片數及最大葉長等項目(圖。
3.不同切割時期之比較
本試驗目的為於不同月份十、一、四及七月進行刻溝雙鱗片繁殖法,以 探討不同切割時期對子球數量及生長情形之影響,找出一年中最佳繁殖之時 期。取‘Lady Jane’周徑 28cm 以上之種球,分別於 96 年十月、97 年一月、
四月及七月初進行刻溝雙鱗片繁殖法,每個處理三重複,每重複 2 球,共 24 球。種球先刻溝 8 等份後靜置一天,種球略為向外張開後,再進行刻溝 雙鱗片處理,於室溫下靜置二天,再將種球種植於植槽中,每植槽種 6 球,
介質使用保濕蛭石,培育於本系簡易溫室。繁殖四個月後再將子球取出,調 查子球形成數、子球直徑、小植株重、葉片數及最大葉長等項目。
4.靜置不同天數之比較
本試驗目的為探討繁殖初期無栽培介質且靜置不同天數下,對子球數量
及生長速度之影響,找出種球切割後最佳靜置天數及培育環境,使鱗片腐爛 率降低提高子球形成率。98 年 3 月 12 日取‘Lady Jane’周徑 30cm 以上之種 球,種球先刻溝 8 等份後靜置一天,種球略為向外張開後,再進行刻溝雙鱗 片處理,分別靜置 1 天、2 天、3 天、4 天及 5 天癒傷組織處理,每個處理 三重複,每重複 1 球,共 15 球。靜置不同天數後直接放置於層架上無添加 任何介質,並用塑膠袋覆蓋其上維持栽培環境之溼度 RH85~90%,每日噴 水以保持鱗片潮濕,一個月後待小球長出再移至植槽中,每植槽種 6 球,介 質使用保濕蛭石,培育於本系簡易溫室。二個月後再將子球取出,調查子球 形成數、子球直徑、小植株重、葉片數及最大葉長等項目。
5.鱗片切割不同份數之比較
本試驗目的為利用刻溝雙鱗片繁殖法於不同切割份數下,不同大小繁殖 體對子球數量及生長之影響,找出最種球最適切割份數。98 年 3 月 12 日取
‘Blossom Peacock’和‘Apple Blossom’周徑 30cm 以上之種球,進行刻溝雙鱗 片繁殖法,切割份數分別為 8、8+1/2(切割 8 等份後再於每一等份鱗片之基 盤處劃上一刀)及 16 等份(圖 9),每個處三重複,每重複 2 球,共 18 球。種 球切割後靜置二天,再放置於層架上無添加任何介質,並用塑膠袋覆蓋其上 維持栽培環境之溼度 RH85~90%,每日噴水以保持鱗片潮濕,一個月後待 小球長出再移至植槽中,每植槽種 6 球,介質使用保濕蛭石,培育於本系簡 易溫室。二個月後再將子球取出,調查子球形成數、子球直徑、小植株重、
葉片數及最大葉長等項目。
(三) 刻溝雙鱗片產業大量生產之應用
本試驗利用開發的刻溝雙鱗片繁殖法與久雅植科公司合作,將此繁殖技 術應用於產業商業種球之生產。
1.種球不同大小之比較
於 97 年 10 月 18 日取品種 A210 大球(周徑 26cm)、中球(周徑 19cm)、
小球(周徑 13cm),每個處理四重複,每重複五球,共 60 球。種球進行刻溝 雙鱗片繁殖法。種球切割後種植於保溼的蛭石介質中,繁殖四個月後再將子 球取出,調查子球形成數、子球直徑、小植株重、葉片數及最大葉長。
2.刻溝、瓣狀及刻溝雙鱗片不同繁殖法之比較
於 97 年 12 月 17 日取品種 A210 周徑 21~22cm 之種球,分別進行刻溝、
瓣狀及刻溝雙鱗片三種不同切刻法,每處理 30 球,共 90 球。種球切割後種 植於保溼的蛭石中,繁殖四個月後再將小球取出,調查子球形成數、子球直 徑、小植株重、葉片數及最大葉長。
肆、結果(Results)
一、不同切割繁殖法之比較
本試驗目的為改進孤挺花之鱗片繁殖方法,以增進子球數量及探討其 生長速度。實驗結果如(表 1)所示,因種球切割方式的不同,所繁殖的子 球數量及直徑有明顯的差異。三個品種間皆以刻溝雙鱗片繁殖法所形成之 子球數量最多,在‘Lady Jane’平均子球數 41.3 個為最高(圖 13)。刻溝和刻 溝穿透所形成子球數次之且無顯著差異,子球數為 10.3~17.3 個。頂切法 在‘Lady Jane’和‘Red Lion’子球數量為 0,‘Double Record’子球數為 7.0 個。
表 2 是不同切割繁殖法所形成子球之生長情形,刻溝和刻溝穿透法子 球直徑為 16.0~19.0mm,刻溝雙鱗片法子球直徑為 11.3~14.7mm。在小植株 重方面,各處理有差異,刻溝法三品種依序為 3.6g、3.8g、5.9g,刻溝穿 透法為 4.2g、6.6g、3.7g,刻溝雙鱗片法為 1.3g、3.2g、2.1g 重。由結果顯 示出子球所形成數量愈多,其所形成之子球直徑會較小,相對小植株重也 會愈小。
三個品種間之比較,刻溝、刻溝穿透、及刻溝雙鱗片之子球形成數皆 以’Lady Jane’表現最高,‘Red Lion’次之,‘Double Record’最低(圖 18)。頂 切法只有‘Double Record’形成小球數,‘Lady Jane’和‘Red Lion’小球數皆為 0。刻溝和刻溝穿透在三個品種間子球形成數無顯著差異,但刻溝雙鱗片 在`Lady Jane’和‘Red Lion’下,所形成子球數最多且有特別顯著差異。
二、刻溝雙鱗片繁殖法
(一)、種球不同大小之比較
‘Apple Blossom’四組不同周徑大小之種球利用刻溝雙鱗片繁殖法之結 果如(表 3)所示,周徑 30、26、22、15cm 所形成之平均子球數分別為 44.8、
38.5、30.8、19.3 個,周徑 30cm 最多子球數可達 82 個,周徑 15cm 最多 小球數可達 30 個。周徑 30cm 和 15cm 之種球有顯著差異,26cm 和 22cm 無顯著差異。結果顯示種球周徑愈大,所形成子球數愈多(圖 19)。
種球周徑愈大,其重量顯著的大量增加,因此其所能提供的母球養份 相對也愈多,故將四組不同大小種球重量換算成平均每 50g 單位所形成之 子球數如(表 4),周徑 30cm、26cm、22cm、15cm 每 50g 單位重量所形成 之子球數分別為 7.3、9.2、9.9、17.9 個,以周徑 15cm 所形成之單位小球 數最多,隨著周徑增加,所形成之單位子球數卻呈遞減現象。
四組所形成子球數的大小其直徑分別為 6.1mm、4.5mm、4.1mm 及 5.7mm,種球 30cm 和 15cm 所形成之子球較大且無顯著差異,26cm 和 22cm 所形成之子球較小也無顯著差異。另外四組子球重分別為 0.33g、0.33g、
0.41g、0.27g,葉片數分別為 1.1、0.9、1.3、1.3 片,結果顯示其平均子球 重和葉片數四組均無顯著差異。
(二)、不同切割時期之比較
刻溝雙鱗片於不同月份切割時期對子球數及生長之結果(表 5)(圖 20)。十月、一月、四月、七月所形成之小球數分別為 41.3、51.3、7、47.9 個,以一月子球數最高,七月次之,但一月和七月子球數無顯著差異;
四月所形成子球數最少,且明顯差距甚多。因四月鱗片腐爛率非常高,
導致所形成平均子球數相對地降低許多。
不同月份所形成子球大小之比較,小植株重分別為 1.4g、1.6g、2.1g、
1.9g,以四月份所形成之子球最重,其餘月份無明顯差異。子球之直徑分 別為 11.3、11.5、15.3、12.2mm,以四月份所形成之子球直徑最大,其餘 月份無顯著差異。結果顯示,所形成子球數愈多,因母球養份移轉至子 球時,平均每個子球所分配之養份則會遞減,因此所形成之子球直徑、
子球重則為較小。因一月、四月、七月其種球周徑皆達 30cm 以上,但十 月份種球周徑未達 30cm,若周徑能達到 30cm 以上,所形成之子球數應 會提高,因種球周徑愈大,所形成子球數相對愈多。
由以上結果顯示,刻溝雙鱗片不同切割繁殖時期,以一月、七月最佳,
十月次之,四月表現最差,因四月鱗片腐爛率非常高,而會影響子球之 形成率,故四月為一年中最不適合繁殖之時期。
(三)、靜置不同天數之比較
刻溝雙鱗片切割後放置於蛭石中繁殖四個月後小球生長情形(表 4),品種 Apple Blossom 周徑 30cm 所形成之子球數為 44.8 個,子球直徑 為 6.1mm;若將刻溝雙鱗片切割後放置於無介質層架上繁殖一個月後,
再將鱗片移至蛭石中繁殖,三個月後小球生長之結果(表 6),小球數為 49.3 個,子球直徑為 5.5mm,由以上結果顯示,繁殖初期無介質培育能避免 鱗片腐爛,使子球形成率提高並能縮短小球生長期間,三個月即可達到 四個月蛭石培育之子球大小。再調查過程中,仍可發現許多小鱗莖由鱗 片基盤長出,若再栽培一個月,所形成子球數量及其大小均可再提高。
種球切割後癒傷處理不同天數對小球形成影響之結果如(表 7),切割 後放置 3 天及 4 天的處理小球形成數為 37 個及 34 個,是各處理中的最 高值及次高值,放置一、二天子球數無差異為 27.3、27 個,放置第五天 子球數最少為 24.7 個。癒傷處理放置一~四天其子球直徑無差異,分別 為 8.5、8.9、8.3、8.3mm,第五天最小為 7.1mm;子球重量則以第三天 最佳為 2.5g,第二~四天無顯著差異分別為 2.3、2.5、2.1g,第一天和第 五天最差為 1.9g 和 1.8g。由以上結果顯示切割後癒傷處理第三天繁殖為 最佳之表現。
(四)、鱗片切割不同份數之比較
將種球切割為 8、8+1/2、16 等份不同份數之刻溝雙鱗片繁殖法,其 結果顯示如(表 6)。‘Apple Blossom’ 切割 8、8+1/2、16 等份其所形成子 球數分別為 49.3、49.8、42.3 個,子球形成數以 8+1/2 最多,16 等份最
少,但 8、8+1/2 無顯著差異。子球重分別為 1.0g、1.1g、1.8g;子球直 徑分別為 5.5、5.5、6.2(mm);平均葉片數分別為 1.8、1.9、1.3 個;最大 葉長分別為 20.4、19.4、20.4cm;以上結果顯示不同切割份數對子球重、
子球直徑、葉片數、最大葉長無顯著差異。
另外一個品種‘Blossom Peacock’利用切割為 8、8+1/2、16 等份不同 份數之刻溝雙鱗片繁殖法,其品種在繁殖上鱗片腐爛率高,每一種切法 皆有整顆種球腐爛而沒有形成子球,故導致整體子球形成數量很少。切 割 8、8+1/2、16 等份其所形成子球數分別為 10.5、11.3、12.個,子球重 分別為 2.1g、1.4g、1.8g;子球直徑分別為 8.1、7.3、7.3mm;平均葉片 數分別為 1.5、1.2、1.2 個;最大葉長分別為 14.1、4.8、8.8cm。
‘Apple Blossom’和‘Blossom Peacock’兩品種間比較,品種間子球繁殖率以
‘Apple Blossom’最好,子球生長狀況其葉片數及最大葉長亦最佳,子球 數愈多,其相對子球直徑則會較小。
三、刻溝雙鱗片產業應用之結果
利用刻溝雙鱗片法應用於孤挺花產業繁殖技術上(表 8)(表 9)(圖 17),
其品種 A210 種球分為大球(周徑 26cm)、中球(19cm)及小球(13cm),其所 形成平均子球數分別為 20.2、13.5、6.4 個,大球最多子球數為 37 個;其 子球直徑分別為 12.4、10.5、9.3mm,子球重為 1.7g、1.4g、1.4g,最大 葉長為 13.3、11.9、14.6cm,結果顯示種球周徑愈大,所形成子球數愈多 和子球愈大,周徑 13cm 以上之種球即可具有繁殖子球之能力。
刻溝雙鱗片、刻溝和瓣狀三種不同切割繁殖法之比較(表 8)(表 9)(圖 16),所形成子球數為 36.4、16.1、12.1 個,以刻溝雙鱗片法最佳,刻溝法 次之,瓣狀法最差,子球直徑分別為 7.5、13.6、16.2mm,子球重為 2.2、
6.6、13.1g,以上結果顯示子球數量愈多,其平均子球直徑和子球重量則 相對較小。
刻溝雙鱗片繁殖法第一次應用於產業上,因技術上尚未純熟及品種間 的差異,所形成之繁殖倍率並未如預期中的結果為佳,若能在技術落實及 栽培管理上之改進,相信刻溝雙鱗片繁殖法在孤挺花繁殖上應有更好繁殖 倍率之表現,可將此技術推廣於孤挺花產業界。
伍、討論(Discussion)
(一)、刻溝雙鱗片繁殖法之探討
本研究開發之刻溝雙鱗片法為結合刻溝和雙鱗片所改良之孤挺花 繁殖方式。刻溝繁殖法可形成之子球數為 12~25 個,繁殖四個月後子球 周徑 6~7cm(朱,2001)。雙鱗片繁殖法繁殖倍率高,子球數可達 50~75 個,
但子球生長速度較慢,切割四個月後的子球周徑為 2~3cm (張,1998)。雖 然國外孤挺花商業種球繁殖普遍採用雙鱗片繁殖法,但在台灣的粗放管 理環境下,鱗片的腐爛率比率偏高,操作技術需大量人力,且子球生長 速度慢,因此該方法在台灣使用上會有許多困擾及較高的成本,故目前 台灣孤挺花的繁殖主要是瓣狀鱗片法為主。
本研究開發之刻溝雙鱗片繁殖法所形成子球數量多,繁殖倍率高於 瓣狀及刻溝法,周徑 26cm 以上種球所形成子球數可達 30~60 餘個,最 高可達 82 個,且所形成子球平均直徑為 6mm 以上,鱗片中外層之子球 直徑甚至可達 12~16mm,其所形成之子球較大,可達刻溝和瓣狀法之 子球大小,故子球之生長速度優於雙鱗片繁殖法。故刻溝雙鱗片繁殖法 結合了雙鱗片繁殖倍率高和刻溝法形成子球較大之優勢,實為改進孤挺 花鱗片繁殖之有效方式。
孤挺花鱗片繁殖時,小鱗莖是由鱗片背軸面接近基盤之表皮細胞及 次表皮細胞分裂所形成(Huang,1990)。大部份小鱗莖生長位置是由雙
鱗片之間基部接近兩邊切口處著生,偶會有小鱗莖著生於雙鱗片內側或 外側之基盤與鱗片之間,推測於癒傷組織形成時有較多的內生荷爾蒙蓄 積,刺激近切口處維管束附近的表皮細胞分裂、分化成小鱗莖(張,
1998)。刻傷種球鱗片,癒傷組織會促使基盤產生不定芽小鱗莖。小鱗 莖生長初期其養份是由母球鱗片所供應,因刻溝法不分離母球,因此子 球可獲得較多的養份來提供子球的生長發育,故所形成之子球較大、子 球鮮重和葉長也明顯較佳。雙鱗片繁殖法是將鱗片分離母球,並形成一 片一片小單位的雙鱗片,因此每一組雙鱗片所含有之養份也相對的減少 許多,故所形成之子球生長速度減緩,在相同繁殖期間所形成之子球較 小。綜合上述,刻溝雙鱗片法因不分離母球,故所能提供子球之養份較 雙鱗片為多,且不分離母球其鱗片較不易腐爛,相對可提高子球形成 率,在種球刻溝上做雙鱗片的處理,能促使更多的癒傷組織形成不定 芽,且有更多的生長空間來形成小鱗莖。在操作管理上,雙鱗片需要許 多的人力及栽培空間,但刻溝雙鱗片法操作簡易且繁殖初期所需之栽培 空間較小,在管理上較為簡便,故刻溝雙鱗片法能有效改進孤挺花鱗片 繁殖之方式。
(二)、種球大小、品種差異對子球數量及生長速度之影響
本實驗結果顯示,種球愈大,繁殖體之鱗片愈大,所能提供子球生 長之養份愈多,因此所形成之子球數量也愈多,所形成之子球也愈大。
種球周徑 30cm 以上,所形子球數量最多,約可達 40~60 餘個,最高甚 至可達 80 幾個,周徑 22~29cm 之種球,子球數量約為 30~40 餘個,種
球周徑 15cm,子球數量約為 15~20 餘個。種球愈大,所形成子球之鮮 重和子球直徑也愈大,且葉片數和最大葉長也愈大。本實驗中子球的鮮 重是子球及其葉及根的總重,鮮重較重者也代表葉和根的生長量多,根 和葉的生長旺盛會促進子球的生長速度,使子球快速形成。一般孤挺花 種球周徑達 20cm 即達商業開花球之標準,本實驗種球周徑達 13cm 即 可具有繁殖子球之能力,但子球數量未超過 20 個。種球周徑 20cm 所形 成子球數量可達 30 個以上,繁殖倍率相對刻溝和瓣狀法已大幅提高,
故種球周徑 20cm 即可達商業種球繁殖生產規模。
子球形成數之另一因素為種球切割份數之影響,種球切割愈細,子 球形成數愈多,相對子球周徑也愈小,種球繁殖期間也愈長(朱,2003)。
呂(1995)於金花石蒜雙鱗片試驗中,子球形成率以 8 分割最佳,16 等份 之雙鱗片形成率較低。本實驗將種球切割成 8 等份為較佳,周徑 20~34cm 一般分割為 8 等份最為適當。若切割成 16 等份,鱗片分割太細會造成 鱗片容易腐爛,且在操作上也較為費時。
在不同品種間子球之繁殖率有顯著的差異,本實驗結果以`Apple Blossom’和`Lady Jane’之繁殖率最佳,所形成子球數量可達 40~60 餘 個,`Apple Blossom’最高子球數甚至可達 82 個,`Lady Jane’ 最高子 球數可達 68 個,`Red Lion’和`Blossom Peacock’繁殖率次之,子球數 約為 20~30 幾個,`Double Record’繁殖率最差,子球數約為 13~20 餘 個。另外`Blossom Peacock’在鱗片繁殖時即易腐爛,導致子球形成率 低,然而此品種之自然分球率高可達 6~8 個。`Double Record’則為子
球繁殖率低之品種。`Apple Blossom’雙鱗片有 94.4%子球形成率,
`Red Lion’有 76.4%子球形成率,此結果與田間栽培自然分球率的表現 同(張,1998)。
(三)、切割時期、栽培環境對對子球數量及生長速度之影響
Hayashi 及 Suzuki(1970)指出,孤挺花生長的溫度最低溫限制為 8℃,28/23℃會造成實生苗球根肥大不良,於 23/18℃、18/13℃下球 根重的增加速度最快,30℃前後的高溫易造成鱗片的腐爛及小球生長 速度的減緩。因此台灣平地秋冬至初春的冷涼氣候較適合小球繁殖(表 12)。
本實驗結果刻溝雙鱗片繁殖時期以一月、七月表現最佳無顯著差 異,十月次之,四月最差。本地夏季高溫雖達 30℃以上,但本實驗栽 培環境皆位於溫室,溫室於夏季有遮陰及噴霧系統,在溫度管理上可 避免高溫。四月份繁殖率最差,推測除了五、六月高溫高濕及病蟲害 造成鱗片容易發霉且腐爛率高,而導致子球形成率非常低,另一主要 因素為四月份為孤挺花之開花期,開完花後老舊之花莖在鱗片中極易 腐爛而造成鱗片之感染,且開完花之鱗片因養份移轉,此時種球鱗片 養份會減少,因此四月份春末夏初為一年中較不適合繁殖之時期。
種球切割後以靜置三天待癒傷組織傷口癒合後再進行培育為最 佳,若種球靜置太久會造成鱗片已開始乾枯而影響子球形成率。經本 實驗改良栽培環境,發現鱗片繁殖初期因栽培蛭石介質之溼度管理不
當,而易造成鱗片腐爛率提高,其相對子球形成率因而降低。故繁殖 初期一個月時,先將種球放置於層架上無介質培育,並用塑膠袋覆蓋 其上,每日噴水維持其相對溼度約 85~90%,此改良方法在操作管理 上較為簡便,且能控制鱗片不易腐爛。培育 4 週即可長出小鱗莖(圖 10),待子球生長一個月後再將鱗片移至蛭石介質中繁殖二~三個月。
試驗結果其所形成之子球初期生長速度較為快速,三個月即可達四個 月蛭石栽培之子球大小。孤挺花子球初期生長速度緩慢,子球周徑增 加速度最快的時期為切割後 30~60 天(朱,2003)。孤挺花的鱗片較為肥 厚,本身具有足夠的水份及養份可供應小鱗莖形成發育,繁殖期間介 質水份含量對小球形成率無顯著影響,但對小球大小生長速度有影 響,此乃因小球初期生長發育對水份需求增加,故介質中水份含量增 加,小鱗莖生長速度也較快。因此繁殖初期只要能保持鱗片水份及防 止發霉污染,即可成功誘導小鱗莖形成。繁殖初期只需將鱗片放置於 保溼栽培環境中即可,故在栽培環境方面之改進,繁殖期間若能控制 種球鱗片不腐爛且提供足夠的水份保持溼潤,即能有效提高子球形成 數和加速子球生長之速度。若進一步配合子球生長栽培管理,將可縮 短種球生產期間於兩年內可得孤挺花之開花球。
陸、結論 (Conclusion)
本研究開發的刻溝雙鱗片為孤挺花鱗片繁殖之改良方式,在管理 上具有節省空間、操作簡便、子球初期生長快速及繁殖數量多的優 點,同時在子球繁殖率及生長速度上取得平衡,達最佳生產之經濟利 益,實為孤挺花短期快速增殖之有效方法。
台灣地處副熱帶區其天然氣候條件很適合孤挺花周年生產,若能 自行開發培育新品種,並整合種球繁殖、栽培及花期調控等技術,而 不必全仰賴進口開花球,無論在生產盆花、切花或作庭園花壇觀賞,
皆具有產業之發展潛力,在國際市場上將可具有競爭之優勢。
圖1:孤挺花種子 Fig1.The seed of Hippeastrum
圖2:子球自然分球之位置 Fig2.The location of bulblets
圖3:孤挺花種球8等份刻溝法 Fig3. The 8 notching of Hippeastrum bulb.
圖4:孤挺花種球8等份瓣狀法
Fig4. The 8 chipping of Hippeastrum bulb.
圖5.孤挺花種球雙鱗片繁殖法
Fig5. The twin-scaling of Hippeastrum bulb.