Final Report NSC 91-3112-B-006-009
I. Inflammation in patients with ischemic stroke and allergy correlates with higher ROS production in respiratory burst and but not gene polymorphisms in NADPH oxidase p22phox
II. Preparation of insect cell expression library for molecular cloning with antibodies
Principle Investigator:
Chi-Chang Shieh M.D., Ph.D.
Department of Pediatrics, National Cheng Kung University Medical College, and Department of Neurology, Chi-Mei Foundation Hospital, and Institute of
Molecular Medicine, Tainan, Taiwan
Con-investigator: Huey-Juan Lin, M.D., M.Ph., Yu-Chan Chao, Ph.D. Students and Asistants: Yan-Jun Chen, Shi-Ying Lee, Kuo-Pin Chuang
Table of Contents
Page
Abstract (中文)
………..3
Abstract
………..4
Introduction
………..5
Materials and Methods
………..11
Results
………..19
Tables
………..26
Figures
………..29
NAD(P)H 氧化酵素基因的免疫調節功能及多型性在人類過敏疾病和血管疾病的 作用 NSC 91-3112-B-006-009 成果報告 中文摘要 在本計畫中,我們從病患研究及實驗中探討細胞產生氧自由基的酵素 NADPH 氧化? 在缺血性腦中風和過敏性氣喘中所扮演的角色。我們也開 始建立使用 baculovirus 在昆蟲細胞表現人類分子圖書館(c-DNA library),作為找出在細胞上影響氧自由基分子的工具。 我們用病例對照研究的結果顯示出了 NADPH 氧化? 基因型,白血球氧自 由機產生,和缺血性腦中風以及過敏的發炎反應間的關係。研究的結果 顯示,C 型反應蛋白質和白血球產生的氧自由基在中風病患的值都較對 照組高,但是彼此之間沒有顯著的關係。我們也發現 NADPH 氧化? p22phox 基因的基因單核酐酸多型性變異(SNP)C242T 在病患群組中和 氧自由機產生的量有關。這可能會在病患身上成為另一個危險因子。我 們將繼續追蹤這些缺血性腦中風及過敏性氣喘的病患,觀察我們所研究 的發炎因子是否和長期的疾病預後有關。 此外,本計畫中我們製作了人類內皮細胞的 baculovirus 分子圖書館(c-DNA library)並測試了基因重組及感染的效率。以此為基礎,我們將改 進真核細胞的分子表現系統,目標是發展出一個具有分子組態特異性抗 體的基因辨識工具。
NAD(P)H oxidase genes: their immunoregulatory functions and polymorphisms in human allergic and vascular diseases
NSC 91-3112-B-006-009 Final Report
Abstract:
We investigated the role of ROS producing NADPH oxidase in the
inflammation of ischemic stroke and asthma. We also initiated the work in setting up an expression cloning system to pinpoint molecular targets relevant to the regulation of ROS production.
Our results of this case-control study elucidate the relationship between NADPH oxidase genotypes, leukocyte ROS production, and inflammation in patients with ischemic stroke and asthma. We showed that ROS production and hsCRP appear to independently associate with the occurrence of ischemic stroke. Moreover, a SNP, C242T correlates with leukocyte ROS production and can be an additional risk factor for patients with ischemic stroke. We are continuing to follow up these patients with ischemic stroke and allergy and observe whether long-term clinical course
correlates with the genotypes and inflammatory parameters investigated in this study. We prepared a baculovirus c-DNA library in this project and tested the efficiency of gene recombination and infection. Based on the technique platform setup in this project, we are continuing to improve the expression library using baculovirus system with the goal of establishing a highly efficient system to identify conformation sensitive epitopes recognized by antibodies.
Introduction Stoke
Stroke is the second leading cause of death in Taiwan. Eleven out of 100 Taiwanese who died in the year 2000 were due to stroke. This number is still increasing. Therefore, stroke has become an important issue to be solved in medicine. According to its pathogenesis, stroke is classified into ischemic stroke and hemorrhagic stroke. Ischemic stroke is due to blood vessel obstruction caused by stenosis or thrombosis in the brain. Brain tissues supplied by the obstructed vessels become damaged due to ischemic necrosis. There are many risk factors for ischemic stroke. Hypertension, hyperlipidemia, aging, smoking, and diabetes are among the important ones. Atherosclerosis is a notable common process of these risk factors to induce ischemic stroke.
Atherosclerosis
Atherosclerosis is a multifactorial progressive process. Lesions of atherosclerosis are found in medium- and large-sized arteries, including
coronary arteries, cerebral arteries, and aorta. Atherosclerosis causes ischemia through thickening of vessel wall or thrombosis formation that leads to vascular stenosis or emboli. The vascular obstruction may lead to infarction in brain, heart, or other organs. In a developing atherosclerotic plaque (Ross, 1999), changes in endothelial permeability lead to increase accumulation of
lipoproteins in intima. Native low-density lipoproteins (LDL) are modified to oxidized LDL by reactive oxygen species (ROS). Some adhesion molecules, including P-selectin, E-selectin, ICAM-1, VCAM-1, and PCAM-1 on
monocytes and T lymphocytes roll and adhere at these sites and transmigrate into intima. These monocytes are activated and become macrophages, which in turn uptake oxidized LDL and become foam cells. Smooth muscle cells
stimulated by cytokines such as platelet-derived growth factor (PDGF) also migrate from media to intima and become part of the lesions (Rosenfeld et al., 1990). These lesions gradually become fatty streaks. Many cells within fatty streaks undergo apoptosis. These biological processes continue repeatedly and the fatty streaks thicken to form fibrous plaques. If plaques obstruct normal blood flow or rupture, thromboses or emboli will be generated and may lead to brain or heart damage. As immune cells are involved in this process,
atherosclerosis is considered an inflammatory disease.
Reactive oxygen species (ROS)
Superoxide (O2-), hydrogen peroxide (H2O2), nitric oxide (NO), and hypochlorous acid (HOCl) are collectively referred to as reactive oxygen species (ROS) (Dröge, 2002). Many cellular sources had been identified to produce ROS, including NADPH oxidase, xanthine oxidase, lipoxygenase, and mitochondria oxidase. ROS derived from phagocytes are active in killing certain bacteria and fungi (Babior, 1999). Recent papers suggested that signal transductions from some cellular receptors are affected by ROS. For instance, tyrosine kinases including Lck and ZAP70 are activated under highly oxidative conditions (Brumell et al., 1996; Schreck et al., 1991).
Increased O2- production has been implicated to be associated with various vascular diseases (Alexander et al., 1995; Griendling et al., 1997). An
process of atherosclerosis. NO is known to regulate antiatherosclerosis process, including inhibition of adhesion molecule expression; platelet adhesion and migration; and smooth muscle cells proliferation and migration (Khan et al., 1996; Radomski et al., 1987; Garg et al., 1989). Superoxide reacting with NO rapidly form peroxynitrite (ONOO-) so that increased superoxide would impair NO bioactivity (Kojda et al., 1999). Therefore, the balance between O2- and NO may influence normal physiological states and diseases of blood vessels and potentially contributes to the process of atherosclerosis. In addition,
peroxynitrite could initiate LDL oxidation for promotion of foam cell formation (White et al., 1994).
NADPH oxidase
NADPH oxidase is a multi-component enzyme complex present on plasma membrane and granules of phagocytes, including granulocytes and macrophages, and also on B lymphocytes (Segal et al., 2000. Appendix II). NADPH oxidase consists of six major components. Two of them,
glycoprotein gp91phox (phagocyte oxidase) and p22phox, are membrane-bound and collectively named cytochrome b558. The other four are cytosolic components including p40phox, p67phox, p47phox, and Rac 2 protein. In response to relevant stimuli, cytosolic components translocate to membrane and form the complete active enzyme with the membrane bound components. Active NADPH oxidase transfers one electron from NADPH to O2 via its flavin adenine dinucleotide (FAD) and two heme groups to form O2-.
-Superoxide dismutase (SOD) changes O2- to H2O2 which consequently formed other ROS with stronger microcidal activity (Hampton et al., 1998). This process of massive ROS production is termed “respiratory burst”. Chronic granulomatous disease (CGD) is an inherited disorder caused by defective leukocyte NADPH oxidase in phagocytes. Patients with CGD fail to generate superoxide and its metabolites to fight pathogens (Geiszt et al., 2001).
Recently, nonphagocytic cells including endothelial cells and smooth muscle cells were found to express homologs of leukocyte NADPH oxidases (Meyer et al., 2000; Lassegue et al., 2001). These functional NADPH
oxidases substitute gp91phox subunit with homologues Nox 1, Nox 3, Nox 4, or Nox 5 (Cheng et al., 2001). Superoxide production from these cells appears to play a role in cellular signaling other than anti-pathogen function. Fukui et al found that superoxide productions from NADPH oxidase were elevated in aortas of hypertensive rats (Fukui et al., 1997). Moreover, several recent investigations indicated that elevated expression of NADPH oxidases subunits might increase ROS generation in coronary arteries and contribute to the process of atherosclerosis (Azumi et al., 1999; Sorescu et al., 2002).
Polymorphisms of p22phox
Gene of p22phox is located on chromosome 16q24 and contains about 600 bp open reading frame in six exons (Dinauer et al., 1990). Two single nucleotide polymorphisms (SNPs) were found in p22phox gene from NADPH oxidase (Inoue et al., 1998). G242T polymorphism is located in the coding region in exon 4 of p22phox and lead to a substitution of histidine by tyrosine. This site is considered a potential heme binding site so that G242T
polymorphism may affect the enzyme activity of p22phox. A640G is found in 3’ untranslated region in exon 6. A640G polymorphism may influence
stability and processing of p22phox mRNA (Gardemann et al., 1999). Results from several different groups showed that p22phox gene polymorphisms may have different roles in different populations. In a Japanese study, 242T allele was more frequent in control subjects than in patients with coronary artery disease (CAD) (Inoue et al., 1998). However, the results of an Australian study indicated that 242T allele was more prevalent in CAD patients than in control subjects (Cai et al., 1999). In another study, 242T allele was also found to be more frequent in patients with cerebrovascular disease (CVD) than in control subjects in the Japanese population (Ito et al., 2000).
Cytokines in the pathogenesis of atherosclerosis
In the beginning of the atherosclerosis process, oxidized LDL stimulates endothelial cells to express adhesion molecules and produce monocyte
chemotactic protein-1 (MCP-1) and macrophage colony-stimulating factor (M-CSF), and results in recruitment of monocytes in intima. Interferon- (IFN-) secreted from activated T cells facilitates the expression of scavenger receptors on macrophages. Macrophages produce tumor nerosis factor- (TNF-) for negative feedback of this reaction (Lusis, 2000). Macrophges endocytose oxidized LDL and become foam cells. Foam cells formation is mediated by MCP-1 and interleukin-1. Platelet- deriverd growth factor (PDGF), fibroblast growth factor-2 (FGF-2), and transforming growth factor (TGF-) stimulate the migration of smooth muscle cells and facilitate the process of
that several plasma markers of inflammation were potential tools for monitoring the process of atherosclerosis. Interleukin-6 (IL-6), soluble intercellular
adhesion molecular type 1 (sICAM-1), high sensity C-reactive protein (hs-CRP) and serum amyloid A produced from the liver are some of these parameters of inflammation in the atherosclerosis. Among them, CRP was reported to be the most powerful predictor of CVD (Ridken et al., 2000).
Aim of this study
By collecting peripheral blood, we measured superoxide production of NADPH oxidases from patients with ischemic stroke and control subjects. We also examined SNP in exon 4 and exon 6 of p22phox to study a correlation between genetic background of NADPH oxidase and ischemic stroke. We also tested inflammation markers to investigate the role of inflammation in ischemic stroke. Through these investigations, we intended to elucidate whether ROS produced by leukocyte NADPH oxidase is related to the pathogenesis of ischemic stroke and allergy.
Material and methods Samples
Patients with ischemic stroke were recruited from Chi-Mei Medical Center in Tainan, Taiwan from September 2000 to April 2002. Age and gender
matched control subjects were recruited as well. Asthmatic children were recruited from National Cheng Kung University Hospital Pediatric Allergy Clinic from September 2002 to June 2003. Tests for normal children were performed at Sheng-Li elementary school, Tainan City.
Leukocyte counts
Peripheral blood samples (10ml) from patients and control subjects were collected in tubes containing 0.5 ml of 0.5 mM ethylenediaminetetra-acetic acid (EDTA). Peripheral blood was mixed well and diluted 1:10 with Turk solution (Merck). Ten μl of diluted solution was put in a hemocytometer chamber and light microscopy with 10X objective lens was used to count numbers of WBC / μl per sample.
Peripheral blood smear preparation
Peripheral blood was mixed well. A drop of blood was put on the end of a clean glass slide. Another slide was placed on the drop of blood and this slide was pushed forward with a rapid movement to make a thin blood smear. Liu A solution (Hand sel Technologies, Inc) was added on the dry blood smear for 30 seconds. Then 2 volume of Liu B solution (Hand sel Technologies, Inc) was added for 60 seconds. These dyes were washed away and blood smear was let
air-dried. We used 100X oil objective lens to count proportions of granulocyte, lymphocyte, and monocyte from a sample.
Isolation of granulocytes and peripheral blood mononuclear cells
Ten volume of peripheral blood was mixed well with one volume of ACD (6% dextran) and waited for 60 minutes. The upper leukocyte rich plasma was layer over Ficoll-Hypaque (Amersham Pharmacia) and centrifuged at 2,000 rpm for 20 minutes (brake off). The ratio of leukocyte rich plasma and
Ficoll-Hypaque was 1:1. Peripheral blood mononuclear cells (PBMC) at the interface between serum and Ficoll-Hypaque were taken into a new tube. Suspension buffer (1g/L BSA, 1X HBSS without Ca2+, Mg2+, and 1mM EDTA, pH 7.3) was added in the tube to wash PBMC. The cell suspension was centrifuged at 1,500 rpm for 5 minutes. Supernatant was discarded and the tube was placed on a clean paper towel to remove residual liquid. This process was repeated for one time. PBMC were ready for experiments. The layer containing red blood cells and granulocytes were taken into another new tube. Lysis buffer (155.2 mM NH4Cl, 20 mM NaHCO3, 1mM EDTA, pH 7.3) was added in the tube. The mixture was incubated at 37°C for 3-5 minutes to lyse red blood cells (RBC). The ratio of granulocytes and lysis buffer was about 1:5. The mixture was centrifuged at 1,500 rpm for 5 minutes to collect granulocytes. The tube was reversed quickly to remove supernatant and blotted on a clean paper towel to dry residual liquid. The pallet of granulocytes was washed by suspension buffer for two times as mentioned above. Granulocytes were ready for experiments.
We took whole blood containing 8105
neutrophils in a 15 ml centrifuge tube and added lysis buffer. The ratio of blood and lysis buffer was about 1:10. The mixture was incubated at 37°C for 3-5 minutes to lyse RBC. The mixture was centrifuged at 1,500 rpm for 5 minutes to pellet leukocytes. The tube was reversed quickly to discard supernatant and blotted on a paper towel to dry residual liquid. The pallet was washed by suspension buffer for one time as mentioned above.
Suspension buffer was added to suspend leukocyte pallet. Leukocytes were stimulated with 2.22 μg/L phorbol myristate acetate (PMA, Molecular Probes) and incubated at 37°C for 10 minutes. The stimulated leukocytes were transferred in a steel dish. The dish was then put in the chamber of lucigenin-enhanced chemiluminescence detector (Tohoku Electronic Industrial Co., Ltd) to measure superoxide production. After the first 200 seconds for measuring background chemiluminescence, 10 μM lucigenin (Sigma) was added to react with superoxide and produce chemiluminescence. Chemiluminescence counts were detected continually from 200 to 600 seconds.
DNA isolation
Genomic DNA was prepared with a Puregene DNA isolation kit (Gentra System, Inc). RBC lysis solution was added to whole blood in a 1.5 ml eppendorf tube. The ratio of RBC lysis solution and blood was 3:1. The eppendorf tube was inverted and incubated at room temperature until mixture became clear. The eppendorf was centrifuged at 13,000 rpm for 20 seconds to collect leukocytes. Supernatant was removed. After 300 μl of cell lysis solution was added, we pipetted up and down to lyse cells. Thereafter 100 μl
of protein precipitation solution was added immediately. The eppendorf tube was vortexed vigorously and centrifuged at 13,000 rpm for 3 minutes to
precipitate proteins. The supernatant was poured into a clean 1.5 ml eppendorf tube and mixed with 300 μl of isopropanol. The eppendorf tube was gently inverted for 50 times until white DNA clump was visible. The eppendorf tube was centrifuged at 13,000 rpm for 1 minute and removed supernatant. Then 300 μl of 70 % ethanol was added. The eppendorf was inverted to wash DNA pellet and was centrifuged at 13,000 rpm for 1 minute. Supernatant was
removed and DNA pellet was dried for 15 minutes. Finally 100 μl of DNA hydration solution was added and the eppendorf tube was placed in aseptic hood overnight. We stored DNA sample at 20 °C.
Polymerase chain reaction
We used polymerase chain reaction (PCR) to amplify exon 4 and exon 6 sequences of p22phox.
Sequence of PCR sense primer for exon 4 is: 5’-TGCTTGTGGGTAAACCAAGGCCGGTG.
Sequence of PCR antisense primer for exon 4 is: 5’-AACACTGAGGTAAGTGGGGGTGGCTCCTGT.
Sequence of PCR sense primer for exon 6 is: 5’-AGCAGTGGACGCCCATCGAGCCCAA.
Sequence of PCR antisense primer for exon 6 is:
5’-CGCTGCGTTTATTGCAGGTGGGTGC (Inoue et al., 1997).
The PCR mixture contained 0.5 μM sense primer, 0.5 μM anti-sense primer, 1X PCR buffer without Mg2+ (MBI Fermentas), 0.2 mM dNTP (Protech), 0.656 μg genomic DNA, 1.5 mM Mg2+ (MBI Fermentas), and 1.2 units Taq polymerase (MBI Fermentas) in a final volume of 30 μl.
Amplification was performed in a GenAmp PCR System 2400 (Perkin Elmer) with 10 minutes of pre-denaturation at 97°C, followed by 1 minute of
denaturatuion at 94°C, 2 minutes of annealing at 64°C, and 2 minutes of extension at 72°C for 40cycles, and a final extension at 72°C for 16 minutes. PCR products were analyzed in 2 % agarose gel for check.
Exon 6 of p22phox
The PCR mixture contained 0.5 μM sense primer, 0.5 μM anti-sense primer, 1X PCR buffer without Mg2+ (MBI Fermentas), 0.2 mM dNTP (Protech), 0.656 μg genomic DNA, 1 mM Mg2+(MBI Fermentas), and 1.2 units Taq polymerase (MBI Fermentas) in a final volume of 30 μl.
Amplification was performed in a GenAmp PCR System 2400 (Perkin Elmer) with 10 minutes of pre-denaturation at 97°C, followed by 1 minute of
denaturation at 94°C, 2 minutes of annealing at 64°C, and 2 minutes of extension at 72°C for 40cycles, and a final extension at 72°C for 16 minutes. PCR products were analyzed in 2 % agarose gel for check.
Restriction enzyme digestion Exon 4 of p22phox
The reaction mixture contained X NEBuffer 1, 3 μl of PCR products of exon 4, and 1 μl of RsaI (NEB) in a final volume of 30 μl. The reaction mixture was mixed well and incubated at 37°C for 2 hours. Mixture was then incubated at 65°C for 20 minutes to inactivate restriction enzyme activity. The reaction products were resolved on 8 % acrylamide gel.
2.1.1 Exon 6 of p22phox
The reaction mixture contained 100 μg/ml BSA, X NEBuffer 3, 3 μl of PCR products of exon 6, and 1μl of DraIII (NEB) in a final volume of 30 μl. The reaction mixture was mixed well and incubated at 37°C for 2 hours. The mixture was then incubated at 65°C for 20 minutes to inactivate restriction enzyme activity. The reaction products were checked on 8 % acrylamide gel.
2.2 PCR product purification
QIAquickTM PCR purification kit (Qiagen) was used to purify PCR
products. One volume of PCR product was added with 5 volumes of Buffer PB. The mixture was added in the QIAquick spin column. The spin column was placed on a 1.5 ml eppendorf and centrifuged at 13,000 rpm for 1 minute. The spin column was placed on the same 1.5 ml eppendorf. Then 750 μl of Buffer PE was added in the column. After centrifugation at 13,000 rpm for 1 minute, flow-through was discarded and placed the spin column on the same 1.5 ml eppendorf. The spin column was centrifuged at 13,000 rpm for 1 minute to remove residual Buffer PE. The spin column was placed on a new 1.5 ml eppendorf and 50 μl of Buffer EB was added to the center of a membrane of
the spin column. After 1 minute, the column was centrifuged at 13,000 rpm for 1 minute. We then collected the purified PCR products in the eppendorf.
DNA sequencing
Purified PCR products were sent to the Automated Sequencing Center of NCKUH for sequencing. Results of DNA sequencing were used to confirm the results of restriction enzyme digestion.
Automatic SNP genotyping
Multiplex PCR was performed in five microliter volumes containing 0.1 units of Taq polymerase (Amplitaq Gold, Applied Biosystems), 5 ng genomic DNA, 2.5 pmol of each PCR primer, and 2.5 µmol of dNTP. Thermocycling was at 95 C for 15 minutes followed by 45 cycles of 95 C for 20 s, 56 C for 30s, 72 C for 30 s. Unincorporated dNTPs were deactivated using 0.3U of Shrimp Alkaline Phosphatase (Roche) followed by primer extension using 5.4 pmol of each primer extension probe, 50 µmole of the appropriate dNTP/ddNTP combination, and 0.5 units of Thermosequenase (Amersham Pharmacia). Reactions were cycled at 94 C for 2 minutes, followed by 40 cycles of 94 degrees for 5 s, 50 degrees for 5 s, 72 degrees for 5 s. Following addition of a cation exchange resin to remove residual salt from the reactions, 7 nanoliters of the purified primer extension reaction was loaded onto a matrix pad (3-hydroxypicoloinic acid) of a
SpectroCHIP (Sequenom, San Diego, CA). SpectroCHIPs were analyzed using a Bruker Biflex III MALDI-TOF mass spectrometer (SpectroREADER, Sequenom, San Diego, CA) and spectra processed using SpectroTYPER (Sequenom). Primer sequences for p22phox exon 4 and exon 6 are:
Exon 4 1st: ACGTTGGATGAAAGGAGTCCCGAGTGGGAG 2nd: ACGTTGGATGTAAAGGGCCCGAACAGCTTC Exon 6 1st: ACGTTGGATGTGACCGACGAGGTCGTGTGA 2nd: ACGTTGGATGGGCTTCGCTGCATTTATTGC respectively.
Measurement of inflammation markers
CRP levels were measured in the Clinical Pathology Labotory in Chi-Mei Medical Center. We used two kinds of enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems, Inc.) to measure inflammation markers from samples. One was DuoSet ELISA Development System to detect IL-6, IFN-γ, MCP-1, sICAM-1. The other was precoated Parameter ELISA System to measure sVCAM-1, sP-selectin, and sE-selectin.
Statistical analysis
The Wilcoxon signed-rank nonparametric test was used to assess the results of inflammation marker measurement because data were no normally distributed. Other statistical analyses were performed with Student’s t-test. Data are expressed as meanSEM. A P value of < 0.05 was considered statistically significant.
Results
Clinical characteristics of ischemic stroke and control subjects
Characters and relevant risk factors for ischemic stroke among patients and controls are shown in Table 1. Age and gender were comparable between patients and control subjects. The risk factors including hypertension, diabetes, smoking, and heart diseases were more prominent in patients. Cholesterol levels of the two groups were almost the same. Clinical ischemic stroke subtypes of patients are listed in Table 2. The following inflammatory parameters were measured in patients and controls.
MCP-1:
MCP-1 plays an important role in the recruitment of monocytes into the arterial wall and the formation of arteriosclerosis. The ELISA data of MCP-1 of patients with ischemic stroke and controls are showed in Figure 10. MCP-1 was significantly unregulated in patients than that in controls (P=0.0005).
Soluble P-selectin:
P-selectin is an adhesion molecule expressed on membrane of endothelial cells. P-selectin is unregulated while inflammation progresses. Soluble P-selectin (sP-seletin) was represented the level of P-selectin. We found that sP-selectin of patients with ischemic stroke was larger in patients than that in controls (P=0.046). Figure 11 showed the result of sP-selectin.
Soluble E-selectin:
Another adhesion molecule, selectin, is expressed on endothelial cells. E-selectin was detected by measuring soluble E-E-selectin (sE-seletin). From the ELISA
measurement, sE-selectin levels of control subjects were higher than those in patients (P<0.05, Figure 12).
Soluble ICAM-1:
Soluble ICAM-1 (sICAM-1) was measured to represent the level of ICAM-1. In contrast to sP-selectin and sE-selectin, sICAM-1 levels were not different between patients and controls (P>0.05, Figure 13).
Soluble VCAM-1
VCAM-1 was detected by measuring soluble VCAM-1 (sVCAM-1). We found that sVCAM-1 levels were not different between patients and controls as well (P>0.05, Figure 14).
IL-6 and IFN-γ from patients and controls could not be detected by ELISA methods so that data of these two cytokines were not shown. A list of comparison of these inflammatory markers in the two groups was shown in Table 3.
Chemiluminescence measurements of ROS production by stimulated leukocytes Chemiluminescence test is an ultra sensitive method for ROS measurement. PMA activates NADPH oxidase to produce superoxide through a protein kinase-C (PKC) dependent pathway. Lucigenin reacts with superoxide and produces
chemiluminescence. We measured superoxide production from leukocyte NADPH oxidase with a chemiluminescence test. Figure 1 showed the result of a typical superoxide measurement experiment. Green and blue lines represented measurement of sample with PMA stimulation or without stimulation. Total time range of
measurement was 600 seconds. After 200 seconds of background detection, lucigenin was added into the sample. CL counts from 200 to 600 seconds represented
superoxide amounts of one measurement. For each patient, we recorded two CL counts, one for the value of a sample with PMA stimulation and the other for that without PMA stimulation. CL counts of PMA stimulation minus that without PMA stimulation was the numerical value representing superoxide production of one person after PMA stimulation.
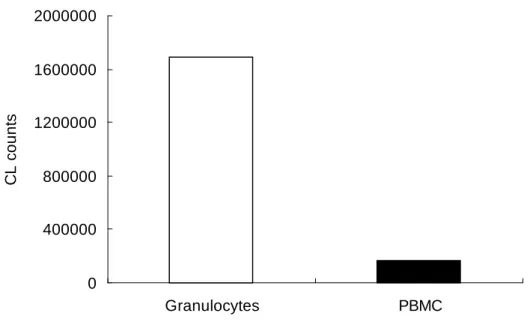
Superoxide production from granulocytes is much higher than that from peripheral blood mononuclear cells.
NADPH oxidase is present in granulocytes, macrophages, and B lymphocytes. We thus assessed which cells are the major sources of superoxide. Using Ficoll-Hypaque, we isolated granulocytes and peripheral blood mononuclear cells (PBMC) from peripheral blood and measured PMA-triggered superoxide from these leukocytes. Figure 2 showed the superoxide production from cells measured by chemiluminescence test. We found that granulocytes produced about ten times more superoxide than PBMC.
Superoxide production was higher in patients with ischemic stroke.
ROS has been considered to be associated with vascular diseases. NADPH oxidase is a major source of ROS production. We sought to investigate the relationship between ROS and ischemic stroke. Using chemiluminescence test, we found that superoxide production of patients was higher than that of control subjects (Figure 3). Superoxide production of follow-up patients in convalescent phase was
measured as well. Compared with patients in acute phase, this elevated superoxide state of patients in convalescent phase did not change significantly (Figure 4).
Gene polymorphism of exon 4 and exon 6 of p22phox in patients with ischemic stroke.
Inoue et al found that there were two SNPs in p22phox of NADPH oxidase (Inoue et al., 1998) and they might be correlated with vascular diseases. To elucidate the relationship between genotypes and development of vascular disease, we sought to study the association between SNPs in p22phox and elevated ROS. We amplified sequences of exon 4 and exon 6 in p22phox through PCR. Two restriction enzymes, RsaI and DraIII, were used to distinguish SNPs of exon4 and exon 6. RsaI cut a recognition sequence (GTAC). PCR products (348 bp) of exon 4 with T allele digested by RsaI lead to 188 bp and 160 bp fragments. On the other hand, DraIII could cleave CACNNNGTG sequence. PCR products (258 bp) of exon 6 with A nucleotide cut by DraIII resulted in 227 bp and 31 bp fragments. PCR products and enzyme digestion products were analyzed in 8% acrylamide gel. A mass
spectrometry-based automatic SNP detection method performed at the Center of Genomic Medicine also was used.
No significant difference was found between ischemic stroke patients and normal controls in these gene polymorphism in p22phox (Fig 7).
Patients with CT or TT genotype on position 242 had higher superoxide production than patients with CC genotype.
We then examined the levels of leukocyte ROS production in patients and controls with different genotypes in p22phox. We found that there is a significantly
lower level of ROS production in patients with CC genotype on position 242 than patients with CT and TT phenotypes. This difference is not apparent in normal controls, who had lower levels of leukocyte ROS production (Fig.8).
Several inflammation markers were increased in patients.
Accumulating investigations suggested that inflammatory processes were involved in each state in atherosclerosis. Several inflammation markers were regarded as predictors of vascular diseases. Therefore, we examed the correlation between ishemic stroke and several markers of inflammation, including CRP, MCP-1, sP-selectin, E-selectin, sICAM-1, and sVCAM-1. Results were expressed as box plots. Minimal and maximal values were represented by circles. Lower and upper sides of boxes represented 25th and 75th percentiles. Medians were showed by the horizontal lines in the middle of boxes.
CRP:
CRP has been found to be a powerful predictor of the risk of cardiovascular diseases (Ridker et al., 2000). Figure 9 showed CRP levels of patients with ischemic stroke and control subjects. CRP levels of patients in acute phase were higher than those of controls (P<0.0001). When patients were in convalescent phase, CRP levels were lower but remained higher than those of controls (P=0.0006).
Relationship between CRP levels and superoxide production in patients with ischemic stroke
Patients and controls were divided into 5 groups according to their CRP values (≦0.5, 0.5-1, 1-3, 3-5, >5 μg/dl). There is no significant differences in leukocyte
ROS production in groups with different CRP levels among patients or normal controls.
Superoxide production was higher in patients with asthma.
We also investigated the relationship between ROS and asthma. Using chemiluminescence test, we found that superoxide production of patients was higher than that of control subjects (Figure 11).
Construction of baculovirus expressing VCAM-1 sequence.
Baculovirus which emits green fluorescence was used to construct
recombinant viruses. Virus with successful recombination emits red fluorescence (Fig.12A). The efficiency of recombination to construct virus capable of infecting insect cells was tested. About 10% of infected cells emit both green and red fluorescence as shown in Fig.12B.
Conclusion and Implications
Our results of this case-control study begin to elucidate the relationship between NADPH oxidase genotypes, leukocyte ROS production, and inflammation in patients with ischemic stroke and asthma. We showed that ROS production and hsCRP appear to independently associate with the occurrence of ischemic stroke. Moreover, a SNP, C242T correlates with leukocyte ROS production and can be an additional risk factor for patients with ischemic stroke. We are continuing to follow up these patients with ischemic stroke and allergy and observe whether long-term clinical course correlates with the genotypes and inflammatory parameters
investigated in this study.
Based on the technique platform setup in this project, we are continuing to improve the expression library using baculovirus system with the goal of establishing a highly efficient system to identify conformation sensitive epitopes recognized by antibodies.
Table 1. Characteristics of patients with ischemic stroke and controls. Patients (N=94) Controls (N=94) P value Age (meanSD), y 54.7 8.5 54.8 9.9 0.95 Men , % 66 66 1.00 Hypertension, % 56 15 <0.001 Diabetes, % 34 6 <0.001 Heart disease, % 15 10 0.27 Smoking, % 37 26 0.08 Cholesterol, mg/dl 20349 20635 0.60
Table 2. Stroke subtypes in 94 patients.
Stroke subtypes Patients
(N=94)
Lacunar, % 48
Large artery atherosclerosis, % 21
Embolism, % 5
Table 3. Inflammatory markers in patients and controls. Patients (N=94) Controls (N=94) P value CRP(mg/L) 2.95 (1.08/7.10) 0.77 (0.36/1.96) <0.0001 sP-selectin(ng/mL) n=82 64.8 (51.2/90.0) 51.7 (39.9/71.9) 0.046 MCP-1(ng/mL) n=76 0.042 (0.030/0.053) 0.021 (0.017/0.038) 0.0005 sICAM-1(ng/mL) n=78 100.98 (20.4/288.66) 89.1 (7.65/177.14) >0.05 sVCAM-1(ng/mL) n=82 341.2 (184.24/709.68) 377 (258.48/550.96) >0.05 sE-selectin(ng/mL) n=76 18.105 (0/85.38) 38.805 (0.329/69.06) <0.05
IFN- Non detectable Non detectable
IL-6 Non detectable Non detectable
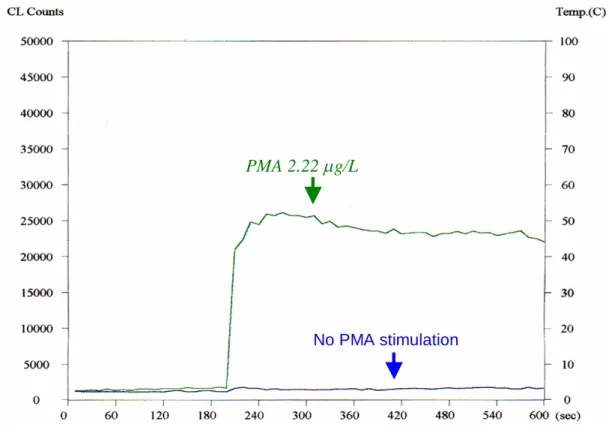
Figure 1. Measurement of superoxide production. The horizontal scale is
time (second). Left vertical scale is level of superoxide production (CL
counts). Right vertical scale is temperature scale (°C). Total
measurement time was 600 seconds. Lucigenin was added when 200
seconds was reached. Blue line represented superoxide level of a sample
without PMA stimulation. Green line was superoxide production after a
sample was incubated with PMA. CL counts from 200 to 600 seconds
were calculated.
PMA 2.22 g/L
Figure 2. Superoxide production from granulocytes and peripheral blood
mononuclear cells (PBMC). After stimulation of PMA (4 g/L) for
10minutes, superoxide generated by granulocytes (, 310
6/ml) and
PBMC (, 310
6/ml) were measured by chemiluminescence test.
Granulocytes produced about much more superoxide than PBMC.
0 400000 800000 1200000 1600000 2000000 Granulocytes PBMC C L co u n ts
Figure 3. Superoxide production from patients with ischemic stroke and
control subjects. Patients with ischemic stroke (, n=94) were in
elevated superoxide production states. Compared with patients,
superoxide levels were lower in control subjects (, n=94). Columns
and error bars stand for means and standard error of means.
0 500000 1000000 1500000 2000000 2500000 Control Patient C L c o u nt s P = 0.0001
Figure 4. Superoxide production from patients in acute and convalescent
phase. Superoxide production from patients in convalescent phase (,
n=38) was as much as that in acute phase (, n=38). Columns and error
bars stand for means and standard error of means.
P > 0.05 0 400000 800000 1200000 1600000
Patients (acute phase) Patients (convalescent phase)
C L c o u n ts
Figure 5. Genotypes of p22phox from patients and control subjects. (A)
RsaI digests PCR products (348 bp) of exon 4 with T allele and makes
188 bp and 160 bp fragments. PCR products (258 bp) of exon 6 with A
nucleotide could be cleavage by DraIII into 227 bp and 31 bp fragments.
(B) PCR products and enzyme digestion products of two exons were
loaded in 8% acrylamide gel. M represents 1kb DNA markers.
A.
Exon 4
Exon 6
——— GGGACAGAAG
C
ACATGA ———
——— GGGACAGAAG
T
ACATGA ———
RsaI site
His
Tyr
——— CTGCCCTCCC
A
CCAGGT ———
——— CTGCCCTCCC
G
CCAGGT ———
DraIII site
B.
M PCR DraIII PCR DraIII PCR DraIII M PCR RsaI PCR RsaI PCR RsaI
Genotype CC CT CT Genotype AG AG AA
A.
B.
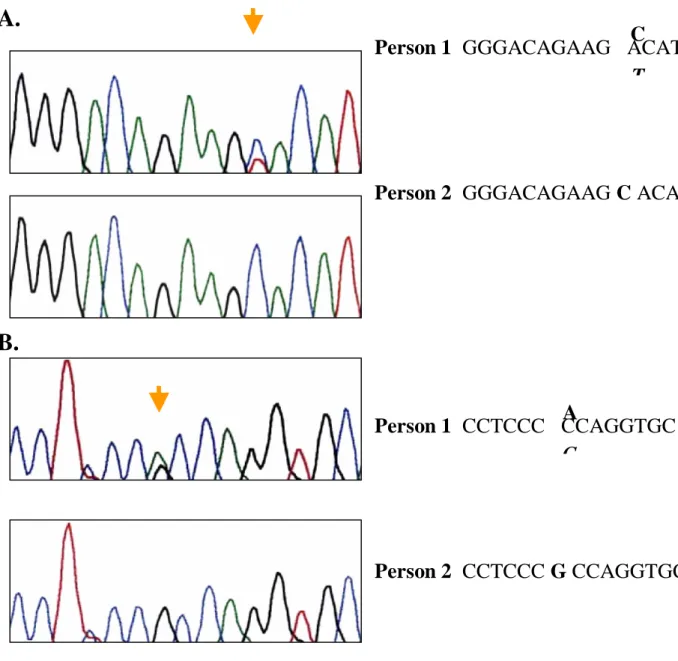
Person 1 GGGACAGAAG ACAT
Person 2 GGGACAGAAG C ACAT
C
T
Figure 6. Sequencing data of exon 4 and exon 6 of p22phox. (A)
Sequences of exon 4 from two persons. Person 1 was heterozygote of CT
and person 2 was CC genotype. (B) Sequences of exon 6 from these two
persons. Person 1 was AG genotype while person 2 was homozygote of
GG.
Person 1 CCTCCC CCAGGTGC
Person 2 CCTCCC G CCAGGTGC
A
A.
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% CC CT TT Control PatientB.
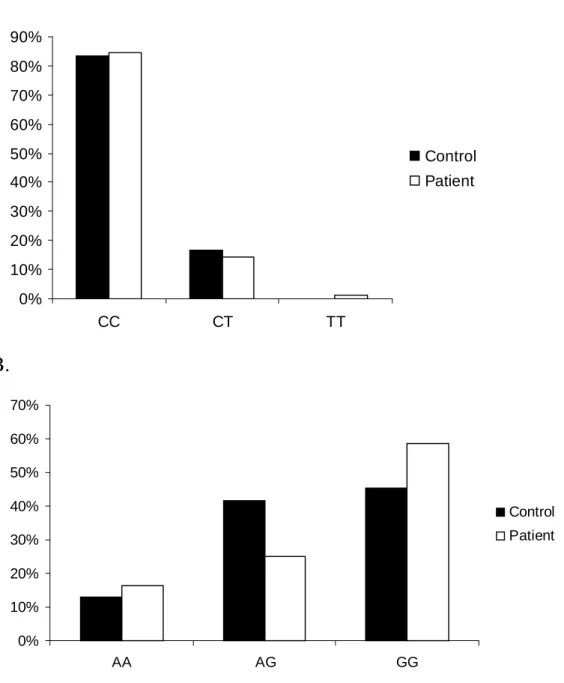
0% 10% 20% 30% 40% 50% 60% 70% AA AG GG Control PatientFigure 7. Single nucleotide polymorphism of p22phox from patients with
ischemic stroke and control subjects. (A) The numbers of C242T
polymorphism in exon 4 from patients (, n=84) and controls (, n=79)
were not different. (B) GG genotypes in patients (, n=84) and controls
(, n=80) were not significantly different (p>0.05).
A. Patients
0 500000 1000000 1500000 2000000 2500000 3000000 3500000 4000000 4500000 CC CT+TT C LB. Control
0 200000 400000 600000 800000 1000000 1200000 CC CT+TT C LFigure 8. Superoxide production by leukocytes exhibiting different
C242T genotypes. There is a significantly lower level of ROS production
in patients with CC genotype on position 242 than patients with CT and
TT phenotypes. This difference is not apparent in normal controls, who
had lower levels of leukocyte ROS production.
P = 0.029
Figure 9. CRP levels of patients and controls. CRP levels (n=257) in
patients were upregulated compared to those in controls. When patients
were in convalescent phase, CRP levels decreased but were higher than
control subjects.
C R P m g /L(0=controls, 1=cases/acute, 2=cases/FU) 0 5 10 20 30 40 50 60 70 80 0 1 2 ( ) Controls Patients (convalescent phase) Patients (acute phase) P < 0.0001 P = 0.002 P = 0.0006
0 1000000 2000000 3000000 4000000 5000000 6000000 7000000 ≦0.5 0.5-1 1-3 3-5 >5 hsCRP value C L Control Patient
Figure 10. CRP levels did not correlated with leukocyte ROS production
in patients and controls. Patients and controls were divided into 5 groups
according to their CRP values (≦0.5, 0.5-1, 1-3, 3-5, >5 μg/dl).
Leukocyte ROS production measured by CL are shown as means
(columns) and standard deviation (error bars).
0
200000
400000
600000
800000
1000000
1200000
1400000
normal
patient
Figure 11. ROS production by leukocytes are higher in asthmatic
childeren when compared with normal controls. Leukocyte ROS
production by patients and normal groups showed significant difference.
P = 0.025A.
B.
Figure 12. Construction of baculovirus expressing VCAM-1 sequence.
Baculovirus which emits green fluorescence was used to construct
recombinant viruses. Virus with successful recombination emits red
fluoresecne. The efficiency of recombination to construct virus capable
of infecting insect cells was tested. About 10% of infected cells emit
both green and red fluorescence as shown in (B).
FITC
References
Ago T, Nunoi H, Ito T, Sumimoto H. Mechanism for phosphorylation-induced
activation of the phagocyte NADPH oxidase protein p47(phox). Triple replacement of serines 303, 304, and 328 with aspartates disrupts the SH3 domain-mediated
intramolecular interaction in p47(phox), thereby activating the oxidase. J Biol Chem 1999; 274(47):33644-33653.
Alexander RW. Theodore Cooper Memorial Lecture. Hypertension and the pathogenesis of atherosclerosis. Oxidative stress and the mediation of arterial inflammatory response: a new perspective. Hypertension 1995; 25(2):155-161. Azumi H, Inoue N, Takeshita S, Rikitake Y, Kawashima S, Hayashi Y, Itoh H, Yokoyama M. Expression of NADH/NADPH oxidase p22phox in human coronary arteries. Circulation 1999; 100(14):1494-1498.
Babior BM. NADPH oxidase: an update. Blood 1999; 93(5):1464-1476.
Blake GJ, Ridker PM. Novel clinical markers of vascular wall inflammation. Circ Res 2001; 89(9):763-771.
Brumell JH, Burkhardt AL, Bolen JB, Grinstein S. Endogenous reactive oxygen intermediates activate tyrosine kinases in human neutrophils. J Biol Chem 1996; 271(3):1455-1461.
Cahilly C, Ballantyne CM, Lim DS, Gotto A, Marian AJ. A variant of p22(phox), involved in generation of reactive oxygen species in the vessel wall, is associated with progression of coronary atherosclerosis. Circ Res 2000; 86(4):391-395.
Cai H, Duarte N, Wilcken DE, Wang XL. NADH/NADPH oxidase p22 phox C242T polymorphism and coronary artery disease in the Australian population. Eur J Clin Invest 1999; 29(9):744-748.
Cheng G, Cao Z, Xu X, van Meir EG, Lambeth JD. Homologs of gp91phox: cloning and tissue expression of Nox3, Nox4, and Nox5. Gene 2001; 269(1-2):131-140. Dinauer MC, Pierce EA, Bruns GA, Curnutte JT, Orkin SH. Human neutrophil cytochrome b light chain (p22-phox). Gene structure, chromosomal location, and mutations in cytochrome-negative autosomal recessive chronic granulomatous disease. J Clin Invest 1990; 86(5):1729-1737.
Droge W. Free radicals in the physiological control of cell function. Physiol Rev 2002; 82(1):47-95.
Dumoutier L, Lejeune D, Colau D, Renauld JC. Cloning and characterization of IL-22 binding protein, a natural antagonist of IL-10-related T cell-derived inducible
Dumoutier L, Louahed J, Renauld JC. Cloning and characterization of IL-10-related T cell-derived inducible factor (IL-TIF), a novel cytokine structurally related to IL-10 and inducible by IL-9. J Immunol 2000; 164(4):1814-1819.
Egashira K, Koyanagi M, Kitamoto S, Ni W, Kataoka C, Morishita R, Kaneda Y, Akiyama C, Nishida KI, Sueishi K, Takeshita A. Anti-monocyte chemoattractant protein-1 gene therapy inhibits vascular remodeling in rats: blockade of MCP-1 activity after intramuscular transfer of a mutant gene inhibits vascular remodeling induced by chronic blockade of NO synthesis. FASEB J 2000; 14(13):1974-1978. Ellson CD, Gobert-Gosse S, Anderson KE, Davidson K, Erdjument-Bromage H, Tempst P, Thuring JW, Cooper MA, Lim ZY, Holmes AB, Gaffney PR, Coadwell J, Chilvers ER, Hawkins PT, Stephens LR. PtdIns(3)P regulates the neutrophil oxidase complex by binding to the PX domain of p40(phox). Nat Cell Biol 2001; 3(7):679-682.
Fickenscher H, Hor S, Kupers H, Knappe A, Wittmann S, Sticht H. The interleukin-10 family of cytokines. Trends Immunol 2002; 23(2):89-96.
Fukui T, Ishizaka N, Rajagopalan S, Laursen JB, Capers Q, Taylor WR, Harrison DG, de Leon H, Wilcox JN, Griendling KK. p22phox mRNA expression and NADPH oxidase activity are increased in aortas from hypertensive rats. Circ Res 1997; 80(1):45-51.
Gardemann A, Mages P, Katz N, Tillmanns H, Haberbosch W. The p22 phox A640G gene polymorphism but not the C242T gene variation is associated with coronary heart disease in younger individuals. Atherosclerosis 1999; 145(2):315-323. Garg UC, Hassid A. Nitric oxide-generating vasodilators and 8-bromo-cyclic guanosine monophosphate inhibit mitogenesis and proliferation of cultured rat vascular smooth muscle cells. J Clin Invest 1989; 83(5):1774-1777.
Geiszt M, Kapus A, Ligeti E. Chronic granulomatous disease: more than the lack of superoxide? J Leukoc Biol 2001; 69(2):191-196.
Griendling KK, Alexander RW. Oxidative stress and cardiovascular disease. Circulation 1997; 96(10):3264-3265.
Griendling KK, Sorescu D, Ushio-Fukai M. NAD(P)H oxidase: role in cardiovascular biology and disease. Circ Res 2000; 86(5):494-501.
Hampton MB, Kettle AJ, Winterbourn CC. Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood 1998; 92(9):3007-3017.
Inoue N, Kawashima S, Kanazawa K, Yamada S, Akita H, Yokoyama M. Polymorphism of the NADH/NADPH oxidase p22 phox gene in patients with coronary artery disease. Circulation 1998; 97(2):135-137.
Ito D, Murata M, Watanabe K, Yoshida T, Saito I, Tanahashi N, Fukuuchi Y. C242T polymorphism of NADPH oxidase p22 PHOX gene and ischemic cerebrovascular disease in the Japanese population. Stroke 2000; 31(4):936-939.
Josephson K, Logsdon NJ, Walter MR. Crystal structure of the IL-10/IL-10R1 complex reveals a shared receptor binding site. Immunity 2001; 15(1):35-46. Khan BV, Harrison DG, Olbrych MT, Alexander RW, Medford RM. Nitric oxide regulates vascular cell adhesion molecule 1 gene expression and redox-sensitive transcriptional events in human vascular endothelial cells. Proc Natl Acad Sci U S A 1996; 93(17):9114-9119.
Kojda G, Harrison D. Interactions between NO and reactive oxygen species: pathophysiological importance in atherosclerosis, hypertension, diabetes and heart failure. Cardiovasc Res 1999; 43(3):562-571.
Kotenko SV, Izotova LS, Mirochnitchenko OV, Esterova E, Dickensheets H,
Donnelly RP, Pestka S. Identification of the functional interleukin-22 (IL-22) receptor complex: the IL-10R2 chain (IL-10Rbeta ) is a common chain of both the IL-10 and IL-22 (IL-10-related T cell-derived inducible factor, IL-TIF) receptor complexes. J Biol Chem 2001; 276(4):2725-2732.
Kotenko SV, Izotova LS, Mirochnitchenko OV, Esterova E, Dickensheets H,
Donnelly RP, Pestka S. Identification, cloning, and characterization of a novel soluble receptor that binds IL-22 and neutralizes its activity. J Immunol 2001; 166(12):7096-7103.
Lassegue B, Sorescu D, Szocs K, Yin Q, Akers M, Zhang Y, Grant SL, Lambeth JD, Griendling KK. Novel gp91(phox) homologues in vascular smooth muscle cells : nox1 mediates angiotensin II-induced superoxide formation and redox-sensitive signaling pathways. Circ Res 2001; 88(9):888-894.
Lusis AJ. Atherosclerosis. Nature 2000; 407(6801):233-241.
Meyer JW, Schmitt ME. A central role for the endothelial NADPH oxidase in atherosclerosis. FEBS Lett 2000; 472(1):1-4.
Powell LJ. C-reactive protein--a review. Am J Med Technol 1979; 45(2):138-142. Radomski MW, Palmer RM, Moncada S. Comparative pharmacology of endothelium-derived relaxing factor, nitric oxide and prostacyclin in platelets. Br J Pharmacol 1987; 92(1):181-187.
Ramos CL, Huo Y, Jung U, Ghosh S, Manka DR, Sarembock IJ, Ley K. Direct demonstration of P-selectin- and VCAM-1-dependent mononuclear cell rolling in early atherosclerotic lesions of apolipoprotein E-deficient mice. Circ Res 1999; 84(11):1237-1244.
Ridker PM, Hennekens CH, Buring JE, Rifai N. C-reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women. N Engl J Med 2000; 342(12):836-843.
Rollins BJ. Chemokines. Blood 1997; 90(3):909-928.
Rosenfeld ME, Palinski W, Yla-Herttuala S, Carew TE. Macrophages, endothelial cells, and lipoprotein oxidation in the pathogenesis of atherosclerosis. Toxicol Pathol 1990; 18(4 Pt 1):560-571.
Ross R. Atherosclerosis--an inflammatory disease. N Engl J Med 1999; 340(2):115-126.
Sato TK, Overduin M, Emr SD. Location, location, location: membrane targeting directed by PX domains. Science 2001; 294(5548):1881-1885.
Segal BH, Leto TL, Gallin JI, Malech HL, Holland SM. Genetic, biological, and clinical features of chronic granulomatous disease. Medicine 2000; 79(3):170-200. Sorescu D, Weiss D, Lassegue B, Clempus RE, Szocs K, Sorescu GP, Valppu L, Quinn MT, Lambeth JD, Vega JD, Taylor WR, Griendling KK. Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation 2002; 105(12):1429-1435.
Stanger O, Renner W, Khoschsorur G, Rigler B, Wascher TC. NADH/NADPH oxidase p22 phox C242T polymorphism and lipid peroxidation in coronary artery disease. Clin Physiol 2001; 21(6):718-722.
Verma S, Li SH, Badiwala MV, Weisel RD, Fedak PW, Li RK, Dhillon B, Mickle DA. Endothelin antagonism and interleukin-6 inhibition attenuate the proatherogenic effects of C-reactive protein. Circulation 2002; 105(16):1890-1896.
Warnholtz A, Nickenig G, Schulz E, Macharzina R, Brasen JH, Skatchkov M, Heitzer T, Stasch JP, Griendling KK, Harrison DG, Bohm M, Meinertz T, Munzel T.
Increased NADH-oxidase-mediated superoxide production in the early stages of atherosclerosis: evidence for involvement of the renin-angiotensin system. Circulation 1999; 99(15):2027-2033.
White CR, Brock TA, Chang LY, Crapo J, Briscoe P, Ku D, Bradley WA, Gianturco SH, Gore J, Freeman BA, . Superoxide and peroxynitrite in atherosclerosis. Proc Natl Acad Sci U S A 1994; 91(3):1044-1048.
Xie MH, Aggarwal S, Ho WH, Foster J, Zhang Z, Stinson J, Wood WI, Goddard AD, Gurney AL. Interleukin (IL)-22, a novel human cytokine that signals through the interferon receptor-related proteins CRF2-4 and IL-22R. J Biol Chem 2000; 275(40):31335-31339.